
Secreted MbovP0145 Promotes IL-8 Expression through Its Interactive β-Actin and MAPK Activation and Contributes to Neutrophil Migration

 , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. In Silico Analyses
2.2. Cultivation of Bacterial Strains and Cell Lines
2.3. Cloning, Expression, and Purification of a Recombinant MbovP0145 Protein
2.4. Construction of Strain Complementary to M. bovis ΔMbov_0145
2.4.1. In Trans Complementation of M. bovis T6.93 via a Plasmid Carrying the Gene Mbov_0145
2.4.2. Western Blot Analysis of MbovP0145 Expression in M. bovis Strains
2.5. EBL Cells Either Infected with M. bovis or Treated with rMbovP0145
2.6. Analysis of IL-8 mRNA Expression with Quantitative Real-Time PCR
2.7. GST Pull-Down Assay to Identify Interactive Protein of MbovP0145
2.8. Immunoprecipitation Assay on Interaction between β-Actin and MbovP0145
2.9. Colocation Analysis with Confocal Laser Fluorescence Microscopy
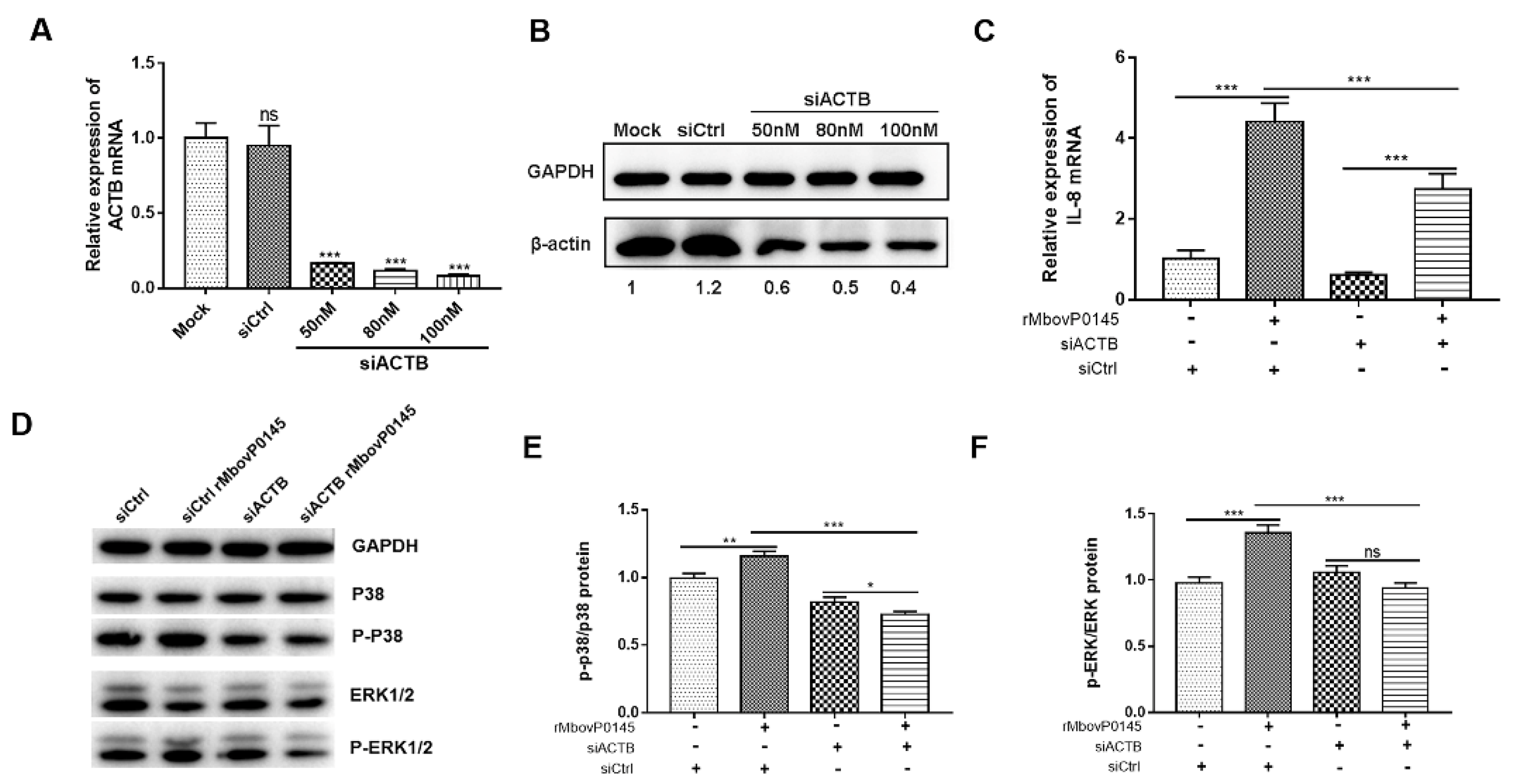
2.10. Knockdown Assay of β-Actin with siRNA Interference
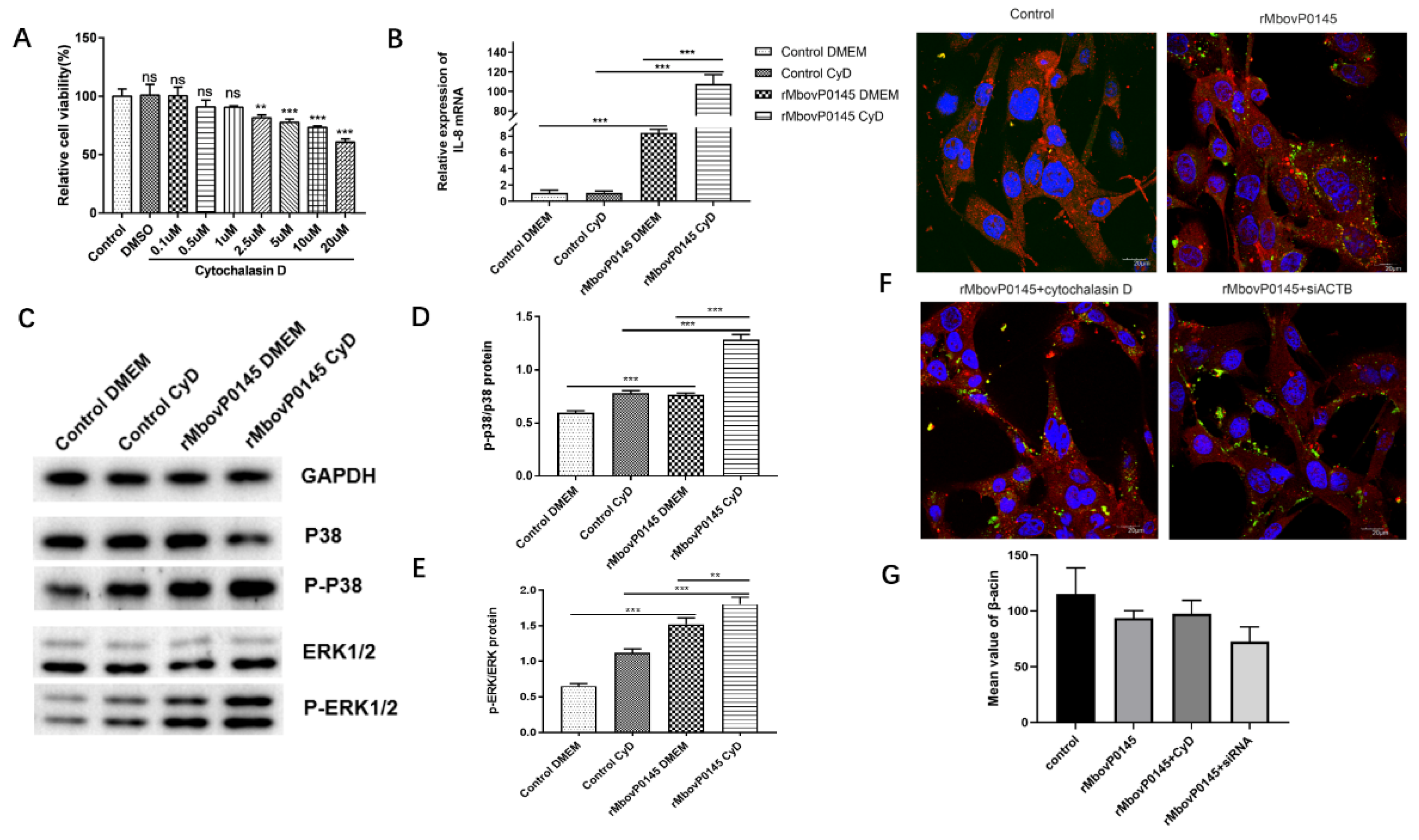
2.11. Inhibition of Actin Polymerization by Cytochalasin D
2.12. Analysis of the Molecules Critical to Signaling Pathways
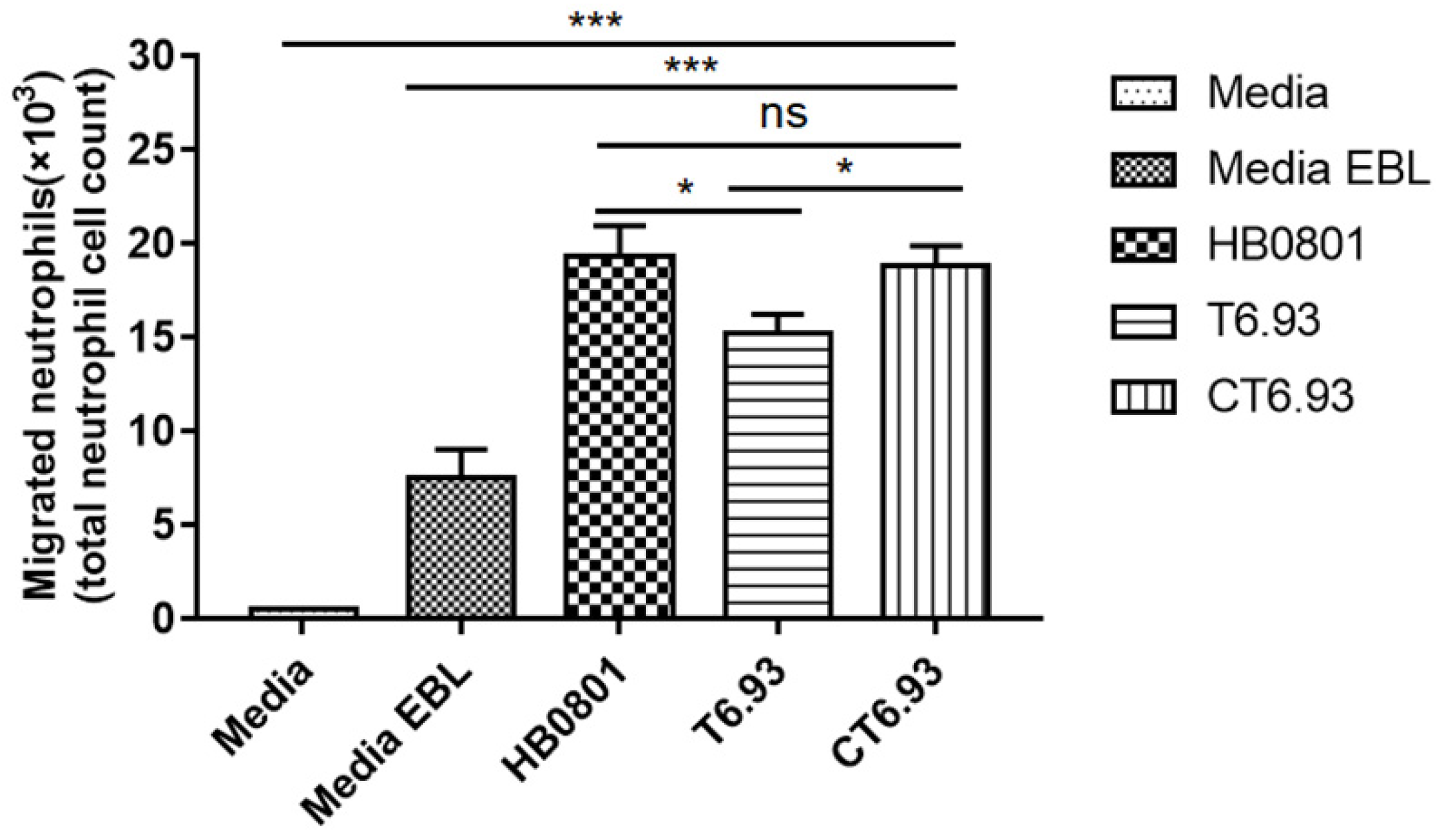
2.13. Neutrophil Chemotaxis Assay
2.14. Statistical Analysis
3. Results
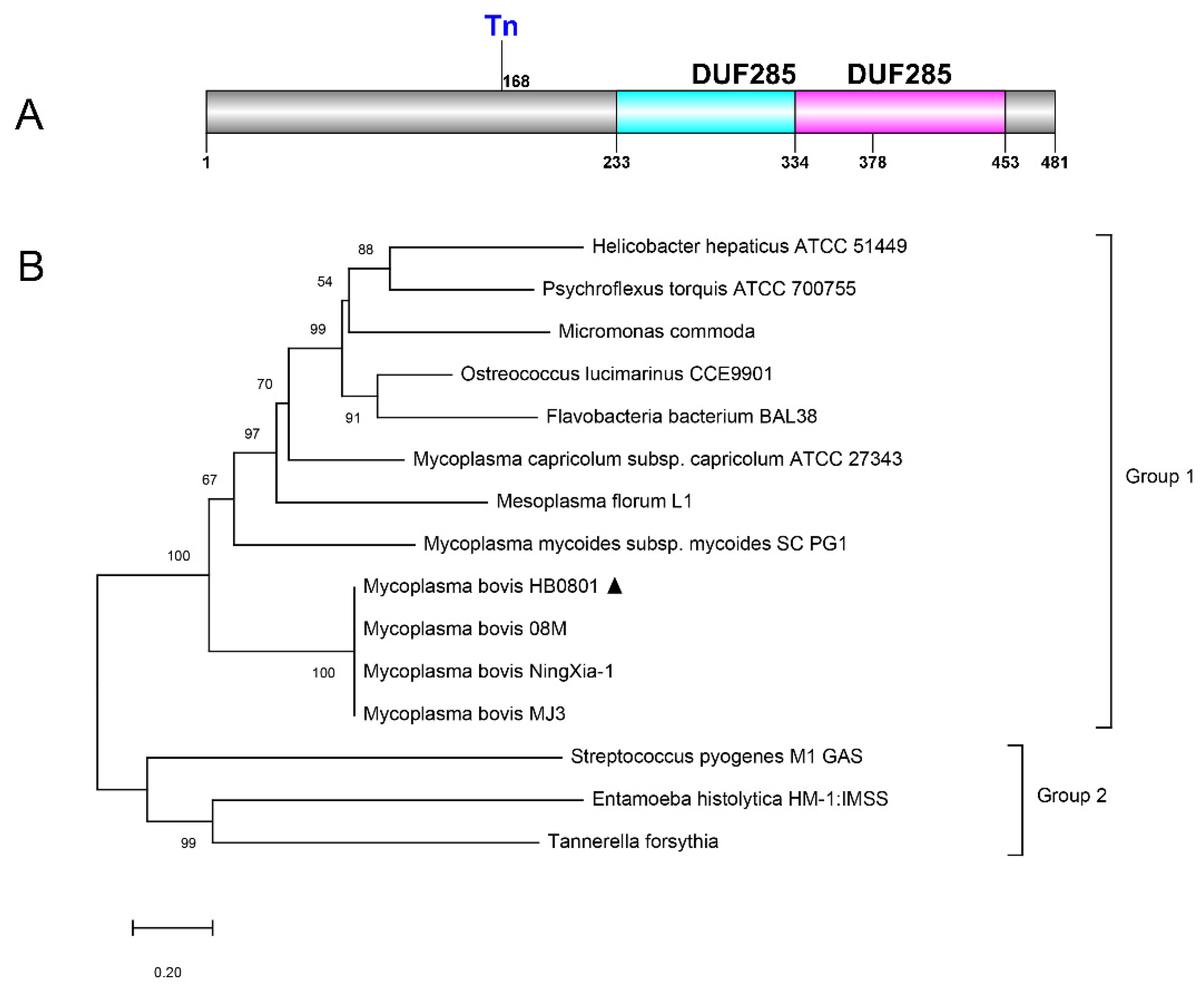
3.1. Bioinformatic Prediction of Genomic Features for the Mbov_0145 Gene
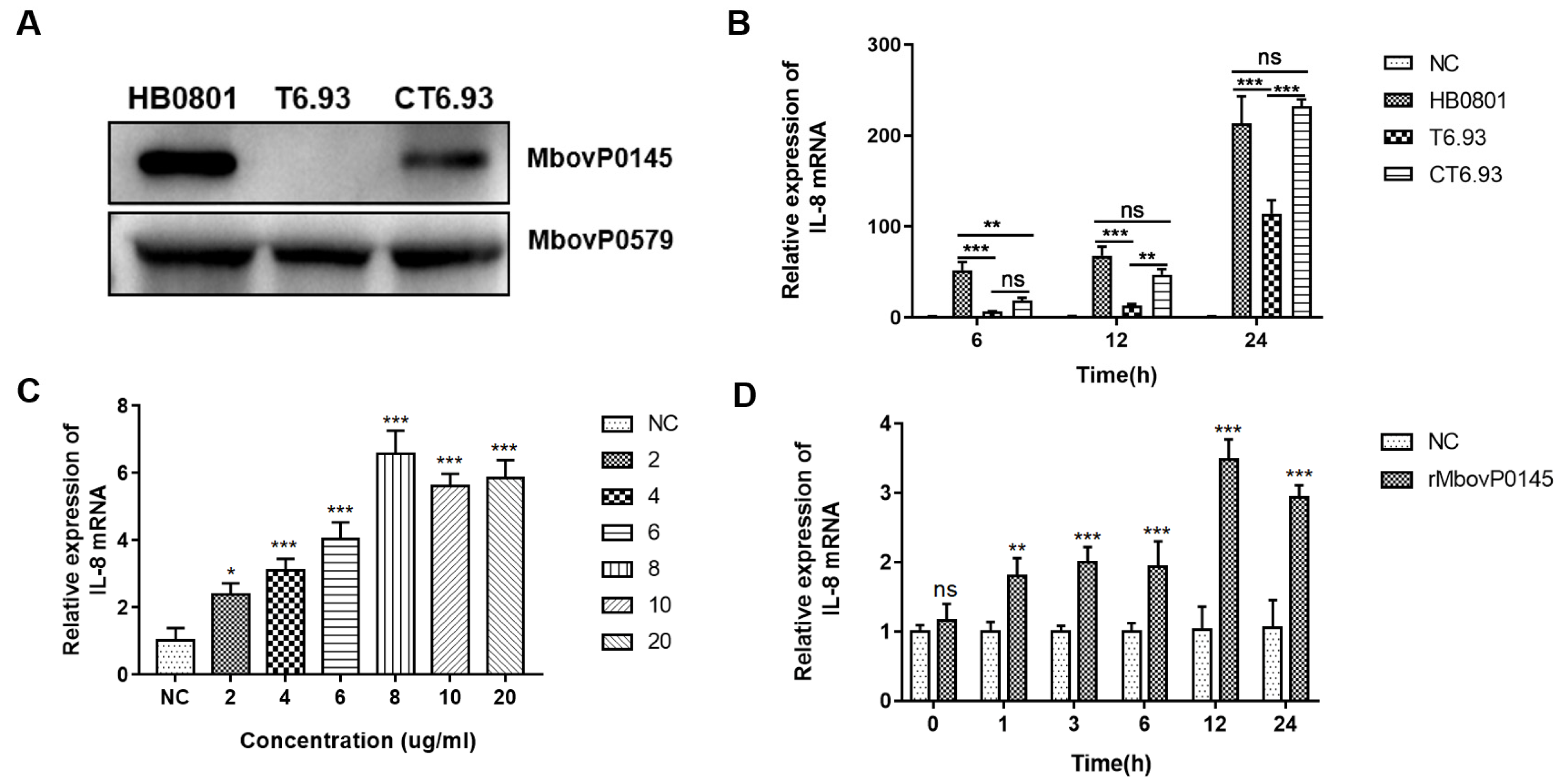
3.2. MbovP0145 Specifically Induced IL-8 Expression in EBL Cells
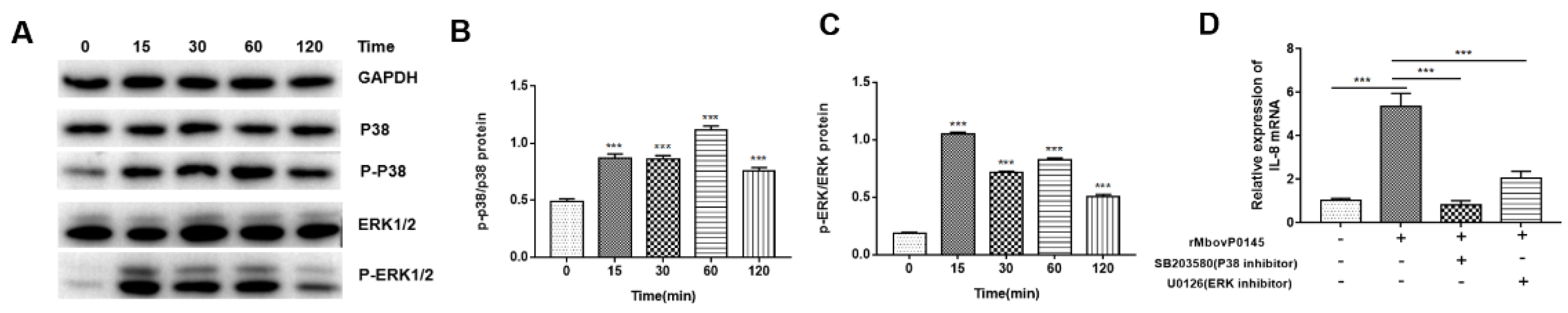
3.3. MbovP0145 Induction of IL-8 Expression Is Regulated by the MAPK Pathway
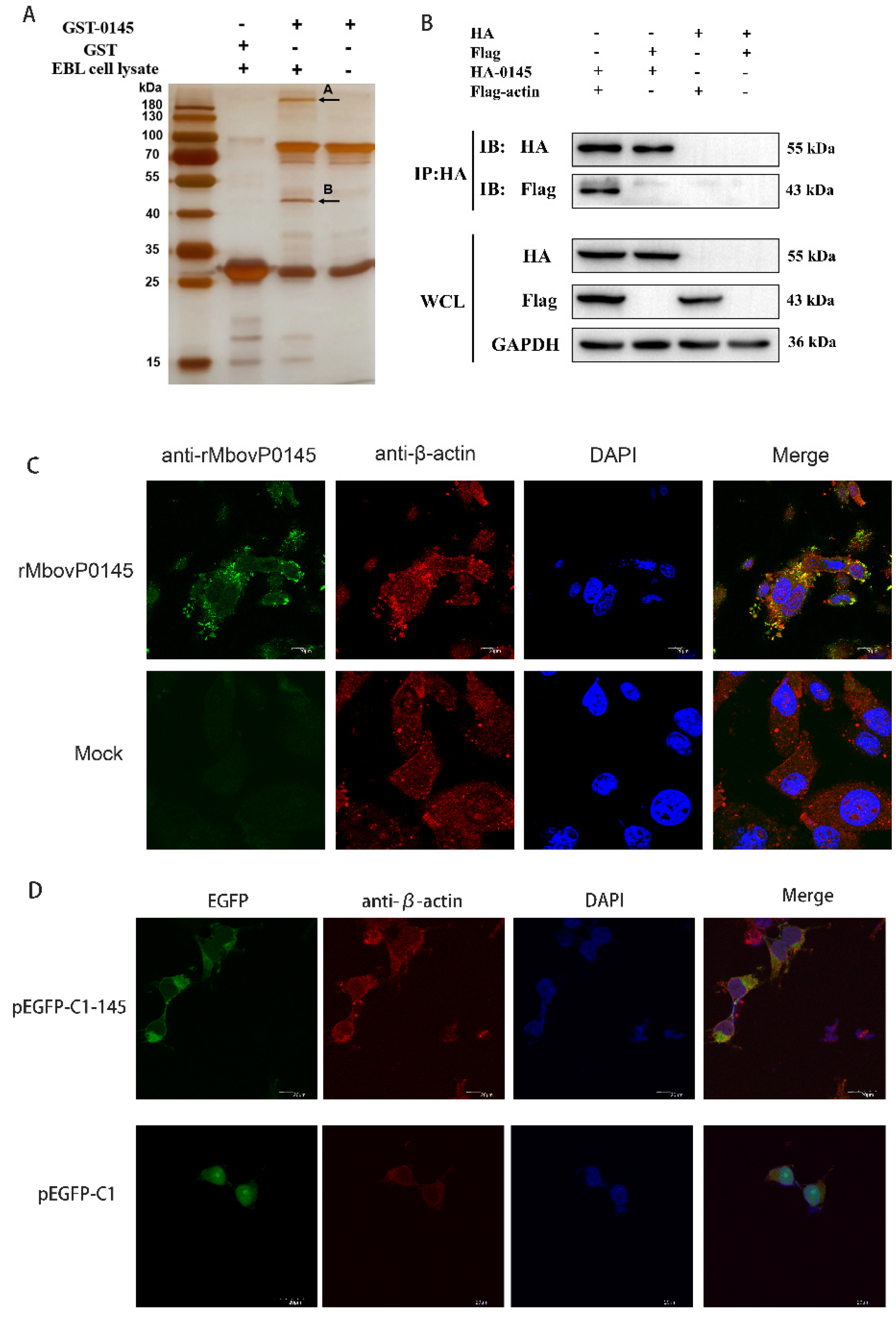
3.4. Preliminarily Identification of MbovP0145 Interactive Proteins
3.5. Confirmation of β-Actin as the Interactive Protein of MbovP0145
3.6. IL-8 Expression Induced by MbovP0145 Depends on Its Interactive β-Actin
3.7. MbovP0145 Induces Neutrophil Migration by Regulating the Production of IL-8
