Quantification of the Animal Tuberculosis Multi-Host Community Offers Insights for Control

 , , ,
, , ,  , ,
, ,
Abstract
:1. Introduction
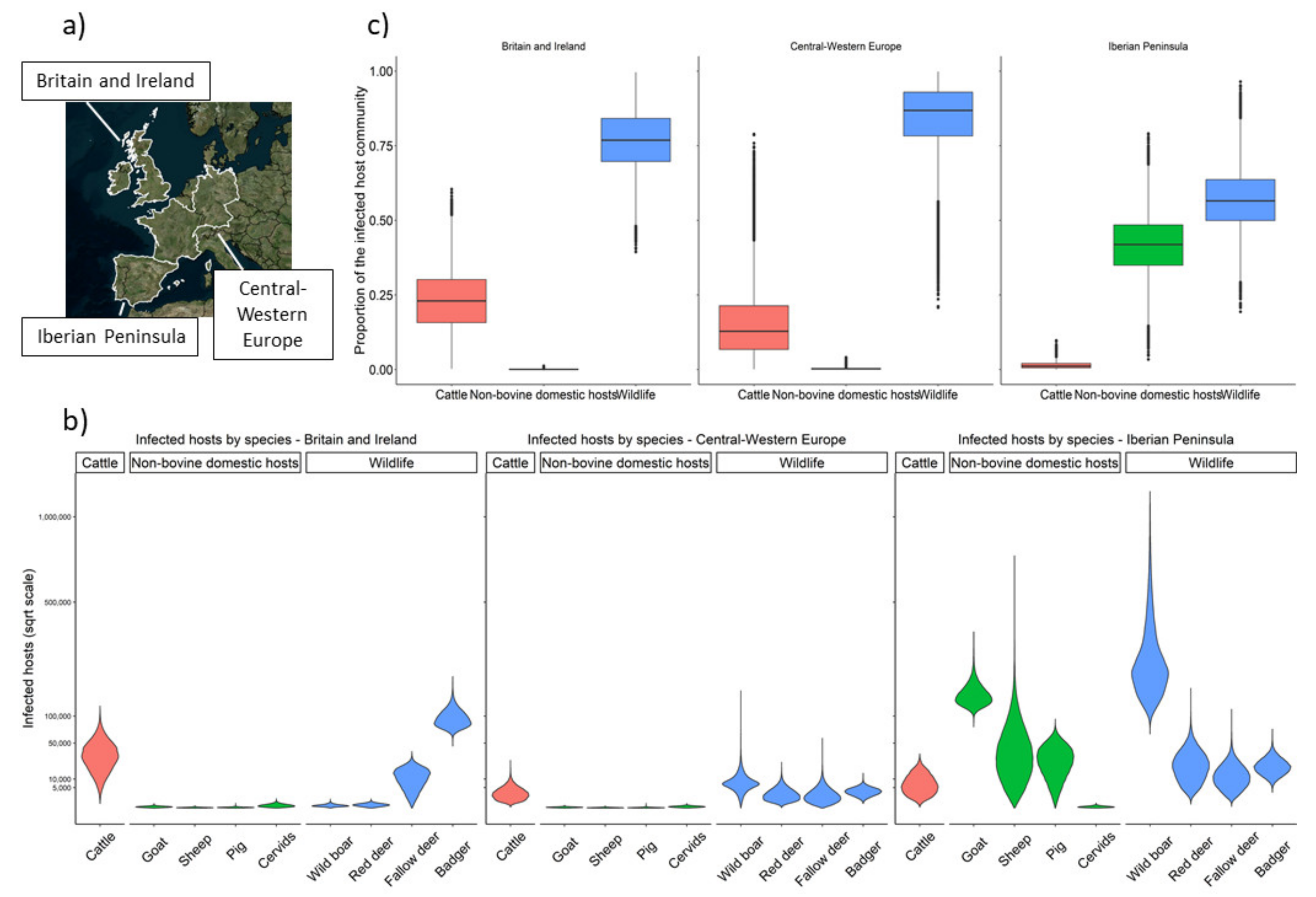
2. Results
3. Discussion
4. Materials and Methods
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Gortázar, C.; Delahay, R.J.; Mcdonald, R.A.; Boadella, M.; Wilson, G.J.; Gavier-Widen, D.; Acevedo, P. The status of tuberculosis in European wild mammals. Mammal Rev. 2012, 42, 193–206. [Google Scholar] [CrossRef]
- Webster, J.P.; Borlase, A.; Rudge, J.W. Who acquires infection from whom and how? Disentangling multi-host and multi-mode transmission dynamics in the ‘elimination’ era. Phil. Trans. R. Soc. B 2017, 372, 20160091. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Haydon, D.T.; Cleaveland, S.; Taylor, L.H.; Laurenson, M.K. Identifying reservoirs of infection: A conceptual and practical challenge. Emerg. Infect. Dis. 2002, 8, 1468–1473. [Google Scholar] [PubMed]
- Crispell, J.; Benton, C.H.; Balaz, D.; De Maio, N.; Ahkmetova, A.; Allen, A.; Biek, R.; Presho, E.L.; Dale, J.; Hewinson, G.; et al. Combining genomics and epidemiology to analyse bi-directional transmission of Mycobacterium bovis in a multi-host system. ELife 2019, 8, e45833. [Google Scholar] [CrossRef] [PubMed]
- Hauer, A.; De Cruz, K.; Cochard, T.; Godreuil, S.; Karoui, C.; Henault, S.; Bulach, T.; Bañuls, A.L.; Biet, F.; Boschiroli, M.L. Genetic evolution of Mycobacterium bovis causing tuberculosis in livestock and wildlife in France since 1978. PLoS ONE 2015, 10, e0117103. [Google Scholar] [CrossRef] [PubMed]
- Broughan, J.M.; Judge, J.; Ely, E.; Delahay, R.J.; Wilson, G.; Clifton-Hadley, R.S.; Goodchild, A.V.; Bishop, H.; Parry, J.E.; Downs, S.H. A review of risk factors for bovine tuberculosis infection in cattle in the UK and Ireland. Epidemiol. Infect. 2016, 144, 2899–2926. [Google Scholar] [CrossRef] [Green Version]
