The Basement Membrane in a 3D Breast Acini Model Modulates Delivery and Anti-Proliferative Effects of Liposomal Anthracyclines

 , , and
, , and
Abstract
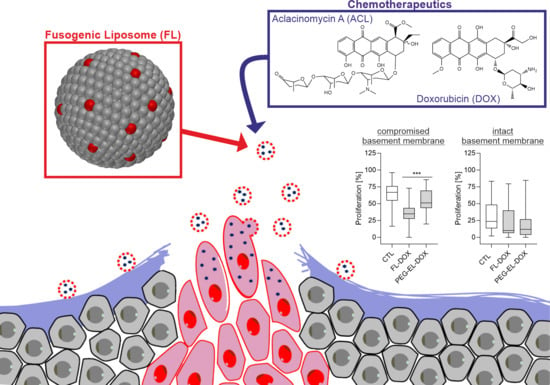
:
1. Introduction
2. Results
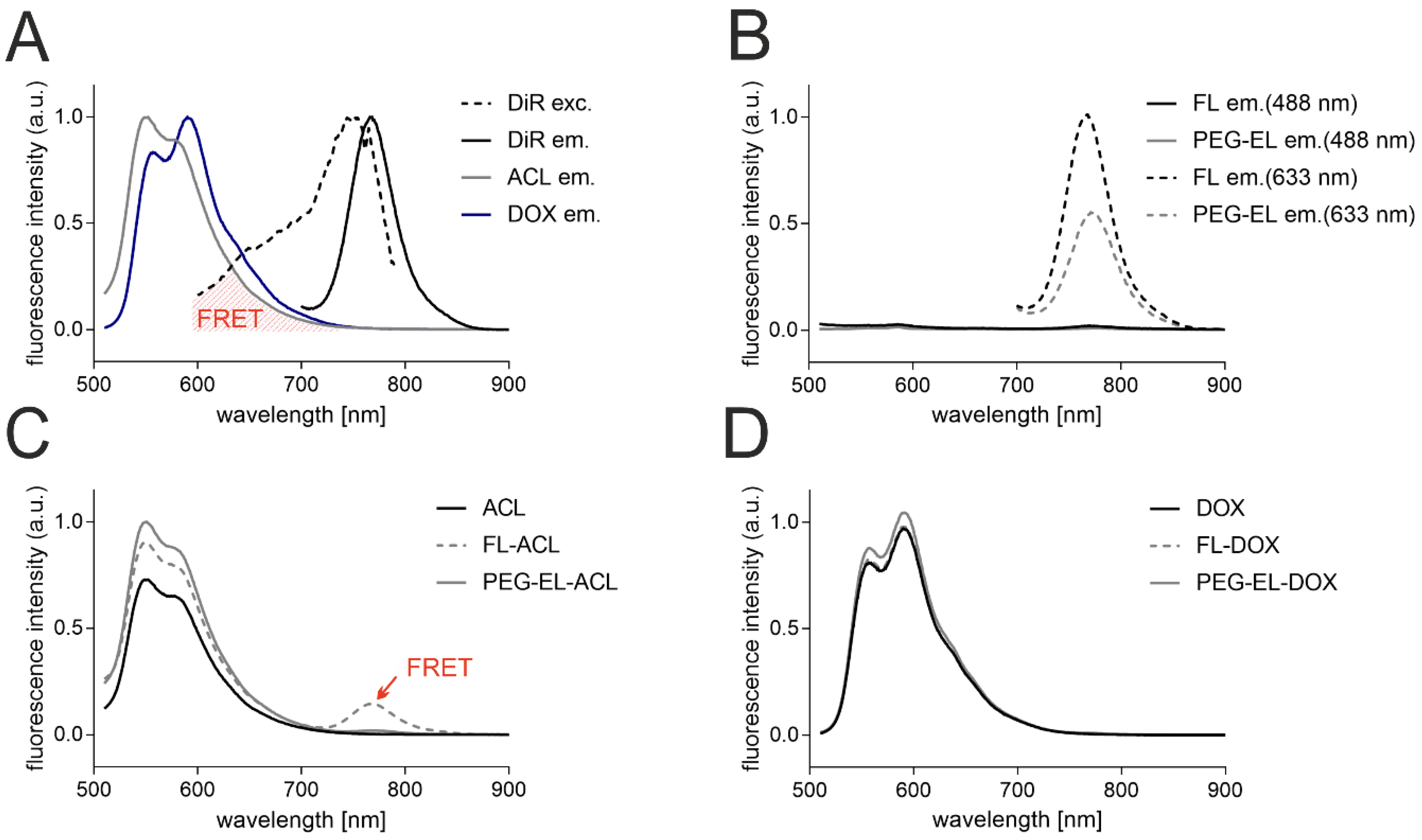
2.1. Characterization of FL and PEG-EL Loaded with ACL and DOX
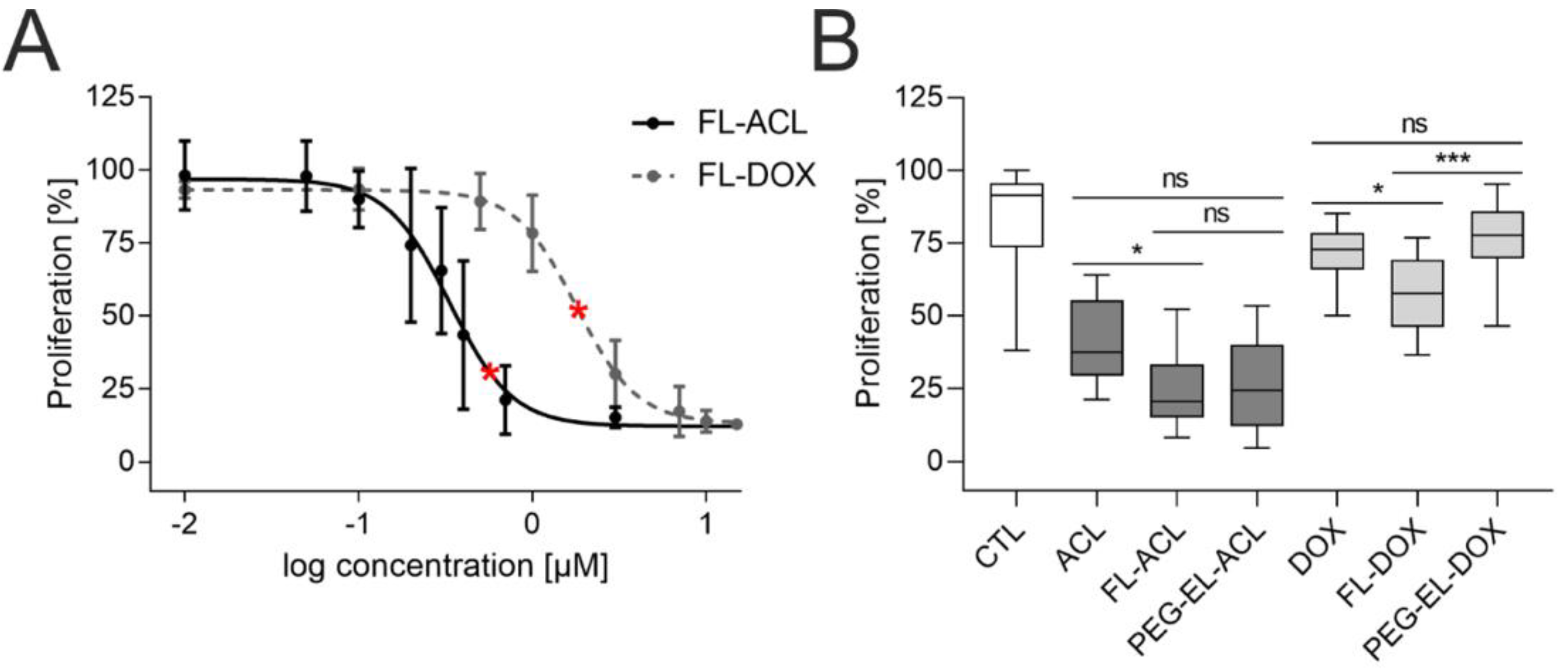
2.2. Uptake Efficiency of ACL and DOX into MCF-10A Cells in 2D Cell Culture
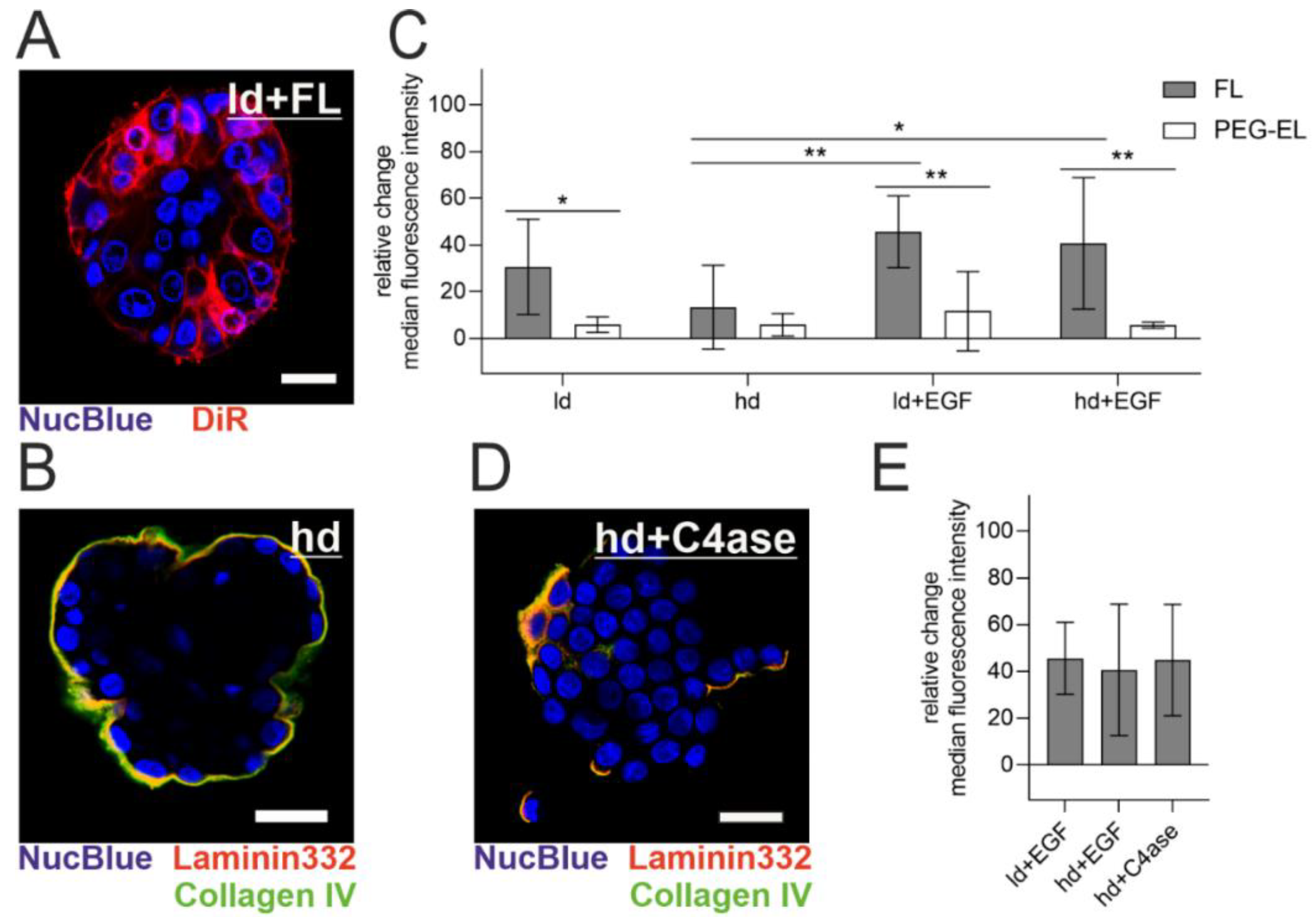