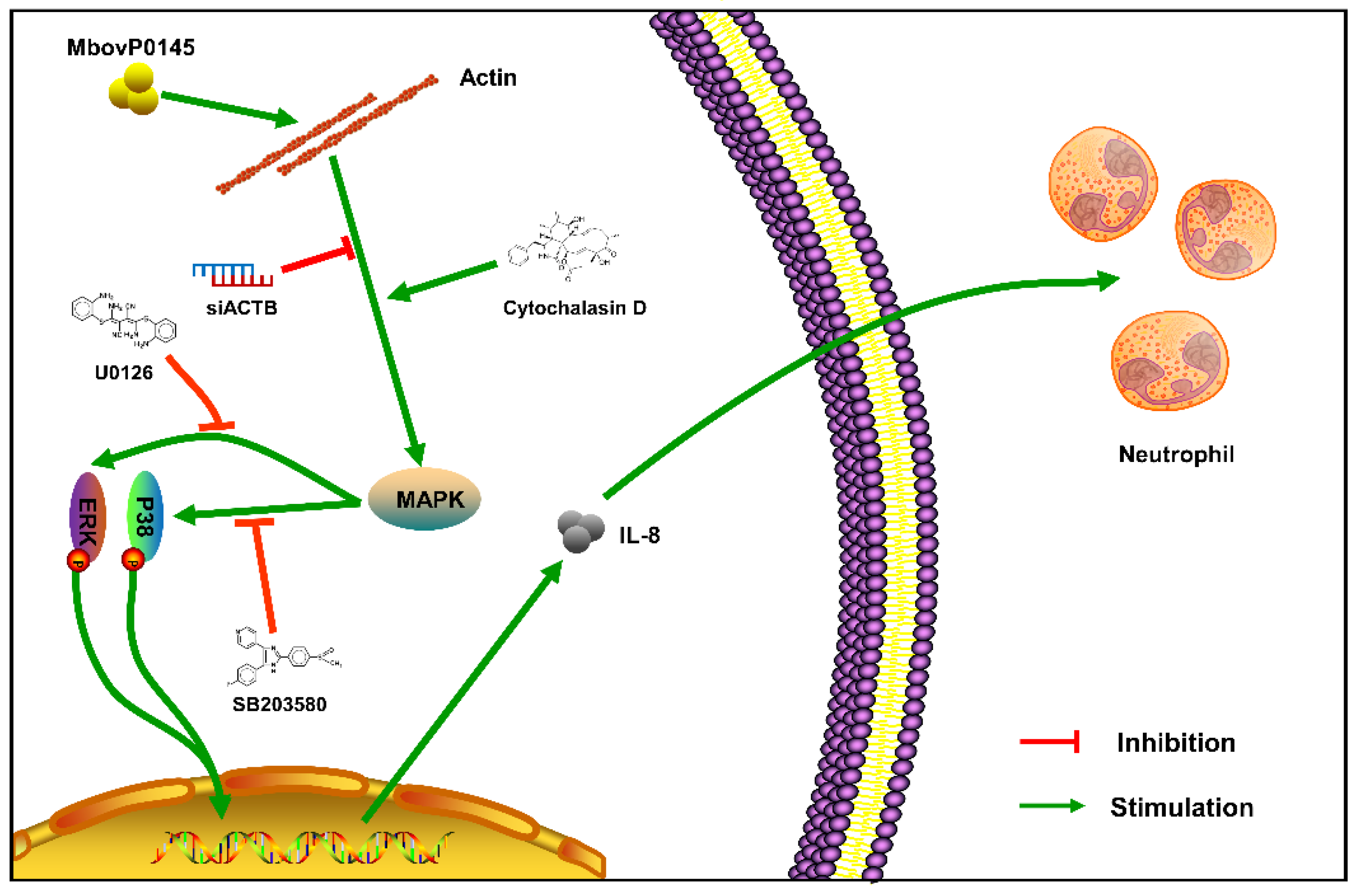
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Citti, C.; Blanchard, A. Mycoplasmas and their host: Emerging and re-emerging minimal pathogens. Trends Microbiol. 2013, 21, 196–203. [Google Scholar] [CrossRef] [PubMed]
- Pitcher, D.G.; Nicholas, R.A.J. Mycoplasma host specificity: Fact or fiction? Vet. J. 2005, 170, 300–306. [Google Scholar] [CrossRef] [PubMed]
- Burki, S.; Frey, J.; Pilo, P. Virulence, persistence and dissemination of Mycoplasma bovis. Vet. Microbiol. 2015, 179, 15–22. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hale, H.H.; Helmboldt, C.F.; Plastridge, W.N.; Stula, E.F. Bovine mastitis caused by a Mycoplasma species. Cornell Vet. 1962, 52, 582–591. [Google Scholar] [PubMed]
- Perez-Casal, J. Pathogenesis and Virulence of Mycoplasma bovis. Vet. Clin. N. Am. Food Anim. Pract. 2020, 36, 269–278. [Google Scholar] [CrossRef]
- Zhao, H.; Zhang, Y.; Wang, Z.; Liu, M.; Wang, P.; Wu, W.; Peng, C. MBOVPG45_0375 Encodes an IgG-Binding Protein and MBOVPG45_0376 Encodes an IgG-Cleaving Protein in Mycoplasma bovis. Front. Vet. Sci. 2021, 8, 644224. [Google Scholar] [CrossRef] [PubMed]
- Chen, S.; Hao, H.; Zhao, P.; Ji, W.; Li, M.; Liu, Y.; Chu, Y. Differential Immunoreactivity to Bovine Convalescent Serum Between Mycoplasma bovis Biofilms and Planktonic Cells Revealed by Comparative Immunoproteomic Analysis. Front. Microbiol. 2018, 9, 379. [Google Scholar] [CrossRef]
- Zhu, X.; Baranowski, E.; Dong, Y.; Li, X.; Hao, Z.; Zhao, G.; Zhang, H.; Lu, D.; Rasheed, M.; Chen, Y.; et al. An emerging role for cyclic dinucleotide phosphodiesterase and nanoRNase activities in Mycoplasma bovis: Securing survival in cell culture. PLoS Pathog 2020, 16, e1008661. [Google Scholar] [CrossRef]
- Tsirigotaki, A.; De Geyter, J.; Sostaric, N.; Economou, A.; Karamanou, S. Protein export through the bacterial Sec pathway. Nat. Rev. Microbiol. 2017, 15, 21–36. [Google Scholar] [CrossRef]
