Fragaria viridis Fruit Metabolites: Variation of LC-MS Profile and Antioxidant Potential during Ripening and Storage



Abstract
:1. Introduction
2. Results and Discussion
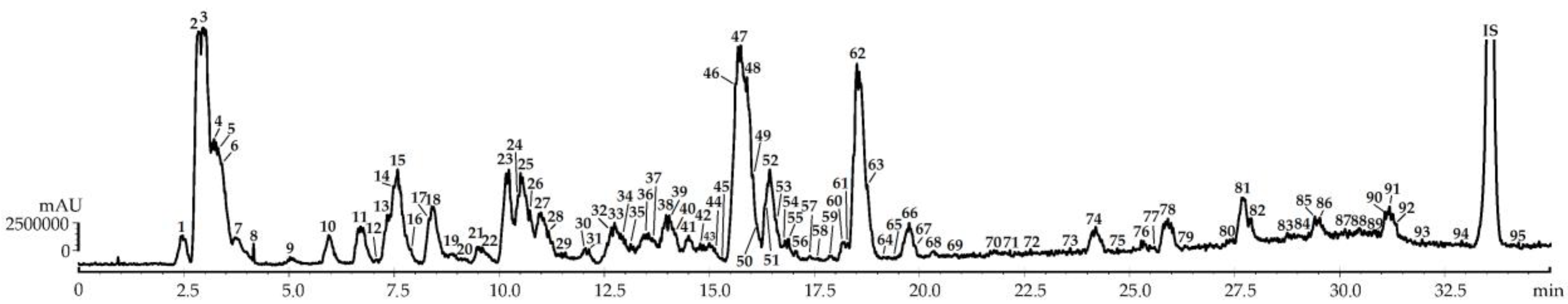
2.1. Metabolites of F. viridis Fruits: LS-MS Profile
2.1.1. Carbohydrates
2.1.2. Organic Acids
2.1.3. Gallic Acid Derivatives
2.1.4. Ellagic Acid Derivatives and Ellagitannins
2.1.5. Hydroxycinnamates and Coumarins
2.1.6. Catechins and Procyanidins
2.1.7. Anthocyanins
2.1.8. Flavonols
2.1.9. Triterpenes
2.2. Quantitative Content and LS-MS Profile Variation of F. viridis Fruits during Ripening
2.3. Antioxidant Potential of F. viridis Fruits: Comparision with Other Strawberries
2.4. Storage Stability of Antioxidants and Antioxidant Potential of F. viridis Ripe Fruits
3. Materials and Methods
3.1. Plant Materials and Chemicals
3.2. Total Extract Preparation from Fragaria Fruits
3.3. High-Performance Liquid Chromatography with Photodiode Array Detection and Electrospray Ionization Triple Quadrupole Mass Spectrometric Detection (HPLC-PDA-ESI-tQ-MS): Metabolite Profiling
3.4. High-Performance Liquid Chromatography with Diode Array Detection (HPLC-DAD): Carbohydrate Analysis
3.5. HPLC-ESI-tQ-MS: Metabolite Quantification
3.6. Antioxidant Activity: In Vitro Assays
3.7. DPPH Radical Scavenging Assisted HPLC-PDA-ESI-tQ-MS Assay
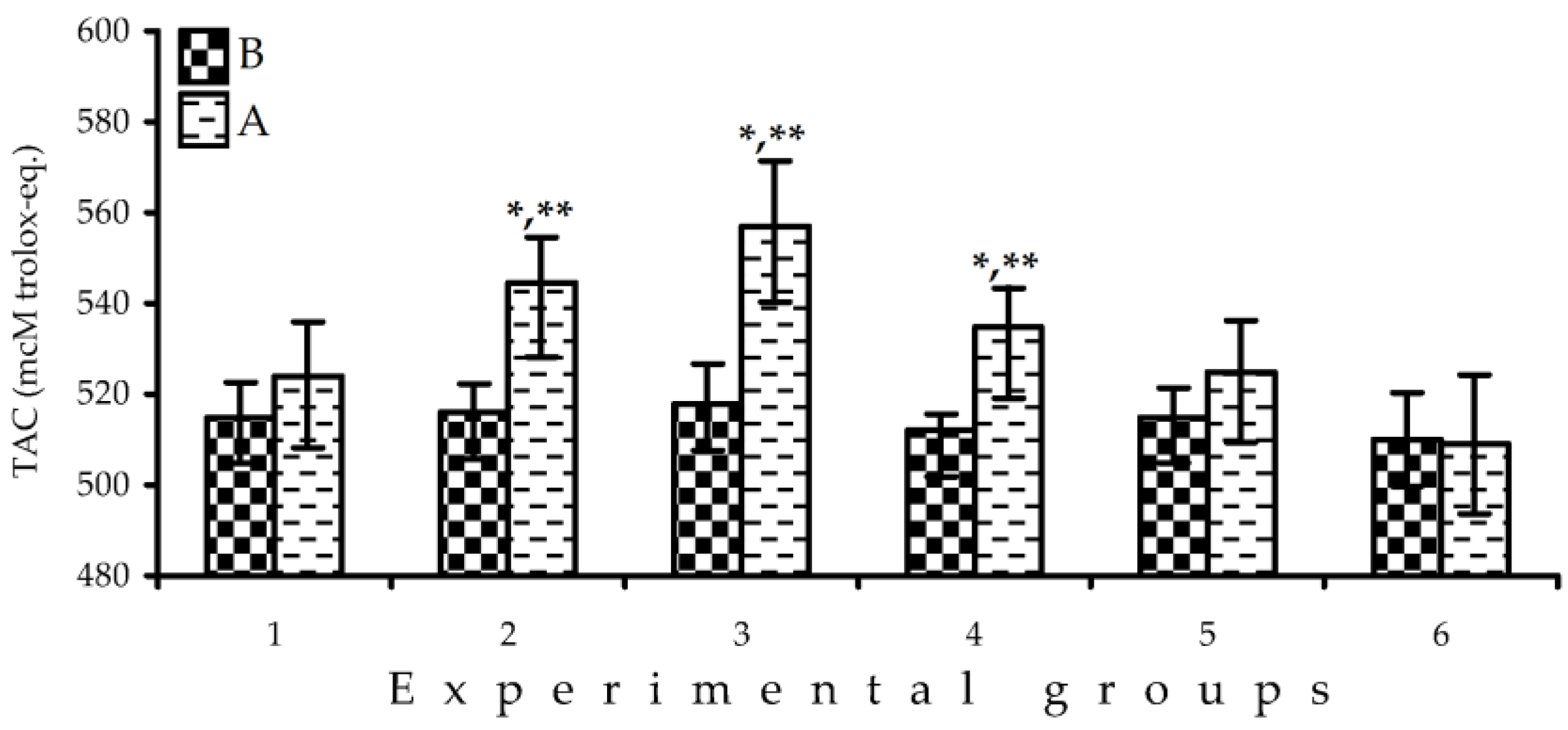
3.8. Serum Total Antioxidant Capacity
3.9. F. viridis Fruit Storage Experiment
3.10. Total Antioxidant Potential of Fresh F. viridis Fruits: Coulometric Assay
3.11. Statistical Analysis
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Liston, A.; Cronn, R.; Ashman, T.L. Fragaria: A genus with deep historical roots and ripe for evolutionary and ecological insights. Am. J. Bot. 2014, 101, 1686–1699. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fierascu, R.C.; Temocico, G.; Fierascu, I.; Ortan, A.; Babeanu, N.E. Fragaria genus: Chemical composition and biological activities. Molecules 2020, 25, 498. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cordenunsi, B.R.; Nascimento, J.R.O.; Lajolo, F.M. Physico-chemical changes related to quality of five strawberry fruit cultivars during cool-storage. Food Chem. 2003, 83, 167–173. [Google Scholar] [CrossRef]
- Pineli, L.L.O.; Moretti, C.L.; dos Santos, M.S.; Campos, A.B.; Brasileiro, A.V.; Cordova, A.C.; Chiarello, M.D. Antioxidants and other chemical and physical characteristics of two strawberry cultivars at different ripeness stages. J. Food Compos. Anal. 2011, 24, 11–16. [Google Scholar] [CrossRef]
- Gasparrini, M.; Giampieri, F.; Forbes-Hernandez, T.Y.; Afrin, S.; Cianciosi, D.; Reboredo-Rodriguez, P.; Varela-Lopez, A.; Zhang, J.; Quiles, J.L.; Mezzetti, B.; et al. Strawberry extracts effiently counteract inflammatory stress induced by the endotoxin lipopolysaccharide in human dermal fibroblast. Food Chem. Toxicol. 2018, 114, 128–140. [Google Scholar] [CrossRef]
- Cardoso, O.; Donato, M.M.; Luxo, C.; Almeida, N.; Liberal, J.; Figueirinha, A.; Batista, M.T. Anti-Helicobacter pylori potential of Agrimonia eupatoria L. and Fragaria vesca. J. Funct. Food. 2018, 44, 299–303. [Google Scholar] [CrossRef]
- Ninomiya, M.; Itoh, T.; Ishikawa, S.; Saiki, M.; Narumiya, K.; Yasuda, M.; Koshikawa, K.; Nozawa, Y.; Koketsu, M. Phenolic constituents isolated from Fragaria ananassa Duch. inhibit antigen-stimulated degranulation through direct inhibition of spleen tyrosine kinase activation. Bioorg. Med. Chem. 2010, 18, 5932–5937. [Google Scholar] [CrossRef]
- Abdulazeez, S.S. Effects of freeze-dried Fragaria × ananassa powder on alloxan-induced diabetic complications in Wistar rats. J. Taibah Univ. Med. Sci. 2014, 9, 268–273. [Google Scholar] [CrossRef] [Green Version]
- Somasagara, R.R.; Hegde, M.; Chiruvella, K.K.; Musini, A.; Choudhary, B.; Raghavan, S.C. Extracts of strawberry fruits induce intrinsic pathway of apoptosis in breast cancer cells and inhibits tumor progression in mice. PLoS ONE 2012, 7, e47021. [Google Scholar] [CrossRef] [Green Version]
- Nowicka, A.; Kucharska, A.Z.; Sokół-Łẹtowska, A.; Fecka, I. Comparison of polyphenol content and antioxidant capacity of strawberry fruit from 90 cultivars of Fragaria × ananassa Duch. Food Chem. 2019, 270, 32–46. [Google Scholar] [CrossRef]
- Komarov, V.L. Flora of USSR; AN SSSR: Moscow, Russia, 1941; Volume X, pp. 58–67. [Google Scholar]
- Gruner, P.; Ulrich, D.; Neinhuis, C.; Olbricht, K. Fragaria viridis Weston: Diversity and breeding potential of an underutilised strawberry species. Acta Horticult. 2017, 1156, 203–208. [Google Scholar] [CrossRef]
- Kirillov, V.; Stikhareva, T.; Atazhanova, G.; Makubayeva, A.; Serafimovich, M.; Kabanova, S.; Rakhimzhanov, A.; Adekenov, S. Composition of essential oil of leaves and fruits of green strawberry (Fragaria viridis Weston) growing wild in Northern Kazakhstan. J. Appl. Bot. Food Qual. 2019, 92, 39–48. [Google Scholar] [CrossRef]
