
Comparative Whole Genome Analysis of an Anaplasma phagocytophilum Strain Isolated from Norwegian Sheep

 , ,
, ,
Abstract
:1. Introduction
2. Results and Discussion
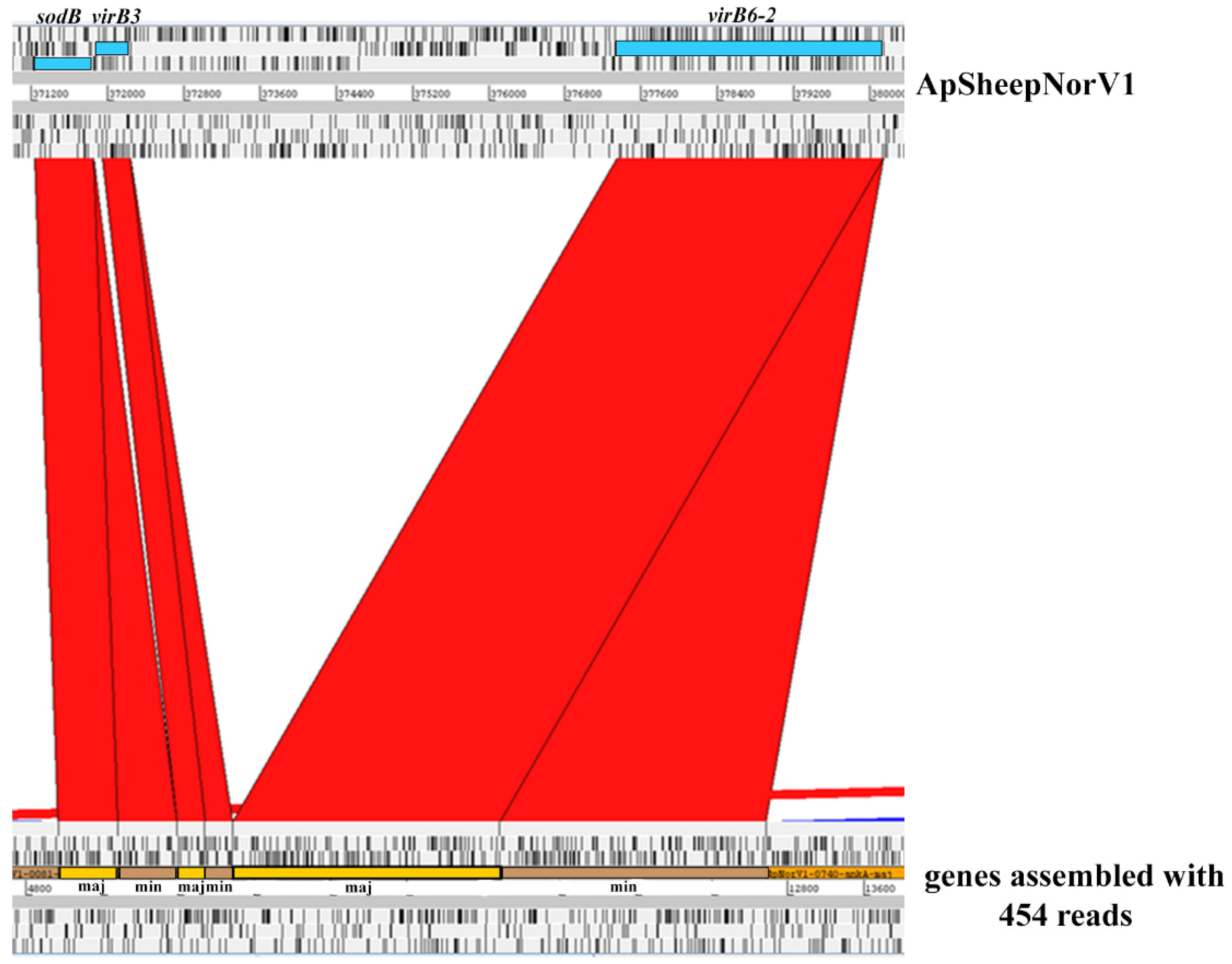
2.1. Characterization of the ApSheep_NorV1 Strain of A. phagocytophilum
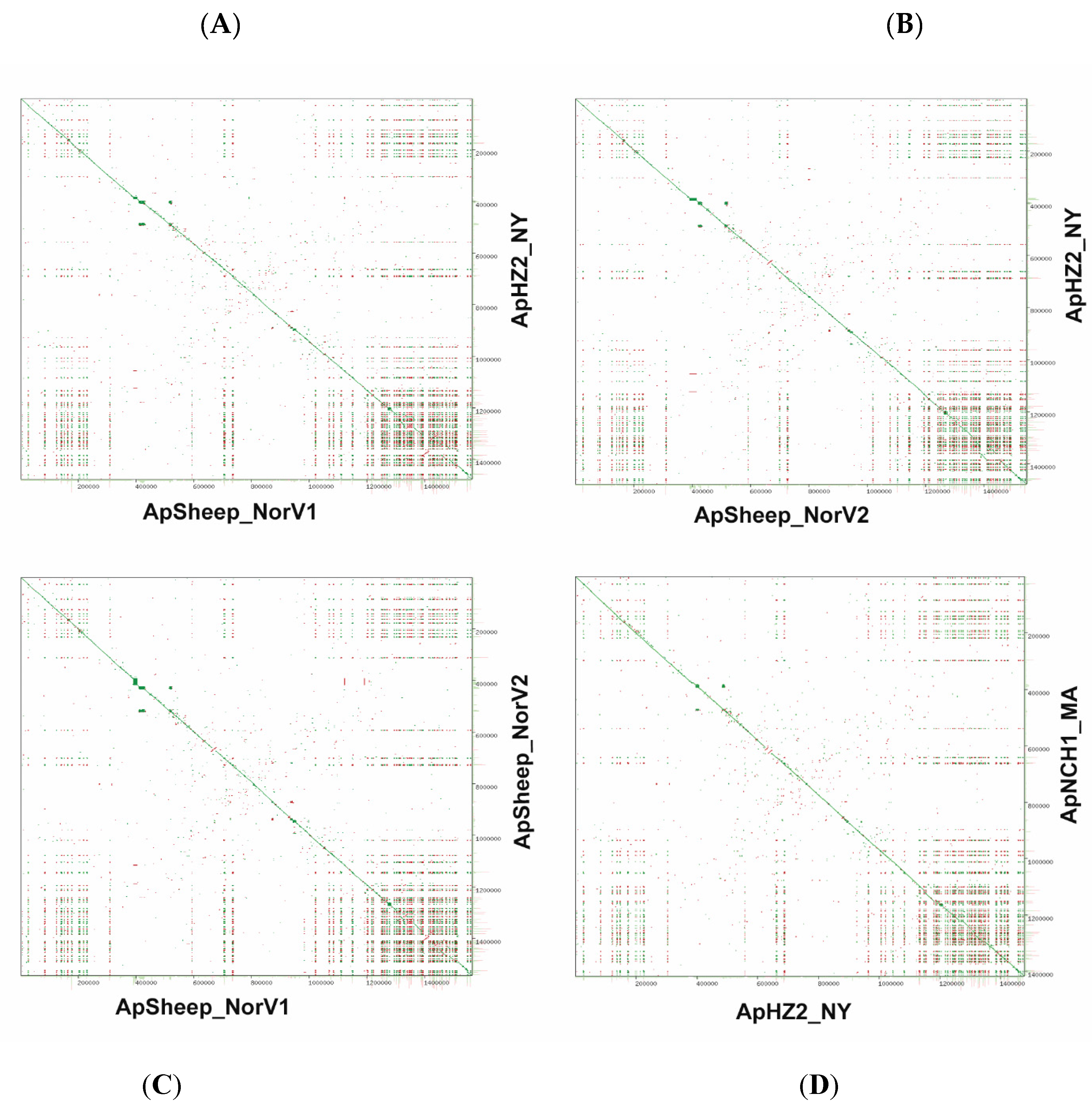
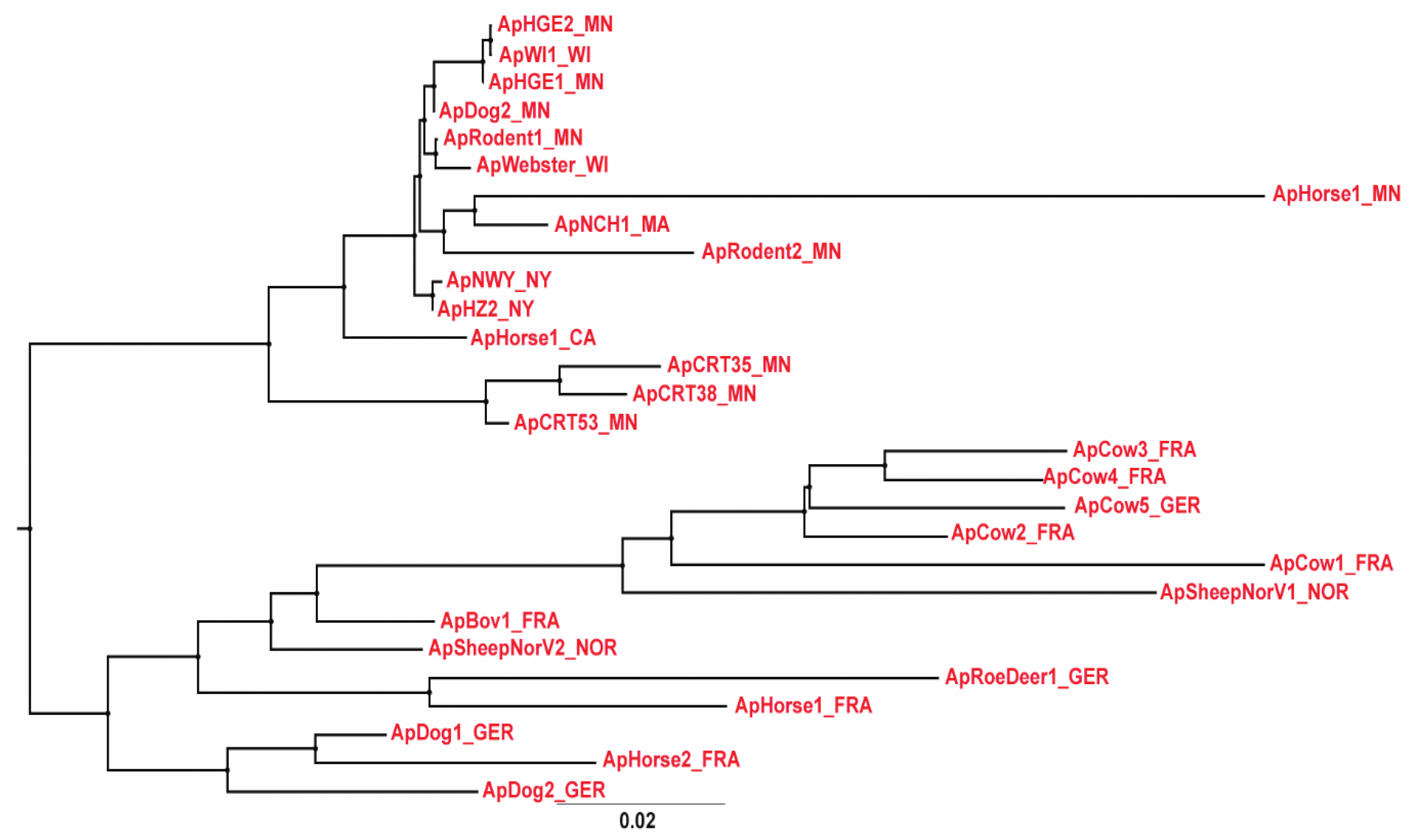
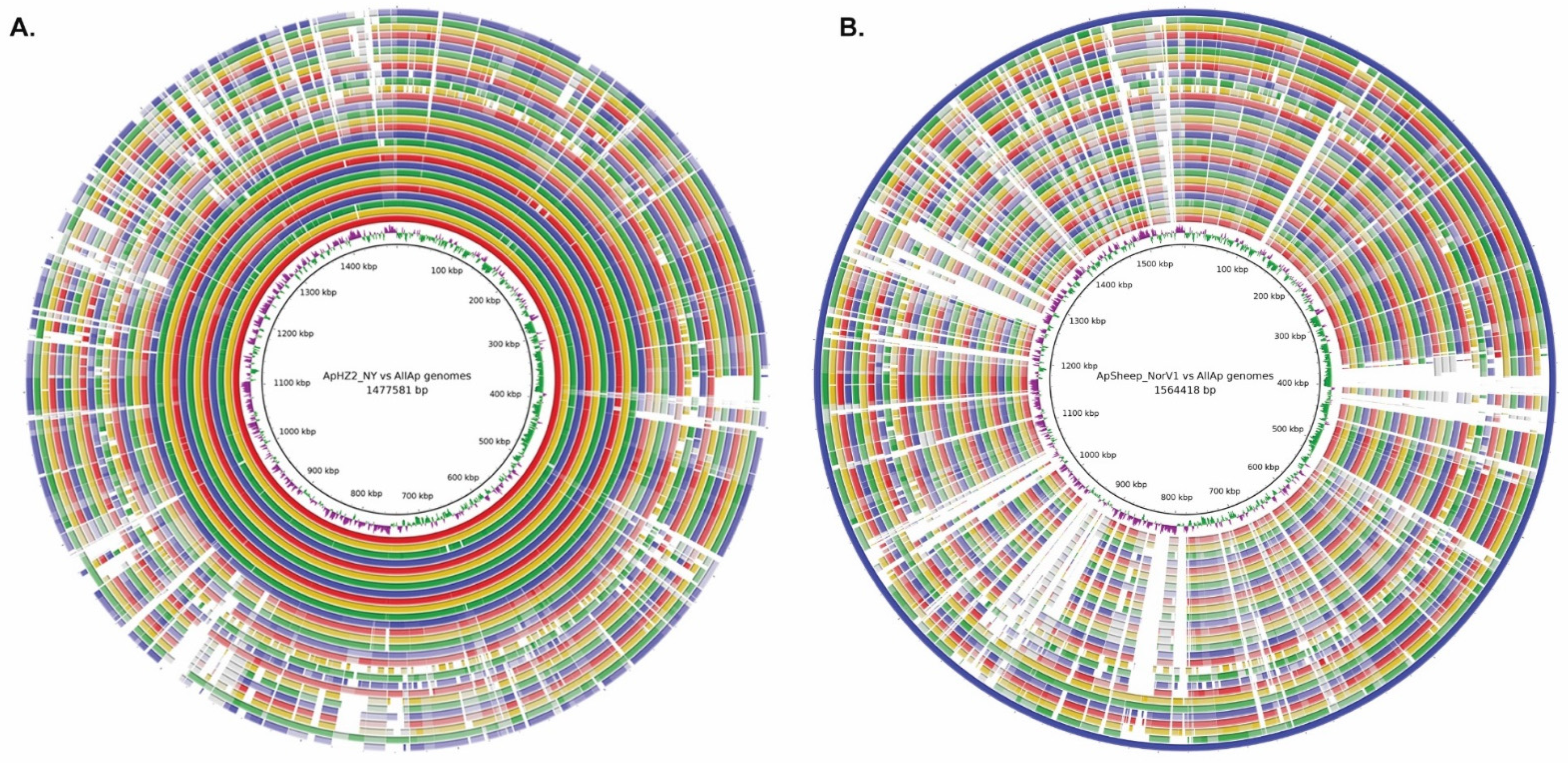
2.2. Comparative Genome Analysis
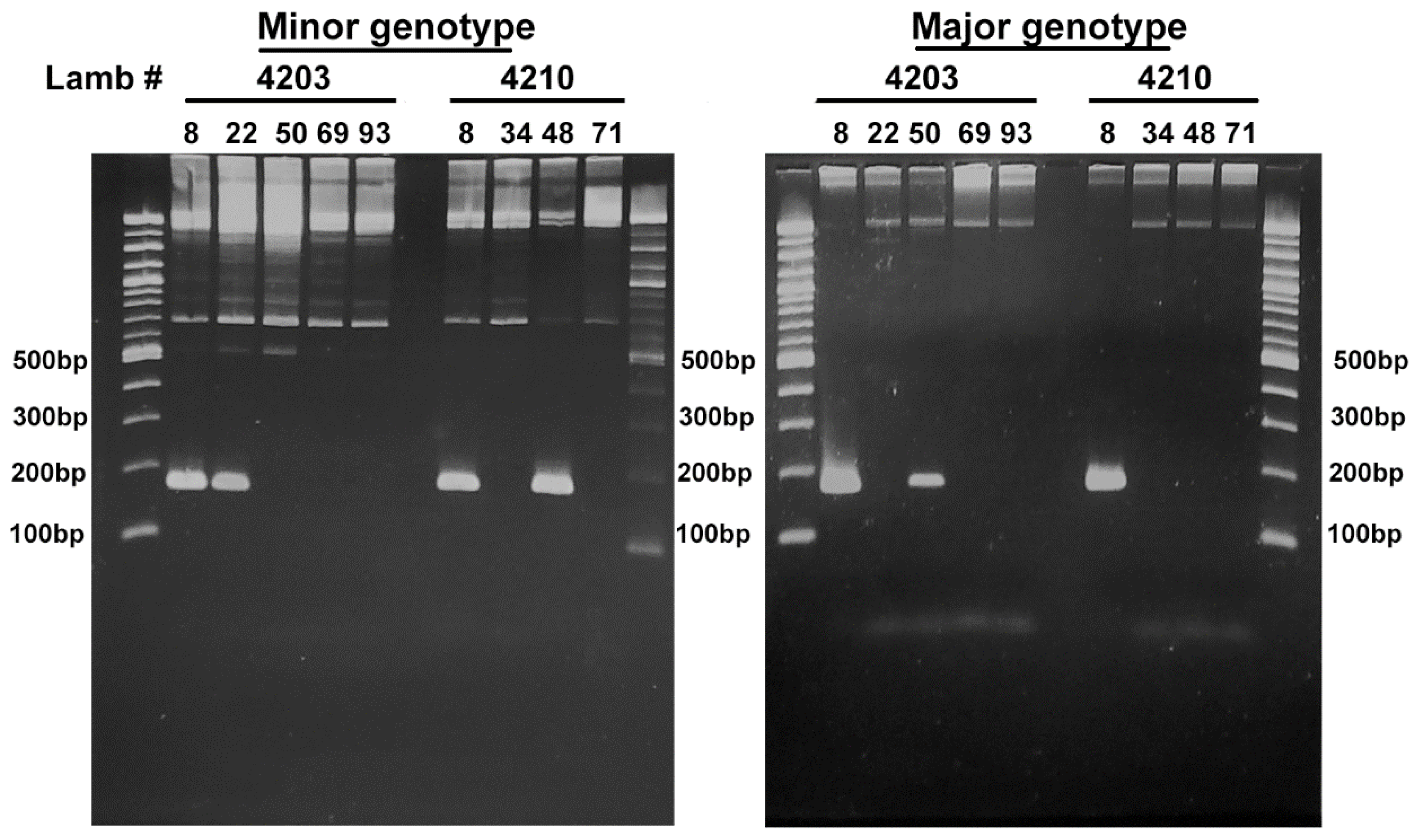
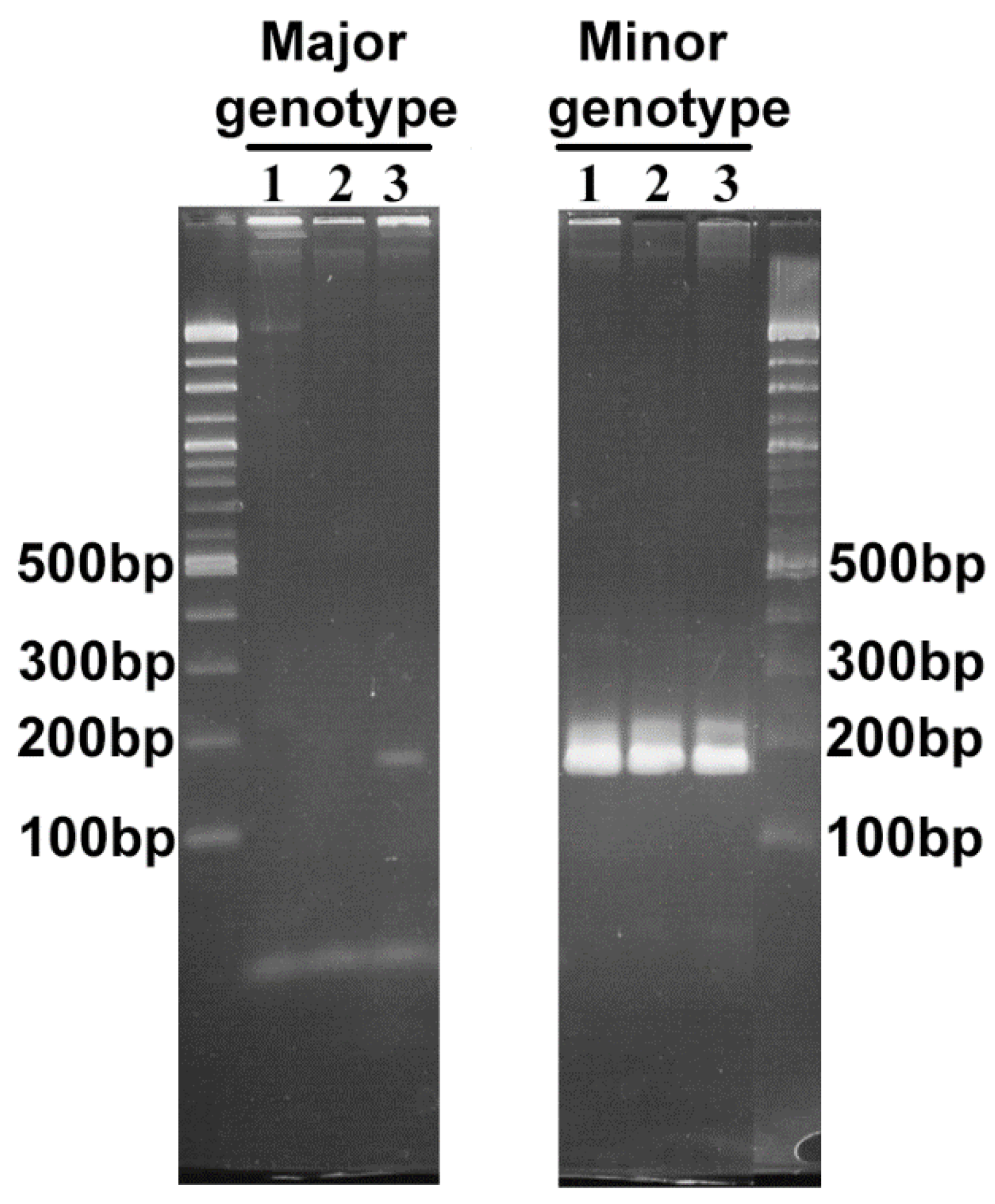
2.3. Comparison of the msp2/p44 Repertoires
2.4. Tick Host Cell Genome
3. Conclusions
4. Materials and Methods
4.1. Anaplasma Phagocytophilum Genome Sequences
4.2. Maintenance of Uninfected Tick Cell Lines and Cultivation of ApNorV1 Mutants
4.3. Preparation of A. phagocytophilum from Infected Sheep
4.4. DNA Extraction and PCR Analysis
4.5. Determination of msp2/p44 Repertoires in Each Strain Genome of A. phagocytophilum
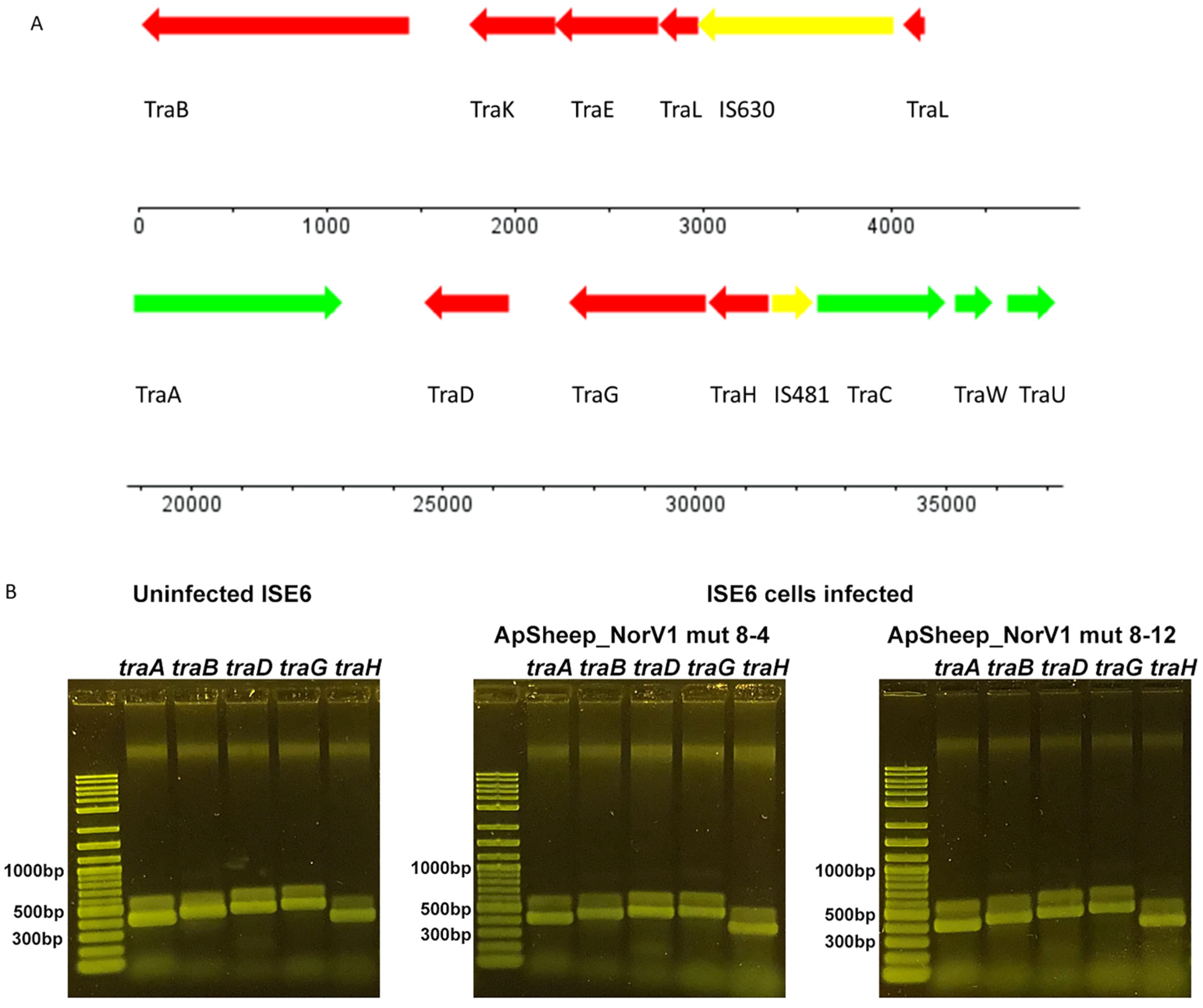
4.6. Identification of Symbiotic Rickettsia tra Genes in ISE6 Cells
4.7. Pan Genome Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Stuen, S.; Bergstrom, K.; Palmer, E. Reduced weight gain due to subclinical Anaplasma phagocytophilum (formerly Ehrlichia phagocytophila) infection. Exp. Appl. Acarol. 2002, 28, 209–215. [Google Scholar] [CrossRef] [PubMed]
- Eskeland, S.; Stuen, S.; Munderloh, U.G.; Barbet, A.; Crosby, L.; Lybeck, K.; Wilhelmsson, P.; Lindgren, P.E.; Makvandi-Nejad, S.; Tollefsen, S.; et al. Clinical and immunological responses in sheep after inoculation with Himar1-transformed Anaplasma phagocytophilum and subsequent challenge with a virulent strain of the bacterium. Vet. Immunol. Immunopathol. 2021, 231, 110165. [Google Scholar] [CrossRef] [PubMed]
- Woldehiwet, Z.; Horrocks, B.K.; Scaife, H.; Ross, G.; Munderloh, U.G.; Bown, K.; Edwards, S.W.; Hart, C.A. Cultivation of an ovine strain of Ehrlichia phagocytophila in tick cell cultures. J. Comp. Pathol. 2002, 127, 142–149. [Google Scholar] [CrossRef] [PubMed]
- Massung, R.F.; Levin, M.L.; Munderloh, U.G.; Silverman, D.J.; Lynch, M.J.; Gaywee, J.K.; Kurtti, T.J. Isolation and propagation of the Ap-Variant 1 strain of Anaplasma phagocytophilum in a tick cell line. J. Clin. Microbiol. 2007, 45, 2138–2143. [Google Scholar] [CrossRef] [Green Version]
