
Serological Evidence of Backyard Pig Exposure to Highly Pathogenic Avian Influenza H5N8 Virus during 2016–2017 Epizootic in France

 , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Herds and Sampling
2.2. ELISA
2.3. Virus Strains Used in Hemagglutination Inhibition Tests
2.4. Production of Negative and Positive Control Sera in Specific Pathogen Free Pigs
2.5. Hemagglutination Inhibition Tests
3. Results
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Vincent, A.; Awada, L.; Brown, I.; Chen, H.; Claes, F.; Dauphin, G.; Donis, R.; Culhane, M.; Hamilton, K.; Lewis, N.; et al. Review of influenza A virus in swine worldwide: A call for increased surveillance and research. Zoonoses Public Health 2014, 61, 4–17. [Google Scholar] [CrossRef]
- Kuntz-Simon, G.; Madec, F. Genetic and antigenic evolution of swine influenza viruses in Europe and evaluation of their zoonotic potential. Zoonoses Public Health 2009, 56, 310–325. [Google Scholar] [CrossRef]
- Nidom, C.A.; Takano, R.; Yamada, S.; Sakai-Tagawa, Y.; Daulay, S.; Aswadi, D.; Suzuki, T.; Suzuki, Y.; Shinya, K.; Iwatsuki-Horimoto, K.; et al. Influenza A (H5N1) viruses from pigs, Indonesia. Emerg. Infect. Dis. 2010, 16, 1515–1523. [Google Scholar] [CrossRef] [PubMed]
- Kwon, T.Y.; Lee, S.S.; Kim, C.Y.; Shin, J.Y.; Sunwoo, S.Y.; Lyoo, Y.S. Genetic characterization of H7N2 influenza virus isolated from pigs. Vet. Microbiol. 2011, 153, 393–397. [Google Scholar] [CrossRef]
- Zhang, G.; Kong, W.; Qi, W.; Long, L.-P.; Cao, Z.; Huang, L.; Qi, H.; Cao, N.; Wang, W.; Zhao, F.; et al. Identification of an H6N6 swine influenza virus in southern China. Infect. Genet. Evol. 2011, 11, 1174–1177. [Google Scholar] [CrossRef] [Green Version]
- Hu, Y.; Liu, X.; Li, S.; Guo, X.; Yang, Y.; Jin, M. Complete genome sequence of a novel H4N1 influenza virus isolated from a pig in central China. J. Virol. 2012, 86, 13879. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, X.; Fu, Y.; Yang, J.; Guo, J.; He, J.; Guo, J.; Weng, S.; Jia, Y.; Liu, B.; Li, X.; et al. Genetic and biological characterization of two novel reassortant H5N6 swine influenza viruses in mice and chickens. Infect. Genet. Evol. 2015, 36, 462–466. [Google Scholar] [CrossRef]
- Scholtissek, C.; Burger, H.; Bachmann, P.A.; Hannoun, C. Genetic relatedness of hemagglutinins of the H1 subtype of influenza A viruses isolated from swine and birds. Virology 1983, 129, 521–523. [Google Scholar] [CrossRef]
- Munoz, O.; De Nardi, M.; van der Meulen, K.; van Reeth, K.; Koopmans, M.; Harris, K.; von Dobschuetz, S.; Freidl, G.; Meijer, A.; Breed, A.; et al. Genetic Adaptation of Influenza A Viruses in Domestic Animals and Their Potential Role in Interspecies Transmission: A Literature Review. Ecohealth 2016, 13, 171–198. [Google Scholar] [CrossRef] [PubMed]