- Raudonis, R.; Raudone, L.; Jakstas, V.; Janulis, V. Comparative evaluation of post-column free radical scavenging and ferric reducing antioxidant power assays for screening of antioxidants in strawberries. J. Chromatogr. A 2012, 1233, 8–15. [Google Scholar] [CrossRef] [PubMed]
- Bagdonaite, E.; Jakstas, V.; Raudonis, R.; Janulis, V. Chlorogenic acid, rutin and hyperoside content in Fragaria vesca, F. viridis and F. moschata in Lithuania. Nat. Prod. Res. 2013, 27, 181–184. [Google Scholar] [CrossRef] [PubMed]
- Olennikov, D.N.; Gadimli, A.I.; Isaev, J.I.; Kashchenko, N.I.; Prokopyev, A.S.; Katayeva, T.N.; Chirikova, N.K.; Vennos, C. Caucasian Gentiana species: Untargeted LC-MS metabolic profiling, antioxidant and digestive enzyme inhibiting activity of six plants. Metabolites 2019, 9, 271. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Olennikov, D.N.; Chirikova, N.K.; Vasilieva, A.G.; Fedorov, I.A. LC-MS profile, gastrointestinal and gut microbiota stability and antioxidant activity of Rhodiola rosea herb metabolites: A comparative study with subterranean organs. Antioxidants 2020, 9, 526. [Google Scholar] [CrossRef]
- Gasperotti, M.; Masuero, D.; Guella, G.; Palmieri, L.; Martinatti, P.; Pojer, E.; Mattivi, F.; Vrhovsek, U. Evolution of ellagitannin content and profile during fruit ripening in Fragaria spp. J. Agric. Food Chem. 2013, 61, 8597–8607. [Google Scholar] [CrossRef] [Green Version]
- Olennikov, D.N.; Chirikova, N.K.; Kashchenko, N.I.; Nikolaev, V.M.; Kim, S.-W.; Vennos, C. Bioactive phenolics of the genus Artemisia (Asteraceae): HPLC-DAD-ESI-TQ-MS/MS profile of the Siberian species and their inhibitory potential against α-amylase and α-glucosidase. Front. Pharmacol. 2018, 9, 756. [Google Scholar] [CrossRef]
- Clifford, M.N.; Wu, W.; Kuhnert, N. The chlorogenic acids of Hemerocallis. Food Chem. 2006, 95, 574–578. [Google Scholar] [CrossRef]
- Sun, J.; Lin, L.; Chen, P. Study of the mass spectrometric behaviors of anthocyanins in negative ionization mode and its applications for characterization of anthocyanins and non-anthocyanin polyphenols. Rapid Commun. Mass Spectrom. 2012, 26, 1123–1133. [Google Scholar] [CrossRef]
- Schuster, B.; Winter, M.; Herrmann, K. 4-O-β-D-Glucosides of hydroxybenzoic and hydroxycinnamic acids—Their synthesis and determination in berry fruit and vegetable. Z. Naturforsch. C. 1986, 41, 511–520. [Google Scholar] [CrossRef]
- Moilanen, J.; Sinkkonen, J.; Salminen, J.-P. Characterization of bioactive plant ellagitannins by chromatographic, spectroscopic and mass spectrometric methods. Chemoecology 2013, 23, 165–179. [Google Scholar] [CrossRef]
- Vrhovsek, U.; Guella, G.; Gasperotti, M.; Pojer, E.; Zancato, M.; Mattivi, F. Clarifying the identity of the main ellagitannin in the fruit of the strawberry, Fragaria vesca and Fragaria ananassa Duch. J. Agric. Food Chem. 2012, 60, 2507–2516. [Google Scholar] [CrossRef]
- Aaby, K.; Mazur, S.; Nes, A.; Skrede, G. Phenolic compounds in strawberry (Fragaria × ananassa Duch.) fruits: Composition in 27 cultivars and changes during ripening. Food Chem. 2012, 132, 86–97. [Google Scholar] [CrossRef]
- Aaby, K.; Ekeberg, D.; Skrede, G. Characterization of phenolic compounds in strawberry (Fragaria x ananassa) fruits by different HPLC detectors and contribution of individual compounds to total antioxidant capacity. J. Agric. Food Chem. 2007, 55, 4395–4406. [Google Scholar] [CrossRef] [PubMed]
- Kajdžanoska, M.; Gjamovski, V.; Stefova, M. HPLC-DAD-ESI-MSn identification of phenolic compounds in cultivated strawberries from Macedonia. Macedon. J. Chem. Chem. Eng. 2010, 29, 181–194. [Google Scholar] [CrossRef] [Green Version]
- Olennikov, D.N.; Kashchenko, N.I.; Chirikova, N.K. Phenolic profile of Potentilla anserina L. (Rosaceae) herb of Siberian origin and development of a rapid method for simultaneous determination of major phenolics in P. anserina pharmaceutical products by microcolumn RP-HPLC-UV. Molecules 2015, 20, 224–248. [Google Scholar] [CrossRef]
- Karlińska, E.; Pecio, Ł.; Macierzyński, J.; Stochmal, A.; Kosmala, M. Structural elucidation of the ellagitannin with a molecular weight of 2038 isolated from strawberry fruit (Fragaria ananassa Duch.) and named fragariin A. Food Chem. 2019, 296, 109–115. [Google Scholar] [CrossRef]
- Olennikov, D.N.; Kashchenko, N.I. New acylated apigenin glycosides from edge flowers of Matricaria chamomilla. Chem. Nat. Comp. 2016, 52, 996–999. [Google Scholar] [CrossRef]
- Olennikov, D.N.; Chirikova, N.K.; Kim, E.; Kim, S.W.; Zul’fugarov, I.S. New glycosides of eriodictyol from Dracocephalum palmatum. Chem. Nat. Comp. 2018, 54, 860–863. [Google Scholar] [CrossRef]
- Olennikov, D.N.; Kashchenko, N.I. New isorhamnetin glycosides and other phenolic compounds from Calendula officinalis. Chem. Nat. Comp. 2013, 49, 833–840. [Google Scholar] [CrossRef]
- Mahmood, T.; Anwar, F.; Abbas, M.; Boyce, M.C.; Saari, N.S. Compositional variation in sugars and organic acids at different maturity stages in selected small fruits from Pakistan. Int. J. Mol. Sci. 2012, 13, 1380–1392. [Google Scholar] [CrossRef] [PubMed]