- Almazan, C.; Fourniol, L.; Rouxel, C.; Alberdi, P.; Gandoin, C.; Lagree, A.C.; Boulouis, H.J.; de la Fuente, J.; Bonnet, S.I. Experimental Ixodes ricinus-Sheep Cycle of Anaplasma phagocytophilum NV2Os Propagated in Tick Cell Cultures. Front. Vet. Sci. 2020, 7, 40. [Google Scholar] [CrossRef] [Green Version]
- Stuen, S.; Bergstrom, K.; Petrovec, M.; Van de Pol, I.; Schouls, L.M. Differences in clinical manifestations and hematological and serological responses after experimental infection with genetic variants of Anaplasma phagocytophilum in sheep. Clin. Diagn. Lab. Immunol. 2003, 10, 692–695. [Google Scholar] [CrossRef] [Green Version]
- Massung, R.F.; Mauel, M.J.; Owens, J.H.; Allan, N.; Courtney, J.W.; Stafford, K.C., III; Mather, T.N. Genetic variants of Ehrlichia phagocytophila, Rhode Island and Connecticut. Emerg. Infect. Dis. 2002, 8, 467–472. [Google Scholar] [CrossRef] [Green Version]
- Dugat, T.; Lagree, A.C.; Maillard, R.; Boulouis, H.J.; Haddad, N. Opening the black box of Anaplasma phagocytophilum diversity: Current situation and future perspectives. Front. Cell Infect. Microbiol. 2015, 5, 61. [Google Scholar] [CrossRef]
- Stuen, S.; Van De Pol, I.; Bergstrom, K.; Schouls, L.M. Identification of Anaplasma phagocytophila (formerly Ehrlichia phagocytophila) variants in blood from sheep in Norway. J. Clin. Microbiol. 2002, 40, 3192–3197. [Google Scholar] [CrossRef] [Green Version]
- Granquist, E.G.; Stuen, S.; Lundgren, A.M.; Braten, M.; Barbet, A.F. Outer membrane protein sequence variation in lambs experimentally infected with Anaplasma phagocytophilum. Infect. Immun. 2008, 76, 120–126. [Google Scholar] [CrossRef] [Green Version]
- Granquist, E.G.; Stuen, S.; Crosby, L.; Lundgren, A.M.; Alleman, A.R.; Barbet, A.F. Variant-specific and diminishing immune responses towards the highly variable MSP2(P44) outer membrane protein of Anaplasma phagocytophilum during persistent infection in lambs. Vet. Immunol. Immunopathol. 2010, 133, 117–124. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stuen, S.; Torsteinbo, W.O.; Bergstrom, K.; Bardsen, K. Superinfection occurs in Anaplasma phagocytophilum infected sheep irrespective of infection phase and protection status. Acta Vet. Scand. 2009, 51, 41. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stuen, S.; Granquist, E.G.; Silaghi, C. Anaplasma phagocytophilum—A widespread multi-host pathogen with highly adaptive strategies. Front. Cell Infect. Microbiol. 2013, 3, 31. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ladbury, G.A.; Stuen, S.; Thomas, R.; Bown, K.J.; Woldehiwet, Z.; Granquist, E.G.; Bergstrom, K.; Birtles, R.J. Dynamic transmission of numerous Anaplasma phagocytophilum genotypes among lambs in an infected sheep flock in an area of anaplasmosis endemicity. J. Clin. Microbiol. 2008, 46, 1686–1691. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Barbet, A.F.; Meeus, P.F.; Belanger, M.; Bowie, M.V.; Yi, J.; Lundgren, A.M.; Alleman, A.R.; Wong, S.J.; Chu, F.K.; Munderloh, U.G.; et al. Expression of multiple outer membrane protein sequence variants from a single genomic locus of Anaplasma phagocytophilum. Infect. Immun. 2003, 71, 1706–1718. [Google Scholar] [CrossRef] [Green Version]
- Noe, L.; Kucherov, G. YASS. Enhancing the sensitivity of DNA similarity search. Nucleic Acids Res. 2005, 33, W540–W543. [Google Scholar] [CrossRef] [Green Version]
- Foley, J.E.; Nieto, N.C.; Barbet, A.; Foley, P. Antigen diversity in the parasitic bacterium Anaplasma phagocytophilum arises from selectively-represented, spatially clustered functional pseudogenes. PLoS ONE 2009, 4, e8265. [Google Scholar] [CrossRef]
- Richter, M.; Rossello-Mora, R. Shifting the genomic gold standard for the prokaryotic species definition. Proc. Natl. Acad. Sci. USA 2009, 106, 19126–19131. [Google Scholar] [CrossRef] [Green Version]
- Richter, M.; Rossello-Mora, R.; Oliver Glockner, F.; Peplies, J. JSpeciesWS: A web server for prokaryotic species circumscription based on pairwise genome comparison. Bioinformatics 2016, 32, 929–931. [Google Scholar] [CrossRef]
- Alikhan, N.F.; Petty, N.K.; Ben Zakour, N.L.; Beatson, S.A. BLAST Ring Image Generator (BRIG): Simple prokaryote genome comparisons. BMC Genom. 2011, 12, 402. [Google Scholar] [CrossRef] [Green Version]