- Zhao, G.; Zhu, X.; Zhang, H.; Chen, Y.; Schieck, E.; Hu, C.; Chen, H.; Guo, A. Novel Secreted Protein of Mycoplasma bovis MbovP280 Induces Macrophage Apoptosis Through CRYAB. Front. Immunol. 2021, 12, 619362. [Google Scholar] [CrossRef]
- Zhang, H.; Zhao, G.; Guo, Y.; Menghwar, H.; Chen, Y.; Chen, H.; Guo, A. Mycoplasma bovis MBOV_RS02825 Encodes a Secretory Nuclease Associated with Cytotoxicity. Int. J. Mol. Sci. 2016, 17, 628. [Google Scholar] [CrossRef]
- Zhang, H.; Hu, G.; Lu, D.; Zhao, G.; Zhang, Y.; Zubair, M.; Chen, Y.; Hu, C.; Chen, X.; Chen, J.; et al. Comparative Secretome Analyses of Mycoplasma bovis Virulent and Attenuated Strains Revealed MbovP0145 as a Promising Diagnostic Biomarker. Front. Vet. Sci. 2021, 8, 583. [Google Scholar] [CrossRef]
- Silvestre, A.; Plaze, A.; Berthon, P.; Thibeaux, R.; Guillen, N.; Labruyere, E. In Entamoeba histolytica, a BspA family protein is required for chemotaxis toward tumour necrosis factor. Microb. Cell 2015, 2, 235–246. [Google Scholar] [CrossRef] [PubMed]
- Shi, L.; Gong, R.; Yin, Z.; Zhou, Y.; Pei, J.; Hu, Z.; Wang, L.; Hu, C.; Liu, T.; Chen, Y.; et al. Preliminary Diagnosis of Cattle infections Mycoplasma bovis Pneumonia. J. Huazhong Agric. Univ. 2008, 27, 572. [Google Scholar]
- Zhu, X.; Dong, Y.; Baranowski, E.; Li, X.; Zhao, G.; Hao, Z.; Zhang, H.; Chen, Y.; Hu, C.; Chen, H.; et al. Mbov_0503 Encodes a Novel Cytoadhesin that Facilitates Mycoplasma bovis Interaction with Tight Junctions. Microorganisms 2020, 8, 164. [Google Scholar] [CrossRef] [Green Version]
- Qi, J.; Guo, A.; Cui, P.; Chen, Y.; Mustafa, R.; Ba, X.; Hu, C.; Bai, Z.; Chen, X.; Shi, L.; et al. Comparative geno-plasticity analysis of Mycoplasma bovis HB0801 (Chinese isolate). PLoS ONE 2012, 7, e38239. [Google Scholar] [CrossRef]
