Metagenomic Characterization of Bacterial Communities on Ready-to-Eat Vegetables and Effects of Household Washing on their Diversity and Composition




Abstract
:1. Introduction
2. Results and Discussion
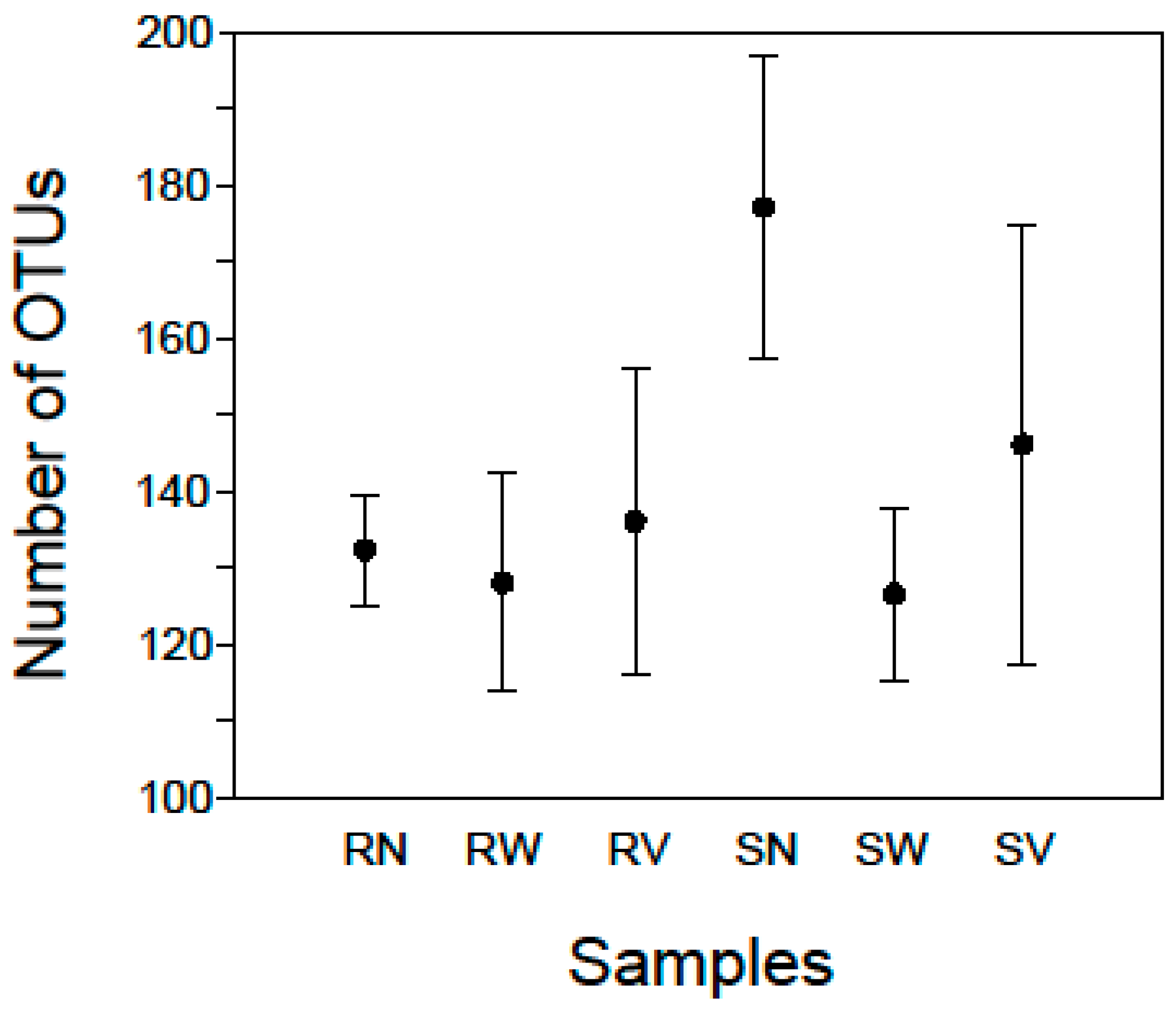
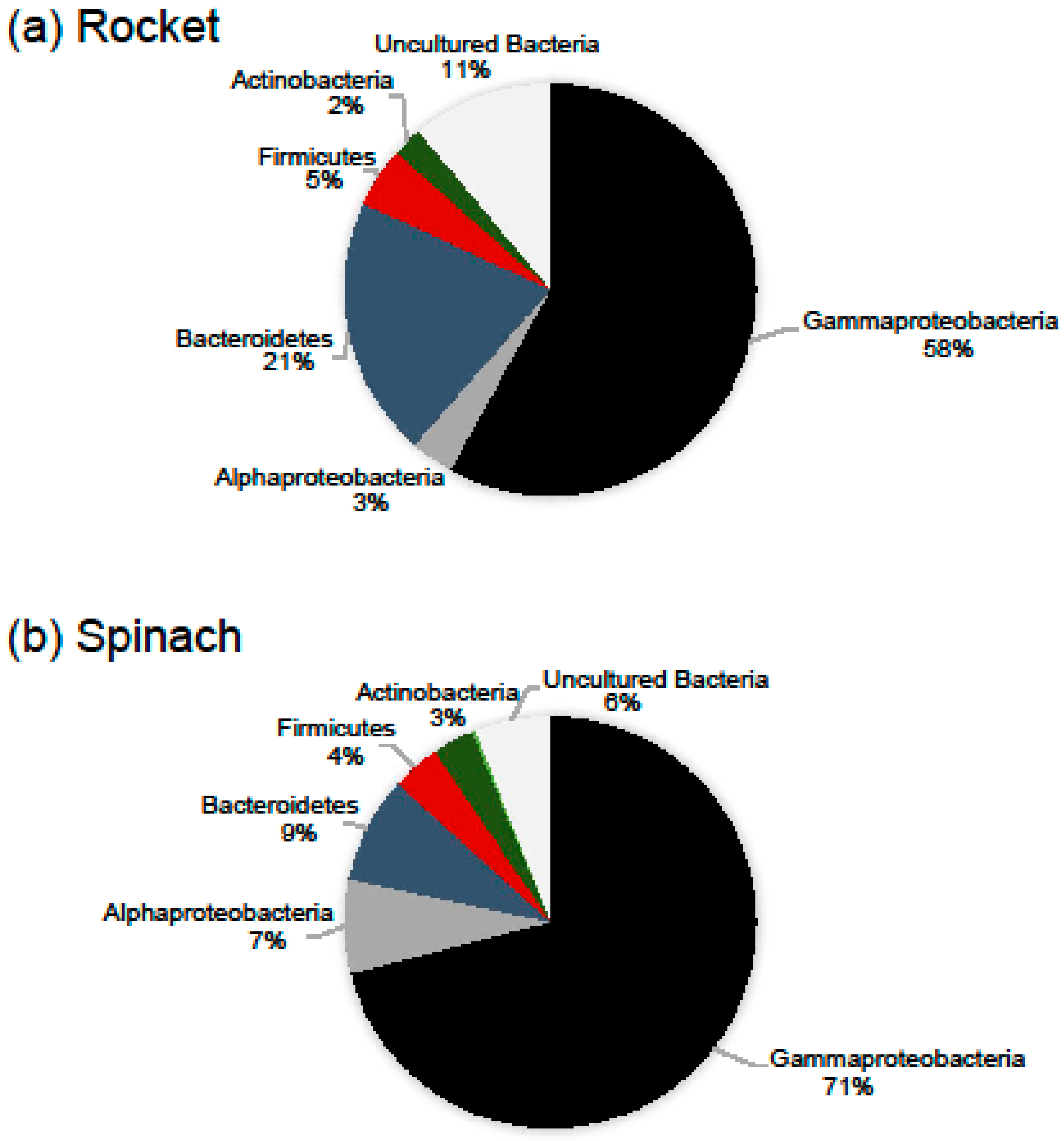
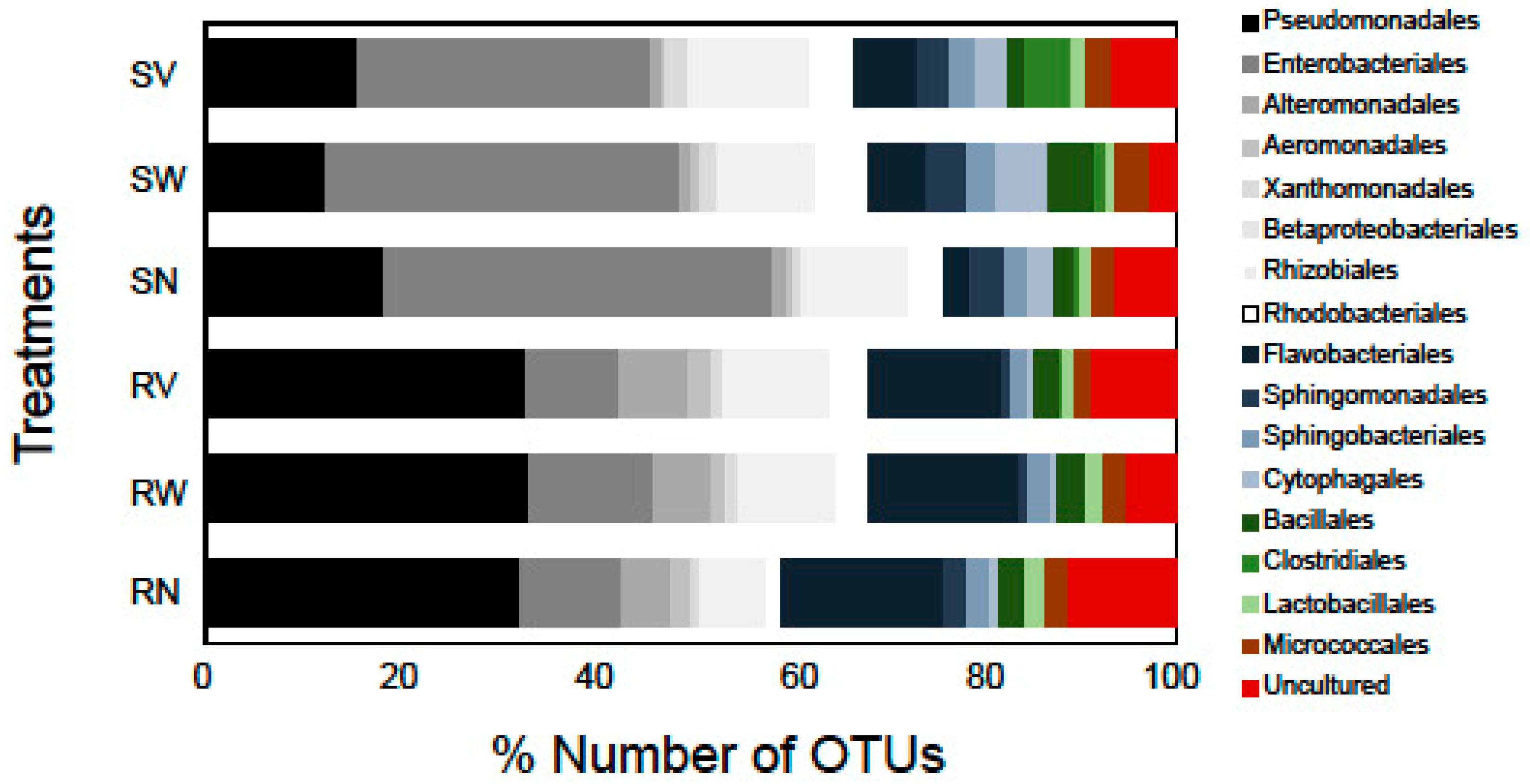
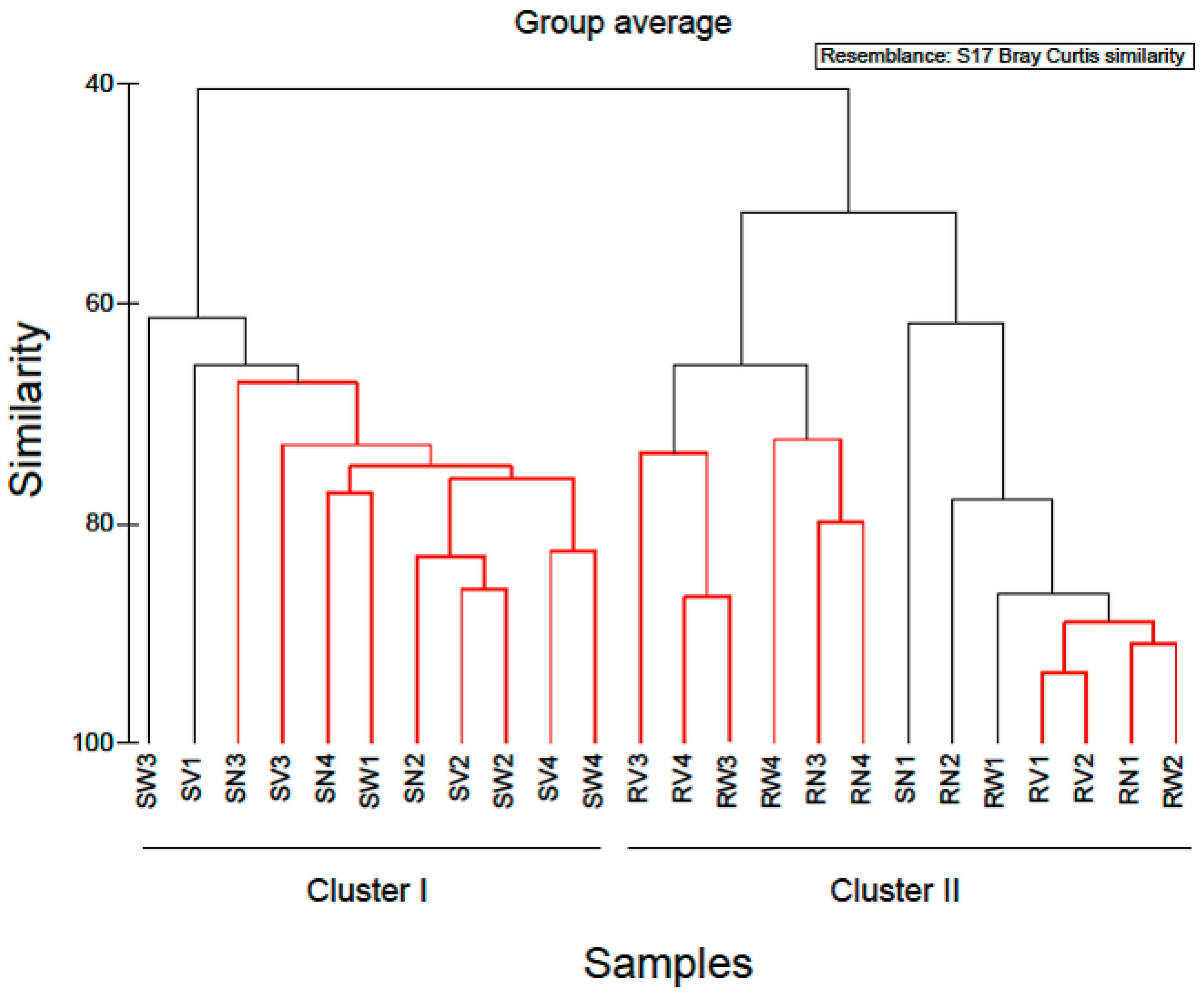
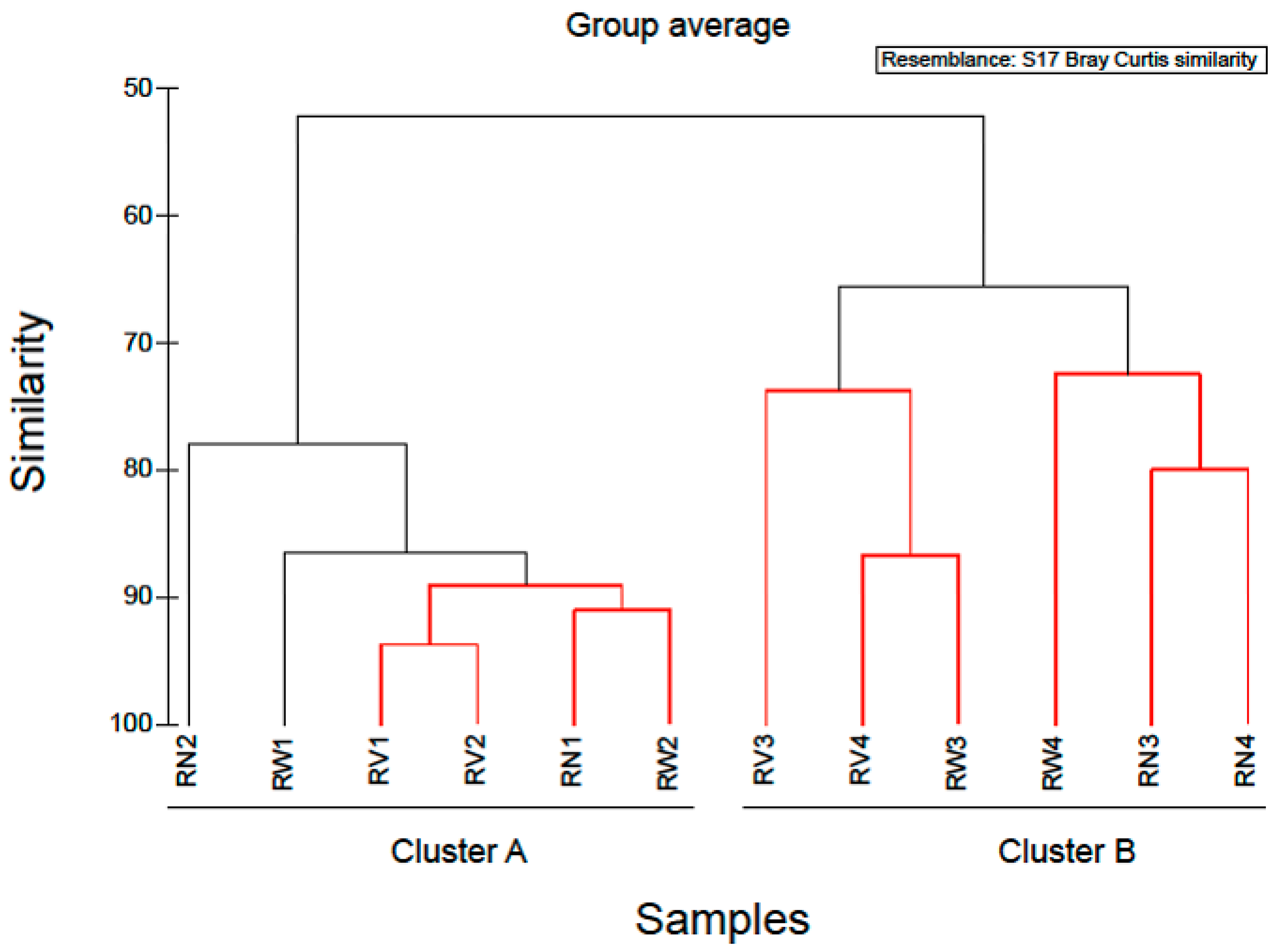
2.1. Bacterial Diversity and Composition of RTE Salads
2.2. Bacterial Diversity and Composition after Household Treatments
2.3. Changes in RTE Salad Microbiome during Refrigerated Storage
