
Identification of Lactate as a Cardiac Protectant by Inhibiting Inflammation and Cardiac Hypertrophy Using a Zebrafish Acute Heart Failure Model


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
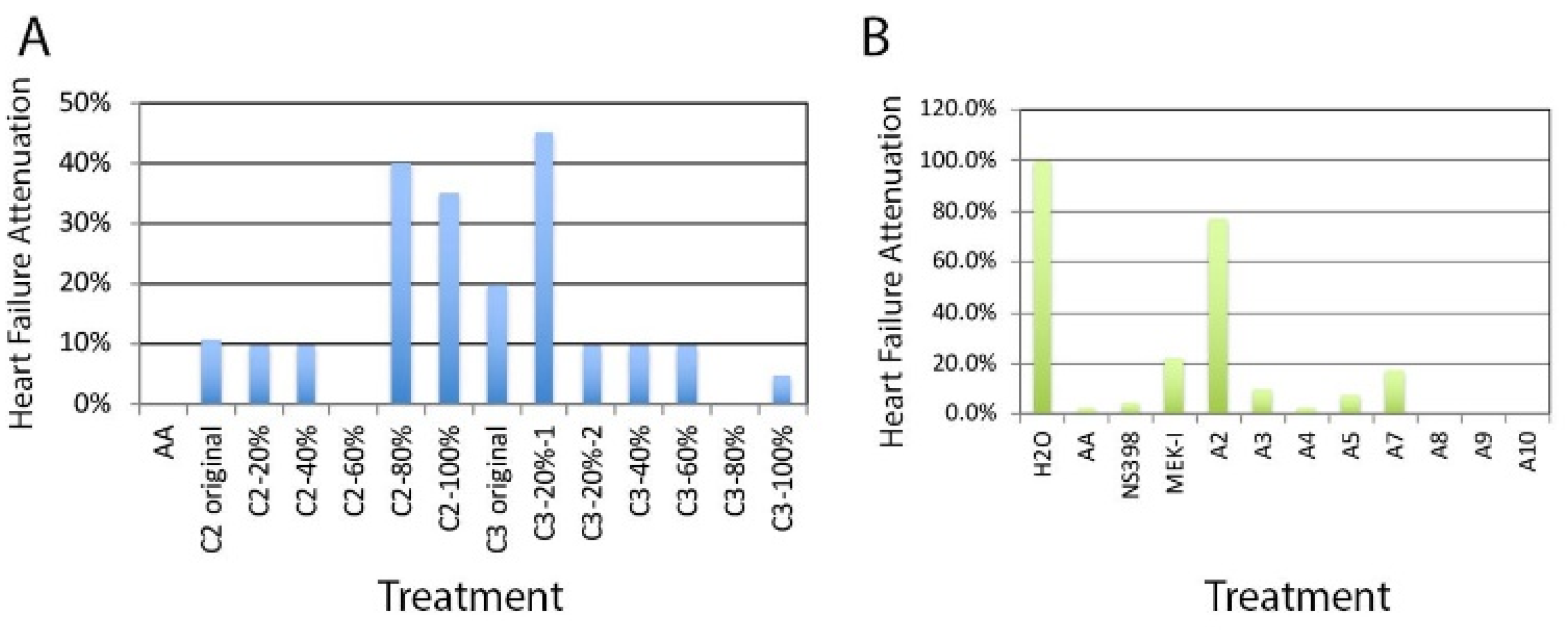
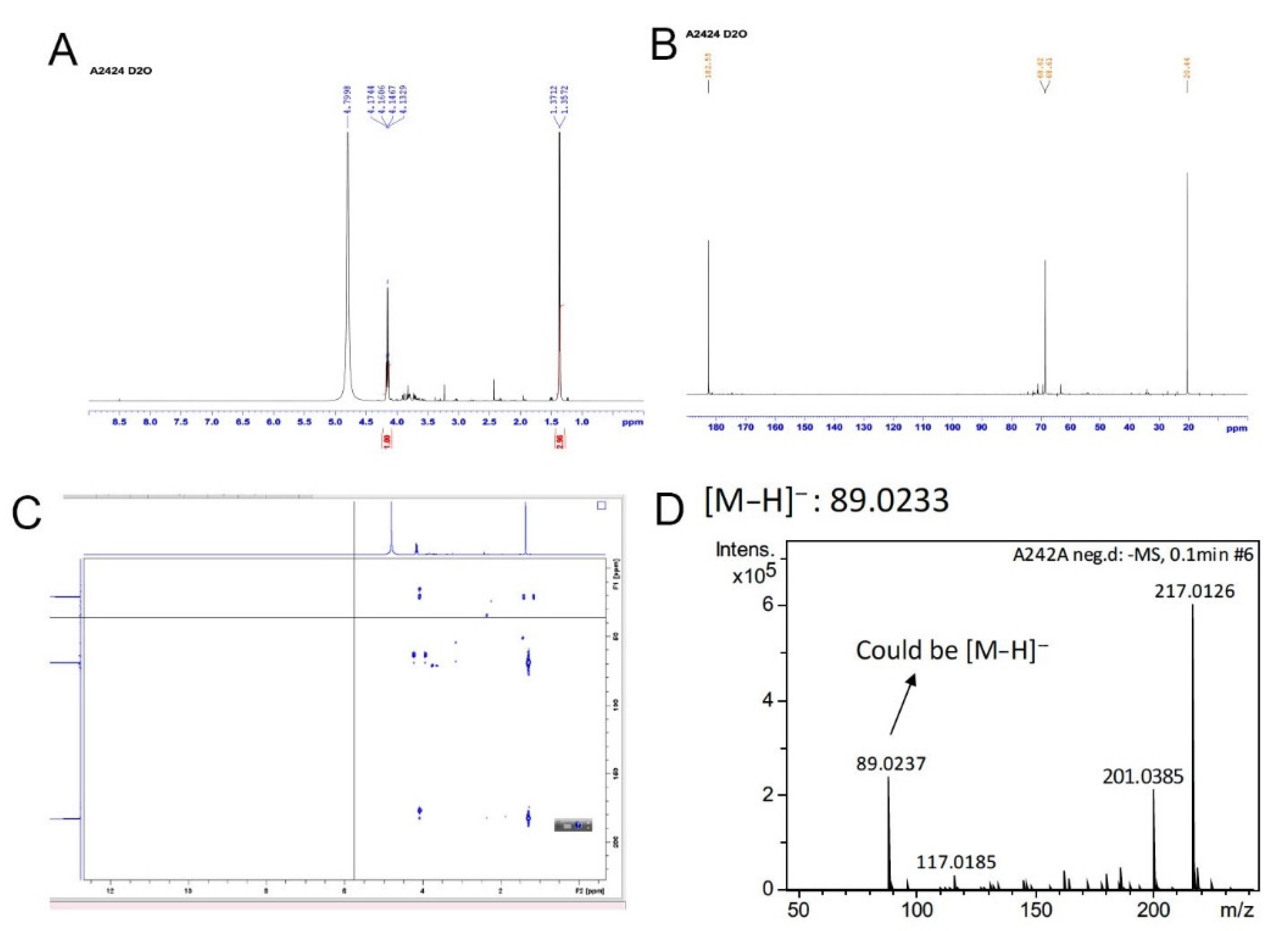
2.1. Chemical Extraction
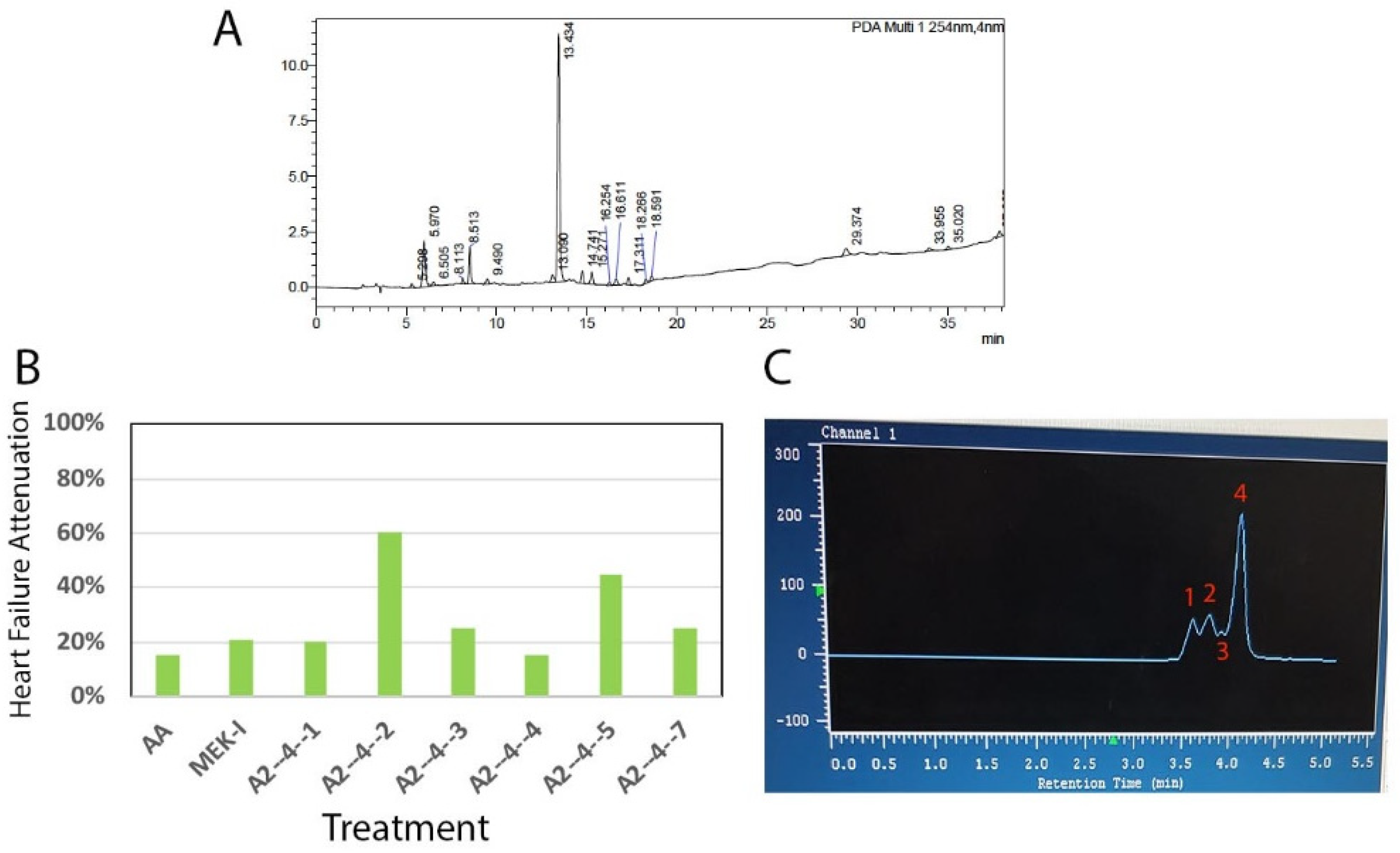
2.2. Bioassay-Guided Chemical Fractionation
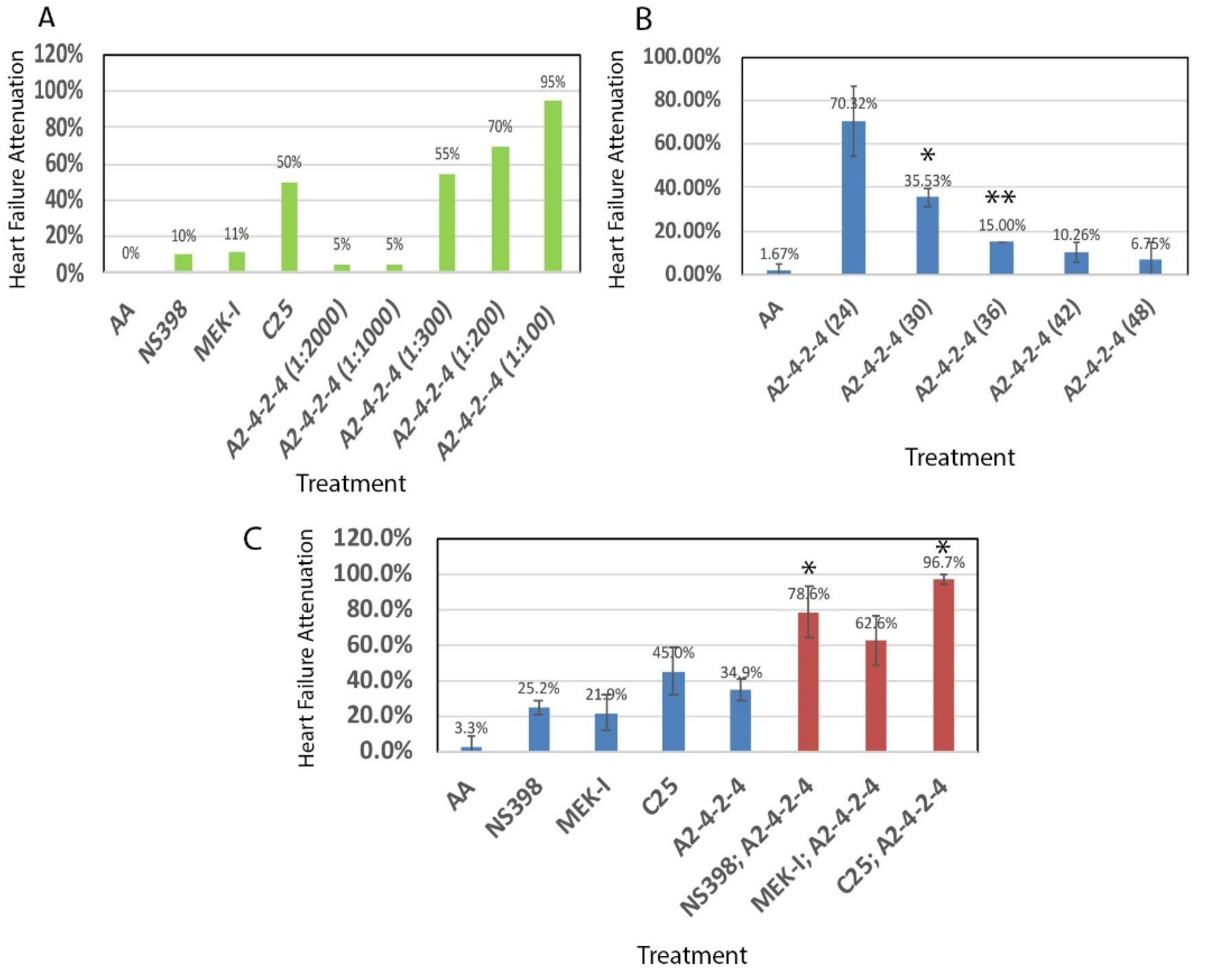
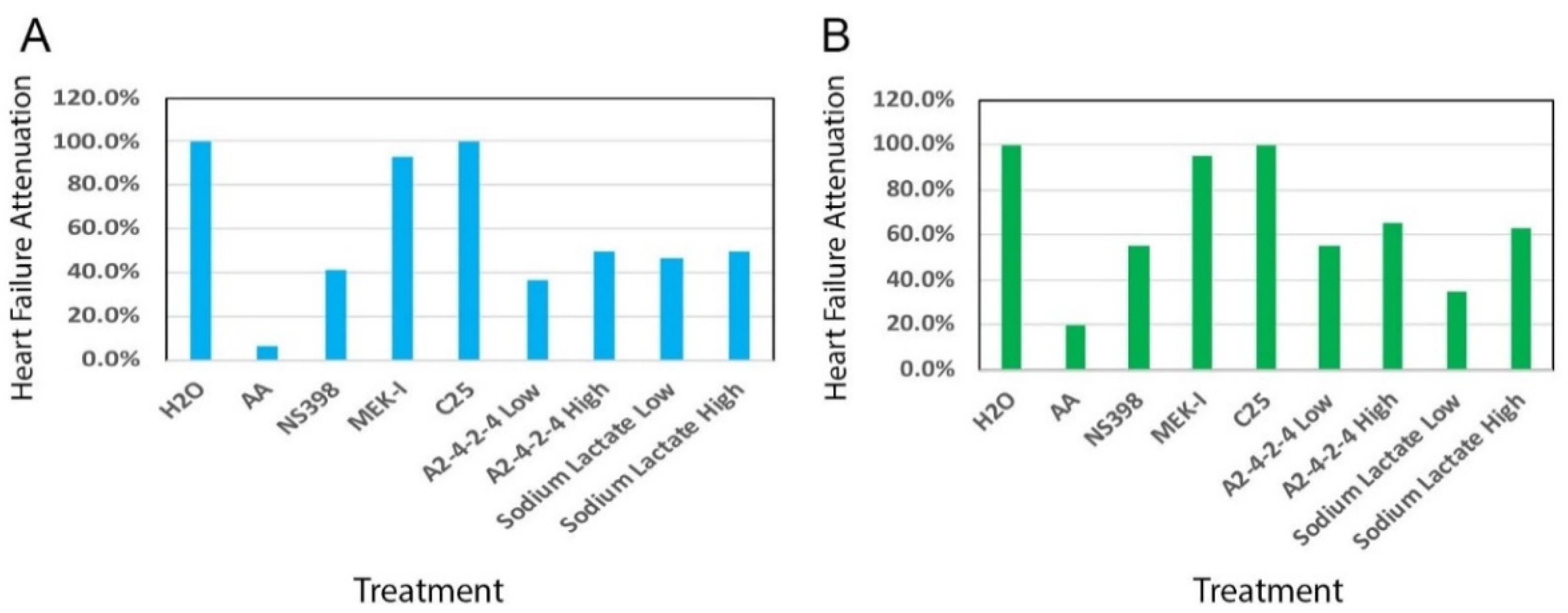
2.3. Comparisons between A2-4-2-4 and Other Drugs
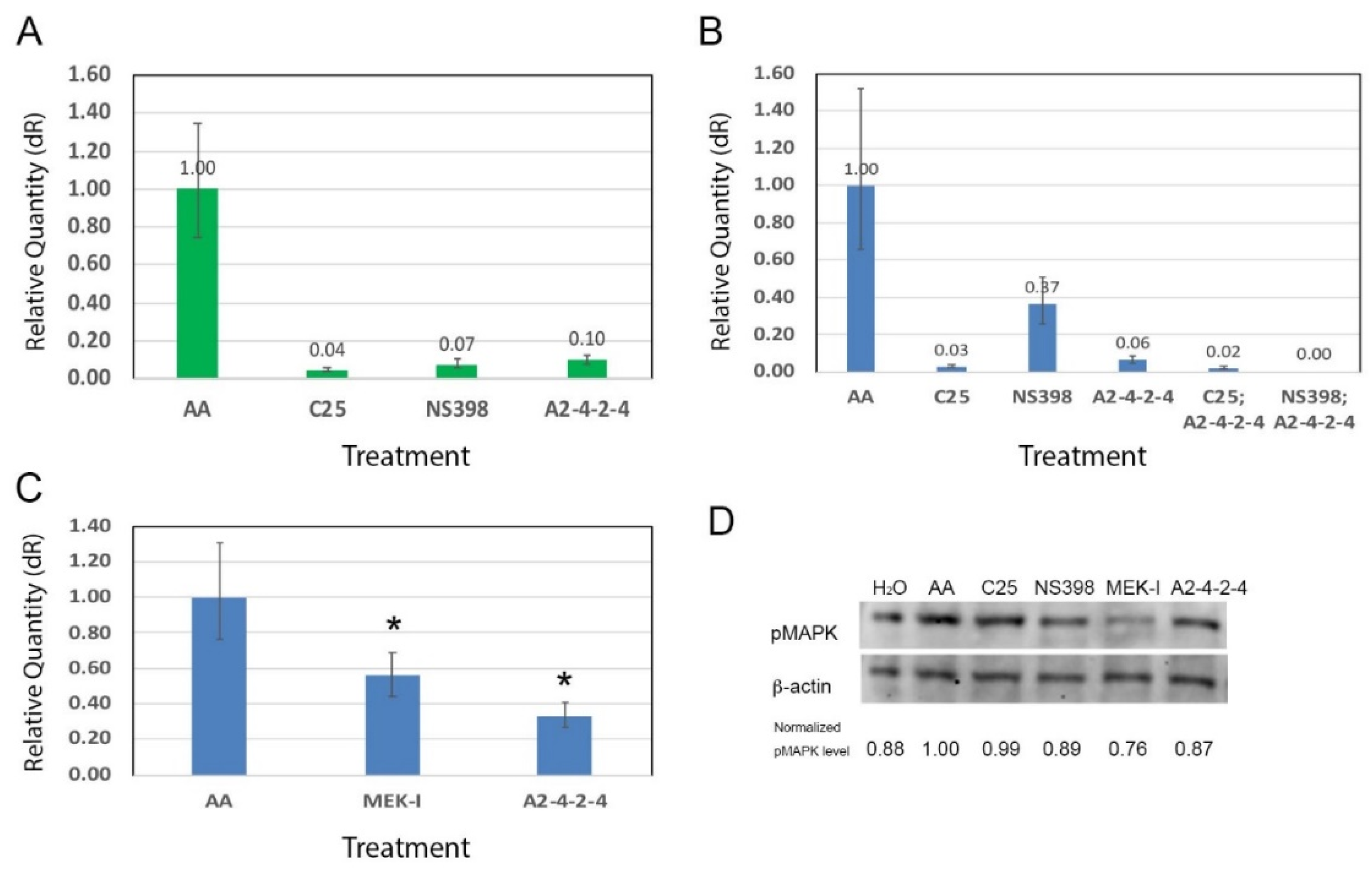
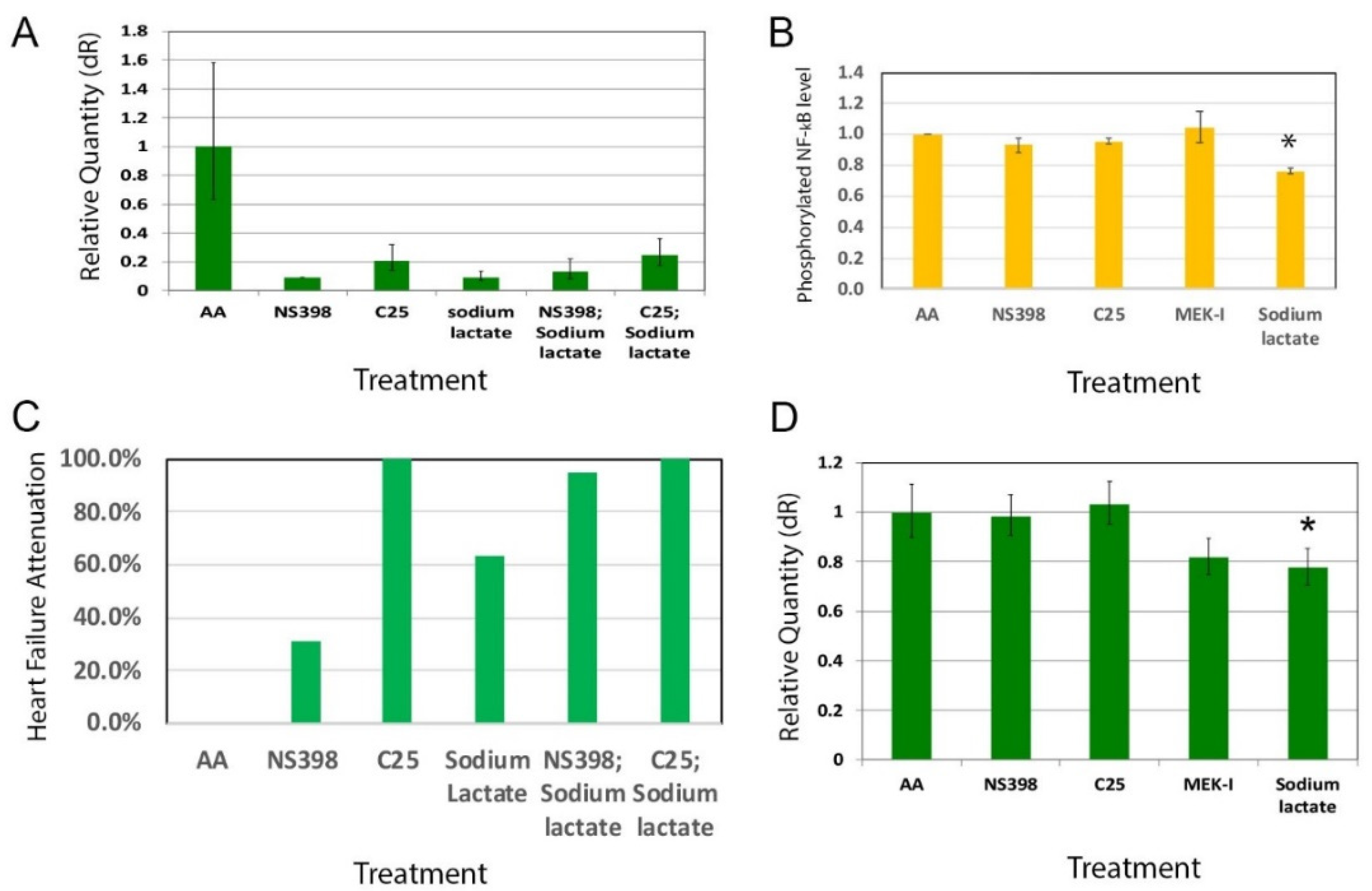
2.4. A2-4-2-4 Attenuates AHF by Inhibiting Inflammation and Cardiac Hypertrophy
2.5. A2-4-2-4 and the MAPK Pathway
3. Discussion
3.1. Drug Discovery for Acute Heart Failure
3.2. Lactate as a Potential Cardiac Protectant
3.3. Molecular Mechanisms of A2-4-2-4/Lactate
3.4. Cellular Mechanisms of A2-4-2-4/Lactate
3.5. Therapeutic Implication
3.6. Advantages of Zebrafish Embryo as a Drug Discovery Model
4. Materials and Methods
4.1. Chemical Extraction and Isolation
4.2. Zebrafish Husbandry and In Vitro Fertilization