- Fu, L.; Han, L.; Xie, C.; Li, W.; Lin, L.; Pan, S.; Zhou, Y.; Li, Z.; Jin, M.; Zhang, A. Identification of extracellular Actin as a ligand for triggering receptor expressed on myeloid Cells-1 signaling. Front. Immunol. 2017, 8, 917. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Khan, F.A.; Faisal, M.; Chao, J.; Liu, K.; Chen, X.; Zhao, G.; Menghwar, H.; Zhang, H.; Zhu, X.; Rasheed, M.A.; et al. Immunoproteomic identification of MbovP579, a promising diagnostic biomarker for serological detection of Mycoplasma bovis infection. Oncotarget 2016, 7, 39376–39395. [Google Scholar] [CrossRef] [Green Version]
- Hou, G.P.; Xue, B.Y.; Li, L.L.; Nan, Y.C.; Zhang, L.; Li, K.K.; Zhao, Q.; Hiscox, J.A.; Stewart, J.P.; Wu, C.Y.; et al. Direct Interaction Between CD163 N-Terminal Domain and MYH9 C-Terminal Domain Contributes to Porcine Reproductive and Respiratory Syndrome Virus Internalization by Permissive Cells. Front. Microbiol. 2019, 10, 1815. [Google Scholar] [CrossRef] [Green Version]
- Singh, R.K.; Furze, R.C.; Birrell, M.A.; Rankin, S.M.; Hume, A.N.; Seabra, M.C. A role for Rab27 in neutrophil chemotaxis and lung recruitment. BMC Cell. Biol. 2014, 15, 39. [Google Scholar] [CrossRef] [Green Version]
- Pilo, P.; Martig, S.; Frey, J.; Vilei, E.M. Antigenic and genetic characterisation of lipoprotein lppC from Mycoplasma mycoides subsp. mycoides SC. Vet. Res. 2003, 34, 761–775. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sharma, A.; Sojar, H.T.; Glurich, I.; Honma, K.; Kuramitsu, H.K.; Genco, R.J. Cloning, expression, and sequencing of a cell surface antigen containing a leucine-rich repeat motif from Bacteroides forsythus ATCC 43037. Infect. Immun. 1998, 66, 5703–5710. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ikegami, A.; Honma, K.; Sharma, A.; Kuramitsu, H.K. Multiple functions of the leucine-rich repeat protein LrrA of Treponema denticola. Infect. Immun. 2004, 72, 4619–4627. [Google Scholar] [CrossRef] [Green Version]