- LaHue, N.P.J.; Vicente, J.; Acevedo, P.; Gortázar, C.; Martínez-López, B. Spatially explicit modeling of animal tuberculosis at the wildlife-livestock interface in Ciudad Real province. Spain Prev. Vet. Med. 2016, 128, 101–111. [Google Scholar]
- Napp, S.; Allepuz, A.; Mercader, I.; Nofrarías, M.; López-Soria, S.; Domingo, M.; Romero, B.; Bezos, J.; De Val, B.P. Evidence of goats acting as domestic reservoirs of bovine tuberculosis. Vet. Rec. 2013, 172, 663. [Google Scholar] [CrossRef] [Green Version]
- Little, T.W.; Naylor, P.F.; Wilesmith, J.W. Laboratory study of Mycobacterium bovis infection in badgers and calves. Vet. Rec. 1982, 111, 550–557. [Google Scholar]
- Donnelly, C.A.; Woodroffe, R.; Cox, D.R.; Bourne, F.J.; Cheeseman, C.L.; Clifton-Hadley, R.S.; Wei, G.; Gettinby, G.; Gilks, P.; Jenkins, H.; et al. Positive and negative effects of widespread badger culling on tuberculosis in cattle. Nature 2006, 439, 843. [Google Scholar] [CrossRef]
- Barasona, J.A.; Gortázar, C.; de la Fuente, J.; Vicente, J. Host richness increases tuberculosis disease risk in game-managed areas. Microorganisms 2019, 7, 182. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brooks-Pollock, E.; Roberts, G.O.; Keeling, M.J.A. A dynamic model of bovine tuberculosis spread and control in Great Britain. Nature 2014, 511, 228. [Google Scholar] [CrossRef] [PubMed]
- Reviriego-Gordejo, F.; Vermeersch, J.P. Towards eradication of bovine tuberculosis in the European Union. Vet. Microbiol. 2006, 112, 101–109. [Google Scholar] [CrossRef] [PubMed]
- European Food Safety Authority; European Centre for Disease Prevention and Control. The European Union summary report on trends and sources of zoonoses, zoonotic agents and food-borne outbreaks in 2017. EFSA J. 2018, 16, 5500. [Google Scholar]
- European Union. Animal Health Funding, National Veterinary Programmes. Available online: https://ec.europa.eu/food/funding/animal-health/national-veterinary-programmes_en (accessed on 22 February 2018).