4.3. Chemical Preparation and Chemical Treatment on Zebrafish Embryos
4.4. RNA Extraction
4.5. Quantitative RT-PCR (qRT-PCR)
4.6. Western Blot
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| AA | aristolochic acid |
| ADHF | acute decompensated heart failure |
| AHF | acute heart failure |
| cAMP | cyclic adenosine monophosphate |
| COX-2 | cyclooxygenase 2 |
| ESIMS | electrospray ionization mass spectrometry |
| GPR | G-protein receptor |
| HPLC | high pressure liquid chromatography |
| JAK-STAT | Janus kinase-signal transducer and activator of transcription pathway |
| MAPK | mitogen-activated protein kinase |
| MEK-I | MAPK/ERK kinase inhibitor |
| MPLC | medium pressure liquid chromatography |
| NF-κB | Nuclear Factor kappa-light-chain-enhancer of activated B cells |
| NMR | nuclear magnetic resonance |
| NSAID | non-steroidal anti-inflammatory drugs |
| PKA | protein kinase A |
| qRT-PCR | quantitative reverse transcription-polymerase chain reaction |
| Tnnt2 | Troponin T2, cardiac type |
References
- Page, R.L., II; O’Bryant, C.L.; Cheng, D.; Dow, T.J.; Ky, B.; Stein, C.M.; Spencer, A.P.; Trupp, R.J.; Lindenfeld, J. Drugs that may cause or exacerbate heart failure. Circulation 2016, 134, e32–e69. [Google Scholar] [CrossRef] [PubMed]
