
Experimental Hybridization in Leishmania: Tools for the Study of Genetic Exchange


Intracellular Parasite Biology Section, Laboratory of Parasitic Diseases, National Institute of Allergy and Infectious Diseases, NIH, Bethesda, MD 20892, USA
*
Author to whom correspondence should be addressed.
Pathogens 2022, 11(5), 580; https://0-doi-org.brum.beds.ac.uk/10.3390/pathogens11050580
Submission received: 19 April 2022
/
Revised: 11 May 2022
/
Accepted: 13 May 2022
/
Published: 14 May 2022
(This article belongs to the Special Issue Leishmania & Leishmaniasis)
Abstract
:Despite major advances over the last decade in our understanding of Leishmania reproductive strategies, the sexual cycle in Leishmania has defied direct observation and remains poorly investigated due to experimental constraints. Here, we summarize the findings and conclusions drawn from genetic analysis of experimental hybrids generated in sand flies and highlight the recent advances in generating hybrids in vitro. The ability to hybridize between culture forms of different species and strains of Leishmania should invite more intensive investigation of the mechanisms underlying genetic exchange and provide a rich source of recombinant parasites for future genetic analyses.
1. Introduction
Kinetoplastid parasites of the Leishmania genus comprise a wide diversity of species, with genetically distinct populations causing a variety of human and zoonotic diseases collectively referred as the leishmaniases; an estimated one billion people are at risk of being infected. Infections can lead to a wide spectrum of clinical presentations ranging from self-healing cutaneous lesions to visceralization and death, largely dependent on the species of parasite and the host’s immunogenetic background. The well-described heteroxenous life cycle of Leishmania involves transmission of promastigote forms through the bite of a phlebotomine sand fly insect to the mammalian host skin, where they differentiate into amastigotes inside phagocytic immune cells [1].
The question of how Leishmania reproduce remains a matter of debate, in large part because while asexual reproduction is readily apparent among life cycle stages found both in the sand fly vector and within phagocytes in the mammalian host, direct observation of sexual reproduction has thus far not been possible. This fact, along with a number of observations based on population genetics, support a predominant if not strictly clonal reproductive strategy [2]. In particular, the high linkage disequilibrium observed among different polymorphic alleles is thought to be due to the absence of mechanisms, namely gene segregation and recombination, which tend to disrupt the common inheritance of alleles on the same or different chromosomes [3]. Population genetic studies, however, have been used to provide evidence for sex, whereby multi locus genotyping and whole genome sequencing analysis of natural isolates have identified hybrids between strains of closely related species and even between divergent species [4,5,6,7,8]. The first experimental demonstration that Leishmania strains can hybridize in a manner consistent with meiotic sex was provided by Akopyants et al. in 2009 [9], in which two recombinant lines of Leishmania major each bearing a different antibiotic resistance gene were used to co-infect sand flies, followed by recovery and cloning of midgut promastigotes that were resistant to both drugs. The hybrid progeny had inherited multiple alleles from both parents. These studies formally demonstrated that Leishmania possess the machinery for genetic exchange and identified promastigotes in the sand fly midgut as a life cycle stage in which hybridization can occur. Following this initial report, experimental intraspecific, interspecific, and intraclonal hybrids have been recovered from or observed in sand flies co-infected with recombinant lines of Leishmania containing different antibiotic resistance and/or fluorescence genes [10,11,12,13,14]. Most recently, both ourselves and others have reported that axenic culture forms of promastigotes can form stable hybrids entirely in vitro [15,16,17]. Possible hybrids formed between intracellular amastigotes residing in a common macrophage vacuole have been described as well [18]. Thus, the continuing debate regarding clonality versus sexuality in Leishmania pertains mainly to the frequency of hybridization, the mode of genetic exchange (whether there a true sexual cycle), and the degree to which recombination has been able to break the prevalent pattern of a clonal population structure [19].
It is important to point out that in the absence of direct observations of cell fusion, nuclear fusion, and meiosis producing haploid gametic cells, conclusions regarding a true sexual cycle in Leishmania based on observations of hybrid genotypes can only be inferred. This review will focus on the biology of the hybrids which have been generated experimentally and which have been analyzed by high resolution whole-genome sequencing in order to inform the mode of genetic exchange. We summarize the approaches current being used to dissect the involvement of meiotic gene orthologs in genetic exchange and to identify and select for mating competent cells. Finally, the potential of experimental hybridization for linkage studies and positional cloning of important genes is critically discussed.
2. Experimental Leishmania Hybrids in Sand Flies
Formal demonstration of genetic exchange in Leishmania spp. during their growth and development in the sand fly vector has been provided by different research groups, and it is widely accepted that inter- and intraspecific hybrids can be generated experimentally in sand flies. The lack of availability of colonized sand flies to most Leishmania research programs and the low frequency at which these hybridizations take place in colonized flies represents a major challenge in the field. In experimental designs where parental lines have a single allele replaced by a selectable marker, only 25% of possible mating events are tracked. In addition, frequencies of hybrid recovery from laboratory sand flies are highly variable and species-dependent (both Leishmania and sand fly). Double drug resistant hybrids have been recovered in 7–26% of infected sand flies, either Phlebotomus duboscqi (natural L. major vector) or Lutzomyia longipalpis (permissive vector) for intraspecies L. major crosses, 34–65% in L. tropica, and 3.2% for inter-species L. major × L. infantum [11,12,13]. The relatively low frequency of hybrid recovery may be related to the use of culture-adapted recombinant parasites for hybrid selection, and it is unknown whether fresh clinical isolates might hybridize with greater efficiency in the vector.
Genetic exchange analyses thus far have relied on the introduction of two different drug resistance markers in each of the parental strains to allow for selection of double drug resistance hybrids. This is an important constraint of the current experimental model, as hybrids must stably grow as promastigotes in the presence of both selection antibiotics for several generations. The use of parental lines that overexpress fluorescent proteins with discernable spectra can provide quick PCR-free initial confirmation of hybridization using flow cytometry analysis or confocal microscopy. However, co-expression of parental fluorescent proteins requires additional confirmation. It is critical to be able to distinguish between a product of genetic exchange and an event of cytoplasmic content sharing. Exchange of flagellar and cytosolic proteins has been shown in T. brucei via transient flagellar membrane fusion or extracellular vesicle endocytosis to give rise to trypanosomes carrying exogenous proteins stable for up to three days [20,21]. Although protein exchange can occur between T. brucei haploid cells, there is no transfer of genetic material during this process [20]. Cell-to-cell communication is an underexplored subject in Leishmania research, and it is possible that it takes place inside restricted compartments, such as in the sand fly thoracic midgut, or intraphagosomally in the case of L. mexicana or L. amazonensis, which share the same host cell vacuole. Single-cell spatial genomic analysis of mixed infections is a potential way forward to acquiring in situ information and avoiding the requirement of parasite growth in culture.
