Droplet Digital PCR for the Detection of Plasmodium falciparum DNA in Whole Blood and Serum: A Comparative Analysis with Other Molecular Methods

 ,
,  , , ,
, , ,
Abstract
:1. Introduction
2. Results
2.1. Clinical Description
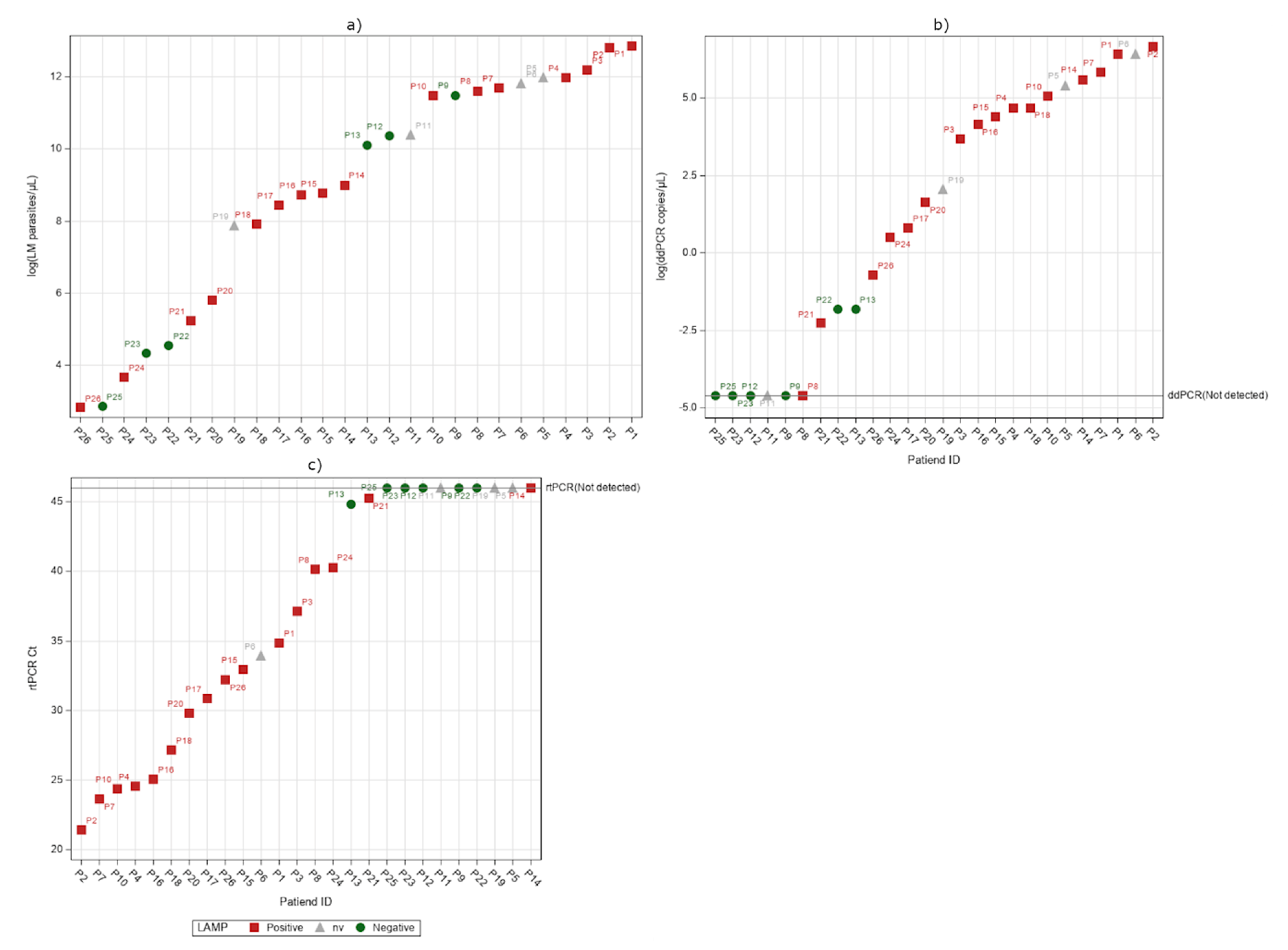
2.2. Limit of Detection Analysis
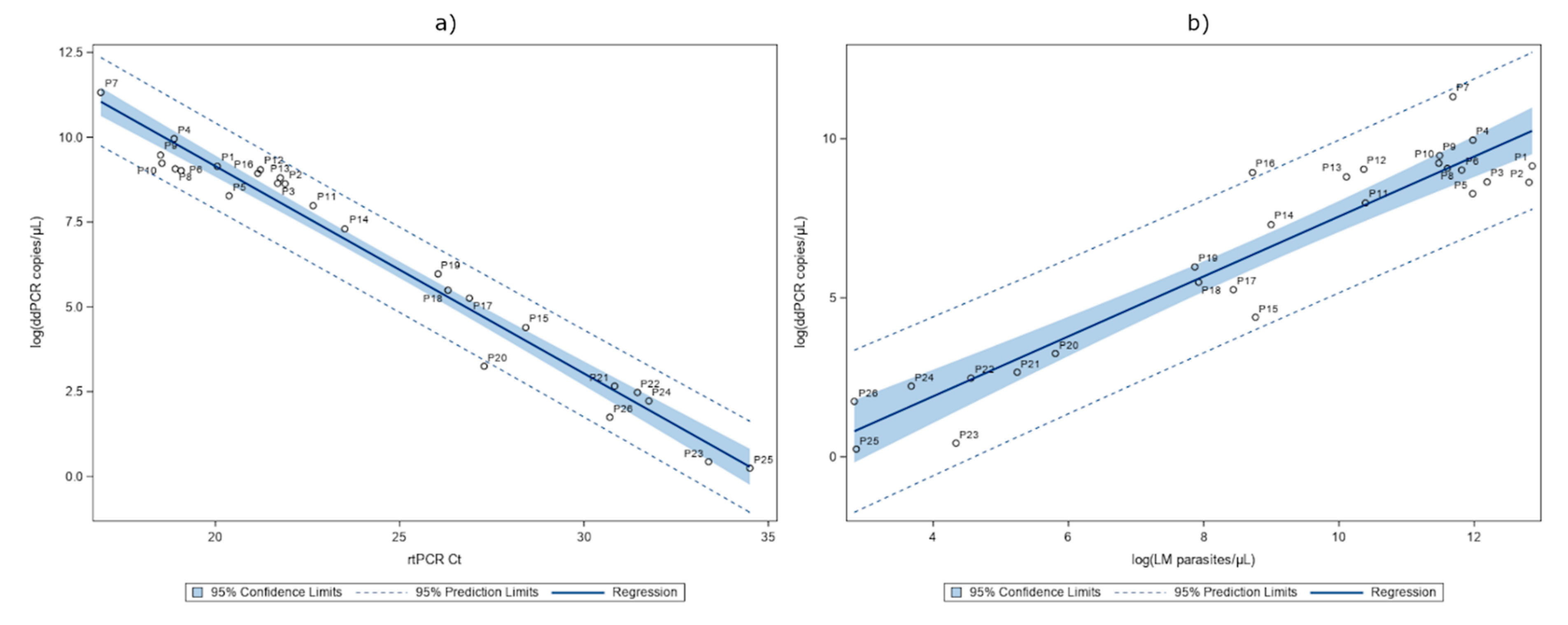
2.3. Detection of P. falciparum DNA in Whole Blood
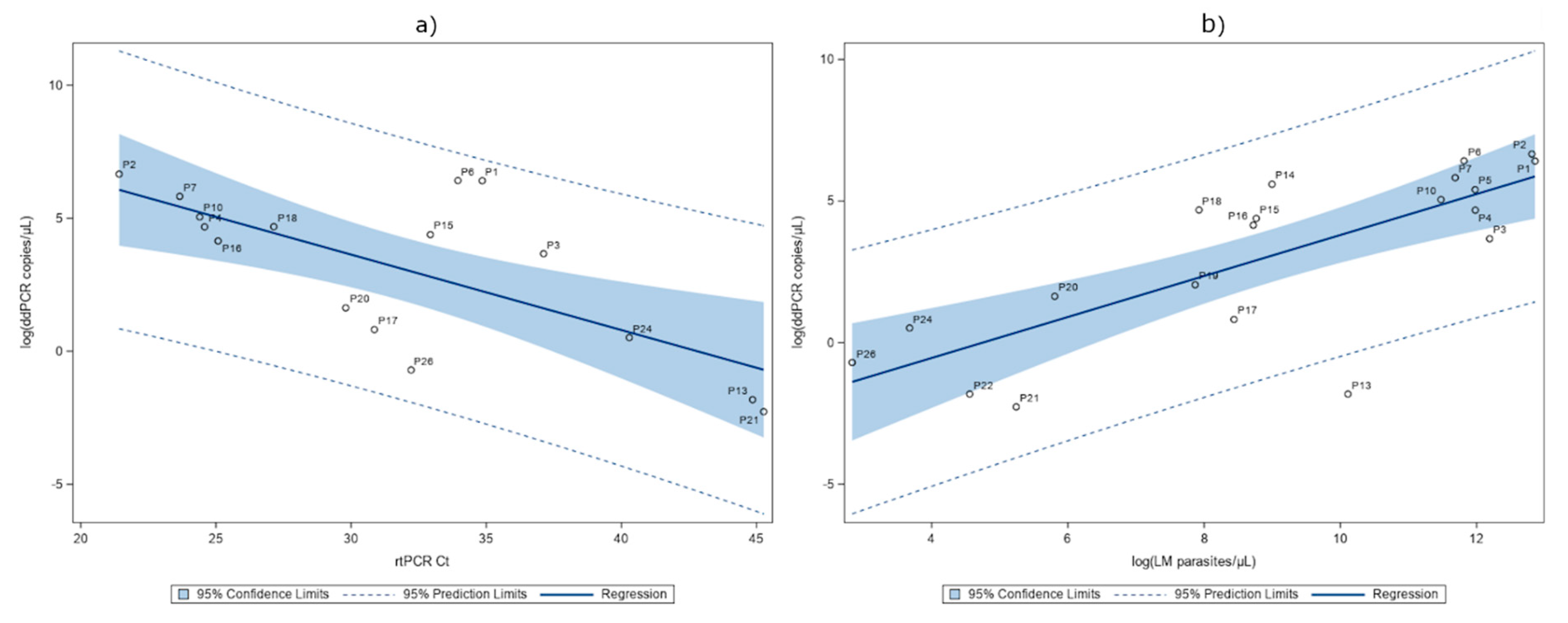
2.4. Detection of P. falciparum DNA in Serum
3. Discussion
4. Materials and Methods
4.1. Setting and Participants
4.2. Diagnostic Analysis
4.3. Analysis of Samples by Microscopy
4.4. Analysis of Samples by RDT
4.5. LAMP
4.6. DNA Extraction
4.7. Real-Time PCR
4.8. ddPCR
4.9. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Carter, R.; Mendis, K.N. Evolutionary and historical aspects of the burden of malaria. Clin. Microbiol. Rev. 2002, 15, 564–594. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Miller, L.H.; Ackerman, H.C.; Su, X.Z.; Wellems, T.E. Malaria biology and disease pathogenesis: Insights for new treatments. Nat. Med. 2013, 19, 156. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- WHO. World Malaria Report 2017; World Health Organization: Geneva, Switzerland, 2017. [Google Scholar]
- Tanner, M.; Greenwood, B.; Whitty, C.J.M.; Ansah, E.K.; Price, R.N.; Dondorp, A.M.; von Seidlein, L.; Baird, J.K.; Beeson, J.G.; Fowkes, F.J.I.; et al. Malaria eradication and elimination: Views on how to translate a vision into reality. BMC Med. 2015, 13, 167. [Google Scholar] [CrossRef] [PubMed]
- Spinazzola, F.; Nicastri, E.; Vlassi, C.; Ghirga, P.; De Marco, M.; Pittalis, S.; Paglia, M.G.; Ferrari, C.; Narciso, P. Imported malaria at Italy’s National Institute for Infectious Diseases Lazzaro Spallanzani, 1984–2003. Eur. J. Clin. Microbiol. Infect. Dis. 2007, 26, 175–179. [Google Scholar] [CrossRef] [PubMed]