- Jordan, J.H.; Castellino, S.M.; Melendez, G.C.; Klepin, H.D.; Ellis, L.R.; Lamar, Z.; Vasu, S.; Kitzman, D.W.; Ntim, W.O.; Brubaker, P.H.; et al. Left ventricular mass change after anthracycline chemotherapy. Circ. Heart Fail. 2018, 11, e004560. [Google Scholar] [CrossRef] [PubMed]
- Anker, M.S.; Lena, A.; Hadzibegovic, S.; Belenkov, Y.; Bergler-Klein, J.; DeBoer, R.A.; Cohen-Solal, A.; Farmakis, D.; von Haehling, S.; Lopez-Fernandez, T.; et al. Modern-day cardio-oncology: A report from the ‘Heart failure and world congress on acute heart failure 2018’. ESC Heart Fail. 2018, 5, 1083–1091. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Arrigo, M.; Jessup, M.; Mullens, W.; Reza, N.; Shah, A.M.; Sliwa, K.; Mebazaa, A. Acute heart failure. Nat. Rev. Dis. Primers 2020, 6, 16. [Google Scholar] [CrossRef]
- Bouchez, B.; Fedele, F.; Giannakoulas, G.; Gustafsson, F.; Harjola, V.-P.; Karason, K.; Kivikko, M.; von Lewinski, D.; Oliva, F.; Papp, Z.; et al. Levosimendan in acute and advanced heart failure: An expert perspective on posology and therapeutic application. Cardiovasc. Drugs Ther. 2018, 32, 617–624. [Google Scholar] [CrossRef] [Green Version]
