
Genotyping and Antimicrobial Susceptibility of Clostridium perfringens and Clostridioides difficile in Camel Minced Meat

 ,
, 
 ,
,  , and
, and 
Abstract
:1. Introduction
2. Results
2.1. Isolation and Identification of C. perfringens and C. difficile
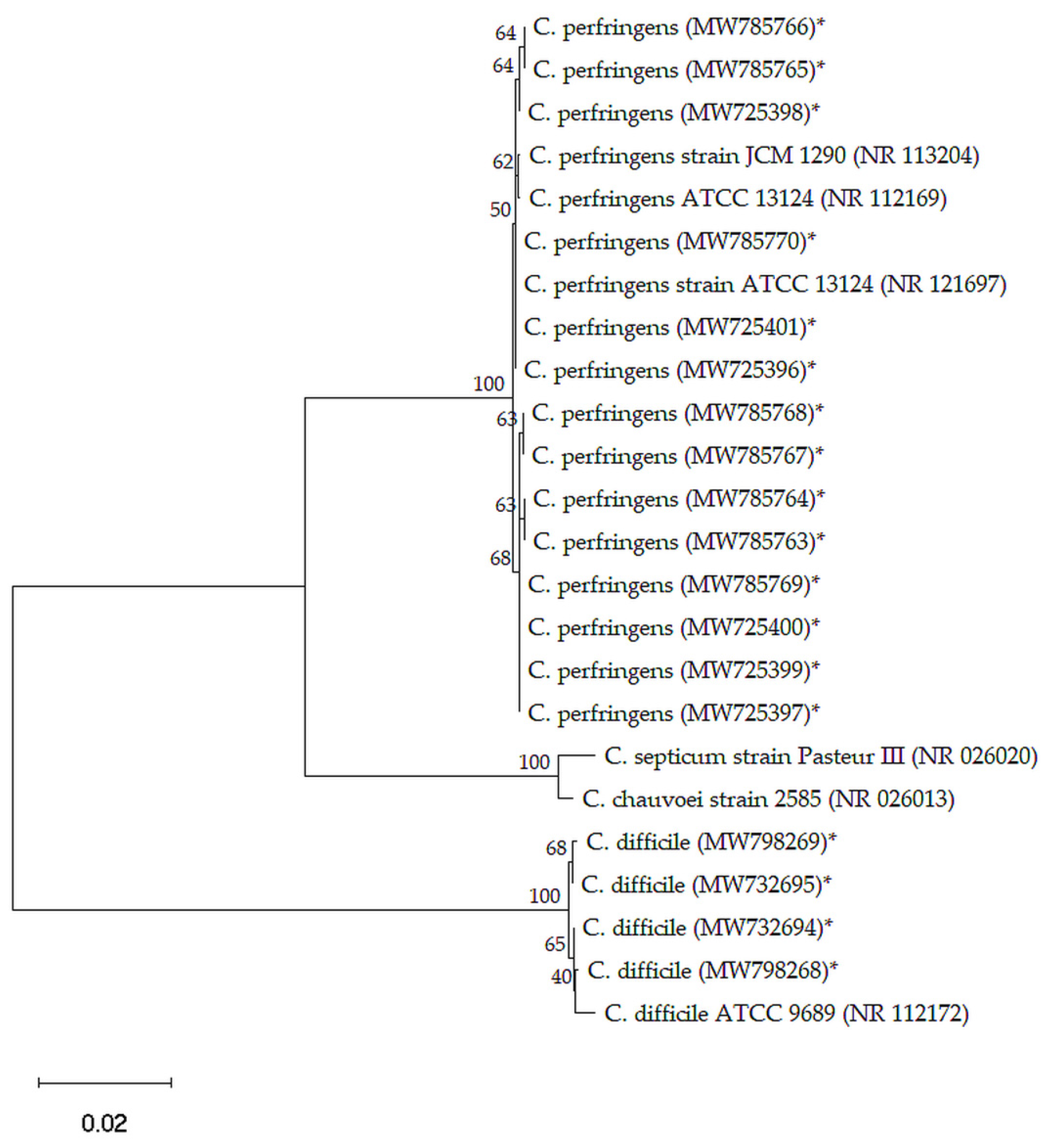
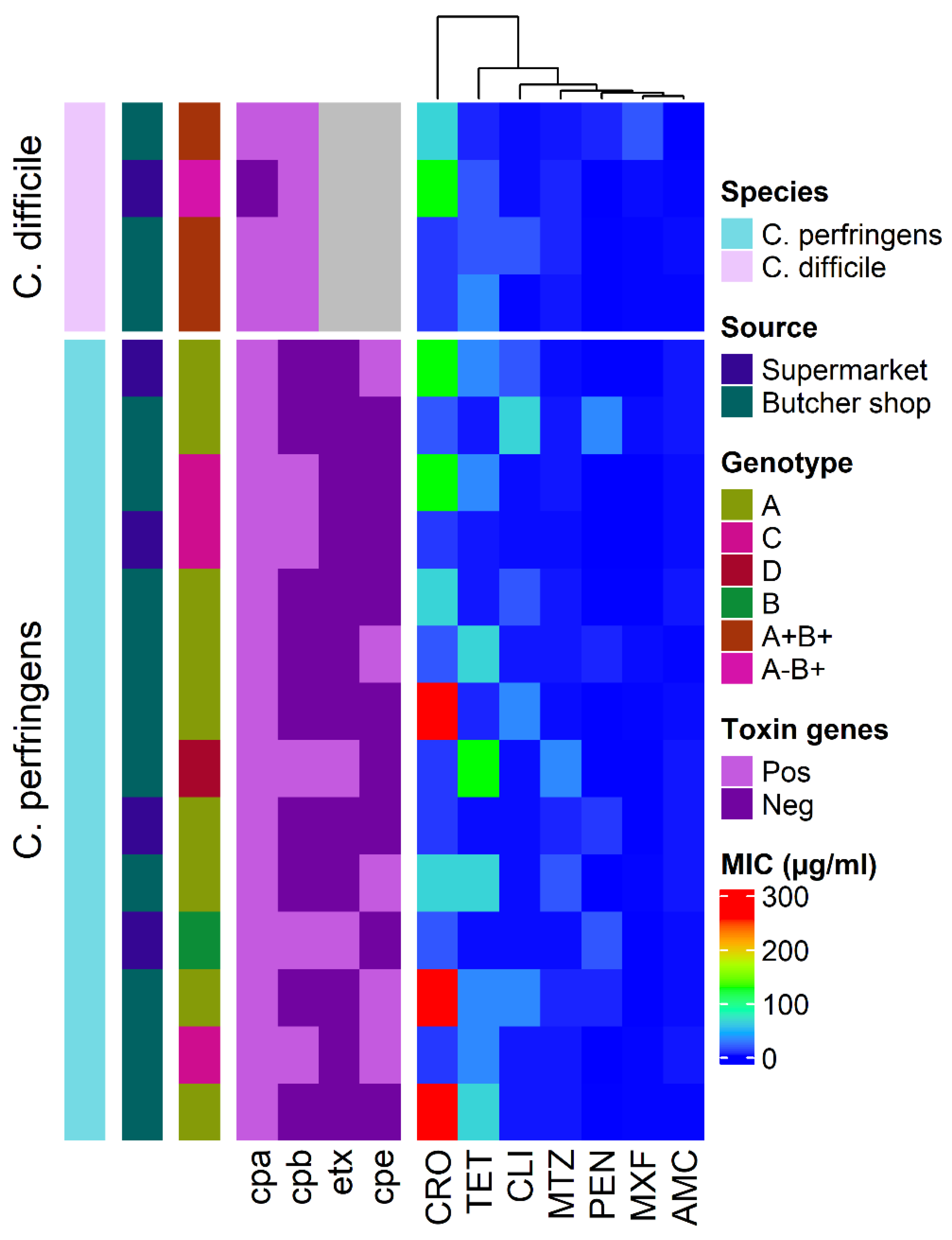
2.2. Genotyping of C. perfringens and C. difficile Isolates
2.3. Detection of C. perfringens and C. difficile Toxin by ELISA
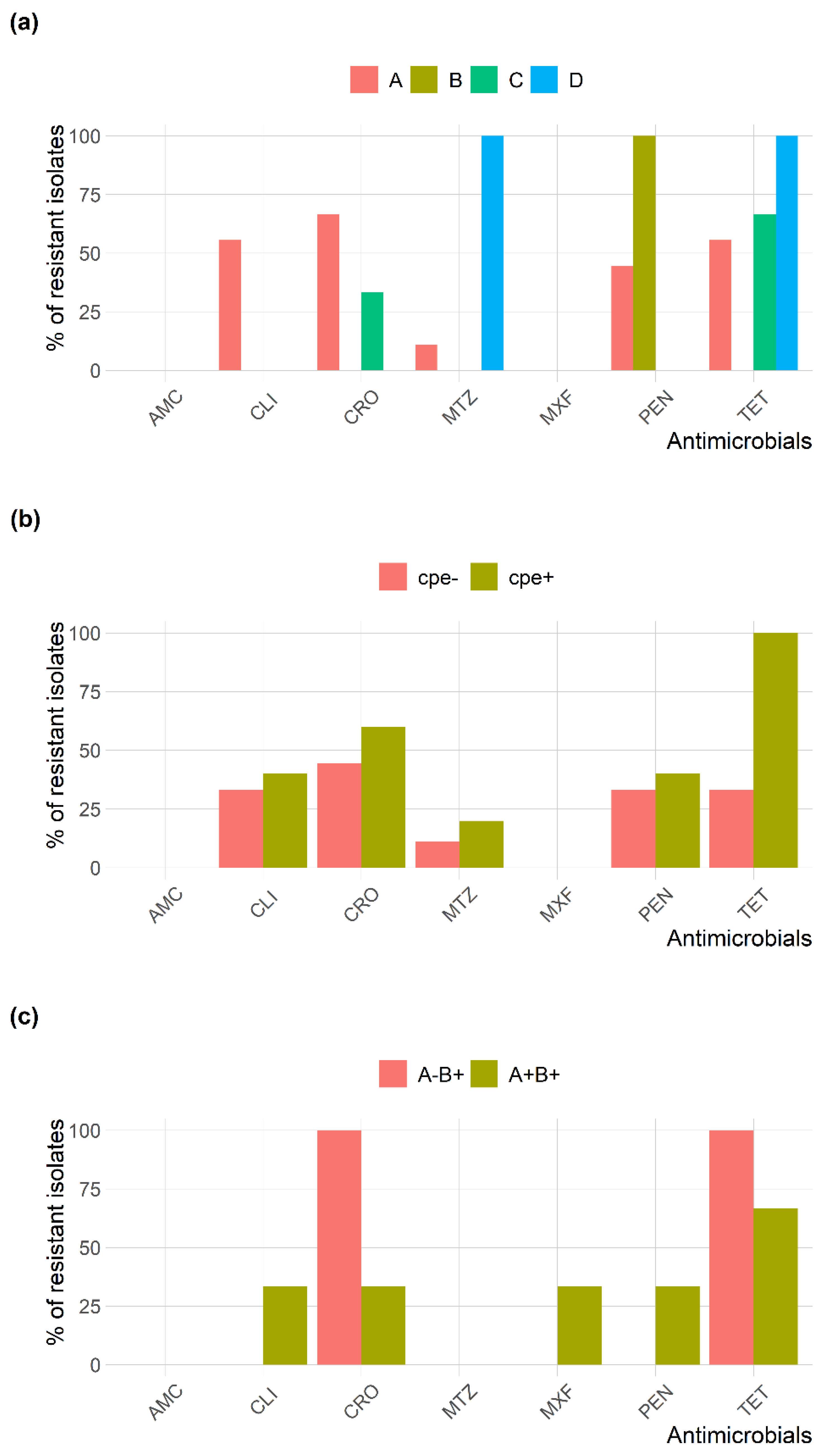
2.4. Antimicrobial Susceptibility
3. Discussion
4. Materials and Methods
4.1. Sample Collection
4.2. Isolation and Counting of C. perfringens
4.3. Isolation of C. difficile
4.4. Molecular Identification and Genotyping
4.4.1. 16S rRNA Gene Amplification and Sequencing
4.4.2. Detection of C. perfringens and C. difficile Toxin Genes by Real-Time PCR
4.5. Detection of C. perfringens and C. difficile Toxin by ELISA
4.6. Antimicrobial Susceptibility Testing