- Watson, S.J.; Langat, P.; Reid, S.M.; Lam, T.T.; Cotten, M.; Kelly, M.; Van Reeth, K.; Qiu, Y.; Simon, G.; Bonin, E.; et al. Molecular Epidemiology and Evolution of Influenza Viruses Circulating within European Swine between 2009 and 2013. J. Virol. 2015, 89, 9920–9931. [Google Scholar] [CrossRef] [Green Version]
- Chiapponi, C.; Ebranati, E.; Pariani, E.; Faccini, S.; Luppi, A.; Baioni, L.; Manfredi, R.; Carta, V.; Merenda, M.; Affanni, P.; et al. Genetic analysis of human and swine influenza A viruses isolated in Northern Italy during 2010-2015. Zoonoses Public Health 2017. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Henritzi, D.; Petric, P.P.; Lewis, N.S.; Graaf, A.; Pessia, A.; Starick, E.; Breithaupt, A.; Strebelow, G.; Luttermann, C.; Parker, L.M.K.; et al. Surveillance of European Domestic Pig Populations Identifies an Emerging Reservoir of Potentially Zoonotic Swine Influenza A Viruses. Cell Host Microbe 2020. [Google Scholar] [CrossRef] [PubMed]
- Sosa Portugal, S.; Cortey, M.; Tello, M.; Casanovas, C.; Mesonero-Escuredo, S.; Barrabés, S.; Pineda, P.; Wacheck, S.; Martín-Valls, G.; Mateu, E. Diversity of influenza A viruses retrieved from respiratory disease outbreaks and subclinically infected herds in Spain (2017–2019). Transbound. Emerg. Dis. 2020. [Google Scholar] [CrossRef]
- Chastagner, A.; Hervé, S.; Quéguiner, S.; Hirchaud, E.; Lucas, P.; Gorin, S.; Béven, V.; Barbier, N.; Deblanc, C.; Blanchard, Y.; et al. Genetic and Antigenic Evolution of European Swine Influenza A Viruses of HA-1C (Avian-Like) and HA-1B (Human-Like) Lineages in France from 2000 to 2018. Viruses 2020, 12, 1304. [Google Scholar] [CrossRef]
- De Vleeschauwer, A.; Van Poucke, S.; Braeckmans, D.; Van Doorsselaere, J.; Van Reeth, K. Efficient transmission of swine-adapted but not wholly avian influenza viruses among pigs and from pigs to ferrets. J. Infect. Dis. 2009, 200, 1884–1892. [Google Scholar] [CrossRef]
- Choi, Y.K.; Nguyen, T.D.; Ozaki, H.; Webby, R.J.; Puthavathana, P.; Buranathal, C.; Chaisingh, A.; Auewarakul, P.; Hanh, N.T.; Ma, S.K.; et al. Studies of H5N1 influenza virus infection of pigs by using viruses isolated in Vietnam and Thailand in 2004. J. Virol. 2005, 79, 10821–10825. [Google Scholar] [CrossRef] [Green Version]
- Wang, J.; Wu, M.; Hong, W.; Fan, X.; Chen, R.; Zheng, Z.; Zeng, Y.; Huang, R.; Zhang, Y.; Lam, T.T.-Y.; et al. Infectivity and Transmissibility of Avian H9N2 Influenza Viruses in Pigs. J. Virol. 2016, 90, 3506–3514. [Google Scholar] [CrossRef] [Green Version]
- Kaplan, B.S.; Torchetti, M.K.; Lager, K.M.; Webby, R.J.; Vincent, A.L. Absence of clinical disease and contact transmission of HPAI H5NX clade 2.3.4.4 from North America in experimentally infected pigs. Influenza Other Respir. Viruses 2017, 11, 464–470. [Google Scholar] [CrossRef] [Green Version]