- Nieminen, M.S.; Brutsaert, D.; Dickstein, K.; Dresler, H.; Follath, F.; Harjola, V.P.; Hochadel, M.; Komajda, M.; Lassus, J.; Lopez-Sendon, J.L.; et al. Euro Heart Failure Survey II (EHFS II): A survey on hospitalized acute heart failure patients: Description of population. Eur. Heart J. 2006, 27, 2725–2736. [Google Scholar] [CrossRef] [Green Version]
- Kurmain, S.; Squire, I. Acute heart failure: Definition, classification and epidemiology. Curr. Heart Fail. Rep. 2017, 14, 385–392. [Google Scholar] [CrossRef] [Green Version]
- Follath, F.; Yilmaz, M.B.; Delgado, J.F.; Parissis, J.T.; Porcher, R.; Gayat, E.; Burrows, N.; McLean, A.; Vilas-Boas, F.; Mebazaa, A. Clinical presentation, management and outcomes in the acute heart failure global survey and standard treatment (ALARM-HF). Intensive Care Med. 2011, 37, 619–626. [Google Scholar] [CrossRef]
- Greene, S.J.; Fonarow, G.C.; Vaduganathan, M.; Khan, S.S.; Butler, J.; Gheorghiade, M. The vulnerable phase after hospitalization for heart failure. Nat. Rev. Cardiol. 2015, 12, 220–229. [Google Scholar] [CrossRef]