- Dias, M.I.; Barros, L.; Morales, P.; Cámara, M.; Alves, M.J.; Oliveira, M.B.P.P.; Santos-Buelga, C.; Ferreira, I.C.F.R. Wild Fragaria vesca L. fruits: A rich source of bioactive phytochemicals. Food Funct. 2016, 7, 4523–4532. [Google Scholar] [CrossRef] [PubMed]
- Kosińska, A.; Diering, S.; Prim, D.; Héritier, J.; Andlauer, W. Phenolic compounds profile of strawberry fruits of Charlotte cultivar. J. Berry Res. 2013, 3, 15–23. [Google Scholar] [CrossRef] [Green Version]
- Liberal, J.; Costa, G.; Carmo, A.; Vitorino, R.; Marques, C.; Domingues, M.R.; Domingues, P.; Gonçalves, A.C.; Alves, R.; Sarmento-Ribeiro, A.B.; et al. Chemical characterization and cytotoxic potential of an ellagitannin-enriched fraction from Fragaria vesca leaves. Arab. J. Chem. 2019, 12, 3652–3666. [Google Scholar] [CrossRef] [Green Version]
- Jordheim, M.; Måge, F.; Andersen, Ø.M. Anthocyanins in berries of Ribes including Gooseberry cultivars with high content of acylated pigments. J. Agricult. Food Chem. 2007, 55, 5529–5535. [Google Scholar] [CrossRef]
- Williams, C.A. Flavone and flavonol O-glycosides. In Flavonoids: Chemistry, Biochemistry, and Applications; Andersen, Ø.M., Markham, K.R., Eds.; CRC Press: Boca Raton, FL, USA, 2006; pp. 749–856. [Google Scholar]
- Zuo, G.-Y.; Liu, S.-L.; Xu, G.-L.; Wang, G.-C.; Zhang, Y.-L.; Zheng, D. Triterpenoids from the Roots of Rubus obcordatus (Rosaceae). Plant Divers. 2008, 30, 381–382. [Google Scholar]
- Yean, M.-H.; Kim, J.-S.; Hyun, Y.-J.; Hyun, J.-W.; Bae, K.-H.; Kang, S.-S. Terpenoids and phenolics from Geum japonicum. Korean J. Pharmacogn. 2012, 43, 107–121. [Google Scholar]
- Basson, C.E.; Groenewald, J.H.; Kossmann, J.; Cronje, C.; Bauer, R. Sugar and acid-related quality attributes and enzyme activities in strawberry fruits: Invertase is the main sucrose hydrolysing enzyme. Food Chem. 2010, 121, 1156–1162. [Google Scholar] [CrossRef]
- Sturm, K.; Koron, D.; Stampar, F. The composition of fruit of different strawberries varieties depending on maturity stage. Food Chem. 2003, 83, 417–422. [Google Scholar] [CrossRef]
- Blanch, M.; Sanchez-Ballesta, M.T.; Escribano, M.I.; Merodio, C. The relationship between bound water and carbohydrate reserves in association with cellular integrity in Fragaria vesca stored under different conditions. Food Bioprocess. Technol. 2015, 8, 875–884. [Google Scholar] [CrossRef]
- Castro, I.; Goncalves, O.; Teixeira, J.A.; Vicente, A.A. Comparative study of selva and camarosa strawberries for the commercial market. J. Food Sci. 2002, 67, 2132–2137. [Google Scholar] [CrossRef]
- Kouyncu, M.A.; Dilmacunal, T. Determination of vitamin C and organic acid changes in strawberry by HPLC during cold storage. Not. Bot. Hort. Agrobot. Cluj 2010, 38, 95–98. [Google Scholar] [CrossRef]
- Palma, J.M.; Corpas, F.J.; Freschi, L.; Valpuesta, V. Fruit ripening: From present knowledge to future development. Front. Plant Sci. 2019, 10, 545. [Google Scholar] [CrossRef]
- Mazur, S.P.; Nes, A.; Wold, A.-B.; Remberg, S.F.; Martinsen, B.K.; Aaby, K. Effects of ripeness and cultivar on chemical composition of strawberry (Fragaria × ananassa Duch.) fruits and their suitability for jam production as a stable product at different storage temperatures. Food Chem. 2014, 146, 412–422. [Google Scholar] [CrossRef]
- Le Fur, Y.; Hory, C.; Bard, M.H.; Olsson, A. Evolution of phytosterols in Chardonnay grape berry skins during last stages of ripening. Vitis 1994, 33, 127–131. [Google Scholar]
- Stiti, N.; Triki, S.; Hartmann, M.A. Formation of triterpenoids throughout Olea europaea fruit ontogeny. Lipids 2007, 42, 55–67. [Google Scholar] [CrossRef]
- Kosma, D.K.; Parsons, E.P.; Isaacson, T.; Lü, S.; Rose, J.K.C.; Jenks, M.A. Fruit cuticle lipid composition during development in tomato ripening mutants. Physiol. Plant. 2010, 139, 107–117. [Google Scholar] [CrossRef]
- Zhu, Q.; Nakagawa, T.; Kishikawa, A.; Ohnuki, K.; Shimizu, K. In vitro bioactivities and phytochemical profile of various parts of the strawberry (Fragaria × ananassa var. Amaou). J. Funct. Food 2015, 13, 38–49. [Google Scholar] [CrossRef]
- Cerezo, A.B.; Cuevas, E.; Winterhalter, P.; Garcia-Parrilla, M.C.; Troncoso, A.M. Isolation, identification, and antioxidant activity of anthocyanin compounds in Camarosa strawberry. Food Chem. 2010, 123, 574–582. [Google Scholar] [CrossRef]
- Álvarez-Fernández, M.A.; Hornedo-Ortega, R.; Cerezo, A.B.; Troncoso, A.M.; García-Parrilla, M.C. Effects of the strawberry (Fragaria ananassa) purée elaboration process on non-anthocyanin phenolic composition and antioxidant activity. Food Chem. 2014, 164, 104–112. [Google Scholar] [CrossRef] [PubMed]
- Cao, G.; Russell, R.M.; Lischner, N.; Prior, R.L. Serum antioxidant capacity is increased by consumption of strawberries, spinach, red wine or vitamin C in elderly women. J. Nutr. 1998, 128, 2383–2390. [Google Scholar] [CrossRef] [PubMed]
- Henning, S.M.; Seeram, N.P.; Zhang, Y.; Li, L.; Gao, K.; Lee, R.-P.; Wang, D.C.; Zerlin, A.; Karp, H.; Thames, G.; et al. Strawberry consumption is associated with increased antioxidant capacity in serum. J. Med. Food 2010, 13, 116–122. [Google Scholar] [CrossRef] [PubMed]