- Onishi, S.; Honma, K.; Liang, S.; Stathopouiou, P.; Kinane, D.; Hajishengallis, G.; Sharma, A. Toll-like receptor 2-mediated interleukin-8 expression in gingival epithelial cells by the Tannerella forsythia leucine-rich repeat protein BspA. Infect. Immun. 2008, 76, 198–205. [Google Scholar] [CrossRef] [Green Version]
- Sharma, A.; Inagaki, S.; Honma, K.; Sfintescu, C.; Baker, P.J.; Evans, R.T. Tannerella forsythia-induced alveolar bone loss in mice involves leucine-rich-repeat BspA protein. J. Dent. Res. 2005, 84, 462–467. [Google Scholar] [CrossRef]
- Zbinden, C.; Pilo, P.; Frey, J.; Bruckmaier, R.M.; Wellnitz, O. The immune response of bovine mammary epithelial cells to live or heat-inactivated Mycoplasma bovis. Vet. Microbiol. 2015, 179, 336–340. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nishi, K.; Gondaira, S.; Okamoto, M.; Nebu, T.; Koiwa, M.; Ohtsuka, H.; Murai, K.; Matsuda, K.; Fujiki, J.; Iwano, H.; et al. Effect of Mycoplasma bovis on expression of inflammatory cytokines and matrix metalloproteinases mRNA in bovine synovial cells. Vet. Immunol. Immunopathol. 2019, 216, 109920. [Google Scholar] [CrossRef]
- Gondaira, S.; Nishi, K.; Fujiki, J.; Iwano, H.; Watanabe, R.; Eguchi, A.; Hirano, Y.; Higuchi, H.; Nagahata, H. Innate immune response in bovine neutrophils stimulated with Mycoplasma bovis. Vet. Res. 2021, 52, 58. [Google Scholar] [CrossRef] [PubMed]
- Lee, K.E.; Kim, K.W.; Hong, J.Y.; Kim, K.E.; Sohn, M.H. Modulation of IL-8 Boosted by Mycoplasma pneumoniae lysate in Human Airway Epithelial Cells. J. Clin. Immunol. 2013, 33, 1117–1125. [Google Scholar] [CrossRef] [PubMed]
- Chmura, K.; Bai, X.Y.; Nakamura, M.; Kandasamy, P.; McGibney, M.; Kuronuma, K.; Mitsuzawa, H.; Voelker, D.R.; Chan, E.D. Induction of IL-8 by Mycoplasma pneumoniae membrane in BEAS-2B cells. Am. J. Physiol.-Lung C 2008, 295, L220–L230. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ojcius, D.M.; Hardy, R.D.; Coalson, J.J.; Peters, J.; Chaparro, A.; Techasaensiri, C.; Cantwell, A.M.; Kannan, T.R.; Baseman, J.B.; Dube, P.H. Analysis of Pulmonary Inflammation and Function in the Mouse and Baboon after Exposure to Mycoplasma pneumoniae CARDS Toxin. PLoS ONE 2009, 4, e7562. [Google Scholar] [CrossRef]