2.4. Potential Pathogens and Other Notable OTUs
3. Materials and Methods
3.1. Sample Processing
3.2. DNA Extraction and Sequencing
3.3. Read Processing
3.4. Data Analysis
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Berg, G.; Erlacher, A.; Smalla, K.; Krause, R. Vegetable microbiomes: Is there a connection among opportunistic infections, human health and our ‘gut feeling’? Microb. Biotechnol. 2014, 7, 487–495. [Google Scholar] [CrossRef] [PubMed]
- Berg, G.; Grube, M.; Schloter, M.; Smalla, K. The plant microbiome and its importance for human health. Front. Microbiol. 2014, 5, 491. [Google Scholar] [CrossRef]
- Brandl, M.T.; Cox, C.E.; Teplitski, M. Salmonella interactions with plants and their associated microbiota. Phytopathology 2013, 103, 316–325. [Google Scholar] [CrossRef]
- Critzer, F.J.; Doyle, M.P. Microbial ecology of foodborne pathogens associated with produce. Curr. Opin. Biotechnol. 2010, 21, 125–130. [Google Scholar] [CrossRef] [PubMed]
- Teplitski, A.; Warriner, K.; Bartz, J.; Schneider, K.R. Untangling metabolic and communication networks: Interactions of enterics with phytobacteria and their implications in produce safety. Trends Microbiol. 2011, 19, 121–127. [Google Scholar] [CrossRef] [PubMed]
- European Food Safety (EFSA). Authority Panel on Biological Hazards. Scientific Opinion on the risk posed by pathogens in food of non-animal origin. Part I. Efsa J. 2013, 11, 3025–3138. [Google Scholar]
- World Health Organization; Food and Agriculture Organization of the United Nations (WHO/FAO). Microbiological Risk Assessment Series No. 14. Microbiological Hazards in Fresh Leafy Vegetables and Herbs. Rome, 2008 (Meeting Report) 151. Available online: http://www.fao.org/3/a-i0452e.pdf (accessed on 26 June 2018).