- Medzhitov, R. Inflammation 2010: New adventures of an old flame. Cell 2010, 140, 771–776. [Google Scholar] [CrossRef] [Green Version]
- Ferrero-Miliani, L.; Nielsen, O.; Andersen, P.; Girardin, S. Chronic inflammation: Importance of NOD2 and NALP3 in interleukin-1 generation. Clin. Exp. Immunol. 2007, 147, 227–235. [Google Scholar] [CrossRef]
- Chen, L.; Den, H.; Cui, H.; Fang, J.; Zuo, Z.; Deng, J.; Li, Y.; Wang, X.; Zhao, L. Inflammatory responses and inflammation-associated diseases in organs. Oncotarget 2018, 9, 7204–7218. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nathan, C.; Ding, Y. Nonresolving inflammation. Cell 2010, 140, 871–882. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cotran, R.S.; Kumar, V.; Robbins, S. Robbins Pathologic Basis of Disease, 8th ed.; Saunders/Elsevier Company: Philadelphia, PA, USA, 2010; pp. 183–258. [Google Scholar]
- Jiang, D.; Liang, J.; Fan, J.; Yu, S.; Chen, S.; Luo, Y.; Prestwich, G.D.; Mascarenhas, M.M.; Garg, H.G.; Quinn, D.A.; et al. Regulation of lung injury and repair by Toll-like receptors and hyaluronan. Nat. Med. 2005, 11, 1173–1179. [Google Scholar] [CrossRef] [PubMed]