- Tettelin, H.; Masignani, V.; Cieslewicz, M.J.; Donati, C.; Medini, D.; Ward, N.L.; Angiuoli, S.V.; Crabtree, J.; Jones, A.L.; Durkin, A.S.; et al. Genome analysis of multiple pathogenic isolates of Streptococcus agalactiae: Implications for the microbial “pan-genome”. Proc. Natl. Acad. Sci. USA 2005, 102, 13950–13955. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Barbet, A.F.; Lundgren, A.M.; Alleman, A.R.; Stuen, S.; Bjoersdorff, A.; Brown, R.N.; Drazenovich, N.L.; Foley, J.E. Structure of the expression site reveals global diversity in MSP2 (P44) variants in Anaplasma phagocytophilum. Infect. Immun. 2006, 74, 6429–6437. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Barbet, A.F.; Al-Khedery, B.; Stuen, S.; Granquist, E.G.; Felsheim, R.F.; Munderloh, U.G. An emerging tick-borne disease of humans is caused by a subset of strains with conserved genome structure. Pathogens 2013, 2, 544–555. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Drown, D.M.; Zee, P.C.; Brandvain, Y.; Wade, M.J. Evolution of transmission mode in obligate symbionts. Evol. Ecol. Res. 2013, 15, 43–59. [Google Scholar] [PubMed]
- Dunning Hotopp, J.C.; Lin, M.; Madupu, R.; Crabtree, J.; Angiuoli, S.V.; Eisen, J.A.; Seshadri, R.; Ren, Q.; Wu, M.; Utterback, T.R.; et al. Comparative genomics of emerging human ehrlichiosis agents. PLoS Genet. 2006, 2, e21. [Google Scholar]
- Miller, J.R.; Koren, S.; Dilley, K.A.; Harkins, D.M.; Stockwell, T.B.; Shabman, R.S.; Sutton, G.G. A draft genome sequence for the Ixodes scapularis cell line, ISE6. F1000Research 2018, 7, 297. [Google Scholar] [CrossRef] [Green Version]
- Kurtti, T.J.; Felsheim, R.F.; Burkhardt, N.Y.; Oliver, J.D.; Heu, C.C.; Munderloh, U.G. Rickettsia buchneri sp. nov., a rickettsial endosymbiont of the blacklegged tick Ixodes scapularis. Int. J. Syst. Evol. Microbiol. 2015, 65, 965–970. [Google Scholar] [CrossRef]
- Hagen, R.; Verhoeve, V.I.; Gillespie, J.J.; Driscoll, T.P. Conjugative transposons and their cargo genes vary across natural populations of Rickettsia buchneri Infecting the tick Ixodes scapularis. Genome. Biol. Evol. 2018, 10, 3218–3229. [Google Scholar] [CrossRef] [Green Version]