- Powell, J.D.; Abente, E.J.; Torchetti, M.K.; Killian, M.L.; Vincent, A.L. An avian influenza virus A(H7N9) reassortant that recently emerged in the United States with low pathogenic phenotype does not efficiently infect swine. Influenza Other Respir Viruses 2019. [Google Scholar] [CrossRef] [PubMed]
- The Commission of the European Communities. Commission Decision of 4 August 2006 Approving a Diagnostic Manual for Avian Influenza as Provided for in Council Directive 2005/94/EC (Notified under Document Number C(2006) 3477) (Text with EEA Relevance); Oj L 237/1; The Commission of the European Communities: Brussels, Belgium, 2006; Volume 2006/437/CE, pp. 1–27. [Google Scholar]
- The French Ministry of Agriculture. Mesures applicables suite à une suspicion ou à la mise en évidence de foyer IAHP en France; The French Ministry of Agriculture: Paris, French, 2016; Volume DGAL/SASPP/2016-1019. [Google Scholar]
- Napp, S.; Majo, N.; Sanchez-Gonzalez, R.; Vergara-Alert, J. Emergence and spread of highly pathogenic avian influenza A(H5N8) in Europe in 2016–2017. Transbound. Emerg. Dis. 2018, 65, 1217–1226. [Google Scholar] [CrossRef] [Green Version]
- Alarcon, P.; Brouwer, A.; Venkatesh, D.; Duncan, D.; Dovas, C.I.; Georgiades, G.; Monne, I.; Fusaro, A.; Dan, A.; Smietanka, K.; et al. Comparison of 2016-17 and Previous Epizootics of Highly Pathogenic Avian Influenza H5 Guangdong Lineage in Europe. Emerg. Infect. Dis. 2018, 24, 2270–2283. [Google Scholar] [CrossRef] [PubMed]
- Adlhoch, C.; Brown, I.H.; Angelova, S.G.; Balint, A.; Bouwstra, R.; Buda, S.; Castrucci, M.R.; Dabrera, G.; Dan, A.; Grund, C.; et al. Highly pathogenic avian influenza A(H5N8) outbreaks: Protection and management of exposed people in Europe, 2014/15 and 2016. Eurosurveillance 2016, 21. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guinat, C.; Nicolas, G.; Vergne, T.; Bronner, A.; Durand, B.; Courcoul, A.; Gilbert, M.; Guerin, J.L.; Paul, M.C. Spatio-temporal patterns of highly pathogenic avian influenza virus subtype H5N8 spread, France, 2016 to 2017. Euro Surveill. 2018, 23. [Google Scholar] [CrossRef] [PubMed]
- Briand, F.-X.; Niqueux, E.; Schmitz, A.; Martenot, C.; Cherbonnel, M.; Massin, P.; Kerbrat, F.; Chatel, M.; Guillemoto, C.; Guillou-Cloarec, C.; et al. Highly Pathogenic Avian Influenza A(H5N8) Virus Spread by Short- and Long-Range Transmission, France, 2016–2017. Emerg. Infect. Dis. J. 2021, 27, 508. [Google Scholar] [CrossRef] [PubMed]
- Lycett, S.J.; Pohlmann, A.; Staubach, C.; Caliendo, V.; Woolhouse, M.; Beer, M.; Kuiken, T. Genesis and spread of multiple reassortants during the 2016/2017 H5 avian influenza epidemic in Eurasia. Proc. Natl. Acad. Sci. USA 2020, 117, 20814–20825. [Google Scholar] [CrossRef]