2.3. Influence of Basement Membrane Integrity on Liposomal Uptake Efficiency
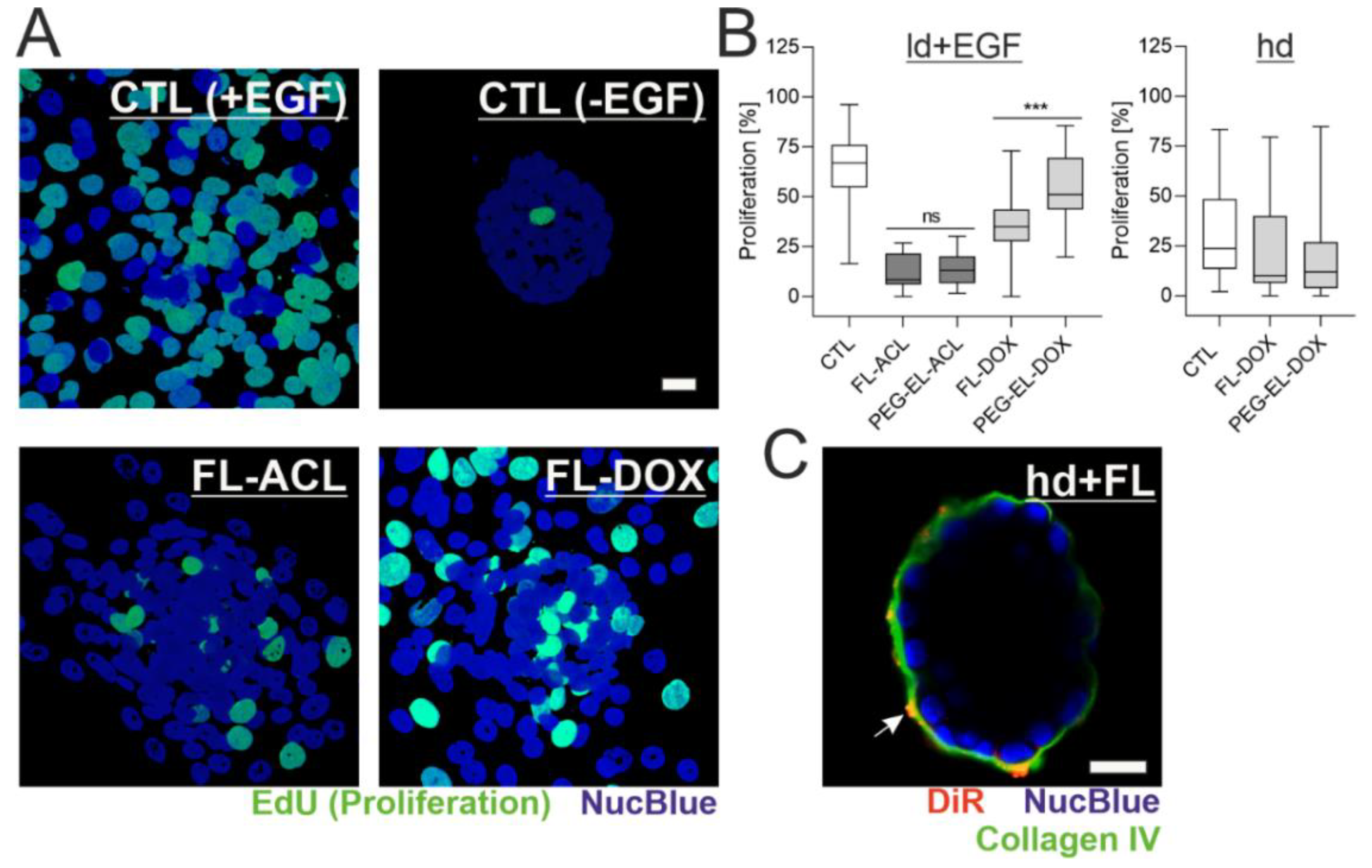
2.4. The Anti-Proliferative Effect of DOX in 3D Breast Acini Models Depends on the Basement Membrane Status
3. Discussion
4. Material and Methods
4.1. Preparation of Fusogenic and Endocytic Liposomes
4.2. Characterization of Liposomes
4.3. Cell Maintenance and Cultivation of MCF-10A Acini
4.4. Liposomal Treatment and Imaging of Cellular Uptake in 2D
4.5. Measurement of the Anti-Proliferative Effects of the Anthracyclines DOX and ACL
4.5.1. Cell Counting Using Flow Cytometry
4.5.2. EdU (5-ethynyl-2′-deoxyuridine) Incorporation Assay
4.6. Evaluation of Membrane Tracer Dye DiR Uptake in Cells of the MCF-10A Acini Model
4.7. Data Presentation and Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
Abbreviations
References
- Zielonke, N.; Gini, A.; Jansen, E.E.L.; Anttila, A.; Segnan, N.; Ponti, A.; Veerus, P.; de Koning, H.J.; van Ravesteyn, N.T.; Heijnsdijk, E.A.M. Evidence for reducing cancer-specific mortality due to screening for breast cancer in Europe: A systematic review. Eur. J. Cancer (Oxf. Engl. 1990) 2020, 127, 191–206. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dafni, U.; Tsourti, Z.; Alatsathianos, I. Breast Cancer Statistics in the European Union: Incidence and Survival across European Countries. Breast Care 2019, 14, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Commission, E. ECIS—European Cancer Information System. Available online: https://ecis.jrc.ec.europa.eu/ (accessed on 17 June 2020).
