
Fox Serum Proteomics Analysis Suggests Host-Specific Responses to Angiostrongylus vasorum Infection in Canids

 and
and
Abstract
:
1. Introduction
2. Results
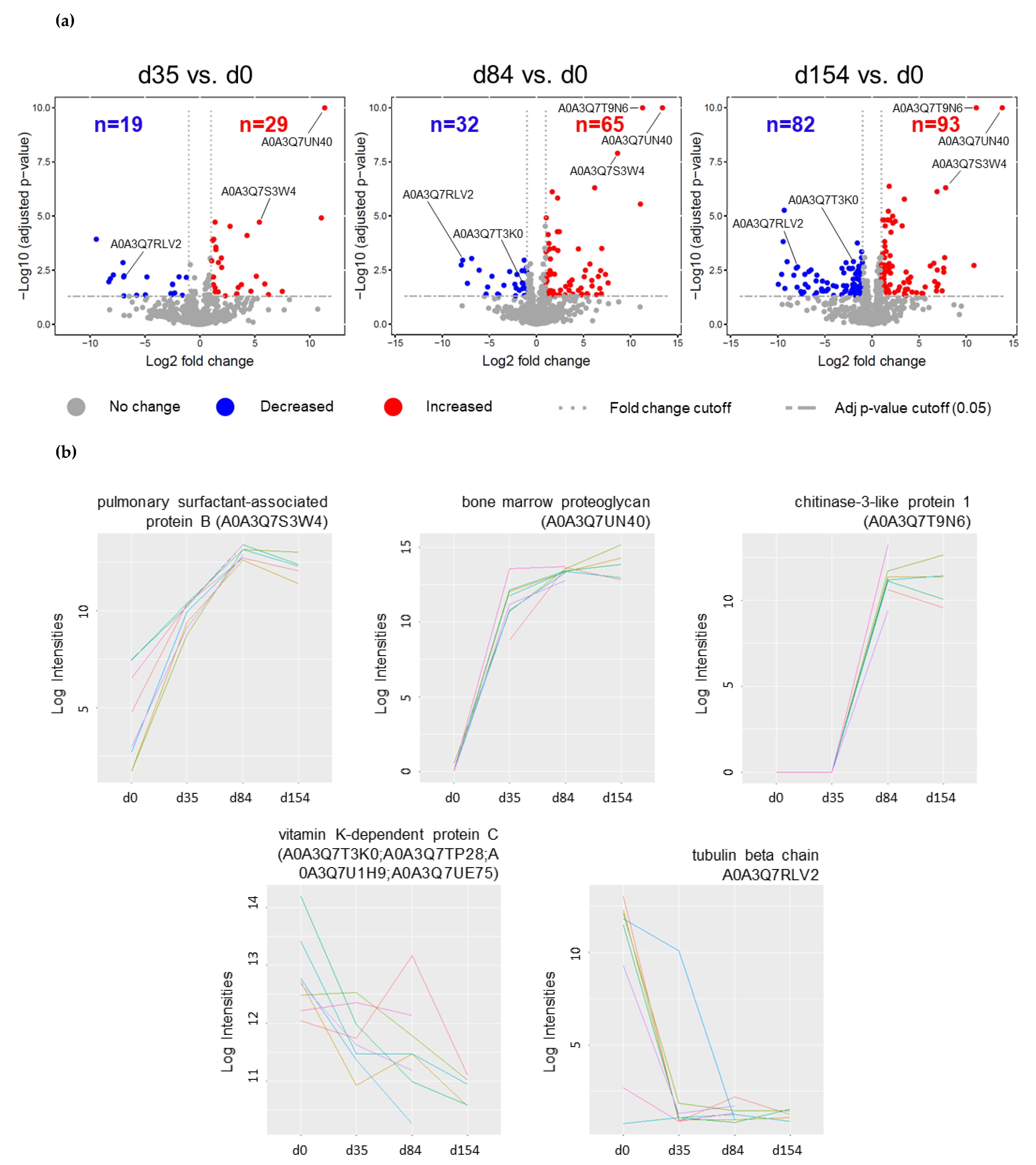
2.1. Differentially Abundant Proteins
2.2. Gene Ontology and Pathway Enrichment
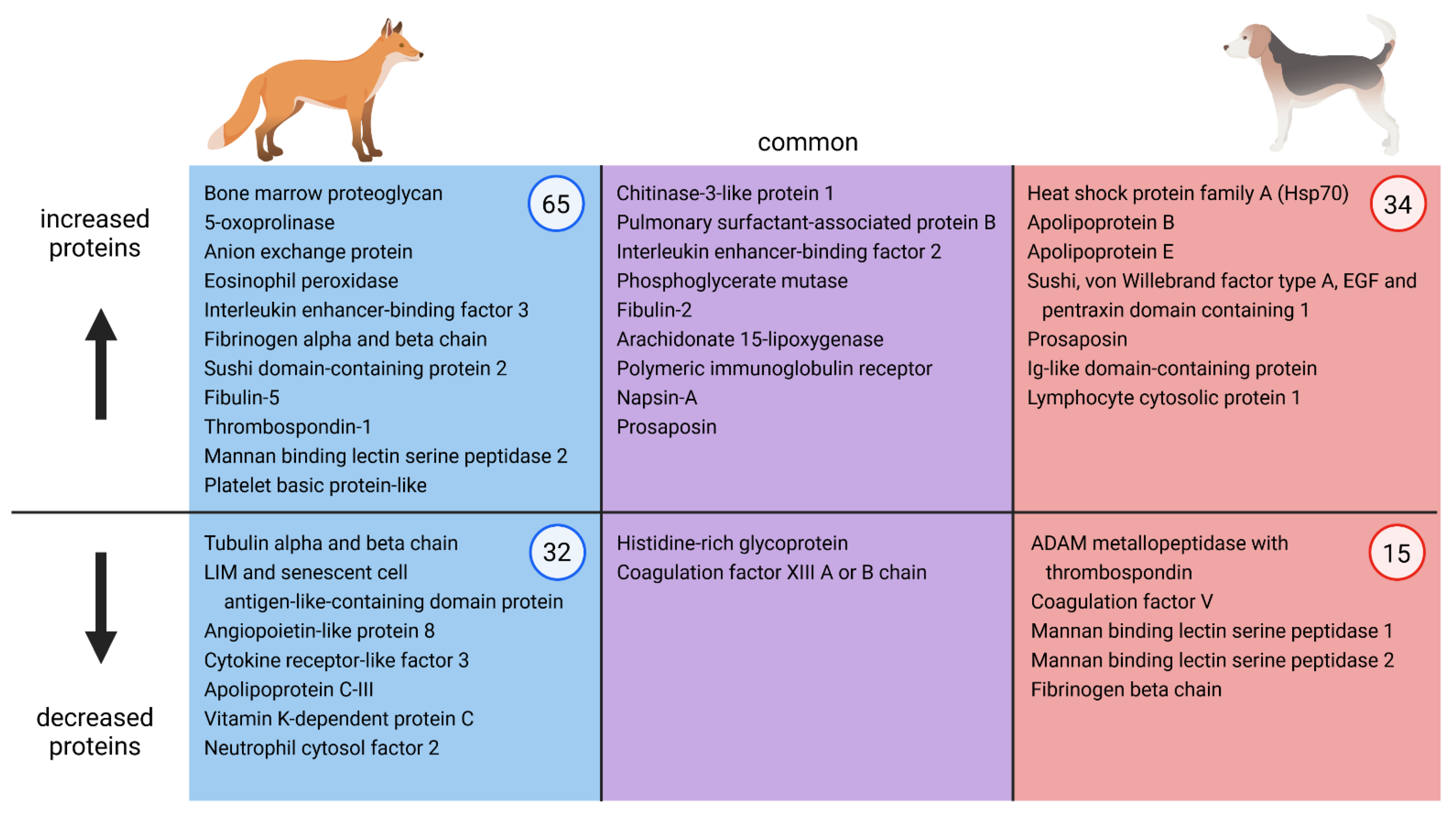
2.3. Qualitative Comparison of Serum Alterations of A. vasorum Experimentally Infected Foxes and Dogs
3. Discussion
4. Materials and Methods
4.1. Sera from Foxes Experimentally Inoculated with A. vasorum
4.2. Serum Sample Preparation
4.3. LC-MS Analysis
4.4. Data Analysis
4.5. Qualitative Comparison of Serum Alterations of Experimentally Infected Foxes and Dogs
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Guilhon, J.; Cens, B. Angiostrongylus vasorum (Baillet, 1866): Étude biologique et morphologique. Ann. Parasitol. Hum. Comp. 1973, 48, 567–596. [Google Scholar] [CrossRef] [Green Version]
- Koch, J.; Willesen, J.L. Canine pulmonary angiostrongylosis: An update. Vet. J. 2009, 179, 348–359. [Google Scholar] [CrossRef]
- Conboy, G.A. Canine angiostrongylosis: The French heartworm: An emerging threat in North America. Vet. Parasitol. 2011, 176, 382–389. [Google Scholar] [CrossRef] [PubMed]
- Penagos-Tabares, F.; Lange, M.K.; Chaparro-Gutiérrez, J.J.; Taubert, A.; Hermosilla, C. Angiostrongylus vasorum and Aelurostrongylus abstrusus: Neglected and underestimated parasites in South America. Parasite Vector 2018, 11, 208. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gillis-Germitsch, N.; Tritten, L.; Hegglin, D.; Deplazes, P.; Schnyder, M. Conquering Switzerland: The emergence of Angiostrongylus vasorum in foxes over three decades and its rapid regional increase in prevalence contrast with the stable occurrence of lungworms. Parasitology 2020, 147, 1071–1079. [Google Scholar] [CrossRef] [PubMed]
- Taylor, C.S.; Garcia Gato, R.; Learmount, J.; Aziz, N.A.; Montgomery, C.; Rose, H.; Coulthwaite, C.L.; McGarry, J.W.; Forman, D.W.; Allen, S.; et al. Increased prevalence and geographic spread of the cardiopulmonary nematode Angiostrongylus vasorum in fox populations in Great Britain. Parasitology 2015, 142, 1190–1195. [Google Scholar] [CrossRef]
- Maksimov, P.; Hermosilla, C.; Taubert, A.; Staubach, C.; Sauter-Louis, C.; Conraths, F.J.; Vrhovec, M.G.; Pantchev, N. GIS-supported epidemiological analysis on canine Angiostrongylus vasorum and Crenosoma vulpis infections in Germany. Parasite Vector 2017, 10, 108. [Google Scholar] [CrossRef] [Green Version]
- Hurnikova, Z.; Miterpakova, M.; Mandelik, R. First autochthonous case of canine Angiostrongylus vasorum in Slovakia. Parasitol. Res. 2013, 112, 3505–3508. [Google Scholar] [CrossRef]
- Jolly, S.; Poncelet, L.; Lempereur, L.; Caron, Y.; Bayrou, C.; Cassart, D.; Grimm, F.; Losson, B. First report of a fatal autochthonous canine Angiostrongylus vasorum infection in Belgium. Parasitol. Int. 2015, 64, 97–99. [Google Scholar] [CrossRef] [Green Version]