- Mazzacano, C.A.; Munderloh, U.G.; Kurtti, T.J. Characterization of a new continuous cell line from the flood water mosquito, Aedes vexans. Cytotechnology 1991, 5, 147–154. [Google Scholar] [CrossRef]
- Drezen, J.M.; Gauthier, J.; Josse, T.; Bezier, A.; Herniou, E.; Huguet, E. Foreign DNA acquisition by invertebrate genomes. J. Invertebr. Pathol. 2017, 147, 157–168. [Google Scholar] [CrossRef]
- Dunning Hotopp, J.C.; Clark, M.E.; Oliveira, D.C.; Foster, J.M.; Fischer, P.; Munoz Torres, M.C.; Giebel, J.D.; Kumar, N.; Ishmael, N.; Wang, S.; et al. Widespread lateral gene transfer from intracellular bacteria to multicellular eukaryotes. Science 2007, 317, 1753–1756. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Weinert, L.A.; Welch, J.J.; Jiggins, F.M. Conjugation gen.nes are common throughout the genus Rickettsia and are transmitted horizontally. Proc. Biol. Sci. 2009, 276, 3619–3627. [Google Scholar] [PubMed] [Green Version]
- Black, W.C.; Piesman, J. Phylogeny of hard-and soft-tick taxa (Acari: Ixodida) based on mitochondrial 16S rDNA sequences. Proc. Natl. Acad. Sci. USA 1994, 91, 10034–10038. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pal, U.; Dai, J.; Li, X.; Neelakanta, G.; Luo, P.; Kumar, M.; Wang, P.; Yang, X.; Anderson, J.F.; Fikrig, E. A differential role for BB0365 in the persistence of Borrelia burgdorferi in mice and ticks. J. Infect. Dis. 2008, 197, 148–155. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Noda, H.; Munderloh, U.G.; Kurtti, T.J. Endosymbionts of ticks and their relationship to Wolbachia spp. and tick-borne pathogens of humans and animals. Appl. Environ. Microbiol. 1997, 63, 3926–3932. [Google Scholar] [CrossRef] [Green Version]
- Munderloh, U.G.; Blouin, E.F.; Kocan, K.M.; Ge, N.L.; Edwards, W.L.; Kurtti, T.J. Establishment of the tick (Acari: Ixodidae)-borne cattle pathogen Anaplasma marginale (Rickettsiales:Anaplasmataceae) in tick cell culture. J. Med. Entomol. 1996, 33, 656–664. [Google Scholar] [CrossRef]