- Humblet, M.F.; Boschiroli, M.L.; Saegerman, C. Classification of worldwide bovine tuberculosis risk factors in cattle: A stratified approach. Vet. Res. 2009, 40, 1–24. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Muñoz-Mendoza, M.; Romero, B.; Del Cerro, A.; Gortázar, C.; García-Marín, J.F.; Menéndez, S.; Mourelo, J.; de Juan, L.; Sáez, J.L.; Delahay, R.J.; et al. Sheep as a potential source of bovine TB: Epidemiology, pathology and evaluation of diagnostic techniques. Transbound. Emerg. Dis. 2015, 63, 635–646. [Google Scholar] [CrossRef] [PubMed]
- Vose, D. Risk Analysis: A Quantitative Guide, 3rd ed.; John Wiley & Sons: Chichester, UK, 2008. [Google Scholar]
- Vicente, J.; Höfle, U.; Garrido, J.M.; Fernández-De-Mera, I.G.; Juste, R.; Barral, M.; Gortázar, C. Wild boar and red deer display high prevalences of tuberculosis-like lesions in Spain. Vet. Res. 2006, 37, 107–119. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Massei, G.; Kindberg, J.; Licoppe, A.; Gačić, D.; Šprem, N.; Kamler, J.; Baubet, E.; Hohmann, U.; Monaco, A.; Ozoliņš, J.; et al. Wild boar populations up, numbers of hunters down? A review of trends and implications for Europe. Pest Manag. Sci. 2015, 71, 492–500. [Google Scholar] [CrossRef]
- Santos, N.; Almeida, V.; Gortázar, C.; Correia-Neves, M. Patterns of Mycobacterium tuberculosis-complex excretion and characterization of super-shedders in naturally-infected wild boar and red deer. Vet. Res. 2015, 46, 129. [Google Scholar] [CrossRef] [Green Version]
- Judge, J.; Wilson, G.J.; Macarthur, R.; McDonald, R.A.; Delahay, R.J. Abundance of badgers (Meles meles) in England and Wales. Sci. Rep. 2017, 7, 276. [Google Scholar] [CrossRef] [PubMed]
- Gallagher, J.; Clifton-Hadley, R.S. Tuberculosis in badgers; a review of the disease and its significance for other animals. Res. Vet. Sci. 2000, 69, 203–217. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Delahay, R.J.; Walker, N.; Smith, G.S.; Wilkinson, D.; Clifton-Hadley, R.S.; Cheeseman, C.L.; Tomlinson, A.J.; Chambers, M.A. Long-term temporal trends and estimated transmission rates for Mycobacterium bovis infection in an undisturbed high-density badger (Meles meles) population. Epidemiol. Infect. 2013, 141, 1445–1456. [Google Scholar] [CrossRef] [PubMed]
- Bouchez-Zacria, M.; Courcoul, A.; Durand, B. The distribution of bovine tuberculosis in cattle farms is linked to cattle trade and badger-mediated contact networks in South-Western France, 2007–2015. Front Vet. Sci. 2018, 5, 173. [Google Scholar] [CrossRef] [PubMed]
- ENETWILD Consortium; Vicente, J.; Plhal, R.; Blanco-Aguiar, J.A.; Sange, M.; Podgórski, T.; Petrovic, K.; Scandura, M.; Nabeiro, A.C.; Body, G.; et al. Analysis of hunting statistics collection frameworks for wild boar across Europe and proposals for improving the harmonization of data collection. EFSA Supp. Pub. 2018, 15, 1523. [Google Scholar]
- Takkouche, B.; Khudyakov, P.; Costa-Bouzas, J.; Spiegelman, D. Confidence intervals for heterogeneity measures in meta-analysis. Am. J. Epid. 2013, 178, 993–1004. [Google Scholar] [CrossRef] [Green Version]