- Traversa, D.; Di Cesare, A.; Meloni, S.; di Regalbono, A.F.; Milillo, P.; Pampurini, F.; Venco, L. Canine angiostrongylosis in Italy: Occurrence of Angiostrongylus vasorum in dogs with compatible clinical pictures. Parasitol. Res. 2013, 112, 2473–2480. [Google Scholar] [CrossRef] [Green Version]
- Bourque, A.; Whitney, H.; Conboy, G. Angiostrongylus vasorum infection in a coyote (Canis latrans) from Newfoundland and Labrador, Canada. J. Wildl. Dis. 2005, 41, 816–819. [Google Scholar] [CrossRef] [Green Version]
- Duarte, F.; Vieira, F.; Louzada, G.; Bessa, E.; Souzalima, S. Occurrence Angiostrongylus vasorum (Baillet, 1866) (Nematoda, Angiostrongylidae) in Cerdocyon thous Linnaeus, 1766 (Carnivora, Canidae) in Minas Gerais State Brazil. Arq. Bras. Med. Vet. Zootec. 2007, 59, 1086–1088. [Google Scholar] [CrossRef] [Green Version]
- Lima, W.; Guimaraes, M.; Lemos, I. Research Note Occurrence of Angiostrongylus vasorum in the lungs of the Brazilian fox Dusicyon vetulus. J. Helminthol. 1994, 68, 87. [Google Scholar] [CrossRef]
- Segovia, J.; Torres, J.; Miquel, J.; Llaneza, L.; Feliu, C. Helminths in the wolf, Canis lupus, from north-western Spain. J. Helminthol. 2001, 75, 183–192. [Google Scholar] [CrossRef] [PubMed]
- Patterson-Kane, J.C.; Gibbons, L.M.; Jefferies, R.; Morgan, E.R.; Wenzlow, N.; Redrobe, S.P. Pneumonia from Angiostrongylus vasorum infection in a red panda (Ailurus fulgens fulgens). J. Vet. Diagn. Investig. 2009, 21, 270–273. [Google Scholar] [CrossRef] [Green Version]
- Torres, J.; Miquel, J.; Motjé, M. Helminth parasites of the eurasian badger (Meles meles L.) in Spain: A biogeographic approach. Parasitol. Res. 2001, 87, 259–263. [Google Scholar] [CrossRef]
- Gillis-Germitsch, N.; Manser, M.B.; Hilbe, M.; Schnyder, M. Meerkats (Suricata suricatta), a new definitive host of the canid nematode Angiostrongylus vasorum. Int. J. Parasitol. Parasites Wildl. 2017, 6, 349–353. [Google Scholar] [CrossRef] [PubMed]
- Chapman, P.S.; Boag, A.K.; Guitian, J.; Boswood, A. Angiostrongylus vasorum infection in 23 dogs (1999–2002). J. Small Anim. Pract. 2004, 45, 435–440. [Google Scholar] [CrossRef] [PubMed]
- Schnyder, M.; Fahrion, A.; Riond, B.; Ossent, P.; Webster, P.; Kranjc, A.; Glaus, T.; Deplazes, P. Clinical, laboratory and pathological findings in dogs experimentally infected with Angiostrongylus vasorum. Parasitol. Res. 2010, 107, 1471–1480. [Google Scholar] [CrossRef] [Green Version]
- Glaus, T.M.; Sigrist, N.; Hofer-Inteeworn, N.; Kuemmerle-Fraune, C.; Mueller, C.; Geissweid, K.; Beckmann, K.; Wenger, M.; Novo Matos, J. Unexplained bleeding as primary clinical complaint in dogs infected with Angiostrongylus vasorum. Schweiz. Arch. Tierheilkd. 2016, 158, 701–709. [Google Scholar] [CrossRef] [Green Version]