- Azzini, E.; Vitaglione, P.; Intorre, F.; Napolitano, A.; Durazzo, A.; Foddai, M.S.; Fumagalli, A.; Catasta, G.; Rossi, L.; Venneria, E.; et al. Bioavailability of strawberry antioxidants in human subject. Brit. J. Nutr. 2010, 104, 1165–1173. [Google Scholar] [CrossRef] [Green Version]
- Basu, A.; Morris, S.; Nguyen, A.; Betts, N.M.; Fu, D.; Lyons, T.J. Effects of dietary strawberry supplementation on antioxidant biomarkers in obese adults with above optimal serum lipids. J. Nutr. Metabol. 2016, 2016, 3910630. [Google Scholar] [CrossRef] [Green Version]
- Pott, D.M.; Vallarino, J.G.; Osorio, S. Metabolite changes during postharvest storage: Effects on fruit quality traits. Metabolites 2020, 10, 187. [Google Scholar] [CrossRef]
- Gazdik, Z.; Zitka, O.; Petrlova, J.; Adam, V.; Zehnalek, J.; Horna, A.; Reznicek, V.; Beklova, M.; Kizek, R. Determination of vitamin C (ascorbic acid) using high performance liquid chromatography coupled with electrochemical detection. Sensors 2008, 8, 7097–7112. [Google Scholar] [CrossRef] [Green Version]
- Rubinskiene, M.; Jasutiene, I.; Venskutonis, P.R.; Viskelis, P. HPLC determination of the composition and stability of blackcurrant anthocyanins. J. Chromatogr. Sci. 2005, 43, 478–482. [Google Scholar] [CrossRef]
- Sójka, M.; Janowski, M.; Grzelak-Błaszczyk, K. Stability and transformations of raspberry (Rubus idaeus L.) ellagitannins in aqueous solutions. Eur. Food Res. Technol. 2019, 245, 1113–1122. [Google Scholar] [CrossRef] [Green Version]
- Szőke, É.; Petroianu, G.; Tekes, K.; Benkő, B.; Szegi, P.; Laufer, R.; Veress, G. HPLC monitoring of the microsomal stability of rutin and quercetin. Acta Chromatogr. 2009, 21, 399–410. [Google Scholar] [CrossRef]
- Octavia, L.; Choo, W.S. Folate, ascorbic acid, anthocyanin and colour changes in strawberry (Fragaria × annanasa) during refrigerated storage. LWT 2017, 86, 652–659. [Google Scholar] [CrossRef]
- Hernández-Herrero, J.A.; Frutos, M.J. Colour and antioxidant capacity stability in grape, strawberry and plum peel model juices at different pHs and temperatures. Food Chem. 2014, 154, 199–204. [Google Scholar] [CrossRef] [PubMed]
- Gössinger, M.; Moritz, S.; Hermes, M.; Wendelin, S.; Scherbichler, H.; Halbwirth, H.; Stich, K.; Berghofer, E. Effects of processing parameters on colour stability of strawberry nectar from puree. J. Food Eng. 2009, 90, 171–178. [Google Scholar] [CrossRef]
- Gil, M.I.; Holcroft, D.M.; Kader, A.A. Changes in strawberry anthocyanins and other polyphenols in response to carbon dioxide treatments. J. Agric. Food Chem. 1997, 45, 1662–1667. [Google Scholar] [CrossRef]
- Kashchenko, N.I.; Olennikov, D.N.; Chirikova, N.K. Ellagitannins in Rosaceous plants from the flora of Sakha (Yakutia) Republic. Butl. Commun. 2014, 39, 127–138. [Google Scholar]
- Olennikov, D.N.; Kruglova, M.Y. New quercetin glucoside and other phenolic compounds from Filipendula genus. Chem. Nat. Comp. 2013, 49, 524–529. [Google Scholar] [CrossRef]
- Olennikov, D.N. Free carbohydrates, glucofructans, and other polysaccharides from Rhaponticum uniflorum. Chem. Nat. Comp. 2018, 54, 751–754. [Google Scholar] [CrossRef]
- Olennikov, D.N.; Zilfikarov, I.N.; Penzina, T.A. Use of microcolumn HPLC for analysis of aloenin in Aloe arborescens raw material and related drugs. Pharm. Chem. J. 2013, 47, 494–497. [Google Scholar] [CrossRef]
- Olennikov, D.N.; Kashchenko, N.I.; Chirikova, N.K.; Gornostai, T.G.; Selyutina, I.Y.; Zilfikarov, I.N. Effect of low temperature cultivation on the phytochemical profile and bioactivity of Arctic plants: A case of Dracocephalum palmatum. Int. J. Mol. Sci. 2017, 18, 2579. [Google Scholar] [CrossRef] [Green Version]
- Olennikov, D.N.; Chirikova, N.K.; Okhlopkova, Z.M.; Zulfugarov, I.S. Chemical composition and antioxidant activity of Tánara Ótó (Dracocephalum palmatum Stephan), a medicinal plant used by the North-Yakutian nomads. Molecules 2013, 18, 14105–14121. [Google Scholar] [CrossRef] [Green Version]
- Pellegrini, N.; Serafini, M.; Colombi, B.; Del Rio, D.; Salvatore, S.; Bianchi, M.; Brighenti, F. Total antioxidant capacity of plant foods, beverages and oils consumed in Italy assessed by three different in vitro assays. J. Nutr. 2003, 133, 2812–2819. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Thaipong, K.; Boonprakob, U.; Crosby, K.; Cisneros-Zevallos, L.; Byrne, D.H. Comparison of ABTS, DPPH, FRAP, and ORAC assays for estimating antioxidant activity from guava fruit extracts. J. Food Comp. Anal. 2006, 19, 669–675. [Google Scholar] [CrossRef]
- Olennikov, D.N.; Kashchenko, N.I.; Chirikova, N.K. Meadowsweet teas as new functional beverages: Comparative analysis of nutrients, phytochemicals and biological effects of four Filipendula species. Molecules 2017, 22, 16. [Google Scholar] [CrossRef] [PubMed] [Green Version]




{kind=link}
{kind=link}
{kind=link}