- Hervé, S.; Deblanc, C.; Gorin, S.; Cador, C.; Andraud, M.; Quéguiner, S.; Barbier, N.; Paboeuf, F.; Rose, N.; Simon, G. Infection grippale en post-sevrage: Impact de l’âge et interférence de l’immunité passive du porcelet sur les paramètres cliniques, virologiques et sérologiques après challenge par un virus H1N1. In Proceedings of the Journées de la Recherche Porcine, Paris, France, 6–7 February 2018; pp. 189–194. [Google Scholar]
- Grech-Angelini, S.; Hervé, S.; Rose, N.; Barbier, N.; Casabianca, F.; Maestrini, O.; Falchi, A.; Simon, G. Serological survey of influenza A viruses in domestic and wild Suidae in Corsica (France), a Mediterranean island environment. Prev. Vet. Med. 2018, 157, 94–98. [Google Scholar] [CrossRef] [PubMed]
- The Commission of the European Communities. Commission Decision of 25 June 2010 on the Implementation by Member States of Surveillance Programmes for Avian Influenza in Poultry and Wild Birds; 2010/367/EU; The Commission of the European Communities: Brussels, Belgium, 2010; Volume OJ L 166/22. [Google Scholar]
- OIE World Organisation for animal Health. Avian influenza (infection with avian influenza viruses). In Manual of Diagnostic Tests and Vaccines for Terrestrial Animals 2018, 8th ed.; World Organization for Animal Health, Ed.; OIE World Organisation for animal Health: Paris, France, 2018; pp. 821–843. ISBN 978-92-95108-18-9. [Google Scholar]
- Simon, G.; Larsen, L.E.; Durrwald, R.; Foni, E.; Harder, T.; Van Reeth, K.; Markowska-Daniel, I.; Reid, S.M.; Dan, A.; Maldonado, J.; et al. European surveillance network for influenza in pigs: Surveillance programs, diagnostic tools and Swine influenza virus subtypes identified in 14 European countries from 2010 to 2013. PLoS ONE 2014, 9, e115815. [Google Scholar] [CrossRef] [PubMed]
- OIE World Organisation for animal Health. Influenza A virus of swine. In Manual of Diagnostic Tests and Vaccines for Terrestrial Animals 2018, 8th ed.; World Organization for Animal Health, Ed.; OIE World Organisation for animal Health: Paris, France, 2018; pp. 1594–1607. ISBN 978-92-95108-18-9. [Google Scholar]
- Anderson, T.K.; Macken, C.A.; Lewis, N.S.; Scheuermann, R.H.; Van Reeth, K.; Brown, I.H.; Swenson, S.L.; Simon, G.; Saito, T.; Berhane, Y.; et al. A Phylogeny-Based Global Nomenclature System and Automated Annotation Tool for H1 Hemagglutinin Genes from Swine Influenza A Viruses. mSphere 2016, 1, e00275-00216. [Google Scholar] [CrossRef] [Green Version]
- Smith, G.J.; Donis, R.O. Nomenclature updates resulting from the evolution of avian influenza A(H5) virus clades 2.1.3.2a, 2.2.1, and 2.3.4 during 2013-2014. Influenza Other Respir Viruses 2015, 9, 271–276. [Google Scholar] [CrossRef]