- Adamantos, S.; Waters, S.; Boag, A. Coagulation status in dogs with naturally occurring Angiostrongylus vasorum infection. J. Small Anim. Pract. 2015, 56, 485–490. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sigrist, N.E.; Hofer-Inteeworn, N.; Jud Schefer, R.; Kuemmerle-Fraune, C.; Schnyder, M.; Kutter, A.P.N. Hyperfibrinolysis and hypofibrinogenemia diagnosed with rotational thromboelastometry in dogs naturally infected with Angiostrongylus vasorum. J. Vet. Intern. Med. 2017, 31, 1091–1099. [Google Scholar] [CrossRef] [Green Version]
- Gredal, H.; Willesen, J.L.; Jensen, H.E.; Nielsen, O.L.; Kristensen, A.T.; Koch, J.; Kirk, R.K.; Pors, S.E.; Skerritt, G.C.; Berendt, M. Acute neurological signs as the predominant clinical manifestation in four dogs with Angiostrongylus vasorum infections in Denmark. Act. Vet. Scand. 2011, 53, 43. [Google Scholar] [CrossRef] [Green Version]
- Simpson, V.R. Angiostrongylus vasorum infection in foxes (Vulpes vulpes) in Cornwall. Vet. Rec. 1996, 139, 443–445. [Google Scholar] [CrossRef]
- Philbey, A.W.; Delgado, D. Detection of Angiostrongylus vasorum in red foxes in Scotland. Vet. Rec. 2013, 173. [Google Scholar] [CrossRef]
- Webster, P.; Monrad, J.; Kapel, C.M.O.; Kristensen, A.T.; Jensen, A.L.; Thamsborg, S.M. The effect of host age and inoculation dose on infection dynamics of Angiostrongylus vasorum in red foxes (Vulpes vulpes). Parasite Vector 2017, 10, 4. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Woolsey, I.D.; Webster, P.; Thamsborg, S.; Schnyder, M.; Monrad, J.; Kapel, C.M.O. Repeated inoculations with the lung and heartworm nematode Angiostrongylus vasorum result in increasing larval excretion and worm burden in the red fox (Vulpes vulpes). Int. J. Parasitol. Parasites Wildl. 2017, 6, 139–145. [Google Scholar] [CrossRef] [PubMed]
- Poli, A.; Arispici, M.; Mancianti, F.; Abramo, F. Pathology of naturally acquired Angiostrongylus vasorum infection in the red fox (Vulpes vulpes). Angew. Parasitol. 1991, 32, 121–126. [Google Scholar]
- Bourque, A.C.; Conboy, G.; Miller, L.M.; Whitney, H. Pathological findings in dogs naturally infected with Angiostrongylus vasorum in Newfoundland and Labrador, Canada. J. Vet. Diagn. Investig. 2008, 20, 11–20. [Google Scholar] [CrossRef] [Green Version]
- McEvoy, F.J.; Buelund, L.; Strathe, A.B.; Willesen, J.L.; Koch, J.; Webster, P.I.A.; Hørlyck, L.; Svalastoga, E. Quantitative computed tomography evaluation of pulmonary disease. Vet. Radiol. Ultrasound 2009, 50, 47–51. [Google Scholar] [CrossRef]
- Dennler, M.; Makara, M.; Kranjc, A.; Schnyder, M.; Ossent, P.; Deplazes, P.; Ohlerth, S.; Glaus, T.M. Thoracic computed tomography findings in dogs experimentally infected with Angiostrongylus vasorum. Vet. Radiol. Ultrasound 2011, 52, 289–294. [Google Scholar] [CrossRef] [PubMed]
- Tritten, L.; Gillis-Germitsch, N.; Kockmann, T.; Schnyder, M. Quantitative proteomics analysis of Angiostrongylus vasorum-induced alterations in dog serum sheds light on the pathogenesis of canine angiostrongylosis. Sci. Rep. 2021, 11, 283. [Google Scholar] [CrossRef] [PubMed]