- Nuñez-García, J.; Downs, S.H.; Parry, J.E.; Abernethy, D.A.; Broughan, J.M.; Cameron, A.R.; Cook, A.J.; de la Rua-Domenech, R.; Goodchild, A.V.; Gunn, J.; et al. Meta-analyses of the sensitivity and specificity of ante-mortem and post-mortem diagnostic tests for bovine tuberculosis in the UK and Ireland. Prev. Vet. Med. 2017, 153, 94–107. [Google Scholar] [CrossRef]
- More, S.J.; Houtsma, E.; Doyle, L.; McGrath, G.; Clegg, T.A.; de la Rua-Domenech, R.; Duignan, A.; Blissitt, M.J.; Dunlop, M.; Schroeder, P.G.; et al. Further description of bovine tuberculosis trends in the United Kingdom and the Republic of Ireland, 2003–2015. Vet. Rec. 2018, 183, 717. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cano-Terriza, D.; Risalde, M.A.; Rodríguez-Hernández, P.; Napp, S.; Fernández-Morente, M.; Moreno, I.; Bezos, J.; Fernánde-Molera, V.; Sáez, J.L.; García-Bocanegra, I. Epidemiological surveillance of Mycobacterium tuberculosis complex in extensively raised pigs in the south of Spain. Prev. Vet. Med. 2018, 159, 87–91. [Google Scholar] [CrossRef]
- Gortázar, C.; Diez-Delgado, I.; Barasona, J.A.; Vicente, J.; De La Fuente, J.; Boadella, M. The wild side of disease control at the wildlife-livestock-human interface: A review. Front. Vet. Sci. 2015, 1, 27. [Google Scholar] [CrossRef] [Green Version]
- Wilson, G.J.; Carter, S.P.; Delahay, R.J. Advances and prospects for management of TB transmission between badgers and cattle. Vet. Microbiol. 2011, 151, 43–50. [Google Scholar] [CrossRef] [Green Version]
- Devleesschauwer, B.; Torgerson, P.; Charlier, J.; Levecke, B.; Praet, N.; Roelandt, S.; Smit, S.; Dorny, P.; Berkvens, D.; Speybroeck, N. Prevalence: Tools for Prevalence Assessment Studies. R Package Version 0.4.0. 2014. Available online: http://cran.r-project.org/package=prevalence (accessed on 12 December 2018).
- R Development Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2008; ISBN 3-900051-07-0. Available online: http://www.R-project.org (accessed on 12 December 2018).
- Pouillot, R.; Delignette-Muller, M.L. Evaluating variability and uncertainty in microbial quantitative risk assessment using two R packages. Int. J. Food Microbiol. 2010, 142, 330–340. [Google Scholar] [CrossRef] [PubMed]
- Geweke, J. Evaluating the accuracy of sampling-based approaches to calculating posterior moments. In Bayesian Statistics, 4th ed.; Bernardo, J.M., Berger, J.O., Dawid, A.P., Smith, A.F.M., Eds.; Oxford University Press: New York, NY, USA, 1992. [Google Scholar]
- Brooks, S.P.; Gelman, A. General methods for monitoring convergence of iterative simulations. J. Comput. Graph. Stat. 1998, 7, 434–455. [Google Scholar]
- Toft, N.; Innocent, G.T.; Gettinby, G.; Reid, S.W. Assessing the convergence of Markov Chain Monte Carlo methods: An example from evaluation of diagnostic tests in absence of a gold standard. Prev. Vet. Med. 2007, 79, 244–256. [Google Scholar] [CrossRef] [PubMed]
- Zhou, X.H.; McClish, D.K.; Obuchowski, N.A. Statistical Methods in Diagnostic Medicine; John Wiley & Sons: New York, NY, USA, 2009. [Google Scholar]
- Eurostat. Livestock and Meat Production Database, 2015 Data. Available online: http://ec.europa.eu/eurostat/web/agriculture/ data/database (accessed on 15 December 2017).