- Eikenberry, S.E.; Gumel, A.B. Mathematical modeling of climate change and malaria transmission dynamics: A historical review. J. Math. Biol. 2018, 77, 857–933. [Google Scholar] [CrossRef]
- Rossati, A.; Bargiacchi, O.; Kroumova, V.; Zaramella, M.; Caputo, A.; Garavelli, P.L. Climate, environment and transmission of malaria. Infez. Med. 2016, 24, 93–104. [Google Scholar]
- Ogden, N.; Gachon, P. Climate change and infectious diseases: What can we expect? Can. Commun. Dis. Rep. 2019, 45, 76. [Google Scholar] [CrossRef]
- Stano, P.; Arzese, A.; Merelli, M.; Mascarello, M.; Maurel, C.; Avolio, M.; Bassetti, M.; De Rosa, R.; Luzzati, R.; Modolo, M.L.; et al. Epidemiological and clinical features of imported malaria at the three main hospitals of the Friuli-Venezia Giulia Region, Italy. Infect. Dis. Health 2018, 23, 17–22. [Google Scholar] [CrossRef] [Green Version]
- Herrador, Z.; Fernández-Martinez, B.; Quesada-Cubo, V.; Diaz-Garcia, O.; Cano, R.; Benito, A.; Gómez-Barroso, D. Imported cases of malaria in Spain: Observational study using nationally reported statistics and surveillance data, 2002–2015. Malar. J. 2019, 18, 230. [Google Scholar] [CrossRef]
- Van geertruyden, J.P. Interactions between malaria and human immunodeficiency virus anno 2014. Clin. Microbiol. Infect. 2014, 20, 278–285. [Google Scholar] [CrossRef] [Green Version]
- Mwingira, F.; Genton, B.; Kabanywanyi, A.N.M.; Felger, I. Comparison of detection methods to estimate asexual Plasmodium falciparum parasite prevalence and gametocyte carriage in a community survey in Tanzania. Malar. J. 2014, 13, 433. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Berzosa, P.; De Lucio, A.; Romay-Barja, M.; Herrador, Z.; González, V.; García, L.; Fernández-Martínez, A.; Santana-Morales, M.; Ncogo, P.; Valladares, B.; et al. Comparison of three diagnostic methods (microscopy, RDT, and PCR) for the detection of malaria parasites in representative samples from Equatorial Guinea 11 Medical and Health Sciences 1108 Medical Microbiology. Malar. J. 2018, 17, 333. [Google Scholar] [CrossRef] [PubMed]