- Cury, M.C.; Lima, W.S.; Guimarães, M.P.; Carvalho, M.G. Hematological and coagulation profiles in dogs experimentally infected with Angiostrongylus vasorum (Baillet, 1866). Vet. Parasitol. 2002, 104, 139–149. [Google Scholar] [CrossRef]
- Huang, L.; Appleton, J.A. Eosinophils in Helminth Infection: Defenders and Dupes. Trends Parasitol. 2016, 32, 798–807. [Google Scholar] [CrossRef] [Green Version]
- Popken-Harris, P.; Checkel, J.; Loegering, D.; Madden, B.; Springett, M.; Kephart, G.; Gleich, G.J. Regulation and processing of a precursor form of eosinophil granule major basic protein (ProMBP) in differentiating eosinophils. Blood 1998, 92, 623–631. [Google Scholar] [CrossRef]
- Thomas, L.L.; Haskell, M.D.; Sarmiento, E.U.; Bilimoria, Y. Distinguishing features of basophil and neutrophil activation by major basic protein. J. Allergy Clin. Immunol. 1994, 94, 1171–1176. [Google Scholar] [CrossRef]
- Rohrbach, M.S.; Wheatley, C.; Slifman, N.; Gleich, G. Activation of platelets by eosinophil granule proteins. J. Exp. Med. 1990, 172, 1271–1274. [Google Scholar] [CrossRef] [Green Version]
- Willesen, J.L.; Jensen, A.L.; Kristensen, A.T.; Koch, J. Haematological and biochemical changes in dogs naturally infected with Angiostrongylus vasorum before and after treatment. Vet. J. 2009, 180, 106–111. [Google Scholar] [CrossRef]
- Zhao, T.; Su, Z.; Li, Y.; Zhang, X.; You, Q. Chitinase-3 like-protein-1 function and its role in diseases. Signal. Transduct. Target. Ther. 2020, 5, 201. [Google Scholar] [CrossRef]
- Sutherland, T.E.; Logan, N.; Rückerl, D.; Humbles, A.A.; Allan, S.M.; Papayannopoulos, V.; Stockinger, B.; Maizels, R.M.; Allen, J.E. Chitinase-like proteins promote IL-17-mediated neutrophilia in a tradeoff between nematode killing and host damage. Nat. Immunol. 2014, 15, 1116–1125. [Google Scholar] [CrossRef] [Green Version]
- Han, S.; Mallampalli, R.K. The role of surfactant in lung disease and host defense against pulmonary infections. Ann. Am. Thorac. Soc. 2015, 12, 765–774. [Google Scholar] [CrossRef] [Green Version]
- Lawler, J. The functions of thrombospondin-1 and-2. Curr. Opin. Cell Biol. 2000, 12, 634–640. [Google Scholar] [CrossRef]
- Fuller, G.M.; Zhang, Z. Transcriptional Control Mechanism of Fibrinogen Gene Expression. Ann. N. Y. Acad. Sci. 2001, 936, 469–479. [Google Scholar] [CrossRef]
- Yan, M.T.S.; Rydz, N.; Goodyear, D.; Sholzberg, M. Acquired factor XIII deficiency: A review. Transfus. Apher. Sci. 2018, 57, 724–730. [Google Scholar] [CrossRef]
- Thomas, A.; Biswas, A.; Dodt, J.; Philippou, H.; Hethershaw, E.; Ensikat, H.J.; Ivaskevicius, V.; Oldenburg, J. Coagulation factor XIIIA subunit missense mutations affect structure and function at the various steps of Factor XIII action. Hum. Mutat. 2016, 37, 1030–1041. [Google Scholar] [CrossRef]