- Chen, Z.; Shao, X.; Dou, X.; Zhang, X.; Wang, Y.; Zhu, C.; Hao, C.; Fan, M.; Ji, W.; Yan, Y. Role of the Mycoplasma pneumoniae/Interleukin-8/Neutrophil Axis in the Pathogenesis of Pneumonia. PLoS ONE 2016, 11, e0146377. [Google Scholar] [CrossRef] [Green Version]
- Pease, J.E.; Sabroe, I. The role of interleukin-8 and its receptors in inflammatory lung disease: Implications for therapy. Am. J. Respir. Med. 2002, 1, 19–25. [Google Scholar] [CrossRef]
- Mukaida, N. Pathophysiological roles of interleukin-8/CXCL8 in pulmonary diseases. Am. J. Physiol.-Lung C 2003, 284, L566–L577. [Google Scholar] [CrossRef] [Green Version]
- Dugina, V.B.; Shagieva, G.S.; Shakhov, A.S.; Alieva, I.B. The Cytoplasmic Actins in the Regulation of Endothelial Cell Function. Int. J. Mol. Sci. 2021, 22, 7836. [Google Scholar] [CrossRef] [PubMed]
- Winder, S.J.; Ayscough, K.R. Actin-binding proteins. J. Cell Sci. 2005, 118, 651. [Google Scholar] [CrossRef] [Green Version]
- Pantaloni, D.; Le Clainche, C.; Carlier, M.F. Mechanism of actin-based motility. Science 2001, 292, 1502–1506. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Raymond, B.B.A.; Madhkoor, R.; Schleicher, I.; Uphoff, C.C.; Turnbull, L.; Whitchurch, C.B.; Rohde, M.; Padula, M.P.; Djordjevic, S.P. Extracellular Actin Is a Receptor for Mycoplasma hyopneumoniae. Front. Cell Infect. Microbiol. 2018, 8, 54. [Google Scholar] [CrossRef] [Green Version]
- Van den Berg, A.; Freitas, J.; Keles, F.; Snoek, M.; van Marle, J.; Jansen, H.M.; Lutter, R. Cytoskeletal architecture differentially controls post-transcriptional processing of IL-6 and IL-8 mRNA in airway epithelial-like cells. Exp. Cell Res. 2006, 312, 1496–1506. [Google Scholar] [CrossRef] [PubMed]