- Hervé, S.; Garin, E.; Calavas, D.; Lecarpentier, L.; Ngwa-Mbot, D.; Poliak, S.; Wendling, S.; Rose, N.; Simon, G. Virological and epidemiological patterns of swine influenza A virus infections in France: Cumulative data from the RESAVIP surveillance network, 2011–2018. Vet. Microbiol. 2019, 239, 108477. [Google Scholar] [CrossRef]
- Loeffen, W.; Boer, E.d.; Koch, G. Transmission of a highly pathogenic avian influenza virus to swine in the Netherlands. In Proceedings of the Internationale Society for Animal Hygiene, Saint Malo, France, 11–13 October 2004. [Google Scholar]
- Santé Publique France. Influenza activity in France, season 2016–2017. Bull. Epidémiol. Hebd. SPF 2017, n 22, 466–475. [Google Scholar]
- Yuk, S.S.; Lee, D.H.; Park, J.K.; Tseren-Ochir, E.O.; Kwon, J.H.; Noh, J.Y.; Song, C.S. Experimental infection of dogs with highly pathogenic avian influenza virus (H5N8). J. Vet. Sci. 2017, 18, 381–384. [Google Scholar] [CrossRef] [Green Version]
- Grund, C.; Hoffmann, D.; Ulrich, R.; Naguib, M.; Schinkothe, J.; Hoffmann, B.; Harder, T.; Saenger, S.; Zscheppang, K.; Tonnies, M.; et al. A novel European H5N8 influenza A virus has increased virulence in ducks but low zoonotic potential. Emerg. Microbes Infect. 2018, 7, 132. [Google Scholar] [CrossRef]
- Lee, Y.N.; Lee, E.K.; Song, B.M.; Heo, G.B.; Woo, S.H.; Cheon, S.H.; Lee, Y.J. Evaluation of the zoonotic potential of multiple subgroups of clade 2.3.4.4 influenza A (H5N8) virus. Virology 2018, 516, 38–45. [Google Scholar] [CrossRef] [PubMed]
- Adlhoch, C.; Dabrera, G.; Penttinen, P.; Pebody, R. Protective Measures for Humans against Avian Influenza A(H5N8) Outbreaks in 22 European Union/European Economic Area Countries and Israel, 2016–2017. Emerg. Infect. Dis. 2018, 24, 1–8. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Long, J.S.; Mistry, B.; Haslam, S.M.; Barclay, W.S. Host and viral determinants of influenza A virus species specificity. Nat. Rev. Microbiol. 2018. [Google Scholar] [CrossRef]
- Harris, K.A.; Freidl, G.S.; Munoz, O.S.; von Dobschuetz, S.; De Nardi, M.; Wieland, B.; Koopmans, M.P.G.; Stärk, K.D.C.; van Reeth, K.; Dauphin, G.; et al. Epidemiological Risk Factors for Animal Influenza A Viruses Overcoming Species Barriers. EcoHealth 2017, 14, 342–360. [Google Scholar] [CrossRef] [PubMed]
- Chastagner, A.; Bonin, E.; Fablet, C.; Quéguiner, S.; Hirchaud, E.; Lucas, P.; Gorin, S.; Barbier, N.; Béven, V.; Garin, E.; et al. Virus persistence in pig herds led to successive reassortment events between swine and human influenza A viruses, resulting in the emergence of a novel triple-reassortant swine influenza virus. Vet. Res. 2019, 50, 77. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cong, Y.; Wang, G.; Guan, Z.; Chang, S.; Zhang, Q.; Yang, G.; Wang, W.; Meng, Q.; Ren, W.; Wang, C.; et al. Reassortant between human-Like H3N2 and avian H5 subtype influenza A viruses in pigs: A potential public health risk. PLoS ONE 2010, 5, e12591. [Google Scholar] [CrossRef] [PubMed]