- Mebius, M.M.; van Genderen, P.J.J.; Urbanus, R.T.; Tielens, A.G.M.; de Groot, P.G.; van Hellemond, J.J. Interference with the host haemostatic system by Schistosomes. PLoS Pathog. 2013, 9, e1003781. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gillis-Germitsch, N.; Kapel, C.M.O.; Thamsborg, S.M.; Deplazes, P.; Schnyder, M. Host-specific serological response to Angiostrongylus vasorum infection in red foxes (Vulpes vulpes): Implications for parasite epidemiology. Parasitology 2017, 144, 1144–1153. [Google Scholar] [CrossRef]
- Schucan, A.; Schnyder, M.; Tanner, I.; Barutzki, D.; Traversa, D.; Deplazes, P. Detection of specific antibodies in dogs infected with Angiostrongylus vasorum. Vet. Parasitol. 2012, 185, 216–224. [Google Scholar] [CrossRef] [Green Version]
- Eleni, C.; Grifoni, G.; Di Egidio, A.; Meoli, R.; De Liberato, C. Pathological findings of Angiostrongylus vasorum infection in red foxes (Vulpes vulpes) from Central Italy, with the first report of a disseminated infection in this host species. Parasitol. Res. 2014, 113, 1247–1250. [Google Scholar] [CrossRef]
- Fischer, C.; Reperant, L.; Weber, J.; Hegglin, D.; Deplazes, P. Echinococcus multilocularis infections of rural, residential and urban foxes (Vulpes vulpes) in the canton of Geneva, Switzerland. Parasite 2005, 12, 339–346. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Coman, B. The age structure of a sample of red foxes (Vulpes vulpes) taken by hunters in Victoria. Wildl. Res. 1988, 15, 223–229. [Google Scholar] [CrossRef]
- Wiśniewski, J.R.; Zougman, A.; Nagaraj, N.; Mann, M. Universal sample preparation method for proteome analysis. Nat. Methods 2009, 6, 359–362. [Google Scholar] [CrossRef] [PubMed]
- Rappsilber, J.; Ishihama, Y.; Mann, M. Stop and go extraction tips for matrix-assisted laser desorption/ionization, nanoelectrospray, and LC/MS sample pretreatment in proteomics. Anal. Chem. 2003, 75, 663–670. [Google Scholar] [CrossRef]
- Vizcaíno, J.A.; Deutsch, E.W.; Wang, R.; Csordas, A.; Reisinger, F.; Rios, D.; Dianes, J.A.; Sun, Z.; Farrah, T.; Bandeira, N. ProteomeXchange provides globally coordinated proteomics data submission and dissemination. Nat. Biotechnol. 2014, 32, 223–226. [Google Scholar] [CrossRef]
- Choi, M.; Chang, C.-Y.; Clough, T.; Broudy, D.; Killeen, T.; MacLean, B.; Vitek, O. MSstats: An R package for statistical analysis of quantitative mass spectrometry-based proteomic experiments. Bioinformatics 2014, 30, 2524–2526. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huang, D.W.; Sherman, B.T.; Lempicki, R.A. Systematic and integrative analysis of large gene lists using DAVID bioinformatics resources. Nat. Protoc. 2009, 4, 44–57. [Google Scholar] [CrossRef] [PubMed]
- Kuleshov, M.V.; Jones, M.R.; Rouillard, A.D.; Fernandez, N.F.; Duan, Q.; Wang, Z.; Koplev, S.; Jenkins, S.L.; Jagodnik, K.M.; Lachmann, A.; et al. Enrichr: A comprehensive gene set enrichment analysis web server 2016 update. Nucleic Acids Res. 2016, 44, W90–W97. [Google Scholar] [CrossRef] [Green Version]



{kind=link}
{kind=link}
{kind=link}