- Peto, R.; Davies, C.; Godwin, J.; Gray, R.; Pan, H.C.; Clarke, M.; Cutter, D.; Darby, S.; McGale, P.; Taylor, C.; et al. Comparisons between different polychemotherapy regimens for early breast cancer: Meta-analyses of long-term outcome among 100,000 women in 123 randomised trials. Lancet (Lond. Engl.) 2012, 379, 432–444. [Google Scholar] [CrossRef] [Green Version]
- Tao, J.J.; Visvanathan, K.; Wolff, A.C. Long term side effects of adjuvant chemotherapy in patients with early breast cancer. Breast 2015, 24 (Suppl. 2), S149–S153. [Google Scholar] [CrossRef] [Green Version]
- Jensen, B.V.; Skovsgaard, T.; Nielsen, S.L. Functional monitoring of anthracycline cardiotoxicity:a prospective, blinded, long-term observational study of outcome in 120 patients. Ann. Oncol. 2002, 13, 699–709. [Google Scholar] [CrossRef]
- Gabizon, A.; Catane, R.; Uziely, B.; Kaufman, B.; Safra, T.; Cohen, R.; Martin, F.; Huang, A.; Barenholz, Y. Prolonged circulation time and enhanced accumulation in malignant exudates of doxorubicin encapsulated in polyethylene-glycol coated liposomes. Cancer Res. 1994, 54, 987–992. [Google Scholar]
- Barenholz, Y. Doxil®—The first FDA-approved nano-drug: Lessons learned. J. Control. Release Off. J. Control. Release Soc. 2012, 160, 117–134. [Google Scholar] [CrossRef]
- Chanan-Khan, A.; Szebeni, J.; Savay, S.; Liebes, L.; Rafique, N.M.; Alving, C.R.; Muggia, F.M. Complement activation following first exposure to pegylated liposomal doxorubicin (Doxil®): Possible role in hypersensitivity reactions. Ann. Oncol. 2003, 14, 1430–1437. [Google Scholar] [CrossRef]
- Patel, J.; Ringley, J.T.; Moore, D.C. Case series of docetaxel-induced dorsal hand-foot syndrome. Adv. Drug Saf. 2018, 9, 495–498. [Google Scholar] [CrossRef] [Green Version]
- Mitra, S.; Gaur, U.; Ghosh, P.C.; Maitra, A.N. Tumour targeted delivery of encapsulated dextran-doxorubicin conjugate using chitosan nanoparticles as carrier. J. Control. Release Off. J. Control. Release Soc. 2001, 74, 317–323. [Google Scholar] [CrossRef]
- Lee, C.C.; Gillies, E.R.; Fox, M.E.; Guillaudeu, S.J.; Fréchet, J.M.; Dy, E.E.; Szoka, F.C. A single dose of doxorubicin-functionalized bow-tie dendrimer cures mice bearing C-26 colon carcinomas. Proc. Natl. Acad. Sci. USA 2006, 103, 16649–16654. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, Y.; Yang, C.; Wang, W.; Liu, J.; Liu, Q.; Huang, F.; Chu, L.; Gao, H.; Li, C.; Kong, D.; et al. Co-delivery of doxorubicin and curcumin by pH-sensitive prodrug nanoparticle for combination therapy of cancer. Sci. Rep. 2016, 6, 21225. [Google Scholar] [CrossRef] [Green Version]
- Hadla, M.; Palazzolo, S.; Corona, G.; Caligiuri, I.; Canzonieri, V.; Toffoli, G.; Rizzolio, F. Exosomes increase the therapeutic index of doxorubicin in breast and ovarian cancer mouse models. Nanomedicine 2016, 11, 2431–2441. [Google Scholar] [CrossRef]
- Bagalkot, V.; Farokhzad, O.C.; Langer, R.; Jon, S. An aptamer-doxorubicin physical conjugate as a novel targeted drug-delivery platform. Angew. Chem. (Int. Ed. Engl.) 2006, 45, 8149–8152. [Google Scholar] [CrossRef] [PubMed]
- Kleusch, C.; Hersch, N.; Hoffmann, B.; Merkel, R.; Csiszár, A. Fluorescent lipids: Functional parts of fusogenic liposomes and tools for cell membrane labeling and visualization. Molecules 2012, 17, 1055–1073. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hoffmann, M.; Hersch, N.; Merkel, R.; Csiszar, A.; Hoffmann, B. Changing the Way of Entrance: Highly Efficient Transfer of mRNA and siRNA via Fusogenic Nano-Carriers. J. Biomed. Nanotechnol. 2019, 15, 170–183. [Google Scholar] [CrossRef] [PubMed]