- Hoque, R.; Farooq, A.; Ghani, A.; Gorelick, F.; Mehal, W.A. Lactate reduces liver and pancreatic injury in Toll-like receptor- and inflammasome-mediated inflammation via GPR81-medicated suppression of innate immunity. Gastroenterology 2014, 146, 1763–1774. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Anker, S.; von Haehling, S. Inflammatory mediators in chronic heart failure: An overview. Heart 2004, 90, 464–470. [Google Scholar] [CrossRef] [Green Version]
- Pucino, V.; Bombardieri, M.; Pitzalis, C.; Mauro, C. Lactate at the crossroads of metabolism, inflammation, and autoimmunity. Eur. J. Immunol. 2017, 47, 14–21. [Google Scholar] [CrossRef]
- Porporato, P.E.; Payen, V.L.; De Saedeleer, C.J.; Preat, V.; Thissen, J.P.; Feron, O.; Sonveaux, P. Lactate stimulates angiogenesis and accelerates the healing of superficial and ischemic wounds in mice. Angiogenesis 2012, 15, 581–592. [Google Scholar] [CrossRef]
- Shen, Z.; Jiang, L.; Yuan, Y.; Deng, T.; Zheng, Y.R.; Zhao, Y.Y.; Li, W.L.; Wu, J.Y.; Gao, J.Q.; Hu, W.W.; et al. Inhibition of G protein-coupled receptor 81 (GPR81) protects against ischemic brain injury. CNS Neurosci. Ther. 2015, 21, 271–279. [Google Scholar] [CrossRef] [PubMed]
- Houser, S.R.; Margulies, K.B.; Murphy, A.M.; Spinale, F.G.; Francis, G.S.; Prabhu, S.D.; Rockman, H.A.; Kass, D.A.; Molkentin, J.D.; Sussman, M.A.; et al. Animal models of heart failure. Circ. Res. 2012, 111, 131–150. [Google Scholar] [CrossRef] [Green Version]
- MacRae, C.A.; Peterson, R.T. Zebrafish as tools for drug discovery. Nat. Rev. Drug. Disc. 2015, 14, 721–731. [Google Scholar] [CrossRef]
- Huang, C.-C.; Chen, P.; Huang, C.; Yu, J. Aristolochic acid induces heart failure in zebrafish embryos that is mediated by inflammation. Toxicol. Sci. 2007, 100, 486–494. [Google Scholar] [CrossRef] [Green Version]
- Huang, C.-C.; Monte, A.P.; Cook, J.M.; Kabir, M.S.; Peterson, K.P. Zebrafish heart failure models for the evaluation of chemical probes and drugs. Assay Drug Dev. 2013, 11, 561–572. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, W.; Elimban, V.; Nijjar, M.S.; Gupta, S.K.; Dhalla, N.S. Role of mitogen-activated protein kinases in cardiac hypertrophy and heart failure. Exp. Clin. Cardiol. 2003, 8, 173–183. [Google Scholar]