| Rank | d35 vs. d0 | d84 vs. d0 | d154 vs. d0 | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Biological Process | Pathway | Biological Process | Pathway | Biological Process | Pathway | |||||||
| Term | Adjusted p-Value | Term | Adjusted p-Value | Term | Adjusted p-Value | Term | Adjusted p-Value | Term | Adjusted p-Value | Term | Adjusted p-Value | |
| 1 | antibacterial humoral response (GO:0019731) | 7.97 × 10−4 | Extracellular matrix organization | 0.0033 | neutrophil mediated immunity (GO:0002446) | 1.29 × 10−10 | Extracellular matrix organization | 1.54 × 10−6 | neutrophil mediated immunity (GO:0002446) | 2.32 × 10−13 | Extracellular matrix organization | 4.58 × 10−13 |
| 2 | extracellular matrix organization (GO:0030198) | 0.0016 | neutrophil activation involved in immune response (GO:0002283) | 1.70 × 10−10 | Elastic fibre formation | 1.28 × 10−4 | neutrophil activation involved in immune response (GO:0002283) | 2.93 × 10−13 | Elastic fibre formation | 1.86 × 10−5 | ||
| 3 | neutrophil mediated immunity (GO:0002446) | 0.0038 | neutrophil degranulation (GO:0043312) | 2.96 × 10−10 | Molecules associated with elastic fibres | 0.0011 | neutrophil degranulation (GO:0043312) | 4.92 × 10−13 | Molecules associated with elastic fibres | 9.80 × 10−5 | ||
| 4 | neutrophil activation involved in immune response (GO:0002283) | 0.0045 | extracellular matrix organization (GO:0030198) | 4.42 × 10−5 | Keratan sulphate degradation | 0.0023 | extracellular matrix organization (GO:0030198) | 9.29 × 10−11 | Metabolism of carbohydrates | 1.39 × 10−4 | ||
| 5 | neutrophil degranulation (GO:0043312) | 0.0056 | sulphur compound catabolic process (GO:0044273) | 0.0035 | Platelet degranulation | 0.0057 | antibacterial humoral response (GO:0019731) | 9.13 × 10−6 | Response to elevated platelet cytosolic Ca2+ | 1.41 × 10−4 | ||
| 6 | cell-matrix adhesion (GO:0007160) | 0.0061 | regulated exocytosis (GO:0045055) | 0.0041 | Response to elevated platelet cytosolic Ca2+ | 0.0059 | cell-matrix adhesion (GO:0007160) | 1.18 × 10−4 | Platelet degranulation | 1.54 × 10−4 | ||
| 7 | regulation of endothelial cell apoptotic process (GO:2000351) | 0.0088 | antibacterial humoral response (GO:0019731) | 0.0042 | Common Pathway of Fibrin Clot Formation | 0.0092 | defence response to bacterium (GO:0042742) | 0.0081 | Keratan sulphate degradation | 0.0034 | ||
| 8 | cellular macromolecular complex assembly (GO:0034622) | 0.0091 | keratan sulphate catabolic process (GO:0042340) | 0.0045 | Metabolism of proteins | 0.018 | platelet degranulation (GO:0002576) | 0.0092 | Axon guidance | 0.0035 | ||
| 9 | regulation of extrinsic apoptotic signalling pathway via death domain receptors (GO:1902041) | 0.010 | cellular protein metabolic process (GO:0044267) | 0.0097 | Keratan sulphate on keratin metabolism | 0.022 | keratan sulphate catabolic process (GO:0042340) | 0.010 | Non-integrin membrane-ECM interactions | 0.0059 | ||