- Simser, J.A.; Palmer, A.T.; Fingerle, V.; Wilske, B.; Kurtti, T.J.; Munderloh, U.G. Rickettsia monacensis sp. nov., a spotted fever group Rickettsia, from ticks (Ixodes ricinus) collected in a European city park. Appl. Environ. Microbiol. 2002, 68, 4559–4566. [Google Scholar] [CrossRef] [Green Version]
- Singu, V.; Peddireddi, L.; Sirigireddy, K.R.; Cheng, C.; Munderloh, U.; Ganta, R.R. Unique macrophage and tick cell-specific protein expression from the p28/p30-outer membrane protein multigene locus in Ehrlichia chaffeensis and Ehrlichia canis. Cell Microbiol. 2006, 8, 1475–1487. [Google Scholar] [CrossRef]
- Kurtti, T.J.; Munderloh, U.G.; Ahlstrand, G.G.; Johnson, R.C. Borrelia burgdorferi in tick cell culture: Growth and cellular adherence. J. Med. Entomol. 1988, 25, 256–261. [Google Scholar] [CrossRef]
- Munderloh, U.G.; Kurtti, T.J. Formulation of medium for tick cell culture. Exp. Appl. Acarol. 1989, 7, 219–229. [Google Scholar] [CrossRef]
- Munderloh, U.G.; Jauron, S.D.; Fingerle, V.; Leitritz, L.; Hayes, S.F.; Hautman, J.M.; Nelson, C.M.; Huberty, B.W.; Kurtti, T.J.; Ahlstrand, G.G.; et al. Invasion and intracellular development of the human granulocytic ehrlichiosis agent in tick cell culture. J. Clin. Microbiol. 1999, 37, 2518–2524. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Al-Khedery, B.; Lundgren, A.M.; Stuen, S.; Granquist, E.G.; Munderloh, U.G.; Nelson, C.M.; Alleman, A.R.; Mahan, S.M.; Barbet, A.F. Structure of the type IV secretion system in different strains of Anaplasma phagocytophilum. BMC Genom. 2012, 13, 678. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Carneiro, M.O.; Russ, C.; Ross, M.G.; Gabriel, S.B.; Nusbaum, C.; DePristo, M.A. Pacific biosciences sequencing technology for genotyping and variation discovery in human data. BMC Genom. 2012, 13, 375. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Koren, S.; Walenz, B.P.; Berlin, K.; Miller, J.R.; Bergman, N.H.; Phillippy, A.M. Canu: Scalable and accurate long-read assembly via adaptive k-mer weighting and repeat separation. Genome Res. 2017, 27, 722–736. [Google Scholar] [CrossRef] [Green Version]
- Bankevich, A.; Nurk, S.; Antipov, D.; Gurevich, A.A.; Dvorkin, M.; Kulikov, A.S.; Lesin, V.M.; Nikolenko, S.I.; Pham, S.; Prjibelski, A.D.; et al. SPAdes: A new genome assembly algorithm and its applications to single-cell sequencing. J. Comput. Biol. 2012, 19, 455–477. [Google Scholar] [CrossRef] [Green Version]