4.7. Data Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- World Health Organization. The Role of food Safety in Health and Development: Report of a Joint FAO/WHO Expert Committee on Food Safety; World Health Organization: Geneva, Switzerland, 1984; p. 79. [Google Scholar]
- Mead, P.S.; Slutsker, L.; Dietz, V.; McCaig, L.F.; Bresee, J.S.; Shapiro, C.; Griffin, P.M.; Tauxe, R.V. Food-related illness and death in the United States. Emerg. Infect. Dis. 1999, 5, 607. [Google Scholar] [CrossRef]
- Flint-Garcia, S.A.; Thuillet, A.C.; Yu, J.; Pressoir, G.; Romero, S.M.; Mitchell, S.E.; Doebley, J.; Kresovich, S.; Goodman, M.M.; Buckler, E.S. Maize association population: A high-resolution platform for quantitative trait locus dissection. Plant J. 2005, 44, 1054–1064. [Google Scholar] [CrossRef]
- McIntyre, L.; Vallaster, L.; Wilcott, L.; Henderson, S.B.; Kosatsky, T. Evaluation of food safety knowledge, attitudes and self-reported hand washing practices in FOODSAFE trained and untrained food handlers in British Columbia, Canada. Food Control 2013, 30, 150–156. [Google Scholar] [CrossRef]
- Havelaar, A.H.; Kirk, M.D.; Torgerson, P.R.; Gibb, H.J.; Hald, T.; Lake, R.J.; Praet, N.; Bellinger, D.C.; De Silva, N.R.; Gargouri, N. World Health Organization global estimates and regional comparisons of the burden of foodborne disease in 2010. PLoS Med. 2015, 12, e1001923. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hoffmann, S.; Devleesschauwer, B.; Aspinall, W.; Cooke, R.; Corrigan, T.; Havelaar, A.; Angulo, F.; Gibb, H.; Kirk, M.; Lake, R. Attribution of global foodborne disease to specific foods: Findings from a World Health Organization structured expert elicitation. PLoS ONE 2017, 12, e0183641. [Google Scholar] [CrossRef] [Green Version]
- CDC. CDC, What Is a Foodborne Disease Outbreak and Why Do They Occur. 2012. Available online: http://www.cdc.gov/foodsafety/facts.html#whatisanoutbreak. (accessed on 20 August 2021).