| 10 | retina homeostasis (GO:0001895) | 0.010 | glycosaminoglycan catabolic process (GO:0006027) | 0.014 | Formation of Fibrin Clot (Clotting Cascade) | 0.037 | sulphur compound catabolic process (GO:0044273) | 0.011 | Degradation of the extracellular matrix | 0.016 | ||
| Rank | d35 vs. d0 | d84 vs. d0 | d154 vs. d0 | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Biological Process | Pathway | Biological Process | Pathway | Biological Process | Pathway | |||||||
| Term | Adjusted p-Value | Term | Adjusted p-Value | Term | Adjusted p-Value | Term | Adjusted p-Value | Term | Adjusted p-Value | Term | Adjusted p-Value | |
| 1 | positive regulation of establishment of protein localization to telomere (GO:1904851) | 7.26 × 10−5 | Prefoldin mediated transfer of substrate to CCT/TriC | 3.12 × 10−11 | positive regulation of establishment of protein localization to telomere (GO:1904851) | 4.79 × 10−4 | Prefoldin mediated transfer of substrate to CCT/TriC | 6.89 × 10−12 | regulation of protein localization to Cajal body (GO:1904869) | 3.05 × 10−5 | Prefoldin mediated transfer of substrate to CCT/TriC | 4.54 × 10−9 |
| 2 | regulation of protein localization to Cajal body (GO:1904869) | 7.27 × 10−5 | Formation of tubulin folding intermediates by CCT/TriC | 3.74 × 10−11 | regulation of protein localization to Cajal body (GO:1904869) | 4.79 × 10−4 | Formation of tubulin folding intermediates by CCT/TriC | 7.48 × 10−12 | positive regulation of protein localization to Cajal body (GO:1904871) | 6.10 × 10−5 | Formation of tubulin folding intermediates by CCT/TriC | 4.95 × 10−9 |
| 3 | regulation of establishment of protein localization to telomere (GO:0070203) | 7.78 × 10−5 | Cooperation of Prefoldin and TriC/CCT in actin and tubulin folding | 6.36 × 10−11 | regulation of establishment of protein localization to telomere (GO:0070203) | 5.12 × 10−4 | Cooperation of Prefoldin and TriC/CCT in actin and tubulin folding | 1.73 × 10−11 | positive regulation of telomerase RNA localization to Cajal body (GO:1904874) | 3.89 × 10−4 | Haemostasis | 6.69 × 10−9 |
| 4 | positive regulation of protein localization to chromosome, telomeric region (GO:1904816) | 8.55 × 10−5 | Chaperonin-mediated protein folding | 4.44 × 10−8 | positive regulation of protein localization to chromosome, telomeric region (GO:1904816) | 5.63 × 10−4 | Chaperonin-mediated protein folding | 3.99 × 10−8 | regulation of telomerase RNA localization to Cajal body (GO:1904872) | 4.32 × 10−4 | Cooperation of Prefoldin and TriC/CCT in actin and tubulin folding | 8.48 × 10−9 |
| 5 | positive regulation of establishment of protein localization (GO:1904951) | 9.50 × 10−5 | Protein folding | 5.17 × 10−8 | positive regulation of establishment of protein localization (GO:1904951) | 6.25 × 10−4 | Protein folding | 4.94 × 10−8 | positive regulation of protein localization to nucleus (GO:1900182) | 4.72 × 10−4 | Folding of actin by CCT/TriC | 1.09 × 10−5 |