- MAGRAMA. Database Encuestas Ganaderas, Ganado Porcino, 2014 Data. Available online: https://www.mapa.gob.es/es/estadistica/temas/estadisticas-agrarias/ganaderia/encuestas-ganaderas/ (accessed on 15 December 2017).
- DGAV. Animal Movement Database Portugal, Swine, 2012 Data. Available online: http://www.dgv.min-agricultura.pt/portal/page/portal/DGV (accessed on 15 December 2017).
- Toïgo, C.; Servanty, S.; Gaillard, J.M.; Brandt, S.; Baubet, E. Disentangling natural from hunting mortality in an intensively hunted wild boar population. J. Wildl. Manag. 2008, 72, 1532–1539. [Google Scholar]
- Keuling, O.; Baubet, E.; Duscher, A.; Ebert, C.; Fischer, C.; Monaco, A.; Podgórski, T.; Prevot, C.; Ronnenberg, K.; Sodeikat, G.; et al. Mortality rates of wild boar Sus scrofa L. in central Europe. Eur. J. Wildl. Res. 2013, 59, 805–814. [Google Scholar] [CrossRef]
- Barasona, J.A.; Acevedo, P.; Diez-Delgado, I.; Queirós, J.; Carrasco-García, R.; Gortázar, C.; Vicente, J. Tuberculosis-associated death among adult wild boars, Spain, 2009–2014. Emerg. Infect. Dis. 2016, 22, 2178. [Google Scholar] [CrossRef] [Green Version]
- Merli, E.; Grignolio, S.; Marcon, A.; Apollonio, M. Wild boar under fire: The effect of spatial behaviour, habitat use and social class on hunting mortality. J. Zool. 2017, 303, 155–164. [Google Scholar] [CrossRef]
- Langvatn, R.; Loison, A. Consequences of harvesting on age structure, sex ratio and population dynamics of red deer Cervus elaphus in central Norway. Wildl. Biol. 1999, 5, 213–223. [Google Scholar]
- Burbaitė, L.; Csányi, S. Red deer population and harvest changes in Europe. Acta Zool. Lit. 2010, 20, 179–188. [Google Scholar] [CrossRef] [Green Version]
- Torres-Porras, J.; Carranza, J.; Pérez-González, J.; Mateos, C.; Alarcos, S. The tragedy of the commons: Unsustainable population structure of Iberian red deer in hunting estates. Eur. J. Wildl. Res. 2014, 60, 351–357. [Google Scholar] [CrossRef]
- Oksanen, J.; Blanchet, F.G.; Friendly, M.; Kindt, R.; Legendre, P.; McGlinn, D.; Minchin, P.R.; O’Hara, R.B.; Simpson, G.L.; Solymos, P.; et al. Package ‘vegan’. Community Ecology Package, Version, 2.5.6. Available online: https://CRAN.R-project.org/package=vegan (accessed on 12 December 2018).
- Simpson, E.H. Measurement of diversity. Nature 1949, 163, 688. [Google Scholar] [CrossRef]
- Wickham, H. ggplot2: Elegant Graphics for Data Analysis; Springer: New York, NY, USA, 2009. [Google Scholar]
- QGIS Development Team. QGIS Geographic Information System. Open Source Geospatial Foundation Project. 2018. Available online: http://qgis.osgeo.org (accessed on 12 December 2018).