- Abente, E.J.; Kitikoon, P.; Lager, K.M.; Gauger, P.C.; Anderson, T.K.; Vincent, A.L. A highly pathogenic avian-derived influenza virus H5N1 with 2009 pandemic H1N1 internal genes demonstrates increased replication and transmission in pigs. J. Gen. Virol. 2017, 98, 18–30. [Google Scholar] [CrossRef] [Green Version]
- Yamaji, R.; Saad, M.D.; Davis, C.T.; Swayne, D.E.; Wang, D.; Wong, F.Y.K.; McCauley, J.W.; Peiris, J.S.M.; Webby, R.J.; Fouchier, R.A.M.; et al. Pandemic potential of highly pathogenic avian influenza clade 2.3.4.4 A(H5) viruses. Rev. Med Virol. 2020, 30, e2099. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xu, W.; Dai, Y.; Hua, C.; Wang, Q.; Zou, P.; Deng, Q.; Jiang, S.; Lu, L. Genomic signature analysis of the recently emerged highly pathogenic A(H5N8) avian influenza virus: Implying an evolutionary trend for bird-to-human transmission. Microbes Infect. 2017, 19, 597–604. [Google Scholar] [CrossRef]
- WHO World Health Organization. Human Infection with Avian Influenza A (H5N8)—the Russian Federation. Available online: https://www.who.int/csr/don/26-feb-2021-influenza-a-russian-federation/en/ (accessed on 18 March 2021).
- Scoizec, A.; Niqueux, E.; Thomas, R.; Daniel, P.; Schmitz, A.; Le Bouquin, S. Airborne Detection of H5N8 Highly Pathogenic Avian Influenza Virus Genome in Poultry Farms, France. Front. Vet. Sci. 2018, 5. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schmitz, A.; Pertusa, M.; Le Bouquin, S.; Rousset, N.; Ogor, K.; LeBras, M.O.; Martenot, C.; Daniel, P.; Belen Cepeda Hontecillas, A.; Scoizec, A.; et al. Natural and Experimental Persistence of Highly Pathogenic H5 Influenza Viruses in Slurry of Domestic Ducks, with or without Lime Treatment. Appl. Environ. Microbiol. 2020, 86. [Google Scholar] [CrossRef] [PubMed]


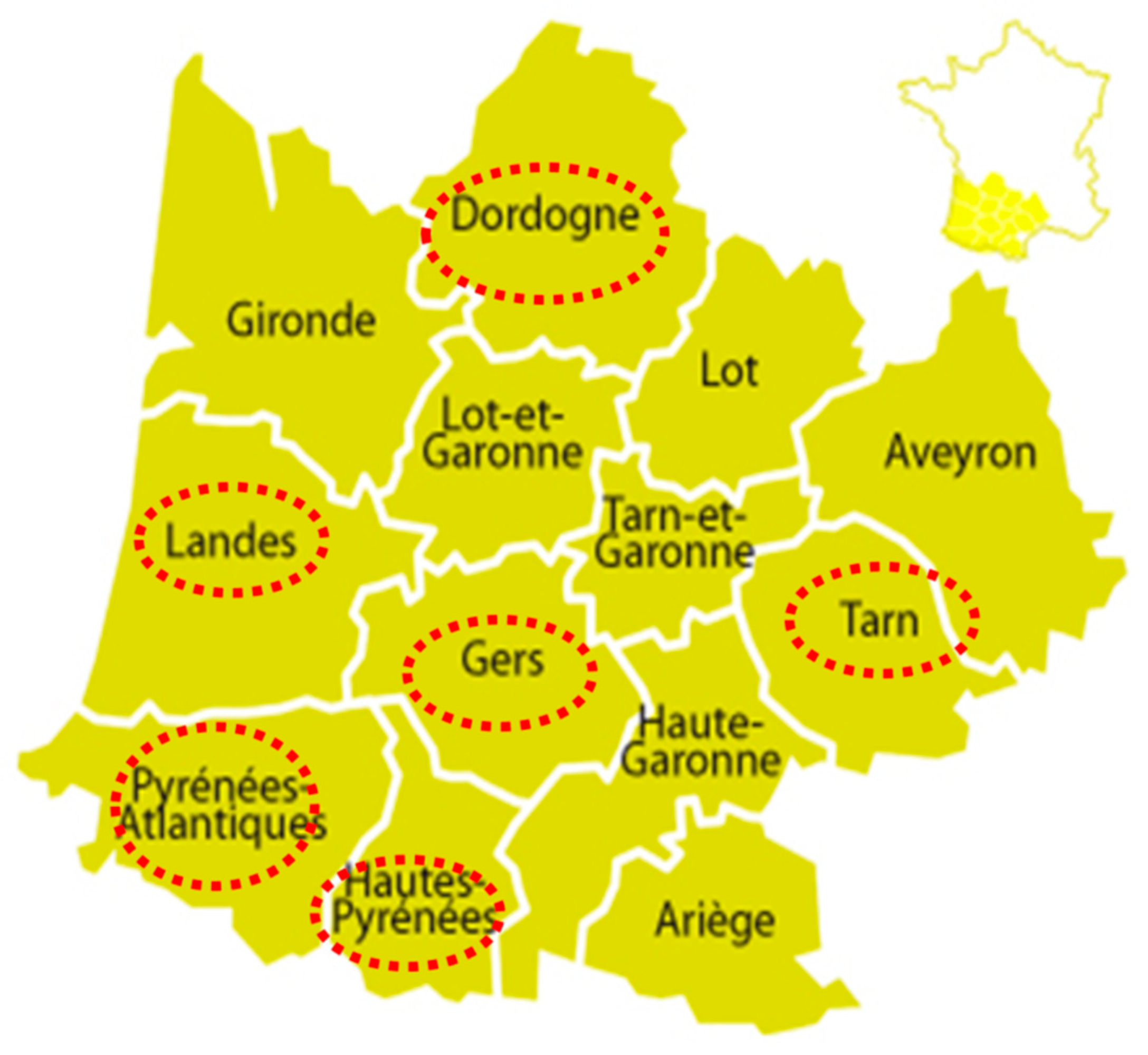{kind=link}
| Herd | Avian Influenza Outbreak | Sampled Pigs | ELISA Results 1 | |||||
|---|---|---|---|---|---|---|---|---|
| ID | Département of Location | HPAI-H5Nx Confirmed Case (Date) | Affected Poultry Species | Type of Pig Breeding | Herd Size | Physiological Stage or Age | Sampling Date (m/d/y) | No. Positive/No. Tested |
| #1 | Dordogne | NA | NA | outdoor farrowing-to-finishing | 134 sows, 500 post-weaning, 2000 finishing | sows | 02/01/2017 | 10/10 |
| #2 | Tarn | H5N8 (12/1/2016) | ducks | outdoor finishing (backyard) | 2 | growing pigs | 12/23/2016 | 1/2 |
| 02/01/2017 | 2/2 | |||||||
| 05/31/2017 | 1/1 | |||||||
| #3 | Tarn | H5N8 (12/2/2016) | ducks, geese | finishing | 185 | 12 weeks | 12/27/2016 | 0/28 |
| 16 weeks | 01/27/2017 | 0/15 | ||||||
| #4 | Gers | H5Nx (12/22/2016) | nd | finishing | nd | 1 year | 01/02/2017 | 0/1 |
| 01/27/2017 | 0/1 | |||||||
| #5 | Gers | H5N8 (12/1/2016) | nd | nd | nd | nd | 01/16/2017 | 0/2 |