| 6 | positive regulation of protein localization to Cajal body (GO:1904871) | 1.45 × 10−4 | Folding of actin by CCT/TriC | 1.55 × 10−5 | positive regulation of protein localization to Cajal body (GO:1904871) | 9.58 × 10−4 | Post-chaperonin tubulin folding pathway | 8.64 × 10−6 | platelet degranulation (GO:0002576) | 4.83 × 10−4 | Chaperonin-mediated protein folding | 1.54 × 10−5 |
| 7 | positive regulation of telomerase RNA localization to Cajal body (GO:1904874) | 1.68 × 10−4 | Cell-extracellular matrix interactions | 6.20 × 10−5 | positive regulation of telomerase RNA localization to Cajal body (GO:1904874) | 0.0011 | Folding of actin by CCT/TriC | 8.78 × 10−5 | regulation of lipoprotein lipase activity (GO:0051004) | 5.10 × 10−4 | Protein folding | 2.02 × 10−5 |
| 8 | regulation of telomerase RNA localization to Cajal body (GO:1904872) | 2.63 × 10−4 | Post-chaperonin tubulin folding pathway | 1.49 × 10−4 | regulation of telomerase RNA localization to Cajal body (GO:1904872) | 0.0017 | Cell-extracellular matrix interactions | 3.56 × 10−4 | hydrogen peroxide metabolic process (GO:0042743) | 5.83 × 10−4 | Platelet degranulation | 2.32 × 10−5 |
| 9 | positive regulation of telomere maintenance via telomerase (GO:0032212) | 0.0014 | Association of TriC/CCT with target proteins during biosynthesis | 7.78 × 10−4 | regulation of lipoprotein lipase activity (GO:0051004) | 0.0021 | Platelet degranulation | 0.0034 | regulated exocytosis (GO:0045055) | 7.10 × 10−4 | Response to elevated platelet cytosolic Ca2+ | 2.84 × 10−5 |
| 10 | positive regulation of telomere maintenance via telomere lengthening (GO:1904358) | 0.0015 | Cooperation of PDCL (PhLP1) and TRiC/CCT in G-protein beta folding | 8.78 × 10−4 | acylglycerol homeostasis (GO:0055090) | 0.0026 | Response to elevated platelet cytosolic Ca2+ | 0.0037 | chylomicron remodelling (GO:0034371) | 0.0019 | Cell-extracellular matrix interactions | 4.23 × 10−5 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gillis-Germitsch, N.; Kockmann, T.; Kapel, C.M.O.; Thamsborg, S.M.; Webster, P.; Tritten, L.; Schnyder, M. Fox Serum Proteomics Analysis Suggests Host-Specific Responses to Angiostrongylus vasorum Infection in Canids. Pathogens 2021, 10, 1513. https://0-doi-org.brum.beds.ac.uk/10.3390/pathogens10111513
Gillis-Germitsch N, Kockmann T, Kapel CMO, Thamsborg SM, Webster P, Tritten L, Schnyder M. Fox Serum Proteomics Analysis Suggests Host-Specific Responses to Angiostrongylus vasorum Infection in Canids. Pathogens. 2021; 10(11):1513. https://0-doi-org.brum.beds.ac.uk/10.3390/pathogens10111513
Chicago/Turabian StyleGillis-Germitsch, Nina, Tobias Kockmann, Christian M. O. Kapel, Stig M. Thamsborg, Pia Webster, Lucienne Tritten, and Manuela Schnyder. 2021. "Fox Serum Proteomics Analysis Suggests Host-Specific Responses to Angiostrongylus vasorum Infection in Canids" Pathogens 10, no. 11: 1513. https://0-doi-org.brum.beds.ac.uk/10.3390/pathogens10111513