- Csiszar, A.; Hersch, N.; Dieluweit, S.; Biehl, R.; Merkel, R.; Hoffmann, B. Novel fusogenic liposomes for fluorescent cell labeling and membrane modification. Bioconjug Chem. 2010, 21, 537–543. [Google Scholar] [CrossRef]
- Kolašinac, R.; Kleusch, C.; Braun, T.; Merkel, R.; Csiszár, A. Deciphering the Functional Composition of Fusogenic Liposomes. Int. J. Mol. Sci. 2018, 19, 346. [Google Scholar] [CrossRef] [Green Version]
- Kube, S.; Hersch, N.; Naumovska, E.; Gensch, T.; Hendriks, J.; Franzen, A.; Landvogt, L.; Siebrasse, J.P.; Kubitscheck, U.; Hoffmann, B.; et al. Fusogenic Liposomes as Nanocarriers for the Delivery of Intracellular Proteins. Langmuir 2017, 33, 1051–1059. [Google Scholar] [CrossRef]
- Wiedenhoeft, T.; Tarantini, S.; Nyúl-Tóth, Á.; Yabluchanskiy, A.; Csipo, T.; Balasubramanian, P.; Lipecz, A.; Kiss, T.; Csiszar, A.; Csiszar, A.; et al. Fusogenic liposomes effectively deliver resveratrol to the cerebral microcirculation and improve endothelium-dependent neurovascular coupling responses in aged mice. Geroscience 2019, 41, 711–725. [Google Scholar] [CrossRef]
- Manzanares, D.; Ceña, V. Endocytosis: The Nanoparticle and Submicron Nanocompounds Gateway into the Cell. Pharmaceutics 2020, 12, 371. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bulbake, U.; Doppalapudi, S.; Kommineni, N.; Khan, W. Liposomal Formulations in Clinical Use: An Updated Review. Pharmaceutics 2017, 9, 12. [Google Scholar] [CrossRef] [PubMed]
- Lipponen, P.; Ji, H.; Aaltomaa, S.; Syrjänen, K. Tumour vascularity and basement membrane structure in breast cancer as related to tumour histology and prognosis. J. Cancer Res. Clin. Oncol. 1994, 120, 645–650. [Google Scholar] [CrossRef] [PubMed]
- Perche, F.; Torchilin, V.P. Cancer cell spheroids as a model to evaluate chemotherapy protocols. Cancer Biol. Ther. 2012, 13, 1205–1213. [Google Scholar] [CrossRef]
- Paškevičiūtė, M.; Petrikaitė, V. Proton Pump Inhibitors Modulate Transport of Doxorubicin and Its Liposomal Form into 2D and 3D Breast Cancer Cell Cultures. Cancer Manag. Res. 2019, 11, 9761–9769. [Google Scholar] [CrossRef] [Green Version]
- Paškevičiūtė, M.; Petrikaitė, V. Application of carbonic anhydrase inhibitors to increase the penetration of doxorubicin and its liposomal formulation into 2D and 3D triple negative breast cancer cell cultures. Am. J. Cancer Res. 2020, 10, 1761–1769. [Google Scholar]
- Gabizon, A.; Shmeeda, H.; Barenholz, Y. Pharmacokinetics of pegylated liposomal Doxorubicin: Review of animal and human studies. Clin. Pharm. 2003, 42, 419–436. [Google Scholar] [CrossRef]
- Friche, E.; Jensen, P.B.; Roed, H.; Skovsgaard, T.; Nissen, N.I. In vitro circumvention of anthracycline--resistance in Ehrlich ascites tumour by anthracycline analogues. Biochem. Pharm. 1990, 39, 1721–1726. [Google Scholar] [CrossRef]
- Gaiko-Shcherbak, A.; Fabris, G.; Dreissen, G.; Merkel, R.; Hoffmann, B.; Noetzel, E. The Acinar Cage: Basement Membranes Determine Molecule Exchange and Mechanical Stability of Human Breast Cell Acini. PLoS ONE 2015, 10, e0145174. [Google Scholar] [CrossRef] [Green Version]
- Fabris, G.; Lucantonio, A.; Hampe, N.; Noetzel, E.; Hoffmann, B.; DeSimone, A.; Merkel, R. Nanoscale Topography and Poroelastic Properties of Model Tissue Breast Gland Basement Membranes. Biophys. J. 2018, 115, 1770–1782. [Google Scholar] [CrossRef] [Green Version]
- Rivankar, S. An overview of doxorubicin formulations in cancer therapy. J. Cancer Res. Ther. 2014, 10, 853–858. [Google Scholar] [CrossRef] [PubMed]