Analysis of parental SNP inheritance has been reliably used for tracking biallelic segregation in Leishmania genetic exchange in several experimental crosses (Figure 1). It has been demonstrated by multilocus genotyping methods ranging from PCR amplification followed by Sanger sequencing, SNP–cleaved amplification polymorphic site (SNP-CAPS) analysis, Southern Blotting, and bulk Whole Genome Sequencing (bWGS) [9,11,22]. In 2009, PCR-based approaches were used to confirm the first demonstration of hybrid generation in laboratory sand flies between two L. major strains, namely, Fn and LV39 [9]. All of the eighteen L. major hybrids showed biparental inheritance of the seven chromosomes tested. These findings were extended to 96 additional hybrids generated in sand flies, involving different pairwise combinations of L. major strains obtained from across the geographic range of this species [11]. Subsequently, the first experimental inter-species hybrids were described, with eleven hybrids found to inherit both L. major and L. infantum parental alleles in all but one of the chromosomes tested [13]. Thus, there is no evidence to date for mating types that might limit genetic exchange; experimentally, at least, there is no species barrier to mating between strains. Importantly, attempts to recover double drug resistant hybrids following co-infection of mice using the same recombinant L. major parental lines that hybridized in the vector were unsuccessful [9], suggesting that the sand fly is where sex happens. The descriptions of possible in vitro hybridization involving amastigotes of the L. mexicana complex residing in a communal vacuole inside macrophages, while extremely interesting, could not be genetically validated because of instability in the growth of the double drug resistant lines [18]. In these studies, co-infection of mice with the parental lines failed to yield recoverable hybrids.
The initial genotyping of the experimental hybrids provided evidence that they were full-genome hybrids based on their bi-parental inheritance of the limited number of markers tested. Subsequently, all of the 36 chromosomes in the genomes of the intra- and interspecific hybrids were investigated in detail using Illumina short-read bWGS [12]. Genome-wide analysis of the SNPs that are homozygous and different between the parents revealed that 99% of the disomic chromosomes in the progeny had a parental-derived SNP frequency close to 0.5, inferring a near full genome hybridization with the allele contributions from each parent matching the inheritance patterns expected under meiosis. The bWGS analysis of the parents and hybrid clones revealed the copy number of the individual chromosomes and the patterns of somy inheritance. While Leishmania are largely diploid, all Leishmania strains show varying degrees of aneuploidy, including mosaic aneuploidy, which refers to somy variations that are present within clonal populations [23]. Thus, the analysis of somy at the population level, as in the published studies, computed average values. In almost every case, chromosomes that were present in close to three copies in either parent were transmitted to the hybrid progeny in either one or two copies and in frequencies expected under meiosis. Finally, bWGS analysis of L. major backcross progeny clones provided the first clear evidence for genome-wide recombination events in Leishmania, demonstrating that classical crossing over occurs at meiosis. In the absence of direct observation of haploid gametic cells, the inheritance patterns revealed by WGS analysis in experimental hybrids remain the strongest argument in favor of meiosis-like sexual recombination in the Leishmania genus.
The conclusions regarding a meiosis-like program controlling genetic exchange in Leishmania is complicated to a certain extent by the unexpected patterns of chromosome inheritance observed in hybrid progeny. The exceptions to Mendelian ratios included chromosomes that showed uniparental inheritance and changes in chromosome copy numbers observed either at the level of individual somy or whole ploidy. Thus, 2% of the chromosomes in the L. major intra-specific hybrids were trisomic despite the chromosomes being disomic in both parents, and 0.7% of the chromosomes showed alleles from only one parent, for which a loss of heterozygosity was inferred [12]. More common to the experimental hybrids was polyploidy, which mirrors findings in T. brucei, for which triploid hybrids are a familiar product of experimental matings [24]. Out of 53 L. major intra-specific hybrids described between different diploid parental combinations, 39 were diploid, 13 were triploid and one was tetraploid [9,11]. Similar variation was shown for inter-species hybrids. From eleven hybrids between L. major and L. infantum, five were diploid, five were triploid, and one was tetraploid [13]. The gene dosage difference due to chromosome copy number variation was reflected in the phenotypic segregation observed, with the hybrids that acquired their extra genome from L. major producing skin lesions and no dissemination, while the hybrids that acquired their extra genome from L. infantum produced strong dissemination to and growth in the viscera no skin pathology. This suggests that tissue tropism is a heritable trait and a potential target for linkage analysis, as discussed in detail below.
Polyploidy is an especially prevalent phenomenon for in vitro generated hybrids. Out of 24 L. tropica in vitro hybrids generated between two diploid parental lines, five were diploid, fifteen were triploid, and four were tetraploid [17]. All three in vitro hybrids from a diploid L. donovani × L. major cross were triploid, and two in vitro hybrids obtained from a diploid L. donovani × triploid L. tropica cross were tetraploid [15]. In the case of a recently published method of irradiation-facilitated in vitro hybridization (discussed below), almost all of the progeny were tetraploid [16]. Although classical meiosis entails the production of haploid gametic cells after reductional division and subsequent fusion of two different gametes, alternative models can be proposed to explain the generation of polyploid hybrids. The generation of triploid progeny may involve the production of haploid gametes in one parent and the failure of meiosis in the other parent such that diploid and haploid cells fuse, as has been proposed for T. brucei [24]. The formation of tetraploid progeny would suggest the fusion of diploid cells, possibly unreduced gametes, or an aberrant form of hybridization that is unrelated to a meiotic program in either parent. It is possible that the polyploid hybrids reflect an unconventional mode of genetic exchange, such as the parasexual process that occurs in certain fungi [25].
3. Kinetoplast DNA Inheritance
Genome-wide analyses revealed biallelic inheritance of the parental nuclear genomes in Leishmania F1 progeny for several different crosses performed experimentally (Figure 1). Conversely, inheritance of Leishmania extranuclear DNA present in the single mitochondrion (kinetoplast DNA or kDNA) has long been considered uniparental, similar to what is seen in late-branching eukaryotes [26]. Recent evidence suggests a different scenario, with components of the kDNA network showing either biparental or uniparental contribution in experimental and putative natural hybrids [8,15].
The Leishmania kDNA is comprised of two main elements: dozens of ~20-kb maxicircles (20–50 molecules in L. tarentolae) and thousands of ~0.9-kb minicircles (10,000–20,000 molecules in L. tarentolae) concatenated together in a mesh network of circular DNA [27]. Maxicircles are the eukaryotic mitochondrial DNA equivalent, carrying eighteen protein-coding genes involved in the respiratory chain and mitoribosome structure, one 12S and one 9S mitochondrial rRNAs. Each minicircle encodes a guide RNA (gRNA) that targets mitochondrial mRNAs in a highly specialized post-transcriptional regulatory mechanism. Twelve of the eighteen maxicircle-encoded mRNAs require the addition or deletion of uridines (U) for expression in an RNA editing process involving the annealing of gRNAs to the target transcript and the activity of an RNA editing core complex [27,28,29].