- McClane, B.A.; Uzal, F.A.; Miyakawa, M.F.; Lyerly, D.; Wilkins, T. The enterotoxic clostridia. Prokaryotes 2006, 4, 698–752. [Google Scholar]
- Miyamoto, K.; Nagahama, M. Clostridium: Food poisoning by Clostridium perfringens. In Encyclopedia of Food and Health; Caballero, B., Finglas, P.M., Toldrá, F., Eds.; Academic Press: Oxford, UK, 2016; pp. 149–154. [Google Scholar]
- Rhodehamel, E.; Harmon, S. BAM: Clostridium perfringens; US FDA: Rockville, MD, USA, 2001. [Google Scholar]
- Rodriguez-Palacios, A.; Staempfli, H.R.; Duffield, T.; Weese, J.S. Clostridium difficile in retail ground meat, Canada. Emerg. Infect. Dis. 2007, 13, 485. [Google Scholar] [CrossRef]
- Songer, J.G.; Trinh, H.T.; Killgore, G.E.; Thompson, A.D.; McDonald, L.C.; Limbago, B.M. Clostridium difficile in retail meat products, USA, 2007. Emerg. Infect. Dis. 2009, 15, 819. [Google Scholar] [CrossRef] [PubMed]
- Odeyemi, O.; Sani, N. Antibiotic resistance and burden of foodborne diseases in developing countries. Future Sci. OA 2016, 2, FSO139. [Google Scholar] [CrossRef] [Green Version]
- Chang, Q.; Wang, W.; Regev-Yochay, G.; Lipsitch, M.; Hanage, W.P. Antibiotics in agriculture and the risk to human health: How worried should we be? Evol. Appl. 2015, 8, 240–247. [Google Scholar] [CrossRef] [Green Version]
- Liu, Y.-Y.; Wang, Y.; Walsh, T.R.; Yi, L.-X.; Zhang, R.; Spencer, J.; Doi, Y.; Tian, G.; Dong, B.; Huang, X. Emergence of plasmid-mediated colistin resistance mechanism MCR-1 in animals and human beings in China: A microbiological and molecular biological study. Lancet Infect. Dis. 2016, 16, 161–168. [Google Scholar] [CrossRef]
- Dawood, A.A.; Alkanhal, M.A. Nutrient composition of Najdi-camel meat. Meat Sci. 1995, 39, 71–78. [Google Scholar] [CrossRef]
- Al-Ani, F. Camel Management and Disease, 1st ed.; Al. Sharq Printing Press: Ar-Rayyan, Qatar; Dar-Ammar Book Publisher: Amman, Jordan, 2004. [Google Scholar]
- Bosilevac, J.M.; Gassem, M.A.; Al Sheddy, I.A.; Almaiman, S.A.; Al-Mohizea, I.S.; Alowaimer, A.; Koohmaraie, M. Prevalence of Escherichia coli O157: H7 and Salmonella in camels, cattle, goats, and sheep harvested for meat in Riyadh. J. Food Prot. 2015, 78, 89–96. [Google Scholar] [CrossRef] [Green Version]
- Raji, M.A.; Garaween, G.; Ehricht, R.; Monecke, S.; Shibl, A.M.; Senok, A. Genetic characterization of Staphylococcus aureus isolated from retail meat in Riyadh, Saudi Arabia. Front. Microbiol. 2016, 7, 911. [Google Scholar] [CrossRef] [PubMed]
- El-Ghareeb, W.R.; Almathen, F.S.; Fayez, M.M.; Alsultan, R.A. Methicillin resistant Staphylococcus aureus (MRSA) in camel meat: Prevalence and antibiotic susceptibility. Slov. Vet. Res. 2019, 56, 249–256. [Google Scholar] [CrossRef] [Green Version]
- El-Ghareeb, W.R.; Abdel-Raheem, S.M.; Al-Marri, T.M.; Alaql, F.A.; Fayez, M.M. Isolation and identification of extended spectrum β-lactamases (ESBLs) Escherichia coli from minced camel meat in Eastern province, Saudi Arabia. Thai J. Vet. Med. 2020, 50, 155–161. [Google Scholar]
- McClane, B.A.; Robertson, S.L.; Li, J. Clostridium perfringens. In Food Microbiology: Fundamentals and Frontiers; Wiley & Sons: New York, NY, USA, 2012; pp. 465–489. [Google Scholar]
- Nowell, V.J.; Poppe, C.; Parreira, V.R.; Jiang, Y.-F.; Reid-Smith, R.; Prescott, J.F. Clostridium perfringens in retail chicken. Anaerobe 2010, 16, 314–315. [Google Scholar] [CrossRef]
- Miki, Y.; Miyamoto, K.; Kaneko-Hirano, I.; Fujiuchi, K.; Akimoto, S. Prevalence and characterization of enterotoxin gene-carrying Clostridium perfringens isolates from retail meat products in Japan. Appl. Environ. Microbiol. 2008, 74, 5366–5372. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wen, Q.; McClane, B.A. Detection of enterotoxigenic Clostridium perfringens type A isolates in American retail foods. Appl. Environ. Microbiol. 2004, 70, 2685–2691. [Google Scholar] [CrossRef] [Green Version]