| No. | tR, min | [M-H]− I, [M-2H]− II, [M-2H]2− III, m/z | MS/MS, m/z | Group a | Compound [ref.] b | Presence in Ripening Stages c |
|---|---|---|---|---|---|---|
| 1 | 2.51 | 341 I | CR | Hexosyl-hexose L [16] | +/+/+ | |
| 2 | 2.94 | 179 I | CR | Hexose L [16] | +/+/+ | |
| 3 | 3.08 | 191 I | OA | Citric acid S | +/+/+ | |
| 4 | 3.33 | 133 I | OA | Malic acid S | +/+/+ | |
| 5 | 3.41 | 149 I | OA | Tartaric acid S | +/+/+ | |
| 6 | 3.50 | 115 I | OA | Fumaric acid S | +/+/+ | |
| 7 | 3.82 | 175 I | OA | Ascorbic acid S | +/+/+ | |
| 8 | 4.22 | 89 I | OA | Oxalic acid S | +/+/+ | |
| 9 | 5.03 | 331 I | 169, 125 | GA | 1-O-Galloyl glucose S [17] | +/+/+ |
| 10 | 5.98 | 169 I | GA | Gallic acid S [17] | +/+/+ | |
| 11 | 6.71 | 783 I; 391 III | 633, 481, 301 | ET | Pedunculagin S [18] | +/+/+ |
| 12 | 7.03 | 161 I | CO | Umbelliferone S [19] | +/+/+ | |
| 13 | 7.43 | 353 I | 191, 179, 173, 135 | HC | 4-O-Caffeoylquinic acid S [20] | +/+/+ |
| 14 | 7.50 | 633 I | 481, 331, 301 | ET | Strictinin S [18] | +/+/+ |
| 15 | 7.58 | 353 I | 191, 165 | HC | 5-O-Caffeoylquinic acid S [20] | +/+/+ [15] |
| 16 | 7.44 | 633 I | 481, 331, 301 | ET | Strictinin isomer L [18] | +/+/+ |
| 17 | 7.83 | 353 I | 191, 179, 135 | HC | 3-O-Caffeoylquinic acid S [20] | +/+/+ |
| 18 | 8.01 | 783 I; 391 III | 633, 481, 301 | ET | Pedunculagin isomer L [18] | +/+/+ |
| 19 | 8.85 | 577 I | 289 | PC | Procyanidin B2 (catechin dimer) S [17] | +/+/+ |
| 20 | 9.15 | 609 II | 447, 285 | CY | Cyanidin 3-O-sophoroside S [21] | −/+/+ |
| 21 | 9.48 | 577 I | 289 | PC | Procyanidin B4 (catechin-epicatechin dimer) S [17] | +/+/+ |
| 22 | 9.67 | 593 II | 431, 269 | CY | Pelargonidin di-O-hexoside L [21] | −/+/+ |
| 23 | 10.14 | 289 I | CT | Catechin S [21] | +/+/+ [14] | |
| 24 | 10.49 | 325 I | 163, 119 | HC | p-Coumaric acid O-hexoside L [22] | +/+/+ |
| 25 | 10.58 | 325 I | 163, 119 | HC | p-Coumaric acid 4-O-glucoside S [22] | +/+/+ |
| 26 | 10.79 | 593 II | 447, 285 | CY | Cyanidin 3-O-rutinoside S [21] | −/+/+ |
| 27 | 11.02 | 447 II | 285 | CY | Cyanidin 3-O-glucoside S [21] | −/+/+ [14] |
| 28 | 11.14 | 577 II | 431, 269 | CY | Pelargonidin 3-O-rutinoside S [21] | −/+/+ |
| 29 | 11.52 | 431 II | 269 | CY | Pelargonidin 3-O-glucoside S [21] | +/+/+ [14] |
| 30 | 12.01 | 933 I; 466 III | 301 | ET | Castalagin S [18,23] | +/+/+ |
| 31 | 12.11 | 865 I | 577, 289 | PC | Procyanidin C2 (catechin trimer) S [17] | +/+/+ |
| 32 | 12.63 | 593 II | 447, 285 | CY | Cyanidin O-p-coumaroyl-O-hexoside L [21] | −/+/+ |
| 33 | 12.78 | 935 I; 467 III | 633, 463, 301 | ET | Casuarictin isomer [18] | +/+/+ |
| 34 | 12.88 | 933 I; 466 III | 301 | ET | Castalagin isomer L [18,23] | +/+/+ |
| 35 | 13.09 | 1103 I; 551 III | 951, 933, 783, 633, 481, 301 | ET | Sanguiin H2 S [18,24] | +/+/+ |
| 36 | 13.41 | 865 I | 577, 289 | PC | Procyanidin trimer (catechin/epicatechin trimer) L [17] | +/+/+ |
| 37 | 13.70 | 577 II | 431, 269 | CY | Pelargonidin O-p-coumaroyl-O-hexoside L [21] | −/+/+ |
| 38 | 13.98 | 489 II | 447, 285 | CY | Cyanidin O-acetyl-O-hexoside L [21] | −/+/+ |
| 39 | 14.03 | 625 I | 463, 301 | FG | Quercetin 3-O-sophoroside S [25] | +/+/+ |
| 40 | 14.11 | 935 I; 467 III | 633, 463, 301 | ET | Casuarictin isomer [18] | +/+/+ |
| 41 | 14.51 | 433 I | 301 | ET | Ellagic acid O-pentoside L [26,27] | +/+/+ |
| 42 | 14.88 | 1567 I; 783 III | 933, 633, 301 | ET | Sanguiin H10 S [18,24] | +/+/+ |
| 43 | 15.01 | 447 I | 301 | ET | Ellagic acid O-desoxyhexoside L [26,27] | +/+/+ |
| 44 | 15.11 | 1103 I; 551 III | 933, 783, 633, 481, 301 | ET | Sanguiin H2 isomer L [18,24] | +/+/+ |
| 45 | 15.34 | 609 I | 463, 301 | FG | Quercetin 3-O-rutinoside S [25] | +/+/+ [15] |
| 46 | 15.53 | 1869 I; 934 III | 1567, 1265, 935, 783, 633, 481, 301 | ET | Sanguiin H6 isomer L [18,24] | +/+/+ |
| 47 | 15.72 | 1401 III | 1235, 933, 783, 633, 301 | ET | Lambertianin C S [18,24] | +/+/+ |
| 48 | 15.81 | 473 II | 431, 269 | CY | Pelargonidin O-acetyl-O-hexoside L [21] | −/+/+ |
| 49 | 15.94 | 463 I | 301 | FG | Quercetin 3-O-glucoside S [25] | +/+/+ [14] |
| 50 | 16.02 | 477 I | 301 | FG | Quercetin 3-O-glucuronide S [25] | +/+/+ |
| 51 | 16.33 | 1103 I; 551 III | 801, 783, 499, 481, 319, 301 | ET | Agrimonic acid A S [28] | +/+/+ |
| 52 | 16.50 | 1869 I; 934 III | 1701, 1567, 1265, 1085, 935, 783, 633, 481, 301 | ET | Sanguiin H6 S [18,24] | +/+/+ |
| 53 | 16.71 | 301 I | 229 | ET | Ellagic acid S [18] | +/+/+ [14] |
| 54 | 16.81 | 1103 I; 551 III | 801, 783, 499, 481, 319, 301 | ET | Agrimonic acid B S [28] | +/+/+ |
| 55 | 16.95 | 433 | 301 | FG | Quercetin 3-O-xyloside S [25] | +/+/+ |
| 56 | 17.07 | 433 | 301 | FG | Quercetin 3-O-arabinoside S [25] | +/+/+ |
| 57 | 17.41 | 593 I | 447, 285 | FG | Kaempferol 3-O-rutinoside S [25] | +/+/+ |
| 58 | 17.56 | 447 I | 285 | FG | Kaempferol 3-O-glucoside S [25] | +/+/+ |
| 59 | 17.92 | 461 I | 285 | FG | Kaempferol 3-O-glucuronide S [25] | +/+/+ |
| 60 | 18.21 | 609 I | 463, 301 | FG | Quercetin 3-O-(6″-O-p-coumaroyl)-glucoside S [25] | +/+/+ |
| 61 | 18.29 | 609 I | 463, 301 | FG | Quercetin O-p-coumaroyl-O-hexoside L [25] | +/+/+ |