- Kotepui, M.; Piwkham, D.; PhunPhuech, B.; Phiwklam, N.; Chupeerach, C.; Duangmano, S. Effects of malaria parasite density on blood cell parameters. PLoS ONE 2015, 10, e0121057. [Google Scholar] [CrossRef] [PubMed]
- Mahende, C.; Ngasala, B.; Lusingu, J.; Yong, T.S.; Lushino, P.; Lemnge, M.; Mmbando, B.; Premji, Z. Performance of rapid diagnostic test, blood-film microscopy and PCR for the diagnosis of malaria infection among febrile children from Korogwe District, Tanzania. Malar. J. 2016, 15, 391. [Google Scholar] [CrossRef] [Green Version]
- Wu, L.; Van Den Hoogen, L.L.; Slater, H.; Walker, P.G.T.; Ghani, A.C.; Drakeley, C.J.; Okell, L.C. Comparison of diagnostics for the detection of asymptomatic Plasmodium falciparum infections to inform control and elimination strategies. Nature 2015, 528, S86. [Google Scholar] [CrossRef] [Green Version]
- Srisutham, S.; Saralamba, N.; Malleret, B.; Rénia, L.; Dondorp, A.M.; Imwong, M. Four human Plasmodium species quantification using droplet digital PCR. PLoS ONE 2017, 12, e0175771. [Google Scholar] [CrossRef]
- Pomari, E.; Piubelli, C.; Perandin, F.B.Z. Digital PCR: A new technology for diagnosis of parasitic infections. Clin. Microbiol. Infect. 2019, 25, 1510–1516. [Google Scholar] [CrossRef]
- De Koninck, A.S.; Cnops, L.; Hofmans, M.; Jacobs, J.; Van Den Bossche, D.; Philippé, J. Diagnostic performance of the loop-mediated isothermal amplification (LAMP) based illumigene® malaria assay in a non-endemic region. Malar. J. 2017, 16, 418. [Google Scholar] [CrossRef] [Green Version]
- Cnops, L.; Boderie, M.; Gillet, P.; Van Esbroeck, M.; Jacobs, J. Rapid diagnostic tests as a source of DNA for Plasmodium species-specific real-time PCR. Malar. J. 2011, 10, 67. [Google Scholar] [CrossRef] [Green Version]