- European Food Safety (EFSA); Authority and Austrian Institute of Technology (AIT) GmbH. Food of Plant Origin Production Methods and Microbiological Hazards Linked to Food—Borne Disease (Food of Plant Origin with High Water Content such as Fruits, Vegetables, Juices and Herbs); Efsa Supporting Publ.: Parma, Italy, 2013. [Google Scholar]
- Allende, A.; Aguayo, E.; Artes, F. Microbial and sensory quality of commercial fresh processed red lettuce throughout the production chain and shelf life. Int. J. Food Microbiol. 2004, 91, 109–117. [Google Scholar] [CrossRef]
- Babic, I.; Roy, S.; Watada, A.E.; Wergin, W.P. Changes in microbial populations on fresh cut spinach. Int. J. Food Microbiol. 1996, 31, 107–119. [Google Scholar] [CrossRef]
- Lopez-Velasco, G. Molecular Characterization of Spinach (Spinacia oleracea) Microbial Community Structure and Its Interaction with Escherichia coli O157:H7 in Modified Atmosphere Conditions. Ph.D. Thesis, Virginia Polytechnic Institute and State University, Blacksburg, VR, USA, 2010; p. 258. [Google Scholar]
- Turner, T.R.; James, E.K.; Poole, P.S. The plant microbiome. Genome Biol. 2013, 14, 209. [Google Scholar] [CrossRef]
- Lindow, S.E.; Brandl, M.T. Microbiology of the phyllosphere. Appl. Environ. Microbiol. 2003, 4, 1875–1883. [Google Scholar] [CrossRef]
- Heaton, J.C.; Jones, K. Microbial contamination of fruit and vegetables and the behaviour of enteropathogens in the phyllosphere: A review. J. Appl. Microbiol. 2007, 104, 613–626. [Google Scholar] [CrossRef] [PubMed]
- Bhatt, V.D.; Ahir, V.B.; Koringa, P.G.; Jakhesara, S.J.; Rank, D.N.; Nauriyal, D.S.; Joshi, C.G. Milk microbiome signatures of subclinical mastitis-affected cattle analysed by shotgun sequencing. J. Appl. Microbiol. 2012, 112, 639–650. [Google Scholar] [CrossRef] [PubMed]
- Ercolini, D. High-throughput sequencing and metagenomics: Moving forward in the culture-independent analysis of food microbial ecology. Appl. Environ. Microb. 2013, 79, 3148–3155. [Google Scholar] [CrossRef]
- Cao, Y.; Fanning, S.; Proos, S.; Jordan, K.; Srikumar, S. A Review on the Application of Next Generation Sequencing Technologies as Applied to Food Related Microbiome Studies. Front. Microbiol. 2017, 8, 1829. [Google Scholar] [CrossRef] [PubMed]
- Rastogi, G.; Sbodio, A.; Tech, J.J.; Suslow, T.V.; Coaker, G.L.; Leveau, J.H. Leaf microbiota in an agroecosystem: Spatiotemporal variation in bacterial community composition on field-grown lettuce. ISME J. 2012, 6, 1812–1822. [Google Scholar] [CrossRef]
- Jackson, C.R.; Stone, B.W.G.; Tyler, H.L. Emerging perspectives on the natural microbiome of fresh produce vegetables. Agriculture 2015, 5, 170–187. [Google Scholar] [CrossRef]
- Lopez-Velasco, G.; Davis, M.; Boyer, R.R.; Mane, S.P.; Ponder, M.A. Changes in spinach phylloepiphytic bacteria communities following minimal processing and refrigerated storage described using pyrosequencing of 16S rRNA amplicons. J. Appl. Microbiol. 2011, 110, 1203–1214. [Google Scholar] [CrossRef] [Green Version]
- Lopez-Velasco, G.; Davis, M.; Boyer, R.R.; Mane, S.P.; Ponder, M.A. Alterations of the phylloepiphytic bacterial community associated with interactions of Escherichia coli O157:H7 during storage of packaged spinach at refrigeration temperatures. Food Microbiol. 2010, 27, 476–486. [Google Scholar] [CrossRef]
- Jackson, C.R.; Randolph, K.C.; Osborn, S.L.; Tyler, H.L. Culture dependent and independent analysis of bacterial communities associated with commercial salad leaf vegetables. Bmc Microbiol. 2013, 13, 274. [Google Scholar] [CrossRef]
- Söderqvist, K.; Osman, O.A.; Wolff, C.; Bertilsson, S.; Vågsholm, I.; Boqvist, S. Emerging microbiota during cold storage and temperature abuse of ready–to-eat salad. Infect. Ecol. Epidemiol. 2017, 7, 1328963. [Google Scholar]
- Nastou, A.J.; Rhoades, P.; Smirniotis, I.; Makri, M.; Kontominas, M.; Likotrafity, E. Efficacy of household washing treatments for the control of Listeria monocytogenes on salad vegetables. Int. J. Food Microbiol. 2012, 159, 247–253. [Google Scholar] [CrossRef] [PubMed]
- Uhlig, E.; Olsson, C.; He, J.; Stark, T.; Sadowska, Z.; Molin, G.; Ahrne, S.; Alsanius, B.; Hakansson, A. Effects of household washing on bacterial load and removal of Escherichia coli from lettuce and ready-to-eat salads. Food Sci. Nutr. 2017, 5, 1215–1220. [Google Scholar] [CrossRef] [PubMed]
- Leff, J.W.; Fierer, N. Bacterial Communities Associated with the Surfaces of Fresh Fruits and Vegetables. PLoS ONE 2013, 8, e59310. [Google Scholar] [CrossRef]
- Rudi, K.; Flateland, S.L.; Hanssen, J.F.; Bengtsson, G.; Nissen, H. Development and evaluation of a16S ribosomal DNA array-based approach for describing complex microbial communities in ready-to-eat vegetable salads packed in modified atmosphere. Appl. Environ. Microb. 2002, 68, 1146–1156. [Google Scholar] [CrossRef]