{kind=link}
| Host Species | Estimated Number of Infected Hosts | ||||||||
|---|---|---|---|---|---|---|---|---|---|
| Britain and Ireland | Central–Western Europe | Iberian Peninsula | Total | ||||||
| Median | Credible Interval95 | Median | Credible Interval95 | Median | Credible Interval95 | Median | Credible Interval95 | ||
| Cattle | Cattle | 29,871 | 1279–89,606 | 2303 | 98–9460 | 6175 | 253–22,356 | 38,349 | 1630–121,422 |
| Non-bovine domestic | Goat | 21 | 1–83 | 9 | 0–36 | 145,906 | 106,874–232,240 | 145,936 | 106,875–232,359 |
| Sheep | 4 | 0–20 | 3 | 0–16 | 32.430 | 1343–146,771 | 32,437 | 1343–146,807 | |
| Pigs | 7 | 0–33 | 3 | 0–22 | 25.392 | 1528–60,716 | 25,402 | 1528–60,771 | |
| Cervids (farmed) | 73 | 3–387 | 26 | 1–101 | 21 | 1–115 | 120 | 5–603 | |
| Wildlife | Wild boar | 74 | 8–325 | 7118 | 846–22,716 | 229,629 | 103,787–693,486 | 236,821 | 104,641–716,527 |
| Red deer | 119 | 5–452 | 1912 | 68–9,033 | 20,134 | 953–62,764 | 22,165 | 1026–72,249 | |
| Fallow deer | 10,621 | 559–24,259 | 0 | 0 | 10,693 | 659–35,678 | 21,314 | 1218–59,937 | |
| Badger | 91,643 | 62,310–148,225 | 3231 | 940–6756 | 20,403 | 8543–39,957 | 115,277 | 71,793–194,938 | |
| Total | 132,433 | 64,165–263,390 | 14,605 | 1953–48,140 | 490,783 | 223,941–1,294,082 | 637,821 | 290,059–1,605,612 | |
| Region | Class of Host | Ratio Non-Bovine Domestic and Wildlife Hosts/Cattle | |
|---|---|---|---|
| Median | Credible Interval95 | ||
| Britain and Ireland | Non-bovine domestic species | 0.004 | 0.0006–0.03 |
| Wildlife | 3.3 | 1.3–19.6 | |
| Total | 3.3 | 1.3–19.6 | |
| Central-Western Europe | Non-bovine domestic species | 0.02 | 0.003–0.2 |
| Wildlife | 6.8 | 1.4–58.3 | |
| Total | 6.8 | 1.4–58.5 | |
| Iberian Peninsula | Non-bovine domestic species | 35.0 | 9.2–342 |
| Wildlife | 49.3 | 11.3–522 | |
| Total | 84.3 | 20.5–864 | |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Santos, N.; Richomme, C.; Nunes, T.; Vicente, J.; Alves, P.C.; de la Fuente, J.; Correia-Neves, M.; Boschiroli, M.-L.; Delahay, R.; Gortázar, C. Quantification of the Animal Tuberculosis Multi-Host Community Offers Insights for Control. Pathogens 2020, 9, 421. https://0-doi-org.brum.beds.ac.uk/10.3390/pathogens9060421
Santos N, Richomme C, Nunes T, Vicente J, Alves PC, de la Fuente J, Correia-Neves M, Boschiroli M-L, Delahay R, Gortázar C. Quantification of the Animal Tuberculosis Multi-Host Community Offers Insights for Control. Pathogens. 2020; 9(6):421. https://0-doi-org.brum.beds.ac.uk/10.3390/pathogens9060421
Chicago/Turabian StyleSantos, Nuno, Céline Richomme, Telmo Nunes, Joaquín Vicente, Paulo C. Alves, José de la Fuente, Margarida Correia-Neves, María-Laura Boschiroli, Richard Delahay, and Christian Gortázar. 2020. "Quantification of the Animal Tuberculosis Multi-Host Community Offers Insights for Control" Pathogens 9, no. 6: 421. https://0-doi-org.brum.beds.ac.uk/10.3390/pathogens9060421