| #6 | Gers | H5N8 (12/22/2016) | ducks | nd | nd | growing pigs | 12/29/2016 | 0/5 |
| #7 | Pyrénées Atlantiques | H5Nx (nd) | nd | nd | nd | 12 weeks | 02/02/2017 | 0/60 |
| #8 | Hautes Pyrénées | H5N8 (12/26/2016) | ducks | post-weaning | nd | 7 weeks | 12/29/2016 | 0/60 |
| #9 | Landes | H5N8 (2/13/2017) | ducks, geese | outdoor post weaning-finishing | nd | 10 weeks | 05/09/2017 | 13/30(A) |
| 13 weeks | 05/29/2017 | 9/30(A) | ||||||
| 17 weeks | 05/09/2017 | 0/30(B) | ||||||
| 20 weeks | 05/29/2017 | 0/30(B) | ||||||
| #10 | Landes | H5N8 (12/27/2016) | ducks, geese | post weaning-finishing | nd | 24 weeks | 05/12/2017 | 0/60 |
| Antigens 1 | HI Titers of Swine Hyperimmune Sera Containing Antibodies Directed Against | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| Swine Influenza A Virus 1 | Avian Influenza A Virus 1 | |||||||||
| HA-Clade [34,35] | 1C.2.1 | 1C.2 | 1A.3.3.2 | 1B.1 | 1B.1.2.3 | H3 | H5-2.3.4.4 | H5-Others | ||
| Subtype | H1avN1 | H1avN2 | H1N1pdm | H1huN2 | H1huN2Δ146–147 | H3N2 | H5N8 | H5N9 | H5N3 | |
| Swine influenza A virus | ||||||||||
| A/sw/Cotes d’Armor/0388/2009 (H1avN1) | 640 | 20 | <10 | <10 | <10 | <10 | <10 £ | <10 £ | <10 £ | |
| A/sw/France/65-150242/2015 (H1avN2) | 80 £ | 1280 £ | <10 £ | <10 £ | <10 £ | <10 £ | nd | nd | nd | |
| A/sw/Sarthe/0255/2010 (H1N1pdm) | 10 | nd | 640 | 10 | <10 | <10 | nd | nd | nd | |
| A/sw/France/57-140136/2014 (H1N1pdm) | 20 | <10 £ | 640 | 20 | <10 | <10 | <10 £ | <10 £ | <10 £ | |
| A/sw/Scotland/410440/1994 (H1huN2) | <10 | <10 £ | 10 | 1280 | 40 | 10 | <10 £ | <10 £ | <10 £ | |
| A/sw/France/22-130212/2013 (H1huN2Δ146–147) | 10 | 10 £ | <10 | 80 | 1280 | 10 | <10 £ | <10 £ | <10 £ | |
| A/sw/Flandres/1/1998 (H3N2) | <10 | 10 £ | <10 | <10 | <10 | 2560 | <10 £ | <10 £ | <10 £ | |
| Avian influenza A virus | ||||||||||
| A/decoyduck/France/161105a/2016 (H5N8) | <10 £ | nd | <10 £ | <10 £ | <10 £ | <10 £ | 50 | <10 | <10 | |
| A/muteswan/Croatia/102/2016 (H5N5) | nd | nd | nd | nd | nd | nd | 20 £ | <10 £ | <10 £ | |
| A/duck/France/150236b/2015 (H5N9) | 10 £ | nd | <10 £ | <10 £ | <10 £ | <10 £ | <10 | 25 | 120 | |
| A/muscovyduck/France/070090b/2007 (H5N3) | <10 £ | nd | <10 £ | <10 £ | <10 £ | <10 £ | <10 | 10 | 160 | |
| A/mallard/France/100204f/2010 (H3N8) | nd | nd | nd | nd | nd | nd | <10 £ | <10 £ | <10 £ | |
| A/pekinduck/France/090173/2009 (H6N8) | nd | nd | nd | nd | nd | nd | <10 £ | <10 £ | <10 £ | |
| Human influenza A virus | ||||||||||
| A/Victoria/361/2011 (H3N2) | <10 £ | nd | <10 £ | 10 £ | nd | <10 £ | <10 £ | <10 £ | <10 £ | |
| Herd ID | Sampling Date | Pig ID | ELISA S/P 2 | HI Titres Obtained with Influenza A Virus Strains 1 of Different Origins, Subtypes and Lineages | ||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Swine IAV | Avian IAV | Human IAV | ||||||||||||||