The inheritance patterns of the full set of genetic components of the kDNA have until recently been under-explored. The use of parental maxicircle polymorphisms in inter-strain crosses as a proxy for overall kDNA inheritance in the early analyses contributed to the inaccurate inference of uniparental kDNA segregation [9]. In fact, recent evidence in T. brucei and Leishmania suggests that while maxicircle contribution is uniparental, minicircle inheritance is biparental [8,30]. The same has been observed for in vitro Leishmania hybrids [15,16]. It is proposed that maxicircle composition in hybrids is derived from both parents upon completion of genetic exchange, while rapid DNA segregation in the following mitotic divisions may lead to the observed loss of contribution by one of the parents. Such loss of complexity is observable in minicircles [8,31], although the effects are less drastic as these are present in thousands of copies and hundreds of minicircle classes (MSCs). MSCs are groups of highly similar minicircle molecules distinguishable within the same Leishmania population by 1–5% sequence mismatches (95–99% sequence identity) [32]. Recent findings confirm the long-held notion of minicircle copy number plasticity and the loss of non-essential minicircles in cultured trypanosomatids due to the lack of specific environmental pressure [31]. Genetic exchange would presumably play a significant role in restoring gRNA complexity, preventing the loss of essential genes, and maintaining correct mitochondrial gene expression and metabolism throughout the parasite’s life cycle [8].
Mito-nuclear discordance is therefore a relevant signature of genetic exchange in trypanosomatids, and its detection provides a marker for past hybridization events in interspecific Leishmania hybrids in nature [32,33]. Extranuclear DNA inheritance in other protists such as Plasmodium is maternal, as male gametes typically lack both plastids and mitochondria [34], similar to products of meiosis in vertebrates. The evidence of parental mitochondrial fusion and the lack of demonstrated heterogamy in Leishmania and T. brucei support the hypothesis that meiosis produces morphologically indistinguishable sexual stages (isogamy) in trypanosomatids. The in vitro hybridization methods recently described in Leishmania (discussed in the next section) may potentially provide a feasible way to study the mechanisms that orchestrate kinetoplast fusion and the kinetics of maxicircle/minicircle segregation. In addition, recent efforts have generated specific bioinformatics packages in R and Python to support the analysis of trypanosomatid minicircle composition from next-generation sequencing data [31,32].
It is unclear whether a more homogeneous minicircle composition could be detected in Leishmania inside the sand fly midgut or in infected host samples. Minicircle diversity is most likely not uniform in the cultured Leishmania promastigote population, as MSCs are expected to change after successive rounds of mitosis in a stochastic manner. For the study of nuclear genome heterogeneity, single-cell WGS (scWGS) has been used to describe the rise of different nuclear karyotypes within a clonal L. donovani culture [35]. Similarly, analysis of single-cell kDNA or single-kinetoplast composition is important for a comprehensive understanding of minicircle heterogeneity, MSC cooperative functioning, and maxicircle–minicircle combinations in Leishmania populations.
4. Novel Tools for Generating Sand Fly-Free Leishmania Hybrids
4.1. Leishmania In Vitro Mating
Sexual reproduction in Leishmania appears to be a non-obligatory part of its transmission cycle, and it is unknown what triggers the commitment to diverge from the mostly unidirectional asexual promastigote growth and differentiation program to a sexual stage in the insect vector. A major obstacle is the availability of laboratory sand fly colonies, which are restricted to a handful of research groups worldwide. Because sand fly rearing is laborious and resource-consuming, the in vitro hybridization methods recently reported may become a helpful tool for dissecting the cryptic Leishmania sexual cycle. In 2020, in vitro generation of bona fide diploid Leishmania full genomic hybrids was confirmed for the first time [17]. The most critical factor was the use of L. tropica as the parental species. As far as we are aware, these are the only crosses in which Leishmania diploid hybrids have been generated by in vitro mating. A second study has recently confirmed the comparatively high mating competency of L. tropica in vitro using a different strain [15]. It is unclear why L. tropica promastigotes have a remarkably high capacity to undergo experimental genetic exchange both in vitro and in vivo (in 34–65% of infected sand flies) [11]. The experimental mating efficiency seems to reflect the elevated genomic diversity of L. tropica isolates and the prevalence of described natural hybrids from Asia carrying an L. tropica haplotype [36,37,38,39] Nevertheless, the hybridization rate of L. tropica in vitro is significantly lower than in sand flies, with an estimated minimum frequency of hybridization in the order of magnitude of 10−8 vs. 10−4, respectively.
In our hands, L. tropica was the only mating-competent species in vitro that did not require exposure to DNA stress conditions. However, in a herculean effort, Gutierrez-Corbo et al. reported the recovery of two L. donovani × L. tropica and three L. donovani × L. major hybrids after testing 3072 culture wells for each cross in different conditions [15]. The in vitro conditions included using promastigotes in different growth phases and culturing the promastigotes in the presence of an insect feeder cell line derived from the sand fly Lutzomyia longipalpis. As the hybridization frequencies were extremely low and not reproducible in independent experiments, it was not possible to draw conclusions from these studies regarding the culture conditions that might promote hybridization.
4.2. DNA Stress as a Trigger for Leishmania Mating In Vitro
In 2022, a modified protocol provided a reproducible enhanced recovery rate of in vitro hybrids by several times over through the treatment of parental Leishmania cells with low levels of DNA-damaging agents prior to co-culture [16]. The rationale for these studies was that in organisms that are known to be facultatively sexual, they can be triggered to initiate their sexual cycle by conditions that produce DNA damage [40,41,42]. In our studies, DNA stress was induced by oxidative damage with hydrogen peroxide, by DNA breaks with methyl methanesulfonate (MMS), or by DNA breaks by ionizing gamma radiation. In contrast to the previous approach, the generation of irradiation-facilitated hybrids (referred to here as iHybrids) is a species-agnostic method (L. major being the exception thus far), supporting the feasible exploration of meiotic gene function in axenic Leishmania culture.
L. tropica exposed to 6.5 Gy of gamma-radiation revealed an increase in hybrid recovery of >100-fold in vitro [16]. The iHybrid approach allowed for the in vitro generation of intraspecific hybrids of L. braziliensis and L. donovani as well as interspecific hybrids between L. tropica and L. infantum (Figure 1). Trypanosomatids are highly tolerant to gamma radiation, and viable parasites can be recovered after exposures of 500 Gy [43]. Stress conditions inside the vector and mammalian host may have contributed to the selection of pathways to withstand DNA damage in these parasites. In the sand fly midgut, Leishmania are exposed to different sources of DNA stress. Blood meal hemoglobin digestion releases heme, which can then be hydrolyzed by heme oxygenase to free Fe2+, leading to reactive oxygen species (ROS) and oxidative DNA damage [44]. We speculate that ROS levels in the digested blood meal below a threshold level that is lethal for the parasites or sand fly [45] may be able to trigger repairable DNA damage and genetic exchange. Alternatively, spontaneous DNA breaks are often associated with replicative stress [46]. In the first two days post-blood meal, the sand fly midgut contains proliferating promastigotes marked by DSBs catalyzed during the S phase. This is consistent with the finding that the frequency of in vitro hybridization has thus far been found to peak when the co-cultures are initiated with parental lines in the exponential growth phase.
The major repair mechanism of DNA double-strand breaks (DSB) involves the non-homologous end joining (NHEJ) pathway, which ligates the ends of the break in order to fully restore it. It is widely accepted that trypanosomatids lack a canonical NHEJ and that homologous recombination (HR) is instead the most important DSB repair pathway, although the mechanisms remain unclear (see review in [46]). The drastic increase in in vitro hybridization post-irradiation suggests an interplay between the DNA break repair machinery and meiosis in these parasites, ultimately leading to upregulation of genes involved in the fusion of gametes and nuclei.