- Dees, M.W.; Lysøe, E.; Nordskog, B.; Brurberg, M.B. Bacterial communities associated with surfaces of leafy greens: Shift in composition and decrease in richness over time. Appl. Environ. Microb. 2015, 81, 1530–1539. [Google Scholar] [CrossRef] [PubMed]
- Ragaert, P.; Devlieghere, F.; Debevere, J. Role of microbiological and physiological spoilage mechanisms during storage of minimally processed vegetables. Postharvest Biol. Technol. 2007, 44, 185–194. [Google Scholar] [CrossRef]
- Adams, M.R.; Moss, M.O. Food Microbiology, 2nd ed.; The Royal Society of Chemistry: Cambridge, UK, 2005; pp. 156–160. [Google Scholar]
- Jay, J.M.; Loessner, M.J.; Golden, D.A. Modern Food Microbiology, 7th ed.; Springer Science+Business Media. Inc.: New York, NY, USA, 2005. [Google Scholar]
- Knight, R.; Jannson, J.; Field, D.; Fierer, N.; Desai, N.; Fuhrman, J.A.; Hugenholtz, P.; van der Lelie, D.; Meyer, F.; Stevens, R.; et al. Unlocking the potential of metagenomics through replicated experimental design. Nat. Biotechnol. 2012, 30, 513–520. [Google Scholar] [CrossRef] [Green Version]
- Medinger, R.; Nolte, V.; Pandey, R.V.; Jost, S.; Ottenwälder, B.; Schlötterer, C.; Boenigk, J. Diversity in a hidden world: Potential and limitation of next-generation sequencing for surveys of molecular diversity of eukaryotic microorganisms. Mol. Ecol. 2010, 19, 32–40. [Google Scholar] [CrossRef]
- Andersen, S.M.; Johnsen, K.; Sørensen, J.; Nielsen, P.; Jacobsen, C.S. Pseudomonas frederiksbergensis sp. nov.; isolated from soil at a coal gasification site. Int. J. Syst. Evol. Microbiol. 2000, 50, 1957–1964. [Google Scholar] [CrossRef]
- Seifert, H.; Strate, A.; Schulze, A.; Pulverer, G. Vascular catheter-related bloodstream infection due to Acinetobacter johnsonii (formerly Acinetobacter calcoaceticus var. lwoffii): Report of 13 Cases. Clin. Infect. Dis. 1993, 17, 632–636. [Google Scholar] [CrossRef]
- Huang, H.C.; Hsieh, T.F.; Erickson, R.S. Biology and epidemiology of Erwinia rhapontici, causal agent of pink seed and crown rot of plants. Plant Pathol. Bull. 2003, 12, 69–76. [Google Scholar]
- Saranraj, P.; Stella, D.; Reetha, D. Microbial Spoilage of vegetables and its control measures: A review. Int. J. Nat. Prod. Sci. 2012, 2, 1–12. [Google Scholar]
- Liao, C.H. Bacterial soft rot. In Microbiology of Fruits and Vegetables; Sapers, G.M., Gorney, J.R., Yousef, A.E., Eds.; CRC Press: Boca Raton, FL, USA, 2005; pp. 117–134. [Google Scholar]
- Ryu, S.H.; Chung, B.S.; Park, M.; Lee, S.S.; Lee, S.-S.; Jeon, C.O. Rheinheimera soli sp. nov.; a gammaproteobacterium isolated from soil in Korea. Int. J. Syst. Evol. Microbiol. 2008, 58, 2271–2274. [Google Scholar] [CrossRef]
- Hao, W. A Potential Energy-Saving Heat Treatment for Re-Circulated Irrigation Water and Its Biological Mechanisms. Ph.D. Thesis, Virginia Polytechnic Institute and State University, Blacksburg, VR, USA, 2012. [Google Scholar]
- Balcht, A.; Smith, R. Pseudomonas aeruginosa: Infections and Treatment; Infectious Disease and Therapy Series Health Care Marcel Dekker, Inc.: New York, NY, USA, 1994; pp. 83–84. [Google Scholar]
- Fusco, V.; Abriouel, H.; Benomar, N.; Kabisch, J.; Chieffi, D.; Cho, G.; Franz, C. Opportunistic food-borne pathogens. In Food Safety Preservation; Grumezescu, A.M., Holban, A.M., Eds.; Academic Press: Cambridge, MA, USA, 2018; pp. 269–306. [Google Scholar]
- Brooke, J.S. Stenotrophomonas maltophilia: An emerging global opportunistic pathogen. Clin. Microbiol. Rev. 2012, 25, 2–41. [Google Scholar] [CrossRef]
- Schwaiger, K.; Helmke, K.; Hölzel, C.S.; Bauer, J. Antibiotic resistance in bacteria isolated from vegetables with regards to the marketing stage (farm vs. supermarket). Int. J. Food Microbiol. 2011, 148, 191–196. [Google Scholar] [CrossRef]
- Fröhling, A.; Rademacher, A.; Rumpold, B.; Klocke, M.; Schlüter, O. Screening of microbial communities associated with endive lettuce during postharvest processing on industrial scale. Heliyon 2018, 4, e00671. [Google Scholar] [CrossRef]
- Jones-Dias, D.; Manageiro, V.; Ferreira, E.; Barreiro, P.; Vieira, L.; Moura, I.B.; Caniça, M. Architecture of class 1, 2, and 3 integrons from gram negative bacteria recovered among fruits and vegetables. Front. Microbiol. 2016, 7, 1400. [Google Scholar] [CrossRef]
- Altwegg, M.; Geiss, H.K. Aeromonas as a human Pathogen. Crc. Cr. Rev. Microbiol. 1989, 16, 253–286. [Google Scholar] [CrossRef]
- Elhariry, H.M. Biofilm Formation by Aeromonas hydrophila on green-leafy vegetables: Cabbage and lettuce. Foodborne Pathog. Dis. 2011, 8, 125–131. [Google Scholar] [CrossRef]
- Gram, L.; Huss, H.H. Microbiological spoilage of fish and fish products. Int. J. Food Microbiol. 1996, 33, 121–137. [Google Scholar] [CrossRef]