- Basma, S.; AM, E.M. Detection of enterotoxigenic Clostridium perfringens type: A in camel meat. Vet. Med. J. 2006, 54, 179–188. [Google Scholar]
- Mohamed, M.E.; I Suelam, I.; Saleh, M.A. The presence of toxin genes of Clostridium perfringens isolated from camels and humans in Egypt. Vet. Arh. 2010, 80, 383–392. [Google Scholar]
- Kamber, U.; Gokce, H.; Elmali, M. Clostridium perfringens and its toxins in minced meat from Kars, Turkey. Food Addit. Contam. 2007, 24, 673–678. [Google Scholar] [CrossRef] [PubMed]
- Jiang, Y.; Ma, Y.; Liu, Q.; Li, T.; Li, Y.; Guo, K.; Zhang, Y. Tracing Clostridium perfringens strains from beef processing of slaughter house by pulsed-field gel electrophoresis, and the distribution and toxinotype of isolates in Shaanxi province, China. Food Microbiol. 2022, 101, 103887. [Google Scholar] [CrossRef] [PubMed]
- Khan, M.A.; Khan, S.B.; Khan, I.U.; Sultan, A.; Khan, R.; Shahid, M.; Aqib, A.I.; Khan, S.; Khan, A.Z.; Hassan, M. The prevalence of Clostridium perfringens in retail meat of Mardan, Pakistan. Turk. J. Vet. Anim. Sci. 2020, 44, 618–623. [Google Scholar] [CrossRef]
- Madoroba, E.; Magwedere, K.; Chaora, N.S.; Matle, I.; Muchadeyi, F.; Mathole, M.A.; Pierneef, R. Microbial Communities of Meat and Meat Products: An Exploratory Analysis of the Product Quality and Safety at Selected Enterprises in South Africa. Microorganisms 2021, 9, 507. [Google Scholar] [CrossRef]
- Jang, Y.-S.; Kim, D.-H.; Bae, D.; Kim, S.-H.; Kim, H.; Moon, J.-S.; Song, K.-Y.; Chon, J.-W.; Seo, K.-H. Prevalence, toxin-typing, and antimicrobial susceptibility of Clostridium perfringens from retail meats in Seoul, Korea. Anaerobe 2020, 64, 102235. [Google Scholar] [CrossRef]
- Guran, H.S.; Vural, A.; Erkan, M.E. The prevalence and molecular typing of Clostridium perfringens in ground beef and sheep meats. J. Verbraucherschutz Lebensmittelsicherheit 2014, 9, 121–128. [Google Scholar] [CrossRef]
- Uzal, F.A.; Songer, J.G. Diagnosis of Clostridium perfringens intestinal infections in sheep and goats. J. Vet. Diagn. Investig. 2008, 20, 253–265. [Google Scholar] [CrossRef] [Green Version]
- Fayez, M.; Elsohaby, I.; Al-Marri, T.; Zidan, K.; Aldoweriej, A.; El-Sergany, E.; Elmoslemany, A. Genotyping and antimicrobial susceptibility of Clostridium perfringens isolated from dromedary camels, pastures and herders. Comp. Immunol. Microbiol. Infect. Dis. 2020, 70, 101460. [Google Scholar] [CrossRef]
- Collie, R.E.; McClane, B.A. Evidence that the enterotoxin gene can be episomal in Clostridium perfringens isolates associated with non-food-borne human gastrointestinal diseases. J. Clin. Microbiol. 1998, 36, 30–36. [Google Scholar] [CrossRef] [Green Version]
- Sparks, S.G.; Carman, R.J.; Sarker, M.R.; McClane, B.A. Genotyping of enterotoxigenic Clostridium perfringens fecal isolates associated with antibiotic-associated diarrhea and food poisoning in North America. J. Clin. Microbiol. 2001, 39, 883–888. [Google Scholar] [CrossRef] [Green Version]
- Brynestad, S.; Granum, P.E. Clostridium perfringens and foodborne infections. Int. J. Food Microbiol. 2002, 74, 195–202. [Google Scholar] [CrossRef]
- Llanco, L.; Nakano, V.; Ferreira, A.; Avila-Campos, M. Toxinotyping and antimicrobial susceptibility of Clostridium perfringens isolated from broiler chickens with necrotic enteritis. Int. J. Food Microbiol. Res. 2012, 4, 290. [Google Scholar]
- Petit, L.; Gibert, M.; Popoff, M.R. Clostridium perfringens: Toxinotype and genotype. Trends Microbiol. 1999, 7, 104–110. [Google Scholar] [CrossRef]
- Freedman, J.C.; Shrestha, A.; McClane, B.A. Clostridium perfringens enterotoxin: Action, genetics, and translational applications. Toxins 2016, 8, 73. [Google Scholar] [CrossRef] [Green Version]
- Yibar, A.; Cetin, E.; Ata, Z.; Erkose, E.; Tayar, M. Clostridium perfringens contamination in retail meat and meat-based products in Bursa, Turkey. Foodborne Pathog. Dis. 2018, 15, 239–245. [Google Scholar] [CrossRef]