4.3. In Vitro Hybridization Provides Novel Insights into Leishmania Mating
Endogenous programmed DSBs comprise the first step in a process that initiates meiotic recombination between homologous chromosomes in eukaryotes in general [47]. At the start of the eukaryotic meiotic prophase I, a zipper-like multiprotein structure named the synaptonemal complex (SC) is formed containing a central element and an axial (lateral) element. The latter includes meiosis-specific factors such as Hop1 (HORMA domain-containing protein 1), Red1, and Rec8/Rad21 for homologous chromosome pairing. Spo11 is the effector topoisomerase-like protein in the DSB machinery, which cleaves the genomic DNA at non-random spots and remains covalently bound to both DNA ends until processing by the Mre11-Rad50-NBS1 complex. The DNA-binding protein RPA covers the single-strand ends, and is later replaced by recombinases Dmc1 (disrupted meiotic cDNA 1) and Rad51 (radiation-sensitive 51), which invade the homologous chromosome and form a D-loop structure mediated by the Hop2-Mnd1 (homologous-pairing protein 2—meiotic nuclear division protein 1) complex [48]. The chromosome synapsis is stabilized by Hop2-Mnd1, which inhibits the interaction between non-homologous chromosomes.
Orthologs of all the meiosis-related proteins referenced here have been found in the Leishmania genome [49,50]. Whether or not they retain their conventional meiosis-specific functions in these parasites is unclear. Thus far, Rad51expression has been shown to be upregulated in L. tropica promastigotes during irradiation-induced mating in vitro and in T. cruzi cell-fused hybrids [16,43]. Apart from the well-conserved components mentioned here, it has been a challenging task to identify homologues of all the meiotic machinery in Leishmania. Recent in silico analyses have identified the putative trypanosomatid orthologs of SYCPs (synaptonemal complex proteins), structural components of the vertebrate axial element of the SC. Interestingly, the highest similarities were found between SYCP2-SYCP3 and KKT16-KKT18 proteins, which form the kinetoplastid kinetochore responsible for the mitotic segregation of sister chromatids. It has been proposed that the ancestral kinetoplastid kinetochore was repurposed from meiotic SC structural components [51].
At the end of prophase I, the Spo11-catalyzed DSB is repaired upon recombination and crossover events are resolved. In protists such as T. brucei, for which classical gametic meiosis has been observed [52], following a pre-meiotic S phase, meiosis I progresses until segregation of chromosome homologs into two haploid 1n2c daughter cells. If both cells enter classical meiosis II, equational division culminates in the generation of four haploid daughter gametic cells (1n1c). The evidence that polyploid experimental Leishmania hybrids are formed along with diploids, particularly in the iHybrid progeny, raises the possibility of two different mechanisms of genetic exchange in Leishmania, one of which does not involve meiosis I; however, definitive evidence that polyploid hybrids are a result of incomplete meiosis or of a non-meiotic process such as that described for Candida albicans parasexual mating remains lacking [25]. In addition to the difficulty of identifying many homologues of eukaryotic meiotic components in Leishmania by their primary amino acid sequence similarity, the lack of a high-throughput experimental model to test null mutants has been a persistent challenge.
The DNA damage-induced approach is not an exclusive feature of trypanosomatids. DSB causing agents have been described as initiating meiosis in multiple microorganisms and partially replacing Spo11 function [40,53,54]. In social amoebas such as dictyostelids, an unknown mechanism completely independent of Spo11 triggers meiotic recombination [55]. The generation of Leishmania iHybrids may bypass the requirement of endogenous Spo11 activity as a trigger of meiosis; thus, evaluation of gene functions directly related to the meiotic DSB machinery may not be appropriate using this method. However, it seems fitting for functional analysis of genes that have a role in later steps such as gamete fusion.
Regardless of the precise mechanism involved, gametic cell membrane fusion (plasmogamy) and nuclei fusion (karyogamy) in protists, fungi, and other microorganisms are marked by the expression of Hap2/Gcs1 and Gex1/Kar5 genes, respectively. Hap2/Gcs1 (Hapless 2/Generative cell specific 1 protein) is an ancestral gamete fusogen found in non-vertebrates which has been widely used as a marker for gametic cells, including Plasmodium and T. brucei [56,57]. Its structure resembles that of flavivirus class II membrane fusion proteins [58]. Gex1/Kar5 (gamete expressed 1/karyogamy protein 5) plays a central role in nuclear envelope fusion during budding yeast mating [59]. Single-cell RNA-sequencing (scRNA-seq) identified Hap2 and Gex1 upregulated in a discrete cell population induced by irradiation treatment of L. tropica parental strains [16]. Sorting of promastigotes expressing a Hap2-mNeonGreen fusion led to the isolation of both mating-competent (Hap2+ cells) and mating-incompetent (Hap2−) promastigotes from axenic culture. Experimental evidence of the implication of other specific genes directly in meiotic recombination using this in vitro model will hopefully help to shed light on the sexual reproduction mechanism in Leishmania.
5. Quantitative Traits and Gene Linkage Analysis
Apart from the potential of in vitro hybridization to inform the underlying mechanisms of genetic exchange in Leishmania, the ability to easily generate large numbers of recombinant parasites holds enormous promise for future genetic analyses seeking to identify the genes controlling important traits. Reverse genetic approaches, in which a gene of interest can be specifically targeted for deletion or overexpression, have thus far had only modest success in identifying the strain- and species-specific genes controlling such critical characters as host range, tissue tropism, and pathogenicity. Future genetic studies offer an unbiased approach to finding causative associations between DNA polymorphisms and phenotypic traits using bi-parental quantitative trait loci (QTL) mapping. QTL mapping can reveal the network of genes that control a complex phenotype. Quantitative traits are frequently more easily examined when gene sequences are isolated from other genomic regions in a different genetic background. To this end, backcrossing is a powerful resource in forward genetics. The evidence that experimental L. major backcrosses are possible, at least in vivo, suggests its use as a potential genetic tool for positional cloning and linkage analyses [12]. Thus far, the ability to produce backcross progeny is restricted to intraspecific crosses. Despite several attempts, backcrosses were not obtained when crossing the L. major × L. infantum F1 hybrids with the L. major parental line, leading to the conclusion that interspecies hybrids might be sterile [12]. It remains to be tested whether the same is true for other interspecies pairwise combinations in vivo or for irradiation-induced hybridization in vitro. Another challenge to be overcome in the in vitro hybrids in order to facilitate their use in future genetic studies is that all iHybrids described to date are polyploid, potentially restricting the number of genomic regions where parental genetic features can be isolated through recombination. Nonetheless, partial loss of heterozygosity was detected in F1 iHybrids in several parts of the genome leading to homozygous contribution of a parental genome [17]. It is hoped that as more effort is directed at manipulating the conditions of in vitro hybridization, the induction of a more complete meiotic program inclusive of the ploidy reduction steps and genome-wide recombinations between homologous chromosomes will be possible.
Finally, the utility of experimental hybrids for QTL mapping requires that the readouts for phenotypes of interest are experimentally available. As the traits of greatest interest pertain to the diversity of clinical outcomes that different strains and species of Leishmania can produce, and which in certain cases can be modeled in a murine host, we have listed the crosses that we have carried out to date in sand flies and in vitro that are suitable for linkage analysis (Table 1).