- Rivera, M.E.D.; Vélez, C.; Zayas, B.; Llamas, K.M. Bacterial assessment on leaves of green vegetable grown on hydroponics and its possible health risks. J. Agric. Environ. Sci. 2015, 4, 1–7. [Google Scholar]
- Bagge, D.; Hjelm, M.; Johansen, C.; Huber, I.; Gram, L. Shewanella putrefaciens adhesion and biofilm formation on food processing surfaces. Appl. Environ. Microb. 2001, 67, 2319–2325. [Google Scholar] [CrossRef] [PubMed]
- Pagani, L.; Lang, A.; Vedovelli, C.; Moling, O.; Rimenti, G.; Pristerà, R.; Mian, P. Soft tissue infection and bacteremia caused by Shewanella putrefaciens. J. Clin. Microbiol. 2003, 41, 2240–2241. [Google Scholar] [CrossRef]
- EC Regulation 1441/2007. Commission Regulation Amending Regulation 2073/2005 on Microbiological Criteria for Foodstuffs; Official Journal of the European Union: Luxembourg, 2007. [Google Scholar]
- Comeau, A.M.; Li, W.K.; Tremblay, J.E.; Carmack, E.C.; Lovejoy, C. Arctic Ocean microbial community structure before and after the 2007 record sea ice minimum. PLoS ONE 2011, 6, e27492. [Google Scholar] [CrossRef] [PubMed]
- Schloss, P.D.; Gevers, D.; Westcott, S.L. Reducing the effects of PCR amplification and sequencing artifacts on 16S rRNA-based studies. PLoS ONE 2011, 6, e27310. [Google Scholar] [CrossRef] [PubMed]
- Schloss, P.D.; Westcott, S.L.; Ryabin, T.; Hall, J.R.; Hartmann, M.; Hollister, E.B.; Lesniewski, R.A.; Oakley, B.B.; Parks, D.H.; Robinson, C.J. Introducing mother: Open-source, platform-independent, community-supported software for describing and comparing microbial communities. Appl. Environ. Microb. 2009, 75, 7537–7541. [Google Scholar] [CrossRef] [PubMed]
- Quast, C.; Pruesse, E.; Yilmaz, P.; Gerken, J.; Schweer, T.; Yarza, P.; Peplies, J.; Glöckner, F.O. The SILVA ribosomal RNA gene database project: Improved data processing and web-based tools. Nucleic Acids Res. 2012, 41, D590–D596. [Google Scholar] [CrossRef]
- Edgar, R.C. Search and clustering orders of magnitude faster than BLAST. Bioinformatics 2010, 26, 2460–2461. [Google Scholar] [CrossRef] [Green Version]
- Kunin, V.; Engelbrektson, A.; Ochman, H.; Hugenholtz, P. Wrinkles in the rare biosphere: Pyrosequencing errors can lead to artificial inflation of diversity estimates. Environ. Microbiol. 2010, 12, 118–123. [Google Scholar] [CrossRef]
- Richards, T.A.; Leonard, G.; Mahé, F.; del Campo, J.; Romac, S.; Jones, M.D.M.; Maguire, F.; Dunthorn, M.; De Vargas, C.; Massana, R.; et al. Molecular diversity and distribution of marine fungi across 130 european environmental samples. Proc. R. Soc. B 2015, 282, 20152243. [Google Scholar] [CrossRef]
- Genitsaris, S.; Monchy, S.; Viscogliosi, E.; Sime-Ngando, T.; Ferreira, S.; Christaki, U. Seasonal variations of marine protist community structure based on taxon-specific traits using the eastern English Channel as a model coastal system. Fems Microbiol. Ecol. 2015, 91, fiv034. [Google Scholar] [CrossRef]
- Christaki, U.; Kormas, K.A.; Genitsaris, S.; Georges, C.; Sime-Ngando, T.; Viscogliosi, E.; Monchy, S. Winter-summer succession of unicellular eukaryotes in a meso-eutrophic coastal system. Microb. Ecol. 2014, 67, 13–23. [Google Scholar] [CrossRef] [PubMed]
- Genitsaris, S.; Monchy, S.; Breton, E.; Lecuyer, E.; Christaki, U. Small-scale variability of protistan communities relative to environmental pressures and biotic interactions at two adjacent coastal stations. Mar. Ecol. Prog. Ser. 2016, 548, 61–75. [Google Scholar] [CrossRef]
- Genitsaris, S.; Stefanidou, N.; Katsiapi, M.; Kormas, K.A.; Sommer, U.; Moustaka-Gouni, M. Variability of airborne bacteria in an urban Mediterranean area (Thessaloniki, Greece). Atmos. Environ. 2017, 157, 101–110. [Google Scholar] [CrossRef]
- Pruesse, E.; Peplies, J.; Glöckner, F.O. SINA: Accurate high-throughput multiple sequence alignment of ribosomal RNA genes. Bioinformatics 2012, 28, 1823–1829. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hammer, O.; Harper, D.A.T.; Ryan, P.D. PAST: Paleontological statistics software package for education and data analysis. Paleontol. Electron. 2001, 4, 9. [Google Scholar]
- Clarke, K.R.; Gorley, R.N. Primer v6: User Manual/Tutorial; PRIMER-E: Plymouth, UK, 2006. [Google Scholar]
- Clarke, K.R.; Warwick, R.M. Similarity-based testing for community pattern: The two-way layout with no replication. Mar. Biol. 1996, 118, 167–176. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| RTE Salad | Expiring Date | Number of Replicates | Treatment | Code |
|---|---|---|---|---|
| Rocket | 20 June 2018 | 2 | No treatment | RN1–RN2 |
| Water | RW1–RW2 | |||
| Acetic Acid Solution | RV1–RV2 | |||
| Rocket | 23 June 2018 | 2 | No treatment | RN3–RN4 |
| Water | RW3–RW4 | |||
| Acetic Acid Solution | RV3–RV4 | |||
| Spinach | 23 June 2018 | 4 | No treatment | SN1–SN4 |