- Tran, T.M.; Aghili, A.; Li, S.; Ongoiba, A.; Kayentao, K.; Doumbo, S.; Traore, B.; Crompton, P.D. A nested real-time PCR assay for the quantification of Plasmodium falciparum DNA extracted from dried blood spots. Malar. J. 2014, 13, 393. [Google Scholar] [CrossRef] [Green Version]
- Gal, S.; Fidler, C.; Turner, S.; Lo, Y.M.; Roberts, D.J.; Wainscoat, J.S. Detection of Plasmodium falciparum DNA in plasma. Ann. N. Y. Acad. Sci. 2001, 945, 234–238. [Google Scholar] [CrossRef] [PubMed]
- Bharti, A.R.; Patra, K.P.; Chuquiyauri, R.; Kosek, M.; Gilman, R.H.; Llanos-Cuentas, A.; Vinetz, J.M. Short report: Polymerase chain reaction detection of Plasmodium vivax and Plasmodium falciparum DNA from stored serum samples: Implications for retrospective diagnosis of malaria. Am. J. Trop. Med. Hyg. 2007, 77, 444–446. [Google Scholar] [CrossRef] [PubMed]
- Lamikanra, A.A.; Dobão, C.; Jiménez, A.; Nhabomba, A.; Tsang, H.P.; Guinovart, C.; Manaca, M.N.; Quinto, L.; Aguilar, R.; Cisterá, P.; et al. A direct comparison of real time PCR on plasma and blood to detect Plasmodium falciparum infection in children. Malar. J. 2012, 11, 201. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Imwong, M.; Woodrow, C.J.; Hendriksen, I.C.E.; Veenemans, J.; Verhoef, H.; Faiz, M.A.; Mohanty, S.; Mishra, S.; Mtove, G.; Gesase, S.; et al. Plasma concentration of parasite DNA as a measure of disease severity in falciparum malaria. J. Infect. Dis. 2015, 211, 1128–1133. [Google Scholar] [CrossRef] [Green Version]
- Koepfli, C.; Nguitragool, W.; Hofmann, N.E.; Robinson, L.J.; Ome-Kaius, M.; Sattabongkot, J.; Felger, I.; Mueller, I. Sensitive and accurate quantification of human malaria parasites using droplet digital PCR (ddPCR). Sci. Rep. 2016, 6, 39183. [Google Scholar] [CrossRef] [Green Version]