6. Conclusions
Hybridization in Leishmania, although inferred by population genetic studies and formally demonstrated by the generation of hybrids in the laboratory, thus far lacks conclusive evidence that it is driven by a sexual process. Meiosis-like sexual recombination is supported by the presence of meiotic orthologs in the genome, and is the most likely system to account for the patterns of chromosome inheritance and recombinations revealed by bWGS of experimental hybrids generated in sand flies. Bi-parental inheritance of kDNA has been observed, although only the maxicircle kDNA from one parent appears to be retained during subsequent mitotic divisions. Progress in understanding the reproductive biology of Leishmania has been hampered by the need for both vector sand flies and genetically engineered parental strains to generate hybrids and the fact that Leishmania are facultatively sexual, yielding only a low frequency of hybrids in the crosses carried out to date. A major recent advance is the ability to generate stable hybrids entirely in vitro. By submitting culture promastigotes to DNA stress conditions, the frequency of hybridization and the ability to hybridize within and between different Leishmania species can be markedly enhanced. A transcriptionally unique population of hybridization-competent promastigotes expressing a number of meiotic gene orthologs has been identified following exposure to gamma radiation. Thus, with this expanded toolset any laboratory able to axenically grow Leishmania insect stages, i.e., virtually every Leishmania laboratory should now be able to generate recombinant parasites for genotype and phenotype analyses. Challenges remain in manipulating culture conditions to promote completion of their meiotic program, as the available in vitro hybrids are predominantly polyploid, in contrast to their diploid counterparts generated in sand flies. Further effort using in vitro protocols will hopefully lead to the identification and enrichment of the putative gametic cells and to direct observations of their meiotic cycle, including reductional division and the fusion of cells, nuclei, and kinetoplasts. Other outstanding questions related to genetic exchange could be addressed as well, including the inheritance of extrachromosmal circular DNA encoding drug resistance and the transmission or purging of intrinsic Leishmania RNA viruses. Perhaps most promising is the exploitation of in vitro hybridization for forward genetic analysis to map genes controlling the extraordinary diversity of the genus with respect to the range of infected hosts, tissue involvement, and pathologies associated with different strains and species of Leishmania.
Author Contributions
Conceptualization, T.R.F. and D.L.S.; writing, T.R.F. and D.L.S. All authors have read and agreed to the published version of the manuscript.
Funding
This work was supported in part by the Intramural Research Program of the National Institute of Allergy and Infectious Diseases, National Institutes of Health.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Kaye, P.; Scott, P. Leishmaniasis: Complexity at the host-pathogen interface. Nat. Rev. Microbiol. 2011, 9, 604–615. [Google Scholar] [CrossRef] [PubMed]
- Rougeron, V.; De Meeus, T.; Banuls, A.L. A primer for Leishmania population genetic studies. Trends Parasitol. 2015, 31, 52–59. [Google Scholar] [CrossRef] [PubMed]
- Tibayrenc, M.; Ben Abderrazak, S.; Guerrini, F.; Banuls, A. Leishmania and the clonal theory of parasitic protozoa. Arch. Inst. Pasteur Tunis 1993, 70, 375–382. [Google Scholar] [PubMed]
- Nolder, D.; Roncal, N.; Davies, C.R.; Llanos-Cuentas, A.; Miles, M.A. Multiple hybrid genotypes of Leishmania (viannia) in a focus of mucocutaneous Leishmaniasis. Am. J. Trop. Med. Hyg. 2007, 76, 573–578. [Google Scholar] [CrossRef] [PubMed]
- Odiwuor, S.; De Doncker, S.; Maes, I.; Dujardin, J.C.; Van der Auwera, G. Natural Leishmania donovani/Leishmania aethiopica hybrids identified from Ethiopia. Infect. Genet. Evol. 2011, 11, 2113–2118. [Google Scholar] [CrossRef] [PubMed]
- Ravel, C.; Cortes, S.; Pratlong, F.; Morio, F.; Dedet, J.P.; Campino, L. First report of genetic hybrids between two very divergent Leishmania species: Leishmania infantum and Leishmania major. Int. J. Parasitol. 2006, 36, 1383–1388. [Google Scholar] [CrossRef]
- Rogers, M.B.; Downing, T.; Smith, B.A.; Imamura, H.; Sanders, M.; Svobodova, M.; Volf, P.; Berriman, M.; Cotton, J.A.; Smith, D.F. Genomic confirmation of hybridisation and recent inbreeding in a vector-isolated Leishmania population. PLoS Genet. 2014, 10, e1004092. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Van den Broeck, F.; Savill, N.J.; Imamura, H.; Sanders, M.; Maes, I.; Cooper, S.; Mateus, D.; Jara, M.; Adaui, V.; Arevalo, J.; et al. Ecological divergence and hybridization of Neotropical Leishmania parasites. Proc. Natl. Acad. Sci. USA 2020, 117, 25159–25168. [Google Scholar] [CrossRef]
- Akopyants, N.S.; Kimblin, N.; Secundino, N.; Patrick, R.; Peters, N.; Lawyer, P.; Dobson, D.E.; Beverley, S.M.; Sacks, D.L. Demonstration of genetic exchange during cyclical development of Leishmania in the sand fly vector. Science 2009, 324, 265–268. [Google Scholar] [CrossRef] [Green Version]
- Calvo-Alvarez, E.; Alvarez-Velilla, R.; Jimenez, M.; Molina, R.; Perez-Pertejo, Y.; Balana-Fouce, R.; Reguera, R.M. First evidence of intraclonal genetic exchange in trypanosomatids using two Leishmania infantum fluorescent transgenic clones. PLoS Negl. Trop. Dis. 2014, 8, e3075. [Google Scholar] [CrossRef]