| 62 | 18.50 | 1869 I; 934 III | 1567, 1265, 1085, 935, 783, 633, 481, 301 | ET | Agrimoniin S [23,29] | +/+/+ |
| 63 | 18.68 | 1018 III | 1691, 1567, 1265, 1209, 935, 783, 633, 481, 301 | ET | Fragariin A L [23,29] | +/+/+ |
| 64 | 19.06 | 549 I | 463, 301 | FG | Quercetin O-malonyl-O-hexoside L [30,31] | +/+/+ |
| 65 | 19.42 | 549 I | 463, 301 | FG | Quercetin 3-O-(6″-O-malonyl)-glucoside S [30,31] | +/+/+ |
| 66 | 19.75 | 593 I | 447, 285 | FG | Kaempferol 3-O-(6″-O-p-coumaroyl)-glucoside S [25] | +/+/+ |
| 67 | 19.83 | 939 I | 787, 635, 483, 331, 169 | GA | 1,2,3,4,6-Penta-O-galloylglucose S [17] | +/+/+ |
| 68 | 20.39 | 533 I | 447, 285 | FG | Kaempferol O-malonyl-O-hexoside L [30,31] | +/+/+ |
| 69 | 20.82 | 533 I | 447, 285 | FG | Kaempferol 3-O-(6″-O-malonyl)-glucoside S [30,31] | +/+/+ |
| 70 | 21.83 | 505 I | 463, 301 | FG | Quercetin 3-O-(2″-O-acetyl)-glucoside S [32] | +/+/+ |
| 71 | 22.14 | 505 I | 463, 301 | FG | Quercetin 3-O-(6″-O-acetyl)-glucoside S [32] | +/+/+ |
| 72 | 22.67 | 489 I | 447, 285 | FG | Kaempferol O-acetyl-O-hexoside L [30,31,32] | +/+/+ |
| 73 | 23.52 | 489 I | 447, 285 | FG | Kaempferol O-acetyl-O-hexoside L [30,31,32] | +/+/+ |
| 74 | 24.21 | 301 I | FG | Quercetin S [25] | +/+/+ [14] | |
| 75 | 24.78 | 811 I | 649, 487 | TR | Tormentic acid di-O-hexoside L [16] | +/+/+ |
| 76 | 25.34 | 285 I | FG | Kaempferol S [25] | +/+/+ | |
| 77 | 25.54 | 795 I | 633, 471 | TR | Pomolic acid di-O-hexoside L [16] | +/+/+ |
| 78 | 25.87 | 649 I | 487 | TR | Tormentic acid O-hexoside L [16] | +/+/+ |
| 79 | 26.41 | 547 I | 505, 463, 301 | FG | Quercetin 3-O-(2″,6″-di-O-acetyl)-glucoside S [32] | +/+/+ |
| 80 | 27.52 | 591 I | 549, 505, 463, 301 | FG | Quercetin O-acetyl-O-malonyl-O-hexoside L [30,31,32] | +/+/+ |
| 81 | 27.73 | 487 I | TR | Tormentic acid S [16] | +/+/+ | |
| 82 | 27.89 | 461 I | 315, 301 | ET | Ellagic acid O-methyl ester-O-desoxyhexoside L [26,27] | +/+/+ |
| 83 | 28.78 | 633 I | 471 | TR | Pomolic acid O-hexoside L [16] | +/+/+ |
| 84 | 29.14 | 695 I | 609, 463, 301 | FG | Quercetin O-malonyl-O-p-coumaroyl-O-hexoside L [30,31,32] | +/+/+ |
| 85 | 29.49 | 695 I | 609, 463, 301 | FG | Quercetin O-malonyl-O-p-coumaroyl-O-hexoside L [30,31,32] | +/+/+ |
| 86 | 29.57 | 475 I | 329, 301 | ET | Ellagic acid di-O-methyl ester-O-desoxyhexoside L [26,27] | +/+/+ |
| 87 | 30.08 | 531 I | 489, 447, 285 | FG | Kaempferol di-O-acetyl-O-hexoside L [30,31,32] | +/+/+ |
| 88 | 30.41 | 651 I | 609, 463, 301 | FG | Quercetin O-acetyl-O-p-coumaroyl-O-hexoside L [30,31,32] | +/+/+ |
| 89 | 30.92 | 575 I | 533, 489, 447, 285 | FG | Kaempferol O-acetyl-O-malonyl-O-hexoside L [30,31,32] | +/+/+ |
| 90 | 31.02 | 679 I | 593, 447, 285 | FG | Kaempferol O-malonyl-O-p-coumaroyl-O-hexoside L [30,31,32] | +/+/+ |
| 91 | 31.22 | 471 I | TR | Pomolic acid S [16] | +/+/+ | |
| 92 | 31.38 | 679 I | 593, 447, 285 | FG | Kaempferol O-malonyl-O-p-coumaroyl-O-hexoside L [30,31,32] | +/+/+ |
| 93 | 31.98 | 635 I | 593, 447, 285 | FG | Kaempferol O-acetyl-O-p-coumaroyl-O-hexoside L [30,31,32] | +/+/+ |
| 94 | 32.86 | 693 I | 651, 609, 463, 301 | FG | Quercetin di-O-acetyl-O-p-coumaroyl-O-hexoside L [30,31,32] | +/+/+ |
| 95 | 34.26 | 737 I | 695, 651, 609, 463, 301 | FG | Quercetin O-acetyl-O-malonyl-O-p-coumaroyl-O-hexoside L [30,31,32] | +/+/+ |
| Compound | Stage of Ripeness | ||
|---|---|---|---|
| Unripe | Intermediate | Ripe | |
| Carbohydrates | |||
| Hexose (glucose+fructose) | 41.10 ± 0.82 | 43.26 ± 0.90 | 45.16 ± 0.92 |
| Hexosyl-hexose (saccharose) | 0.04 ± 0.00 | 0.06 ± 0.00 | 0.11 ± 0.00 |
| Total carbohydrates | 41.14 | 43.32 | 45.27 |
| Organic acids | |||
| Ascorbic acid | 0.62 ± 0.02 | 0.86 ± 0.02 | 1.12 ± 0.02 |
| Citric acid | 2.83 ± 0.06 | 3.18 ± 0.07 | 5.63 ± 0.11 |
| Malic acid | 0.42 ± 0.01 | 0.45 ± 0.01 | 0.59 ± 0.02 |
| Tartaric acid | 0.37 ± 0.01 | 0.40 ± 0.01 | 0.42 ± 0.01 |
| Fumaric acid | 0.01 ± 0.00 | 0.03 ± 0.00 | 0.07 ± 0.00 |
| Oxalic acid | traces | traces | 0.05 ± 0.00 |
| Total organic acids | 4.25 | 4.92 | 7.88 |
| Gallic acid derivatives | |||
| Gallic acid | traces | 0.01 ± 0.00 | 0.01 ± 0.00 |
| 1-O-Galloyl glucose | 0.05 ± 0.00 | 0.03 ± 0.00 | 0.03 ± 0.00 |
| 1,2,3,4,6-Penta-O-galloylglucose | traces | traces | traces |
| Total gallic acid derivatives | 0.05 | 0.04 | 0.04 |
| Hydroxycinnamates and coumarins | |||
| p-Coumaric acid 4-O-glucoside | 0.35 ± 0.01 | 0.33 ± 0.01 | 0.29 ± 0.00 |
| p-Coumaric acid O-hexoside 24 | 0.14 ± 0.00 | 0.11 ± 0.00 | 0.08 ± 0.00 |
| 3-O-Caffeoylquinic acid | 0.12 ± 0.00 | 0.08 ± 0.00 | 0.04 ± 0.00 |
| 4-O-Caffeoylquinic acid | 0.08 ± 0.00 | 0.07 ± 0.00 | 0.05 ± 0.00 |
| 5-O-Caffeoylquinic acid | 0.28 ± 0.00 | 0.21 ± 0.00 | 0.14 ± 0.00 |
| Umbelliferone | traces | traces | traces |
| Total hydroxycinnamates and coumarins | 0.97 | 0.80 | 0.60 |
| Ellagic acid derivatives and ellagitannins | |||
| Ellagic acid | 0.10 ± 0.00 | 0.10 ± 0.00 | 0.12 ± 0.00 |
| Ellagic acid O-pentoside 41 | 0.05 ± 0.00 | 0.09 ± 0.00 | 0.11 ± 0.00 |