- Costa, F.T.M.; Avril, M.; Nogueira, P.A.; Gysin, J. Cytoadhesion of Plasmodium falciparum-infected erythrocytes and the infected placenta: A two-way pathway. Braz. J. Med. Biol. Res. 2006, 39, 1525–1536. [Google Scholar] [CrossRef]
- Wampfler, R.; Mwingira, F.; Javati, S.; Robinson, L.; Betuela, I.; Siba, P.; Beck, H.P.; Mueller, I.; Felger, I. Strategies for Detection of Plasmodium species Gametocytes. PLoS ONE 2013, 8, e76316. [Google Scholar] [CrossRef] [Green Version]
- Vafa Homann, M.; Emami, S.N.; Yman, V.; Stenström, C.; Sondén, K.; Ramström, H.; Karlsson, M.; Asghar, M.; Färnert, A. Detection of Malaria Parasites After Treatment in Travelers: A 12-months Longitudinal Study and Statistical Modelling Analysis. EBioMedicine 2017, 25, 66–72. [Google Scholar] [CrossRef] [Green Version]
- White, N.J.; Turner, G.D.H.; Day, N.P.J.; Dondorp, A.M. Lethal Malaria: Marchiafava and Bignami Were Right. J. Infect. Dis. 2013, 208, 192–198. [Google Scholar] [CrossRef]
- White, N.J. Malaria parasite clearance. Malar. J. 2017, 16, 88. [Google Scholar] [CrossRef] [Green Version]
- Franke-Fayard, B.; Fonager, J.; Braks, A.; Khan, S.M.; Janse, C.J. Sequestration and tissue accumulation of human malaria parasites: Can we learn anything from rodent models of malaria? PLoS Pathog. 2010, 6, e1001032. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Perandin, F.; Manca, N.; Calderaro, A.; Piccolo, G.; Galati, L.; Ricci, L.; Medici, M.C.; Arcangeletti, M.C.; Snounou, G.; Dettori, G.; et al. Development of a Real-Time PCR Assay for Detection of Plasmodium falciparum, Plasmodium vivax, and Plasmodium ovale for Routine Clinical Diagnosis. J. Clin. Microbiol. 2004, 42, 1214–1219. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pan, X.B.; Han, J.C.; Cong, X.; Wei, L. BST2/Tetherin Inhibits Dengue Virus Release from Human Hepatoma Cells. PLoS ONE 2012, 7, e51033. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