- Lessa, F.C.; Mu, Y.; Bamberg, W.M.; Beldavs, Z.G.; Dumyati, G.K.; Dunn, J.R.; Farley, M.M.; Holzbauer, S.M.; Meek, J.I.; Phipps, E.C. Burden of Clostridium difficile infection in the United States. N. Engl. J. Med. 2015, 372, 825–834. [Google Scholar] [CrossRef] [Green Version]
- Kola, A.; Wiuff, C.; Akerlund, T.; van Benthem, B.H.; Coignard, B.; Lyytikäinen, O.; Weitzel-Kage, D.; Suetens, C.; Wilcox, M.H.; Kuijper, E.J. Survey of Clostridium difficile infection surveillance systems in Europe, 2011. Eurosurveillance 2016, 21, 30291. [Google Scholar] [CrossRef] [Green Version]
- Bouttier, S.; Barc, M.-C.; Felix, B.; Lambert, S.; Collignon, A.; Barbut, F. Clostridium difficile in ground meat, France. Emerg. Infect. Dis. 2010, 16, 733. [Google Scholar] [CrossRef]
- Von Abercron, S.M.M.; Karlsson, F.; Wigh, G.T.; Wierup, M.; Krovacek, K. Low occurrence of Clostridium difficile in retail ground meat in Sweden. J. Food Protect. 2009, 72, 1732–1734. [Google Scholar] [CrossRef]
- De Boer, E.; Zwartkruis-Nahuis, A.; Heuvelink, A.E.; Harmanus, C.; Kuijper, E.J. Prevalence of Clostridium difficile in retailed meat in the Netherlands. Int. J. Food Microbiol. 2011, 144, 561–564. [Google Scholar] [CrossRef] [PubMed]
- Hofer, E.; Haechler, H.; Frei, R.; Stephan, R. Low occurrence of Clostridium difficile in fecal samples of healthy calves and pigs at slaughter and in minced meat in Switzerland. J. Food Protect. 2010, 73, 973–975. [Google Scholar] [CrossRef] [PubMed]
- Jöbstl, M.; Heuberger, S.; Indra, A.; Nepf, R.; Köfer, J.; Wagner, M. Clostridium difficile in raw products of animal origin. Int. J. Food Microbiol. 2010, 138, 172–175. [Google Scholar] [CrossRef]
- Dubberke, E.R.; Haslam, D.B.; Lanzas, C.; Bobo, L.D.; Burnham, C.A.; Gröhn, Y.T.; Tarr, P.I. The ecology and pathobiology of Clostridium difficile infections: An interdisciplinary challenge. Zoonoses Public health 2011, 58, 4–20. [Google Scholar] [CrossRef] [Green Version]
- Rodriguez-Palacios, A.; Pickworth, C.; Loerch, S.; LeJeune, J.T. Transient fecal shedding and limited animal-to-animal transmission of Clostridium difficile by naturally infected finishing feedlot cattle. Appl. Environ. Microbiol. 2011, 77, 3391–3397. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Weese, J.S.; Rousseau, J.; Deckert, A.; Gow, S.; Reid-Smith, R.J. Clostridium difficile and methicillin-resistant Staphylococcus aureus shedding by slaughter-age pigs. BMC Vet. Res. 2011, 7, 1–7. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- EFSA, E. European Food Safety Authority and European Centre for Disease Prevention and Control. EFSA J. 2015, 13, 4329. [Google Scholar]
- Janezic, S.; Zidaric, V.; Pardon, B.; Indra, A.; Kokotovic, B.; Blanco, J.L.; Seyboldt, C.; Diaz, C.R.; Poxton, I.R.; Perreten, V. International Clostridium difficile animal strain collection and large diversity of animal associated strains. BMC Microbiol. 2014, 14, 1–10. [Google Scholar] [CrossRef] [Green Version]
- Knight, D.R.; Elliott, B.; Chang, B.J.; Perkins, T.T.; Riley, T.V. Diversity and evolution in the genome of Clostridium difficile. Clin. Microbiol. Rev. 2015, 28, 721–741. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Immerseel, F.V.; Buck, J.D.; Pasmans, F.; Huyghebaert, G.; Haesebrouck, F.; Ducatelle, R. Clostridium perfringens in poultry: An emerging threat for animal and public health. Avian Pathol. 2004, 33, 537–549. [Google Scholar] [CrossRef] [PubMed]
- Hosseinzadeh, S.; Bahadori, M.; Poormontaseri, M.; Dehghani, M.; Fazeli, M.; Nazifi, S. Molecular characterization of Clostridium perfringens isolated from cattle and sheep carcasses and its antibiotic resistance patterns in Shiraz slaughterhouse, southern Iran. Vet. Arh. 2018, 88, 581–591. [Google Scholar] [CrossRef]