| Water | SW1–SW4 | |||
| Acetic Acid Solution | SV1–SV4 |
| OTUs | Putative High-Level Taxonomic Affiliation | Closest Cultured Relative (% Similarity) [Accession Number] | Isolation Source | SIMPER Contribution (%) | SIMPER Cumulative Contribution (%) | RTE Salad | Relative Abundance (%) |
|---|---|---|---|---|---|---|---|
| OTU001 | Gammaproteobacteria | Pseudomonas frederiksbergensis [MH144327] | Soil at a coal gasification site | 20.51 | 20.51 | All | 35.2 |
| OTU003 | Gammaproteobacteria | Erwinia rhapontici [LC424328] | Barkey | 10.78 | 31.28 | All | 7.4 |
| OTU002 | Gammaproteobacteria | Acinetobacter johnsonii [MK294307] | Mine tailings | 8.77 | 40.05 | All | 7.9 |
| OTU004 | Gammaproteobacteria | Rheinheimera sp. [LC270228] | Wastewater stream | 7.58 | 47.64 | All rocket samples & SN2 SV1 SV3 SW1 SW2 | 4.8 |
| OTU008 | Gammaproteobacteria | Serratia sp. [KU750792] | Rhizosphere from Lepidium meyenii | 5.08 | 52.72 | All | 3.7 |
| OTU0010 | Gammaproteobacteria | Pantoea agglomerans [MG681225] | Commercial Cucumber Fermentation Cover brine | 4.51 | 57.23 | All | 2.8 |
| OTU007 | Bacteroidetes | Flavobacterium resistens [MH549189] | Insuyu cave | 4.32 | 61.55 | All | 3.4 |
| OTU0011 | Gammaproteobacteria | Pantoea agglomerans [MH101508] | Chelidonium majus (medical herb) | 4 | 65.56 | All | 2.5 |
| OTU0014 | Gammaproteobacteria | Pantoea sp. B1(2013) [KF010367] | Plant root | 3 | 68.55 | All spinach samples & RN3RN4RV3RV4RW2RW3RW4 | 1.8 |
| OTU005 | Gammaproteobacteria | Pseudomonas aeruginosa [MF838682] | Dairy product | 2.72 | 71.27 | All | 4.3 |
| OTU0015 | Gammaproteobacteria | Enterobacter sp. [MG681230] | Commercial Cucumber Fermentation Cover brine | 2.08 | 73.35 | All | 1.7 |
| OTU0016 | Gammaproteobacteria | Kluyvera intermedia [MH620740] | - | 1.8 | 75.16 | All | 1.5 |
| OTU009 | Gammaproteobacteria | Pseudomonas viridiflava [MG972916] | Fresh-cut escarole | 1.69 | 76.85 | All | 2.8 |
| OTU0012 | Gammaproteobacteria | Janthinobacterium sp. [MF774126] | Himalayan region | 1.5 | 78.35 | All | 2.1 |
| OTU0013 | Gammaproteobacteria | Duganella zoogloeoides [KT983992] | Fresh water | 1.49 | 79.84 | All | 2.1 |
| OTU0017 | Bacteroidetes | Chryseobacterium indoltheticum [MK138643] | Marine mud | 1.31 | 81.14 | All | 1.2 |
| OTU0019 | Alphaproteobacteria | Sphingomonas faeni [MH482321] | Indoor dusts in animal sheds | 1.28 | 82.42 | All except RN1. RW1. RV1 | 0.8 |
| OTU0018 | Gammaproteobacteria | Aeromonas hydrophila [MK038972] | Freshwater cage culture system | 1.03 | 83.46 | All except SW2 | 0.9 |
| OTU0021 | Firmicutes | Paenibacillus sp. [MH769399] | - | 0.98 | 84.44 | All | 0.7 |
| OTU0023 | Gammaproteobacteria | Comamonas jiangduensis [MH712950] | “Oncorhynchus tshawytscha” (Salmon) | 0.94 | 85.38 | All except SW2. SW4. SV2 | 0.7 |
| OTU0026 | Gammaproteobacteria | Enterobacter ludwigii [MH137696] | “Plutella xylostella” (diamondback moth) | 0.83 | 86.21 | All | 0.5 |
| OTU0031 | Gammaproteobacteria | Pantoea ananatis [KX891513] | - | 0.77 | 86.98 | All spinach samples & RN2RN3RV3RV4RW3RW4 | 0.5 |
| OTU0020 | Gammaproteobacteria | Massilia aurea [KY047391] | Wetland. Drinking water distribution system | 0.65 | 87.63 | All | 0.7 |
| OTU0027 | Gammaproteobacteria | Pectobacterium zantedeschiae [MG761827] | “Zantedeschia sp. (calla lily)” Tubers | 0.61 | 88.24 | All | 0.6 |
| OTU0032 | Gammaproteobacteria | Erwinia sp. [MG859640] | Skin | 0.57 | 88.81 | All | 0.4 |
| OTU0030 | Gammaproteobacteria | Acidovorax sp. [JQ723711] | Chelidonium majus root | 0.52 | 89.33 | All except SW4 | 0.4 |
| OTU0024 | Firmicutes | Exiguobacterium antarcticum [MH125158] | Soil | 0.5 | 89.83 | All | 0.6 |
| OTU0022 | Bacteroidetes | Sphingobacterium faecium [MK100919] | Rhizosphere and roots of wheat | 0.5 | 90.33 | All | 0.7 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tatsika, S.; Karamanoli, K.; Karayanni, H.; Genitsaris, S. Metagenomic Characterization of Bacterial Communities on Ready-to-Eat Vegetables and Effects of Household Washing on their Diversity and Composition. Pathogens 2019, 8, 37. https://0-doi-org.brum.beds.ac.uk/10.3390/pathogens8010037
Tatsika S, Karamanoli K, Karayanni H, Genitsaris S. Metagenomic Characterization of Bacterial Communities on Ready-to-Eat Vegetables and Effects of Household Washing on their Diversity and Composition. Pathogens. 2019; 8(1):37. https://0-doi-org.brum.beds.ac.uk/10.3390/pathogens8010037
Chicago/Turabian StyleTatsika, Soultana, Katerina Karamanoli, Hera Karayanni, and Savvas Genitsaris. 2019. "Metagenomic Characterization of Bacterial Communities on Ready-to-Eat Vegetables and Effects of Household Washing on their Diversity and Composition" Pathogens 8, no. 1: 37. https://0-doi-org.brum.beds.ac.uk/10.3390/pathogens8010037