| Ellagic acid O-desoxyhexoside 43 | 0.01 ± 0.00 | 0.04 ± 0.00 | 0.07 ± 0.00 |
| Ellagic acid O-methyl ester-O-desoxyhexoside 82 | 0.14 ± 0.00 | 0.16 ± 0.00 | 0.24 ± 0.00 |
| Ellagic acid di-O-methyl ester-O-desoxyhexoside 86 | 0.10 ± 0.00 | 0.18 ± 0.00 | 0.30 ± 0.00 |
| Pedunculagin | 0.26 ± 0.00 | 0.30 ± 0.01 | 0.32 ± 0.01 |
| Pedunculagin isomer 18 | 0.05 ± 0.00 | 0.07 ± 0.00 | 0.11 ± 0.00 |
| Strictinin isomer 14 | 0.10 ± 0.00 | 0.10 ± 0.00 | 0.12 ± 0.00 |
| Strictinin isomer 16 | 0.11 ± 0.00 | 0.12 ± 0.00 | 0.18 ± 0.00 |
| Castalagin isomer 30 | traces | 0.02 ± 0.00 | 0.04 ± 0.00 |
| Castalagin isomer 34 | traces | traces | 0.01 ± 0.00 |
| Casuarictin isomer 33 | traces | traces | 0.02 ± 0.00 |
| Casuarictin isomer 40 | 0.06 ± 0.00 | 0.08 ± 0.00 | 0.14 ± 0.00 |
| Sanguiin H2 | traces | 0.01 ± 0.00 | 0.05 ± 0.00 |
| Sanguiin H2 isomer 44 | 0.09 ± 0.00 | 0.05 ± 0.00 | 0.02 ± 0.00 |
| Sanguiin H6 | 0.36 ± 0.01 | 0.25 ± 0.00 | 0.22 ± 0.00 |
| Sanguiin H6 isomer 46 | 0.45 ± 0.01 | 0.43 ± 0.01 | 0.40 ± 0.01 |
| Sanguiin H10 | 0.21 ± 0.00 | 0.15 ± 0.00 | 0.08 ± 0.00 |
| Lambertianin C | 1.86 ± 0.04 | 1.42 ± 0.03 | 1.20 ± 0.02 |
| Agrimonic acid A | 0.02 ± 0.00 | 0.05 ± 0.00 | 0.08 ± 0.00 |
| Agrimonic acid B | 0.01 ± 0.00 | 0.03 ± 0.00 | 0.10 ± 0.00 |
| Agrimoniin | 2.63 ± 0.05 | 2.03 ± 0.04 | 1.41 ± 0.03 |
| Fragariin A | 0.93 ± 0.02 | 0.69 ± 0.02 | 0.63 ± 0.01 |
| Total ellagic acid derivatives and ellagitannins | 7.54 | 6.37 | 5.97 |
| Catechins and procyanidins | |||
| Catechin | 0.11 ± 0.00 | 0.05 ± 0.00 | 0.05 ± 0.00 |
| Procyanidin B2 | 0.09 ± 0.00 | 0.05 ± 0.00 | 0.02 ± 0.00 |
| Procyanidin B4 | 0.02 ± 0.00 | 0.01 ± 0.00 | traces |
| Procyanidin C2 | 0.05 ± 0.00 | 0.03 ± 0.00 | 0.01 ± 0.00 |
| Procyanidin trimer 36 | 0.02 ± 0.00 | 0.01 ± 0.00 | 0.01 ± 0.00 |
| Total catechins and procyanidins | 0.29 | 0.15 | 0.09 |
| Anthocyanins | |||
| Pelargonidin 3-O-glucoside | traces | 0.02 ± 0.00 | 0.06 ± 0.00 |
| Pelargonidin 3-O-rutinoside | n.d. | n.d. | 0.03 ± 0.00 |
| Pelargonidin di-O-hexoside 22 | n.d. | n.d. | traces |
| Pelargonidin O-acetyl-O-hexoside 48 | n.d. | n.d. | traces |
| Pelargonidin O-p-coumaroyl-O-hexoside 37 | n.d. | n.d. | traces |
| Cyanidin 3-O-glucoside | n.d. | 0.01 ± 0.00 | 0.05 ± 0.00 |
| Cyanidin 3-O-rutinoside | n.d. | n.d. | traces |
| Cyanidin 3-O-sophoroside | n.d. | n.d. | traces |
| Cyanidin O-acetyl-O-hexoside 38 | n.d. | n.d. | traces |
| Cyanidin O-p-coumaroyl-O-hexoside 32 | n.d. | n.d. | traces |
| Total anthocyanins | traces | 0.03 | 0.14 |
| Flavonols and flavonol glycosides | |||
| Kaempferol | traces | traces | 0.01 ± 0.00 |
| Kaempferol 3-O-glucoside | traces | 0.05 ± 0.00 | 0.09 ± 0.00 |
| Kaempferol 3-O-glucuronide | traces | traces | 0.08 ± 0.00 |
| Kaempferol 3-O-rutinoside | 0.28 ± 0.00 | 0.23 ± 0.00 | 0.11 ± 0.00 |
| Kaempferol O-acetyl-O-hexoside 72 | traces | traces | traces |
| Kaempferol O-acetyl-O-hexoside 73 | traces | traces | traces |
| Kaempferol di-O-acetyl-O-hexoside 87 | traces | traces | traces |
| Kaempferol O-malonyl-O-hexoside 68 | 0.01 ± 0.00 | traces | traces |
| Kaempferol O-malonyl-O-hexoside 69 | 0.02 ± 0.00 | 0.01 ± 0.00 | traces |
| Kaempferol 3-O-(6″-O-p-coumaroyl)-glucoside | 0.08 ± 0.00 | 0.04 ± 0.00 | 0.04 ± 0.00 |
| Kaempferol O-acetyl-O-malonyl-O-hexoside 89 | 0.02 ± 0.00 | 0.01 ± 0.00 | 0.01 ± 0.00 |
| Kaempferol O-malonyl-O-p-coumaroyl-O-hexoside 90 | traces | traces | traces |
| Kaempferol O-malonyl-O-p-coumaroyl-O-hexoside 92 | traces | traces | traces |
| Kaempferol O-acetyl-O-p-coumaroyl-O-hexoside 93 | 0.01 ± 0.00 | 0.01 ± 0.00 | 0.01 ± 0.00 |
| Total kaempferol derivatives | 0.42 | 0.35 | 0.35 |
| Quercetin | traces | traces | 0.02 ± 0.00 |
| Quercetin 3-O-xyloside | traces | traces | 0.03 ± 0.00 |
| Quercetin 3-O-arabinoside | traces | traces | 0.01 ± 0.00 |
| Quercetin 3-O-glucoside | traces | 0.04 ± 0.00 | 0.08 ± 0.00 |
| Quercetin 3-O-glucuronide | traces | 0.05 ± 0.00 | 0.11 ± 0.00 |
| Quercetin 3-O-rutinoside | 0.32 ± 0.00 | 0.28 ± 0.00 | 0.25 ± 0.00 |
| Quercetin 3-O-sophoroside | 0.11 ± 0.00 | 0.08 ± 0.00 | 0.03 ± 0.00 |
| Quercetin 3-O-(2″-O-acetyl)-glucoside | 0.06 ± 0.00 | 0.03 ± 0.00 | 0.01 ± 0.00 |
| Quercetin 3-O-(6″-O-acetyl)-glucoside | 0.03 ± 0.00 | 0.02 ± 0.00 | 0.02 ± 0.00 |
| Quercetin 3-O-(2″,6″-di-O-acetyl)-glucoside | 0.03 ± 0.00 | 0.01 ± 0.00 | 0.01 ± 0.00 |
| Quercetin 3-O-(6″-O-malonyl)-glucoside | 0.04 ± 0.00 | 0.04 ± 0.00 | 0.02 ± 0.00 |
| Quercetin O-malonyl-O-hexoside 64 | 0.01 ± 0.00 | traces | traces |
| Quercetin 3-O-(6″-O-p-coumaroyl)-glucoside | 0.11 ± 0.00 | 0.06 ± 0.00 | 0.05 ± 0.00 |
| Quercetin O-p-coumaroyl-O-hexoside 61 | 0.02 ± 0.00 | 0.02 ± 0.00 | 0.01 ± 0.00 |
| Quercetin O-acetyl-O-malonyl-O-hexoside 80 | 0.01 ± 0.00 | 0.01 ± 0.00 | traces |
| Quercetin O-malonyl-O-p-coumaroyl-O-hexoside 84 | 0.02 ± 0.00 | 0.01 ± 0.00 | traces |
| Quercetin O-malonyl-O-p-coumaroyl-O-hexoside 85 | 0.01 ± 0.00 | 0.01 ± 0.00 | traces |
| Quercetin O-acetyl-O-p-coumaroyl-O-hexoside 88 | 0.01 ± 0.00 | traces | traces |