- Beran, V.; Chmelar, D.; Vobejdova, J.; Konigova, A.; Nemec, J.; Tvrdik, J. Sensitivity to antibiotics of Clostridium difficile toxigenic nosocomial strains. Folia Microbiol. 2014, 59, 209–215. [Google Scholar] [CrossRef]
- Hmood, A.M.; Al-Shukri, M.S.; Al-Charrakh, A.H. Molecular detection and antimicrobial resistance of Clostridium perfringens isolated from diabetic patients and bullet wounds. J. Appl. Biol. Biotechnol. 2019, 7, 54–59. [Google Scholar]
- Hu, W.-S.; Kim, H.; Koo, O.K. Molecular genotyping, biofilm formation and antibiotic resistance of enterotoxigenic Clostridium perfringens isolated from meat supplied to school cafeterias in South Korea. Anaerobe 2018, 52, 115–121. [Google Scholar] [CrossRef]
- Gharaibeh, S.; Al Rifai, R.; Al-Majali, A. Molecular typing and antimicrobial susceptibility of Clostridium perfringens from broiler chickens. Anaerobe 2010, 16, 586–589. [Google Scholar] [CrossRef] [PubMed]
- Yadav, J.P.; Das, S.C.; Dhaka, P.; Vijay, D.; Kumar, M.; Mukhopadhyay, A.K.; Chowdhury, G.; Chauhan, P.; Singh, R.; Dhama, K. Molecular characterization and antimicrobial resistance profile of Clostridium perfringens type A isolates from humans, animals, fish and their environment. Anaerobe 2017, 47, 120–124. [Google Scholar] [CrossRef]
- Kiu, R.; Hall, L.J. An update on the human and animal enteric pathogen Clostridium perfringens. Emerg. Microbes Infect. 2018, 7, 1–15. [Google Scholar] [CrossRef] [Green Version]
- Mooyottu, S.; Flock, G.; Kollanoor-Johny, A.; Upadhyaya, I.; Jayarao, B.; Venkitanarayanan, K. Characterization of a multidrug resistant C. difficile meat isolate. Int. J. Food Microbiol. 2015, 192, 111–116. [Google Scholar] [CrossRef] [PubMed]
- Zhang, W.-Z.; Li, W.-G.; Liu, Y.-Q.; Gu, W.-P.; Zhang, Q.; Li, H.; Liu, Z.-J.; Zhang, X.; Wu, Y.; Lu, J.-X. The molecular characters and antibiotic resistance of Clostridioides difficile from economic animals in China. BMC Microbiol. 2020, 20, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Zowawi, H.M. Antimicrobial resistance in Saudi Arabia: An urgent call for an immediate action. Saudi Med. J. 2016, 37, 935. [Google Scholar] [CrossRef]
- Ngamwongsatit, B.; Tanomsridachchai, W.; Suthienkul, O.; Urairong, S.; Navasakuljinda, W.; Janvilisri, T. Multidrug resistance in Clostridium perfringens isolated from diarrheal neonatal piglets in Thailand. Anaerobe 2016, 38, 88–93. [Google Scholar] [CrossRef]
- Doyle, M.E. Multidrug-resistant pathogens in the food supply. Foodborne Pathog. Dis. 2015, 12, 261–279. [Google Scholar] [CrossRef] [PubMed]
- Weese, J.S.; Avery, B.P.; Rousseau, J.; Reid-Smith, R.J. Detection and enumeration of Clostridium difficile spores in retail beef and pork. Appl. Environ. Microbiol. 2009, 75, 5009–5011. [Google Scholar] [CrossRef] [Green Version]
- Woo, P.; Lau, S.; Chan, K.; Fung, A.; Tang, B.; Yuen, K. Clostridium bacteraemia characterised by 16S ribosomal RNA gene sequencing. J. Clin. Pathol. 2005, 58, 301–307. [Google Scholar] [CrossRef] [Green Version]
- Gurjar, A.; Hegde, N.; Love, B.; Jayarao, B. Real-time multiplex PCR assay for rapid detection and toxintyping of Clostridium perfringens toxin producing strains in feces of dairy cattle. Mol. Cell. Probes 2008, 22, 90–95. [Google Scholar] [CrossRef]
- Duncan, C.L.; Strong, D.H. Improved medium for sporulation of Clostridium perfringens. Appl. Microbiol. 1968, 16, 82–89. [Google Scholar] [CrossRef] [PubMed]
- CLSI. Clinical and Laboratory Standards Institute. Methods for Antimicrobial Susceptibility Testing of Anaerobic Bacteria, 9th ed.; CLSI standard M11; Clinical and Laboratory Standards Institute: Wayne, PA, USA, 2018. [Google Scholar]
- CLSI. Clinical and Laboratory Standards Institute. Performance Standards for Antimicrobial Susceptibility Testing, 30th ed.; CLSI Supplement M100; Clinical and Laboratory Standards Institute: Wayne, PA, USA, 2020. [Google Scholar]