- Wei, G.; Ni, W.; Chiao, J.W.; Cai, Z.; Huang, H.; Liu, D. A meta-analysis of CAG (cytarabine, aclarubicin, G-CSF) regimen for the treatment of 1029 patients with acute myeloid leukemia and myelodysplastic syndrome. J. Hematol. Oncol. 2011, 4, 46. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Binaschi, M.; Farinosi, R.; Austin, C.A.; Fisher, L.M.; Zunino, F.; Capranico, G. Human DNA topoisomerase IIalpha-dependent DNA cleavage and yeast cell killing by anthracycline analogues. Cancer Res. 1998, 58, 1886–1892. [Google Scholar] [PubMed]
- Doroshow, J.H. Effect of anthracycline antibiotics on oxygen radical formation in rat heart. Cancer Res. 1983, 43, 460–472. [Google Scholar] [PubMed]
- Sato, S.; Iwaizumi, M.; Handa, K.; Tamura, Y. Electron spin resonance study on the mode of generation of free radicals of daunomycin, adriamycin, and carboquone in NAD(P)H-microsome system. GANN Jpn. J. Cancer Res. 1977, 68, 603–608. [Google Scholar]
- Dong, J.; Naito, M.; Tatsuta, T.; Seimiya, H.; Johdo, O.; Tsuruo, T. Difference between the resistance mechanisms of aclacinomycin- and adriamycin-resistant P388 cell lines. Oncol. Res. 1995, 7, 245–252. [Google Scholar]
- McGowan, J.V.; Chung, R.; Maulik, A.; Piotrowska, I.; Walker, J.M.; Yellon, D.M. Anthracycline Chemotherapy and Cardiotoxicity. Cardiovasc. Drugs 2017, 31, 63–75. [Google Scholar] [CrossRef] [Green Version]
- Eckardt, J.R.; Campbell, E.; Burris, H.A.; Weiss, G.R.; Rodriguez, G.I.; Fields, S.M.; Thurman, A.M.; Peacock, N.W.; Cobb, P.; Rothenberg, M.L.; et al. A phase II trial of DaunoXome, liposome-encapsulated daunorubicin, in patients with metastatic adenocarcinoma of the colon. Am. J. Clin. Oncol. 1994, 17, 498–501. [Google Scholar] [CrossRef]
- Batist, G.; Barton, J.; Chaikin, P.; Swenson, C.; Welles, L. Myocet (liposome-encapsulated doxorubicin citrate): A new approach in breast cancer therapy. Expert Opin. Pharmacother. 2002, 3, 1739–1751. [Google Scholar] [CrossRef]
- Förster, T. Zwischenmolekulare Energiewanderung und Fluoreszenz. Ann. Der. Phys. 1948, 437, 55–75. [Google Scholar] [CrossRef]
- Stryer, L.; Haugland, R.P. Energy transfer: A spectroscopic ruler. Proc. Natl. Acad. Sci. USA 1967, 58, 719–726. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kolašinac, R.; Jaksch, S.; Dreissen, G.; Braeutigam, A.; Merkel, R.; Csiszár, A. Influence of Environmental Conditions on the Fusion of Cationic Liposomes with Living Mammalian Cells. Nanomaterials 2019, 9, 1025. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zenebergh, A.; Baurain, R.; Trouet, A. Cellular pharmacokinetics of aclacinomycin A in cultured L1210 cells. Comparison with daunorubicin and doxorubicin. Cancer Chemother. Pharmacol. 1982, 8, 243–249. [Google Scholar] [CrossRef] [PubMed]
- Ndolo, R.A.; Luan, Y.; Duan, S.; Forrest, M.L.; Krise, J.P. Lysosomotropic Properties of Weakly Basic Anticancer Agents Promote Cancer Cell Selectivity In Vitro. PLoS ONE 2012, 7, e49366. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gatseva, A.; Sin, Y.Y.; Brezzo, G.; Van Agtmael, T. Basement membrane collagens and disease mechanisms. Essays Biochem. 2019, 63, 297–312. [Google Scholar] [CrossRef] [Green Version]
- Bissell, M.J.; Radisky, D. Putting tumours in context. Nat. Rev. Cancer 2001, 1, 46–54. [Google Scholar] [CrossRef] [Green Version]
- Bergamaschi, A.; Tagliabue, E.; Sørlie, T.; Naume, B.; Triulzi, T.; Orlandi, R.; Russnes, H.; Nesland, J.; Tammi, R.; Auvinen, P.; et al. Extracellular matrix signature identifies breast cancer subgroups with different clinical outcome. J. Pathol. 2008, 214, 357–367. [Google Scholar] [CrossRef]