| HA Clade | 1C.2.1 | 1A.3.3.2 | 1B.1 | 1B.1.2.3 | H3 | H5-2.3.4.4 | H5-Others | Hx(N8) | H3 | |||||||
| Subtype | H1avN1 | H1N1pdm | H1huN2 | H1huN2 Δ146–147 | H3N2 | H5N8 | H5N5 | H5N9 | H5N3 | H3N8 | H6N8 | H3N2 | ||||
| #1 | 01 Feb | #1 | 21.8 | 10 | <10 | <10 | <10 | <10 | <10 | nd | <10 | <10 | nd | nd | nd | |
| #2 | 4.7 | 80 | 80 | <10 | <10 | <10 | <10 | nd | <10 | <10 | nd | nd | nd | |||
| #3 | 13.0 | 20 | <10 | <10 | <10 | <10 | <10 | nd | <10 | <10 | nd | nd | nd | |||
| #4 | 18.8 | 20 | 40 | <10 | <10 | <10 | <10 | nd | <10 | <10 | nd | nd | nd | |||
| #5 | 22.3 | <10 | <10 | <10 | <10 | <10 | <10 | nd | <10 | <10 | nd | nd | nd | |||
| #6 | 5.1 | 20 | 20 | <10 | <10 | <10 | <10 | nd | <10 | <10 | nd | nd | nd | |||
| #7 | 5.9 | 40 | 80 | <10 | <10 | <10 | <10 | nd | <10 | <10 | nd | nd | nd | |||
| #8 | 18.2 | <10 | 20 | <10 | <10 | <10 | <10 | nd | <10 | <10 | nd | nd | nd | |||
| #9 | 5.4 | 40 | 160 | <10 | <10 | <10 | <10 | nd | <10 | <10 | nd | nd | nd | |||
| #10 | 5.4 | 10 | 10 | <10 | <10 | <10 | <10 | nd | <10 | <10 | nd | nd | nd | |||
| #2 | 23 Dec | #1 | 10.2 | <10 | <10 | <10 | <10 | <10 | 40 | 20 | <10 | <10 | <10 | <10 | 10 | |
| 23 Dec | #2 | 58.5 | <10 | <10 | <10 | <10 | <10 | <10 | <10 | <10 | <10 | <10 | <10 | 20 | ||
| 01 Feb | #1 | 24.8 | <10 | <10 | <10 | <10 | <10 | 40 | 20 | <10 | <10 | <10 | <10 | <10 | ||
| 01 Feb | #2 | 9.1 | <10 | <10 | <10 | <10 | <10 | <10 | <10 | <10 | <10 | <10 | <10 | 20 | ||
| 31 May | nd | 41.7 | <10 | <10 | <10 | nd | <10 | <10 | nd | <10 | <10 | nd | nd | 10 | ||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hervé, S.; Schmitz, A.; Briand, F.-X.; Gorin, S.; Quéguiner, S.; Niqueux, É.; Paboeuf, F.; Scoizec, A.; Le Bouquin-Leneveu, S.; Eterradossi, N.; et al. Serological Evidence of Backyard Pig Exposure to Highly Pathogenic Avian Influenza H5N8 Virus during 2016–2017 Epizootic in France. Pathogens 2021, 10, 621. https://0-doi-org.brum.beds.ac.uk/10.3390/pathogens10050621
Hervé S, Schmitz A, Briand F-X, Gorin S, Quéguiner S, Niqueux É, Paboeuf F, Scoizec A, Le Bouquin-Leneveu S, Eterradossi N, et al. Serological Evidence of Backyard Pig Exposure to Highly Pathogenic Avian Influenza H5N8 Virus during 2016–2017 Epizootic in France. Pathogens. 2021; 10(5):621. https://0-doi-org.brum.beds.ac.uk/10.3390/pathogens10050621
Chicago/Turabian StyleHervé, Séverine, Audrey Schmitz, François-Xavier Briand, Stéphane Gorin, Stéphane Quéguiner, Éric Niqueux, Frédéric Paboeuf, Axelle Scoizec, Sophie Le Bouquin-Leneveu, Nicolas Eterradossi, and et al. 2021. "Serological Evidence of Backyard Pig Exposure to Highly Pathogenic Avian Influenza H5N8 Virus during 2016–2017 Epizootic in France" Pathogens 10, no. 5: 621. https://0-doi-org.brum.beds.ac.uk/10.3390/pathogens10050621