| Quercetin di-O-acetyl-O-p-coumaroyl-O-hexoside 94 | traces | traces | traces |
| Quercetin O-acetyl-O-malonyl-O-p-coumaroyl-O-hexoside 95 | 0.04 ± 0.00 | 0.02 ± 0.00 | 0.01 ± 0.00 |
| Total quercetin derivatives | 0.82 | 0.68 | 0.66 |
| Total flavonols and flavonol glycosides | 1.24 | 1.03 | 1.01 |
| Triterpenes | |||
| Pomolic acid | traces | traces | 0.01 ± 0.00 |
| Pomolic acid O-hexoside 83 | traces | traces | traces |
| Pomolic acid di-O-hexoside 77 | traces | traces | traces |
| Tormentic acid | traces | traces | 0.02 ± 0.00 |
| Tormentic acid O-hexoside 78 | traces | traces | 0.01 ± 0.00 |
| Tormentic acid di-O-hexoside 75 | traces | traces | traces |
| Total triterpenes | traces | traces | 0.04 |
| Assay a | F. viridis | F. vesca (Ripe) | F. ananassa (Ripe) | ||
|---|---|---|---|---|---|
| Unripe | Intermediate | Ripe | |||
| DPPH | 29.2 ± 0.6 d,e | 28.4 ± 0.5 c | 27.5 ± 0.5 c,d | 15.2 ± 0.3 b | 9.3 ± 0.2 a |
| ABTS | 35.1 ± 0.8 h,i | 35.3 ± 0.8 h | 36.2 ± 0.9 i | 19.7 ± 0.4 f,g | 14.7 ± 0.3 f |
| FRAP | 42.6 ± 1.0 l | 45.4 ± 1.0 l,m | 47.1 ± 1.0 m | 27.1 ± 0.5 k | 21.1 ± 0.4 j |
| ORAC | 33.6 ± 0.8 p,q | 32.8 ± 0.7 o,p | 33.0 ± 0.8 p | 25.1 ± 0.5 n,o | 18.9 ± 0.4 n |
| Compound | T, °C | Day of Storage | |||||||
|---|---|---|---|---|---|---|---|---|---|
| 0 | 1 | 2 | 3 | 4 | 5 | 6 | 7 | ||
| Ascorbic acid | 4 | 1.14 ± 0.02 | 1.02 ± 0.02 | 0.95 ± 0.02 | 0.89 ± 0.02 | 0.86 ± 0.02 | 0.73 ± 0.02 | 0.55 ± 0.02 | 0.51 ± 0.02 |
| 20 | 0.85 ± 0.02 | 0.54 ± 0.01 | 0.36 ± 0.01 | n.a. | n.a. | n.a. | n.a. | ||
| Ellagic acid | 4 | 0.10 ± 0.00 | 0.10 ± 0.00 | 0.10 ± 0.00 | 0.12 ± 0.00 | 0.14 ± 0.00 | 0.15 ± 0.00 | 0.17 ± 0.00 | 0.19 ± 0.00 |
| 20 | 0.10 ± 0.00 | 0.12 ± 0.00 | 0.25 ± 0.00 | n.a. | n.a. | n.a. | n.a. | ||
| Pedunculagin | 4 | 0.33 ± 0.01 | 0.34 ± 0.01 | 0.34 ± 0.01 | 0.35 ± 0.01 | 0.35 ± 0.01 | 0.38 ± 0.01 | 0.38 ± 0.01 | 0.40 ± 0.01 |
| 20 | 0.35 ± 0.01 | 0.38 ± 0.01 | 0.49 ± 0.01 | n.a. | n.a. | n.a. | n.a. | ||
| Sanguiin H6 | 4 | 0.20 ± 0.00 | 0.20 ± 0.00 | 0.20 ± 0.00 | 0.21 ± 0.00 | 0.22 ± 0.00 | 0.24 ± 0.00 | 0.25 ± 0.00 | 0.25 ± 0.00 |
| 20 | 0.20 ± 0.00 | 0.21 ± 0.00 | 0.30 ± 0.00 | n.a. | n.a. | n.a. | n.a. | ||
| Lambertianin C | 4 | 1.26 ± 0.02 | 1.24 ± 0.02 | 1.20 ± 0.02 | 1.15 ± 0.02 | 1.11 ± 0.02 | 0.99 ± 0.02 | 0.97 ± 0.02 | 0.93 ± 0.02 |
| 20 | 1.04 ± 0.02 | 0.90 ± 0.02 | 0.72 ± 0.02 | n.a. | n.a. | n.a. | n.a. | ||
| Agrimoniin | 4 | 1.47 ± 0.03 | 1.45 ± 0.03 | 1.40 ± 0.03 | 1.37 ± 0.03 | 1.35 ± 0.03 | 1.25 ± 0.02 | 1.22 ± 0.02 | 1.17 ± 0.02 |
| 20 | 1.33 ± 0.03 | 1.08 ± 0.02 | 0.93 ± 0.02 | n.a. | n.a. | n.a. | n.a. | ||
| Fragariin A | 4 | 0.65 ± 0.02 | 0.65 ± 0.02 | 0.62 ± 0.02 | 0.60 ± 0.02 | 0.59 ± 0.01 | 0.55 ± 0.01 | 0.53 ± 0.02 | 0.51 ± 0.01 |
| 20 | 0.42 ± 0.01 | 0.34 ± 0.01 | 0.22 ± 0.00 | n.a. | n.a. | n.a. | n.a. | ||
| Pelargonidin 3-O-glucoside | 4 | 0.07 ± 0.00 | 0.07 ± 0.00 | 0.07 ± 0.00 | 0.07 ± 0.00 | 0.06 ± 0.00 | 0.06 ± 0.00 | 0.05 ± 0.00 | 0.05 ± 0.00 |
| 20 | 0.04 ± 0.00 | 0.02 ± 0.00 | traces | n.a. | n.a. | n.a. | n.a. | ||
| Cyanidin 3-O-glucoside | 4 | 0.05 ± 0.00 | 0.05 ± 0.00 | 0.05 ± 0.00 | 0.04 ± 0.00 | 0.04 ± 0.00 | 0.02 ± 0.00 | 0.02 ± 0.00 | 0.02 ± 0.00 |
| 20 | 0.02 ± 0.00 | traces | traces | n.a. | n.a. | n.a. | n.a. | ||
| Quercetin 3-O-glucoside | 4 | 0.10 ± 0.00 | 0.10 ± 0.00 | 0.10 ± 0.00 | 0.10 ± 0.00 | 0.10 ± 0.00 | 0.10 ± 0.00 | 0.11 ± 0.00 | 0.11 ± 0.00 |
| 20 | 0.10 ± 0.00 | 0.11 ± 0.00 | 0.12 ± 0.00 | n.a. | n.a. | n.a. | n.a. | ||
| Quercetin 3-O-glucuronide | 4 | 0.15 ± 0.00 | 0.15 ± 0.00 | 0.15 ± 0.00 | 0.16 ± 0.00 | 0.17 ± 0.00 | 0.17 ± 0.00 | 0.17 ± 0.00 | 0.17 ± 0.00 |
| 20 | 0.15 ± 0.00 | 0.15 ± 0.00 | 0.17 ± 0.00 | n.a. | n.a. | n.a. | n.a. | ||
| Quercetin 3-O-rutinoside | 4 | 0.24 ± 0.00 | 0.24 ± 0.00 | 0.24 ± 0.00 | 0.24 ± 0.00 | 0.24 ± 0.00 | 0.22 ± 0.00 | 0.21 ± 0.00 | 0.20 ± 0.00 |
| 20 | 0.24 ± 0.00 | 0.20 ± 0.00 | 0.18 ± 0.00 | n.a. | n.a. | n.a. | n.a. | ||
| Total antioxidant potential | 4 | 4.12 ± 0.09 | 4.10 ± 0.08 | 4.07 ± 0.08 | 4.02 ± 0.08 | 3.97 ± 0.08 | 3.86 ± 0.08 | 3.59 ± 0.07 | 3.27 ± 0.07 |
| 20 | 2.88 ± 0.05 | 1.72 ± 0.04 | 0.52 ± 0.02 | n.a. | n.a. | n.a. | n.a. | ||
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Olennikov, D.N.; Vasilieva, A.G.; Chirikova, N.K. Fragaria viridis Fruit Metabolites: Variation of LC-MS Profile and Antioxidant Potential during Ripening and Storage. Pharmaceuticals 2020, 13, 262. https://0-doi-org.brum.beds.ac.uk/10.3390/ph13090262
Olennikov DN, Vasilieva AG, Chirikova NK. Fragaria viridis Fruit Metabolites: Variation of LC-MS Profile and Antioxidant Potential during Ripening and Storage. Pharmaceuticals. 2020; 13(9):262. https://0-doi-org.brum.beds.ac.uk/10.3390/ph13090262
Chicago/Turabian StyleOlennikov, Daniil N., Aina G. Vasilieva, and Nadezhda K. Chirikova. 2020. "Fragaria viridis Fruit Metabolites: Variation of LC-MS Profile and Antioxidant Potential during Ripening and Storage" Pharmaceuticals 13, no. 9: 262. https://0-doi-org.brum.beds.ac.uk/10.3390/ph13090262