- Kustermans, G.; El Mjiyad, N.; Horion, J.; Jacobs, N.; Piette, J.; Legrand-Poels, S. Actin cytoskeleton differentially modulates NF-kappaB-mediated IL-8 expression in myelomonocytic cells. Biochem. Pharmacol. 2008, 76, 1214–1228. [Google Scholar] [CrossRef] [PubMed]
- Gao, M.; Wu, S.; Ji, J.; Zhang, J.; Liu, Q.; Yue, Y.; Liu, L.; Liu, X.; Liu, W. The influence of actin depolymerization induced by Cytochalasin D and mechanical stretch on interleukin-8 expression and JNK phosphorylation levels in human retinal pigment epithelial cells. BMC Ophthalmol. 2017, 17, 43. [Google Scholar] [CrossRef] [Green Version]
- Nemeth, Z.H.; Deitch, E.A.; Davidson, M.T.; Szabo, C.; Vizi, E.S.; Hasko, G. Disruption of the actin cytoskeleton results in nuclear factor-kappa B activation and inflammatory mediator production in cultured human intestinal epithelial cells (vol 200, pg 71, 2004). J. Cell. Physiol. 2004, 200, 167–168. [Google Scholar] [CrossRef]
- Xythalis, D.; Frewin, M.B.; Gudewicz, P.W. Inhibition of IL-8-mediated MAPK activation in human neutrophils by beta(1) integrin ligands. Inflammation 2002, 26, 83–88. [Google Scholar] [CrossRef]
- Vicente-Manzanares, M.; Choi, C.K.; Horwitz, A.R. Integrins in cell migration--the actin connection. J. Cell Sci. 2009, 122, 199–206. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, Y.; Liu, S.; Zhang, Y.; Yang, J. Myosin Heavy Chain 9: Oncogene or Tumor Suppressor Gene? Med. Sci. Monit. 2019, 25, 888–892. [Google Scholar] [CrossRef] [PubMed]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Names | Primer Sequences (5′→3′) 1 |
|---|---|
| IL-8-F | GAAGAGAGCTGAGAAGCAAGATCC |
| IL-8-R | ACCCACACAGAACATGAGGC |
| GAPDH-F | TGGTGAAGGTCGGAGTGAAC |
| GAPDH-R | ATGGCGACGATGTCCACTTT |
| pOH/P-0145-F1 | ATTTGCGGCCGCACGGGGCTAAAGAAGCTGATAT (NotI) |
| pOH/P-0145-R1 | TAGCAAAAAGCATAATTATTTATATCCTTTTCTT |
| pOH/P-0145-F2 | TAAATAATTATGCTTTTTGCTAGTTCACTTCCTTT |
| pOH/P-0145-R2 | AATTGCGGCCGCTTATTTAGATACTTGCCTAAAA (NotI) |