- Inbar, E.; Akopyants, N.S.; Charmoy, M.; Romano, A.; Lawyer, P.; Elnaiem, D.E.; Kauffmann, F.; Barhoumi, M.; Grigg, M.; Owens, K.; et al. The Mating Competence of Geographically Diverse Leishmania major Strains in Their Natural and Unnatural Sand Fly Vectors. PLoS Genet. 2013, 9, e1003672. [Google Scholar] [CrossRef] [PubMed]
- Inbar, E.; Shaik, J.; Iantorno, S.A.; Romano, A.; Nzelu, C.O.; Owens, K.; Sanders, M.J.; Dobson, D.; Cotton, J.A.; Grigg, M.E.; et al. Whole genome sequencing of experimental hybrids supports meiosis-like sexual recombination in Leishmania. PLoS Genet. 2019, 15, e1008042. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Romano, A.; Inbar, E.; Debrabant, A.; Charmoy, M.; Lawyer, P.; Ribeiro-Gomes, F.; Barhoumi, M.; Grigg, M.; Shaik, J.; Dobson, D.; et al. Cross-species genetic exchange between visceral and cutaneous strains of Leishmania in the sand fly vector. Proc. Natl. Acad. Sci. USA 2014, 111, 16808–16813. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sadlova, J.; Yeo, M.; Seblova, V.; Lewis, M.D.; Mauricio, I.; Volf, P.; Miles, M.A. Visualisation of Leishmania donovani fluorescent hybrids during early stage development in the sand fly vector. PLoS ONE 2011, 6, e19851. [Google Scholar] [CrossRef]
- Gutierrez-Corbo, C.; Dominguez-Asenjo, B.; Perez-Pertejo, Y.; Garcia-Estrada, C.; Bello, F.J.; Balana-Fouce, R.; Reguera, R.M. Axenic interspecies and intraclonal hybrid formation in Leishmania: Successful crossings between visceral and cutaneous strains. PLoS Negl. Trop. Dis. 2022, 16, e0010170. [Google Scholar] [CrossRef]
- Louradour, I.; Ferreira, T.R.; Duge, E.; Karunaweera, N.; Paun, A.; Sacks, D. Stress conditions promote Leishmania hybridization in vitro marked by expression of the ancestral gamete fusogen HAP2 as revealed by single-cell RNA-seq. Elife 2022, 11, e73488. [Google Scholar] [CrossRef]
- Louradour, I.; Ferreira, T.R.; Ghosh, K.; Shaik, J.; Sacks, D. In Vitro Generation of Leishmania Hybrids. Cell Rep. 2020, 31, 107507. [Google Scholar] [CrossRef]
- Telittchenko, R.; Descoteaux, A. Study on the Occurrence of Genetic Exchange among Parasites of the Leishmania mexicana Complex. Front. Cell Infect. Microbiol. 2020, 10, 607253. [Google Scholar] [CrossRef]
- Tibayrenc, M.; Ayala, F.J. How clonal are Trypanosoma and Leishmania? Trends Parasitol. 2013, 29, 264–269. [Google Scholar] [CrossRef]
- Imhof, S.; Fragoso, C.; Hemphill, A.; von Schubert, C.; Li, D.; Legant, W.; Betzig, E.; Roditi, I. Flagellar membrane fusion and protein exchange in trypanosomes; a new form of cell-cell communication? F1000Research 2016, 5, 682. [Google Scholar] [CrossRef] [Green Version]
- Szempruch, A.J.; Dennison, L.; Kieft, R.; Harrington, J.M.; Hajduk, S.L. Sending a message: Extracellular vesicles of pathogenic protozoan parasites. Nat. Rev. Microbiol. 2016, 14, 669–675. [Google Scholar] [CrossRef] [PubMed]
- Coelho, A.C.; Leprohon, P.; Ouellette, M. Generation of Leishmania hybrids by whole genomic DNA transformation. PLoS Negl. Trop. Dis. 2012, 6, e1817. [Google Scholar] [CrossRef] [PubMed]
- Sterkers, Y.; Lachaud, L.; Bourgeois, N.; Crobu, L.; Bastien, P.; Pages, M. Novel insights into genome plasticity in Eukaryotes: Mosaic aneuploidy in Leishmania. Mol. Microbiol. 2012, 86, 15–23. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gibson, W.; Garside, L. Genetic exchange in Trypanosoma brucei brucei: Variable chromosomal location of housekeeping genes in different trypanosome stocks. Mol. Biochem. Parasitol. 1991, 45, 77–89. [Google Scholar] [CrossRef]
- Forche, A.; Alby, K.; Schaefer, D.; Johnson, A.D.; Berman, J.; Bennett, R.J. The parasexual cycle in Candida albicans provides an alternative pathway to meiosis for the formation of recombinant strains. PLoS Biol. 2008, 6, e110. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Birky, C.W., Jr. Uniparental inheritance of mitochondrial and chloroplast genes: Mechanisms and evolution. Proc. Natl. Acad. Sci. USA 1995, 92, 11331–11338. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Simpson, L.; Douglass, S.M.; Lake, J.A.; Pellegrini, M.; Li, F. Comparison of the Mitochondrial Genomes and Steady State Transcriptomes of Two Strains of the Trypanosomatid Parasite, Leishmania tarentolae. PLoS Negl. Trop. Dis. 2015, 9, e0003841. [Google Scholar] [CrossRef]
- Blum, B.; Bakalara, N.; Simpson, L. A model for RNA editing in kinetoplastid mitochondria: “guide” RNA molecules transcribed from maxicircle DNA provide the edited information. Cell 1990, 60, 189–198. [Google Scholar] [CrossRef]
- Urrea, D.A.; Triana-Chavez, O.; Alzate, J.F. Mitochondrial genomics of human pathogenic parasite Leishmania (Viannia) panamensis. PeerJ 2019, 7, e7235. [Google Scholar] [CrossRef] [Green Version]
- Kay, C.; Peacock, L.; Williams, T.A.; Gibson, W. Signatures of hybridization in Trypanosoma brucei. PLoS Pathog. 2022, 18, e1010300. [Google Scholar] [CrossRef]
- Cooper, S.; Wadsworth, E.S.; Ochsenreiter, T.; Ivens, A.; Savill, N.J.; Schnaufer, A. Assembly and annotation of the mitochondrial minicircle genome of a differentiation-competent strain of Trypanosoma brucei. Nucleic Acids Res. 2019, 47, 11304–11325. [Google Scholar] [CrossRef]