- Rissman, A.I. Reordering contigs of draft genomes using the Mauve Aligner. Bioinformatics 2009, 25, 2071–2073. [Google Scholar] [CrossRef]
- Seemann, T. Prokka: Rapid prokaryotic genome annotation. Bioinformatics 2014, 30, 2068–2069. [Google Scholar] [CrossRef]
- Page, A.J.; Cummins, C.A.; Hunt, M.; Wong, V.K.; Reuter, S.; Holden, M.T.; Fookes, M.; Falush, D.; Keane, J.A.; Parkhill, J. Roary: Rapid large-scale prokaryote pan genome analysis. Bioinformatics 2015, 31, 3691–3693. [Google Scholar] [CrossRef]
- Price, M.N.; Dehal, P.S.; Arkin, A.P. FastTree 2—Approximately maximum-likelihood trees for large alignments. PLoS ONE 2010, 5, e9490. [Google Scholar] [CrossRef]
- Page, A.J.; Cummins, C.A.; Hunt, M.; Wong, V.K.; Reuter, S.; Holden, M.T.; Fookes, M.; Falush, D.; Keane, J.A.; Parkhill, J. Phandango: An interactive viewer for bacterial population genomics. Bioinformatics 2018, 34, 292–293. [Google Scholar]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Conserved Flanking Sequences | Major Read Type/Total Reads | Minor Read Type/Total Reads |
|---|---|---|---|
| omp1X | CTGGACGTGTTCTCTGCT……GGGAAGTATAATCCTTCA | 31/36 | 5/36 |
| omp1X | GTAAAGCATAATATAGCG……CAGAGTATCAGCACTGAT | 30/32 | 2/32 |
| virB6-2 | CACATAAGTGGTGGTGGT……TTACTAAGAAAAGAGGAG | 17/22 | 5/22 |
| virB6-2 | ACACTTGTGCCTAAATCG……ATTGCAGGCAGTTACACT | 17/23 | 2//34 |
| Total | 17/24 | 14/124 | |
| Percentage | 17/25 | 11% |
| Tick Cell Line and Passage # (p.) | (a)Tick F/R | (b)17kDa | traA | traB | traD | traG | traH |
|---|---|---|---|---|---|---|---|
| ISE6 p.28 | + | − | + | + | + | + | + |
| ISE6 p. 133A | + | − | + | + | + | + | + |
| ISE6 p. 133B | + | − | + | + | + | + | + |
| ISE18 p. 43 | + | − | − | − | − | − | − |
| ISE24 p. 6 A | + | − | − | − | − | − | − |
| ISE24 p.6 B | + | − | − | − | − | − | − |
| IDE2 p. 61 | + | − | − | − | − | − | − |
| IDE8 p. 36 A,B | + | − | − | − | − | − | − |
| IRE11 p. 78 A,B | + | − | − | − | − | − | − |
| AAE2 p. 14 | + | − | − | − | − | − | − |
| BME26 p. 89 | + | − | − | − | − | − | − |