- Yurchenco, P.D.; Cheng, Y.S.; Colognato, H. Laminin forms an independent network in basement membranes. J. Cell Biol. 1992, 117, 1119–1133. [Google Scholar] [CrossRef] [Green Version]
- Harvey, S.J.; Jarad, G.; Cunningham, J.; Rops, A.L.; van der Vlag, J.; Berden, J.H.; Moeller, M.J.; Holzman, L.B.; Burgess, R.W.; Miner, J.H. Disruption of glomerular basement membrane charge through podocyte-specific mutation of agrin does not alter glomerular permselectivity. Am. J. Pathol. 2007, 171, 139–152. [Google Scholar] [CrossRef] [Green Version]
- Goode, N.P.; Shires, M.; Davison, A.M. The glomerular basement membrane charge-selectivity barrier: An oversimplified concept? Nephrol. Dial. Transpl. 1996, 11, 1714–1716. [Google Scholar] [CrossRef]
- Woodle, M.C.; Newman, M.S.; Martin, F.J. Liposome leakage and blood circulation: Comparison of adsorbed block copolymers with covalent attachment of PEG. Int. J. Pharm. 1992, 88, 327–334. [Google Scholar] [CrossRef]
- Ota, Z.; Makino, H.; Takaya, Y.; Ofuji, T. Molecular sieve in renal glomerular and tubular basement membranes as revealed by electron microscopy. Ren. Physiol. 1980, 3, 317–323. [Google Scholar] [CrossRef]
- Persky, B.; Grganto, D.M. Diffusion of dextrans and microspheres in the human amniotic basement membrane model. Clin. Exp. Metastasis 1987, 5, 321–328. [Google Scholar] [CrossRef] [PubMed]
- Albrechtsen, R.; Nielsen, M.; Wewer, U.; Engvall, E.; Ruoslahti, E. Basement membrane changes in breast cancer detected by immunohistochemical staining for laminin. Cancer Res. 1981, 41, 5076–5081. [Google Scholar] [PubMed]
- Farid, M.; Faber, T.; Dietrich, D.; Lamprecht, A. Cell membrane fusing liposomes for cytoplasmic delivery in brain endothelial cells. Colloids Surf. B Biointerfaces 2020, 194, 111193. [Google Scholar] [CrossRef]
- Schindelin, J.; Arganda-Carreras, I.; Frise, E.; Kaynig, V.; Longair, M.; Pietzsch, T.; Preibisch, S.; Rueden, C.; Saalfeld, S.; Schmid, B.; et al. Fiji: An open-source platform for biological-image analysis. Nat. Methods 2012, 9, 676–682. [Google Scholar] [CrossRef] [PubMed] [Green Version]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Size (nm) | ζ Potential (mV) | EE (%) | |
|---|---|---|---|
| FL | 116 ± 45 | 70 ± 6 | - |
| FL-ACL | 87 ± 14 | 64 ± 25 | 92 ± 6 |
| FL-DOX | 85 ± 4 | 69 ± 5 | 88 ± 6 |
| PEG-EL | 163 ± 78 | −29 ± 18 | - |
| PEG-EL-ACL | 152 ± 46 | −47 ± 5 | 89 ± 4 |
| PEG-EL-DOX | 142 ± 44 | −48 ± 10 | 90 ± 6 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wiedenhoeft, T.; Braun, T.; Springer, R.; Teske, M.; Noetzel, E.; Merkel, R.; Csiszár, A. The Basement Membrane in a 3D Breast Acini Model Modulates Delivery and Anti-Proliferative Effects of Liposomal Anthracyclines. Pharmaceuticals 2020, 13, 256. https://0-doi-org.brum.beds.ac.uk/10.3390/ph13090256
Wiedenhoeft T, Braun T, Springer R, Teske M, Noetzel E, Merkel R, Csiszár A. The Basement Membrane in a 3D Breast Acini Model Modulates Delivery and Anti-Proliferative Effects of Liposomal Anthracyclines. Pharmaceuticals. 2020; 13(9):256. https://0-doi-org.brum.beds.ac.uk/10.3390/ph13090256
Chicago/Turabian StyleWiedenhoeft, Tabea, Tobias Braun, Ronald Springer, Michael Teske, Erik Noetzel, Rudolf Merkel, and Agnes Csiszár. 2020. "The Basement Membrane in a 3D Breast Acini Model Modulates Delivery and Anti-Proliferative Effects of Liposomal Anthracyclines" Pharmaceuticals 13, no. 9: 256. https://0-doi-org.brum.beds.ac.uk/10.3390/ph13090256