- Geerts, M.; Schnaufer, A.; Van den Broeck, F. rKOMICS: An R package for processing mitochondrial minicircle assemblies in population-scale genome projects. BMC Bioinform. 2021, 22, 468. [Google Scholar] [CrossRef] [PubMed]
- Kato, H.; Caceres, A.G.; Gomez, E.A.; Tabbabi, A.; Mizushima, D.; Yamamoto, D.S.; Hashiguchi, Y. Prevalence of Genetically Complex Leishmania Strains with Hybrid and Mito-Nuclear Discordance. Front. Cell Infect. Microbiol. 2021, 11, 625001. [Google Scholar] [CrossRef] [PubMed]
- Okamoto, N.; Spurck, T.P.; Goodman, C.D.; McFadden, G.I. Apicoplast and mitochondrion in gametocytogenesis of Plasmodium falciparum. Eukaryot. Cell 2009, 8, 128–132. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Negreira, G.H.; Monsieurs, P.; Imamura, H.; Maes, I.; Kuk, N.; Yagoubat, A.; Van den Broeck, F.; Sterkers, Y.; Dujardin, J.C.; Domagalska, M.A. High throughput single-cell genome sequencing gives insights into the generation and evolution of mosaic aneuploidy in Leishmania donovani. Nucleic Acids Res. 2022, 50, 293–305. [Google Scholar] [CrossRef]
- Charyyeva, A.; Cetinkaya, U.; Ozkan, B.; Sahin, S.; Yaprak, N.; Sahin, I.; Yurchenko, V.; Kostygov, A.Y. Genetic diversity of Leishmania tropica: Unexpectedly complex distribution pattern. Acta Trop. 2021, 218, 105888. [Google Scholar] [CrossRef] [PubMed]
- Iantorno, S.A.; Durrant, C.; Khan, A.; Sanders, M.J.; Beverley, S.M.; Warren, W.C.; Berriman, M.; Sacks, D.L.; Cotton, J.A.; Grigg, M.E. Gene Expression in Leishmania Is Regulated Predominantly by Gene Dosage. MBio 2017, 8, e01393-17. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lypaczewski, P.; Zhang, W.W.; Matlashewski, G. Evidence that a naturally occurring single nucleotide polymorphism in the RagC gene of Leishmania donovani contributes to reduced virulence. PLoS Negl. Trop. Dis. 2021, 15, e0009079. [Google Scholar] [CrossRef]
- Salloum, T.; Moussa, R.; Rahy, R.; Al Deek, J.; Khalifeh, I.; El Hajj, R.; Hall, N.; Hirt, R.P.; Tokajian, S. Expanded genome-wide comparisons give novel insights into population structure and genetic heterogeneity of Leishmania tropica complex. PLoS Negl. Trop. Dis. 2020, 14, e0008684. [Google Scholar] [CrossRef]
- Bernstein, C.; Johns, V. Sexual reproduction as a response to H2O2 damage in Schizosaccharomyces pombe. J. Bacteriol. 1989, 171, 1893–1897. [Google Scholar] [CrossRef] [Green Version]
- Ram, Y.; Hadany, L. Condition-dependent sex: Who does it, when and why? Philos. Trans. R. Soc. B Biol. Sci. 2016, 371, 20150539. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schoustra, S.; Rundle, H.D.; Dali, R.; Kassen, R. Fitness-associated sexual reproduction in a filamentous fungus. Curr. Biol. 2010, 20, 1350–1355. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alves, C.L.; Repoles, B.M.; da Silva, M.S.; Mendes, I.C.; Marin, P.A.; Aguiar, P.H.N.; Santos, S.D.S.; Franco, G.R.; Macedo, A.M.; Pena, S.D.J.; et al. The recombinase Rad51 plays a key role in events of genetic exchange in Trypanosoma cruzi. Sci. Rep. 2018, 8, 13335. [Google Scholar] [CrossRef] [PubMed]
- Graca-Souza, A.V.; Maya-Monteiro, C.; Paiva-Silva, G.O.; Braz, G.R.; Paes, M.C.; Sorgine, M.H.; Oliveira, M.F.; Oliveira, P.L. Adaptations against heme toxicity in blood-feeding arthropods. Insect Biochem. Mol. Biol. 2006, 36, 322–335. [Google Scholar] [CrossRef]
- Pruzinova, K.; Sadlova, J.; Myskova, J.; Lestinova, T.; Janda, J.; Volf, P. Leishmania mortality in sand fly blood meal is not species-specific and does not result from direct effect of proteinases. Parasites Vectors 2018, 11, 37. [Google Scholar] [CrossRef] [Green Version]
- da Silva, M.S. DNA Double-Strand Breaks: A Double-Edged Sword for Trypanosomatids. Front. Cell Dev. Biol. 2021, 9, 669041. [Google Scholar] [CrossRef]
- de Massy, B. Initiation of meiotic recombination: How and where? Conservation and specificities among eukaryotes. Annu. Rev. Genet. 2013, 47, 563–599. [Google Scholar] [CrossRef]
- Yadav, V.K.; Claeys Bouuaert, C. Mechanism and Control of Meiotic DNA Double-Strand Break Formation in S. cerevisiae. Front. Cell Dev. Biol. 2021, 9, 642737. [Google Scholar] [CrossRef]
- Inbar, E.; Hughitt, V.K.; Dillon, L.A.; Ghosh, K.; El-Sayed, N.M.; Sacks, D.L. The Transcriptome of Leishmania major Developmental Stages in Their Natural Sand Fly Vector. MBio 2017, 8, e00029-17. [Google Scholar] [CrossRef] [Green Version]
- Ramesh, M.A.; Malik, S.B.; Logsdon, J.M., Jr. A phylogenomic inventory of meiotic genes; evidence for sex in Giardia and an early eukaryotic origin of meiosis. Curr. Biol. 2005, 15, 185–191. [Google Scholar] [CrossRef] [Green Version]
- Tromer, E.C.; Wemyss, T.A.; Ludzia, P.; Waller, R.F.; Akiyoshi, B. Repurposing of synaptonemal complex proteins for kinetochores in Kinetoplastida. Open Biol. 2021, 11, 210049. [Google Scholar] [CrossRef] [PubMed]
- Peacock, L.; Ferris, V.; Sharma, R.; Sunter, J.; Bailey, M.; Carrington, M.; Gibson, W. Identification of the meiotic life cycle stage of Trypanosoma brucei in the tsetse fly. Proc. Natl. Acad. Sci. USA 2011, 108, 3671–3676. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dernburg, A.F.; McDonald, K.; Moulder, G.; Barstead, R.; Dresser, M.; Villeneuve, A.M. Meiotic recombination in C. elegans initiates by a conserved mechanism and is dispensable for homologous chromosome synapsis. Cell 1998, 94, 387–398. [Google Scholar] [CrossRef] [Green Version]
- Thorne, L.W.; Byers, B. Stage-specific effects of X-irradiation on yeast meiosis. Genetics 1993, 134, 29–42. [Google Scholar] [CrossRef]