| AVE1 p. 9 | + | − | − | − | − | − | − |
| Primers Targeting A. phagocytophilum Variants | |||
|---|---|---|---|
| Sequence (5’ to 3’) | Amplicon size (bp) | Target | |
| AB1751 | GGTGCTCTTAAAAAACCTAGCGC | 169 | Major variant |
| AB1752 | CAGCCGCAAAGTTCTTTACTCTAT | ||
| AB1753 | GGGCTTTTAAAGCATAGCTT | 167 | Minor variant |
| AB1754 | TAGCTGCAGGATCCTTTACTCCCC | ||
| Primers using for Rickettsia-tra genes in ISE6 cells analysis | |||
| AB2111 | ATGAAGCAGGGATGGTAGGT | 415 | traA |
| AB2112 | AGTAACTCCCTGATGCCTTGA | ||
| AB2113 | TTCCAACCACCACACCAGTA | 450 | traB |
| AB2114 | AGGTCAAAACTTCCCGAGGT | ||
| AB2115 | AACGCAGCCACCATATTTCC | 473 | traD |
| AB2116 | AGTTGGTACTCGCTGAAGGA | ||
| AB2117 | TGGTGAGTACTTGCCCACAT | 491 | traG |
| AB2118 | GCCTTGCAGCAGATGAATTC | ||
| AB2119 | AATTGAGGCAGATTCTCCGC | 383 | traH |
| AB2120 | AGGGCGATTGTGAAATGACT | ||
| T1 forward | CCGGTCTGAACTCAGATCAAGT | 460 | Mitochondrial 16S rRNA [33] |
| T1 reverse | CTGCTCAATGATTTTTTAAATTGCTGTGG | ||
| T2 forward | GGTATCGTGCTCGACTC | 450 | Tick β-actin [34] |
| T2 reverse | ATCAGGTAGTCGGTCAGG | ||
| Rr17.61p | GCTCTTGCAACTTCTATGTT | 434 | 17kDa [35] |
| Rr17.492n | CATTGTTCGTCAGGTTGGCG | ||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Crosby, F.L.; Eskeland, S.; Bø-Granquist, E.G.; Munderloh, U.G.; Price, L.D.; Al-Khedery, B.; Stuen, S.; Barbet, A.F. Comparative Whole Genome Analysis of an Anaplasma phagocytophilum Strain Isolated from Norwegian Sheep. Pathogens 2022, 11, 601. https://0-doi-org.brum.beds.ac.uk/10.3390/pathogens11050601
Crosby FL, Eskeland S, Bø-Granquist EG, Munderloh UG, Price LD, Al-Khedery B, Stuen S, Barbet AF. Comparative Whole Genome Analysis of an Anaplasma phagocytophilum Strain Isolated from Norwegian Sheep. Pathogens. 2022; 11(5):601. https://0-doi-org.brum.beds.ac.uk/10.3390/pathogens11050601
Chicago/Turabian StyleCrosby, Francy L., Sveinung Eskeland, Erik G. Bø-Granquist, Ulrike G. Munderloh, Lisa D. Price, Basima Al-Khedery, Snorre Stuen, and Anthony F. Barbet. 2022. "Comparative Whole Genome Analysis of an Anaplasma phagocytophilum Strain Isolated from Norwegian Sheep" Pathogens 11, no. 5: 601. https://0-doi-org.brum.beds.ac.uk/10.3390/pathogens11050601