| Patients with P. falciparum Malaria N (%) or Median(IQR) | |
|---|---|
| Patients | 26 (100) |
Country of origin | Country of exposition |
| Sex M | 20 (76.92) |
| Age by sex Male: Female: | 49 (37–58) 37 (26–38) |
| Parasitaemia (trophozoites/μL of blood) at diagnosis | From 438,900 to 17 |
| Microscopy identification P. falciparum: | 26 (100) |
Plasmodium antigen test result (BinaxNOW® Malaria) | |
| LAMP (results on blood) Plasmodium: | 26 (100) |
| Severe clinical manifestations | 3 (11.54) |
| Pregnancy | 1 (3.85) |
| Patient ID | Trophozoites/μL Blood | %iRBCs | LAMP Wholeblood | LAMP Serum | rtPCR (Ct) Wholeblood | rtPCR (Ct) Serum | ddPCR (Copies/μL Blood) Wholeblood | ddPCR (Copies/μL Serum) Serum |
|---|---|---|---|---|---|---|---|---|
| P1 | 383,460 | 9.15 | Positive | Positive | 20.05 | 34.85 | 9350 | 608.178 |
| P2 | 366,000 | 9.5 | Positive | Positive | 21.89 | 21.42 | 5588 | 780.267 |
| P3 | 196,930 | 4.7 | Positive | Positive | 21.69 | 37.12 | 5654 | 39.111 |
| P4 | 159,544 | 2.96 | Positive | Positive | 18.88 | 24.58 | 21,109 | 107.36 |
| P5 | 159,120 | 5.2 | Positive | nv | 20.37 | nd | 3916 | 221.467 |
| P6 | 135,135 | 3.85 | Positive | nv | 19.07 | 33.95 | 8228 | 611.6 |
| P7 | 118,800 | 3.37 | Positive | Positive | 16.89 | 23.66 | 82,500 | 337.92 |
| P8 | 109,470 | 2.05 | Positive | Positive | 18.91 | 40.16 | 8668 | nd |
| P9 | 97,600 | 2.44 | Positive | Negative | 18.51 | nd | 13,002 | nd |
| P10 | 96,400 | 2.41 | Positive | Positive | 18.55 | 24.4 | 10,230 | 155.76 |
| P11 | 32,560 | 0.55 | Positive | nv | 22.65 | nd | 2926 | nd |
| P12 | 31,790 | 0.85 | Positive | Negative | 21.22 | nd | 8448 | nd |
| P13 | 24,610 | 1.15 | Positive | Negative | 21.76 | 44.85 | 6644 | 0.152 |
| P14 | 8076 | 0.184 | Positive | Positive | 23.51 | nd | 1476.2 | 268.4 |
| P15 | 6409 | 0.15 | Positive | Positive | 28.42 | 32.93 | 80.52 | 80.178 |