- Singh, S.; Yadav, A.S.; Singh, S.M.; Bharti, P. Prevalence of Salmonella in chicken eggs collected from poultry farms and marketing channels and their antimicrobial resistance. Food Res. Int. 2010, 43, 2027–2030. [Google Scholar] [CrossRef]
- Magiorakos, A.-P.; Srinivasan, A.; Carey, R.; Carmeli, Y.; Falagas, M.; Giske, C.; Harbarth, S.; Hindler, J.; Kahlmeter, G.; Olsson-Liljequist, B. Multidrug-resistant, extensively drug-resistant and pandrug-resistant bacteria: An international expert proposal for interim standard definitions for acquired resistance. Clin. Microbiol. Infect. 2012, 18, 268–281. [Google Scholar] [CrossRef] [Green Version]
- Gu, Z.; Eils, R.; Schlesner, M. Complex heatmaps reveal patterns and correlations in multidimensional genomic data. Bioinformatics 2016, 32, 2847–2849. [Google Scholar] [CrossRef] [PubMed] [Green Version]




{kind=link}
{kind=link}
{kind=link}
| Sample Source | No. of Tested Samples | No. (%) of | |
|---|---|---|---|
| C. perfringens | C. difficile | ||
| Butcher shops | 50 | 10 (71.4) | 3 (75) |
| Supermarkets | 50 | 4 (28.6) | 1 (25) |
| Total | 100 | 14 (14%) | 4 (4%) |
| Clostridium spp. | Type | N | Toxin Gene | No. (%) |
|---|---|---|---|---|
| C. perfringens | A | 9 | cpa+ | 5 (35.7) |
| cpa+, cpe+ | 4 (28.6) | |||
| B | 1 | cpa+, cpb+, etx+ | 1 (7.1) | |
| C | 3 | cpa+, cpb+ | 2 (14.3) | |
| cpa+, cpb+, cpe+ | 1 (7.1) | |||
| D | 1 | cpa+, etx+ | 1 (7.1) | |
| C. difficile | A+B+ | 3 | tcdA, tcdB | 3 (75) |
| A−B+ | 1 | tcdB | 1 (25) |
| Genotype | Source | Accession No. | Resistance Patterns 1 | MAR Index 2 |
|---|---|---|---|---|
| C. perfringens type C | Supermarket | MW725399 | - | 0.00 |
| C. perfringens type A | Supermarket | MW785765 | PEN | 0.14 |
| C. perfringens type B | Supermarket | MW785767 | PEN | 0.14 |
| C. perfringens type C | Butcher shop | MW785769 | TET | 0.14 |
| C. perfringens type A | Butcher shop | MW725397 | PEN, CLI | 0.29 |
| C. perfringens type C | Butcher shop | MW725398 | CRO, TET | 0.29 |
| C. perfringens type A | Butcher shop | MW725400 | CRO, CLI | 0.29 |
| C. perfringens type A | Butcher shop | MW725401 | PEN, TET | 0.29 |
| C. perfringens type A | Butcher shop | MW785763 | CRO, CLI | 0.29 |
| C. perfringens type D | Butcher shop | MW785764 | MTZ, TET | 0.29 |
| C. perfringens type A | Butcher shop | MW785770 | CRO, TET | 0.29 |
| C. perfringens type A | Supermarket | MW725396 | CRO, CLI, TET | 0.43 |
| C. perfringens type A | Butcher shop | MW785766 | CRO, MTZ, TET | 0.43 |
| C. perfringens type A | Butcher shop | MW785768 | PEN, CRO, CLI, TET | 0.57 |
| C. difficile type A+B+ | Butcher shop | MW798269 | TET | 0.14 |
| C. difficile type A+B+ | Butcher shop | MW798268 | CLI, TET | 0.29 |
| C. difficile type A−B+ | Supermarket | MW732695 | CRO, TET | 0.29 |
| C. difficile type A+B+ | Butcher shop | MW732694 | PEN, CRO, MXF | 0.43 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Fayez, M.; El-Ghareeb, W.R.; Elmoslemany, A.; Alsunaini, S.J.; Alkafafy, M.; Alzahrani, O.M.; Mahmoud, S.F.; Elsohaby, I. Genotyping and Antimicrobial Susceptibility of Clostridium perfringens and Clostridioides difficile in Camel Minced Meat. Pathogens 2021, 10, 1640. https://0-doi-org.brum.beds.ac.uk/10.3390/pathogens10121640
Fayez M, El-Ghareeb WR, Elmoslemany A, Alsunaini SJ, Alkafafy M, Alzahrani OM, Mahmoud SF, Elsohaby I. Genotyping and Antimicrobial Susceptibility of Clostridium perfringens and Clostridioides difficile in Camel Minced Meat. Pathogens. 2021; 10(12):1640. https://0-doi-org.brum.beds.ac.uk/10.3390/pathogens10121640
Chicago/Turabian StyleFayez, Mahmoud, Waleed R. El-Ghareeb, Ahmed Elmoslemany, Saleem J. Alsunaini, Mohamed Alkafafy, Othman M. Alzahrani, Samy F. Mahmoud, and Ibrahim Elsohaby. 2021. "Genotyping and Antimicrobial Susceptibility of Clostridium perfringens and Clostridioides difficile in Camel Minced Meat" Pathogens 10, no. 12: 1640. https://0-doi-org.brum.beds.ac.uk/10.3390/pathogens10121640