| GST-0145-F | TTCCAGGGGCCCCTGGGATCCATGCTGTTTGCCTCAAGCCTG (BamHI) |
| GST-0145-R | GTCACGATGCGGCCGCTCGAGTTATTTGCTAACCTGACGAAAATTCG (XhoI) |
| HA-0145-F | GTTCCAGATTACGCTGAATTCATGCTGTTTGCCTCAAGCCTG (EcoRI) |
| HA-0145-R | ATTAAGATCTGCTAGCTCGAGTTATTTGCTAACCTGACGAAAATTCG (XhoI) |
| Flag-actin-F | AAGCTTGCGGCCGCGAATTCAATGGATGATGATATTGCTGCGC (EcoRI) |
| Flag-actin-R | CAGGGATGCCACCCGGGATCCCTAGAAGCATTTGCGGTGGAC (BamHI) |
| EGFP-0145 F | AGTCCGGACTCAGATCTCGAGATATGCTGTTTGCCTCAAGCCTG (XhoI) |
| EGFP-0145 R | TTATCTAGATCCGGTGGATCCTTATTTGCTAACCTGACGAAAATTCG (BamHI) |
| Band | Protein Name | Accession No.a | Unique Peptide Count b | Percentage Coverage c | MW (KDa) d | PI e |
|---|---|---|---|---|---|---|
| A | Myosin heavy chain 9 | F1MQ37 | 92 | 35.62% | 227,201.32 | 5.49 |
| B | Actin, cytoplasmic 1 | P60712 | 10 | 24.00% | 41,736.29 | 5.29 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lu, D.; Zhang, H.; Zhang, Y.; Zhao, G.; Anwar Khan, F.; Chen, Y.; Hu, C.; Yang, L.; Chen, H.; Guo, A. Secreted MbovP0145 Promotes IL-8 Expression through Its Interactive β-Actin and MAPK Activation and Contributes to Neutrophil Migration. Pathogens 2021, 10, 1628. https://0-doi-org.brum.beds.ac.uk/10.3390/pathogens10121628
Lu D, Zhang H, Zhang Y, Zhao G, Anwar Khan F, Chen Y, Hu C, Yang L, Chen H, Guo A. Secreted MbovP0145 Promotes IL-8 Expression through Its Interactive β-Actin and MAPK Activation and Contributes to Neutrophil Migration. Pathogens. 2021; 10(12):1628. https://0-doi-org.brum.beds.ac.uk/10.3390/pathogens10121628
Chicago/Turabian StyleLu, Doukun, Hui Zhang, Yiqiu Zhang, Gang Zhao, Farhan Anwar Khan, Yingyu Chen, Changmin Hu, Liguo Yang, Huanchun Chen, and Aizhen Guo. 2021. "Secreted MbovP0145 Promotes IL-8 Expression through Its Interactive β-Actin and MAPK Activation and Contributes to Neutrophil Migration" Pathogens 10, no. 12: 1628. https://0-doi-org.brum.beds.ac.uk/10.3390/pathogens10121628