| P16 | 6133.5 | 0.1557 | Positive | Positive | 21.15 | 25.08 | 7601 | 63.184 |
| P17 | 4620 | 0.092 | Positive | Positive | 26.89 | 30.86 | 191.4 | 2.249 |
| P18 | 2764 | 0.0507 | Positive | Positive | 26.31 | 27.14 | 242 | 108.044 |
| P19 | 2613 | 0.0549 | Positive | nv | 26.04 | nd | 391.6 | 7.676 |
| P20 | 332.5 | 0.0088 | Positive | Positive | 27.29 | 29.8 | 25.74 | 5.104 |
| P21 | 189 | 0.0043 | Positive | Positive | 30.83 | 45.26 | 14.3 | 0.093 |
| P22 | 95.4 | 0.0023 | Positive | Negative | 31.45 | nd | 11.88 | 0.152 |
| P23 | 76.5 | 0.0014 | Positive | Negative | 33.38 | nd | 1.54 | nd |
| P24 | 39.5 | 0.0008 | Positive | Positive | 31.76 | 40.29 | 9.24 | 1.662 |
| P25 | 17.5 | 0.0003 | Positive | Negative | 34.5 | nd | 1.276 | nd |
| P26 | 17 | 0.0004 | Positive | Positive | 30.7 | 32.22 | 5.72 | 0.484 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pomari, E.; Silva, R.; Moro, L.; La Marca, G.; Perandin, F.; Verra, F.; Bisoffi, Z.; Piubelli, C. Droplet Digital PCR for the Detection of Plasmodium falciparum DNA in Whole Blood and Serum: A Comparative Analysis with Other Molecular Methods. Pathogens 2020, 9, 478. https://0-doi-org.brum.beds.ac.uk/10.3390/pathogens9060478
Pomari E, Silva R, Moro L, La Marca G, Perandin F, Verra F, Bisoffi Z, Piubelli C. Droplet Digital PCR for the Detection of Plasmodium falciparum DNA in Whole Blood and Serum: A Comparative Analysis with Other Molecular Methods. Pathogens. 2020; 9(6):478. https://0-doi-org.brum.beds.ac.uk/10.3390/pathogens9060478
Chicago/Turabian StylePomari, Elena, Ronaldo Silva, Lucia Moro, Giulia La Marca, Francesca Perandin, Federica Verra, Zeno Bisoffi, and Chiara Piubelli. 2020. "Droplet Digital PCR for the Detection of Plasmodium falciparum DNA in Whole Blood and Serum: A Comparative Analysis with Other Molecular Methods" Pathogens 9, no. 6: 478. https://0-doi-org.brum.beds.ac.uk/10.3390/pathogens9060478