- Bloomfield, G. Spo11-Independent Meiosis in Social Amoebae. Annu. Rev. Microbiol. 2018, 72, 293–307. [Google Scholar] [CrossRef]
- Liu, Y.; Tewari, R.; Ning, J.; Blagborough, A.M.; Garbom, S.; Pei, J.; Grishin, N.V.; Steele, R.E.; Sinden, R.E.; Snell, W.J.; et al. The conserved plant sterility gene HAP2 functions after attachment of fusogenic membranes in Chlamydomonas and Plasmodium gametes. Genes Dev. 2008, 22, 1051–1068. [Google Scholar] [CrossRef] [Green Version]
- Peacock, L.; Kay, C.; Farren, C.; Bailey, M.; Carrington, M.; Gibson, W. Sequential production of gametes during meiosis in trypanosomes. Commun. Biol. 2021, 4, 555. [Google Scholar] [CrossRef]
- Fedry, J.; Liu, Y.; Pehau-Arnaudet, G.; Pei, J.; Li, W.; Tortorici, M.A.; Traincard, F.; Meola, A.; Bricogne, G.; Grishin, N.V.; et al. The Ancient Gamete Fusogen HAP2 Is a Eukaryotic Class II Fusion Protein. Cell 2017, 168, 904–915.e10. [Google Scholar] [CrossRef] [Green Version]
- Beh, C.T.; Brizzio, V.; Rose, M.D. KAR5 encodes a novel pheromone-inducible protein required for homotypic nuclear fusion. J. Cell Biol. 1997, 139, 1063–1076. [Google Scholar] [CrossRef] [Green Version]
- Neva, F.A.; Wyler, D.; Nash, T. Cutaneous leishmaniasis—A case with persistent organisms after treatment in presence of normal immune response. Am. J. Trop. Med. Hyg. 1979, 28, 467–471. [Google Scholar] [CrossRef]
- Lee, S.H.; Sacks, D.L. Tissue-resident macrophages as replicative niches for intracellular pathogens. Emerg. Top. Life Sci. 2017, 1, 621–626. [Google Scholar] [CrossRef] [PubMed]
- Kariyawasam, K.; Selvapandiyan, A.; Siriwardana, H.; Dube, A.; Karunanayake, P.; Senanayake, S.; Dey, R.; Gannavaram, S.; Nakhasi, H.L.; Karunaweera, N.D. Dermotropic Leishmania donovani in Sri Lanka: Visceralizing potential in clinical and preclinical studies. Parasitology 2018, 145, 443–452. [Google Scholar] [CrossRef] [PubMed] [Green Version]
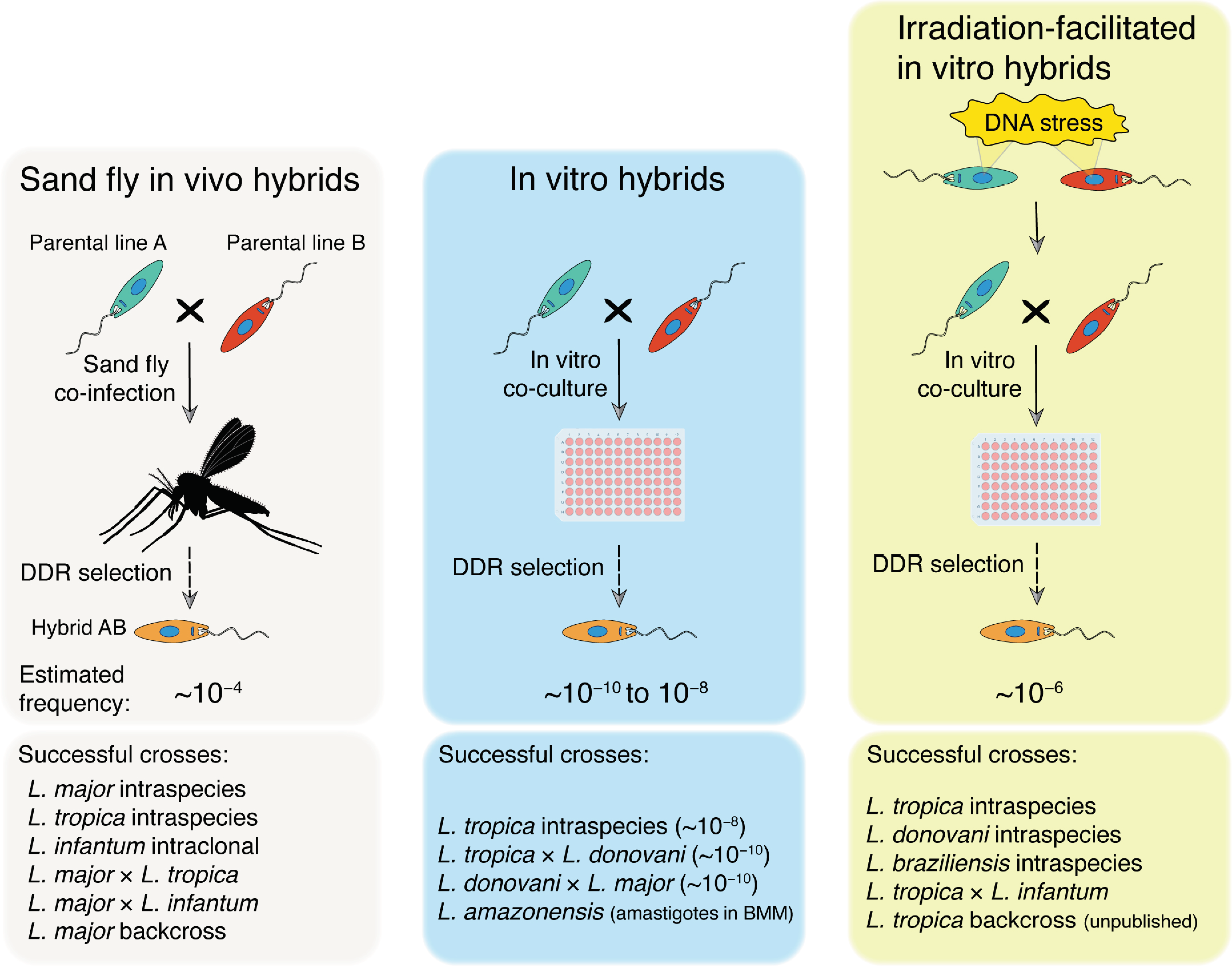
Figure 1.
Experimental Leishmania hybridization. Three major methodologies for generation of Leishmania hybrids have been described using two drug-resistant parental promastigote cell lines: (1) sand fly co-infection [9,10,11,12,13] (left panel); (2) in vitro co-culture [15,17] (middle panel); and (3) in vitro co-culture of gamma-radiation treated promastigotes [16] (right panel). DNA stress may be induced by H2O2 or methyl methanosulfonate (MMS) treatment to increase mating competency. In all cases, hybrids were selected by double drug resistance (DDR) in culture. Estimated average minimum frequencies of mating-competent cells are shown for each method. Successful experimental crosses described using each method are listed in the bottom part of each panel. BMM: mouse bone marrow-derived macrophages.
Figure 1.
Experimental Leishmania hybridization. Three major methodologies for generation of Leishmania hybrids have been described using two drug-resistant parental promastigote cell lines: (1) sand fly co-infection [9,10,11,12,13] (left panel); (2) in vitro co-culture [15,17] (middle panel); and (3) in vitro co-culture of gamma-radiation treated promastigotes [16] (right panel). DNA stress may be induced by H2O2 or methyl methanosulfonate (MMS) treatment to increase mating competency. In all cases, hybrids were selected by double drug resistance (DDR) in culture. Estimated average minimum frequencies of mating-competent cells are shown for each method. Successful experimental crosses described using each method are listed in the bottom part of each panel. BMM: mouse bone marrow-derived macrophages.


{kind=link}
Table 1.
Leishmania crosses and disease models in mice currently available for linkage studies.
| Leishmania Cross | Parental Disease Phenotype in Humans | Parental Disease Phenotype in Mouse Models |
|---|---|---|
| L. major Fn × L. major Sd [11] | Healing cutaneous lesion Chronic cutaneous lesion [60] | C57Bl/6: healing ear dermal lesion [61] C57Bl/6: non-healing dermal lesion [61] |
| L. major Fn × L. infantum LLM-320 [13] | Healing cutaneous lesion Visceral leishmaniasis | C57Bl/6: healing ear dermal lesion; poor dissemination to or growth in the spleen [13] C57Bl/6: no dermal pathology; good dissemination to and growth in the spleen [13] |
| L. major Fn × L. tropica L747 (unpublished) | Healing cutaneous lesion Chronic cutaneous lesion | BALB/c: ear dermal lesion BALB/c: no dermal pathology (unpublished) |
| L. donovani SL2706 × L. donovani Mongi [16] | Cutaneous lesion [62] Visceral leishmaniasis | BALB/c: ear dermal lesion; no dissemination to or growth in the spleen [62] BALB/c: no dermal pathology; good dissemination to and growth in the spleen |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Ferreira, T.R.; Sacks, D.L. Experimental Hybridization in Leishmania: Tools for the Study of Genetic Exchange. Pathogens 2022, 11, 580. https://0-doi-org.brum.beds.ac.uk/10.3390/pathogens11050580
AMA Style
Ferreira TR, Sacks DL. Experimental Hybridization in Leishmania: Tools for the Study of Genetic Exchange. Pathogens. 2022; 11(5):580. https://0-doi-org.brum.beds.ac.uk/10.3390/pathogens11050580
Chicago/Turabian StyleFerreira, Tiago R., and David L. Sacks. 2022. "Experimental Hybridization in Leishmania: Tools for the Study of Genetic Exchange" Pathogens 11, no. 5: 580. https://0-doi-org.brum.beds.ac.uk/10.3390/pathogens11050580
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.