- Delgado, R.M., III; Nawar, M.A.; Zewail, A.M.; Kar, B.; Vaughn, W.K.; Wu, K.K.; Aleksic, N.; Sivasubramanian, N.; McKay, K.; Mann, D.L.; et al. Cyclooxygenase-2 inhibitor treatment improves left ventricular function and mortality in a murine model of doxorubicin-induced heart failure. Circulation 2004, 109, 1428–1433. [Google Scholar] [CrossRef] [Green Version]
- Bueno, O.F.; Molkentin, J.D. Involvement of extracellular signal-regulated kinases 1/2 in cardiac hypertrophy and cell death. Circ. Res. 2002, 91, 776–781. [Google Scholar] [CrossRef] [PubMed]
- Ahmed, K.; Tunaru, S.; Tang, C.; Muller, M.; Gille, A.; Sassmann, A.; Hanson, J.; Offermanns, S. An autocrine lactate loop mediates insulin-dependent inhibition of lipolysis through GPR81. Cell Metab. 2010, 11, 311–319. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Farooq, A.; Hoque, R.; Ouyang, X.; Farooq, A.; Ghani, A.; Ahsan, K.; Guerra, M.; Mehal, W.Z. Activation of N-methyl-d-aspartate receptor downregulates inflammasome activity and liver inflammation via a β-arrestin-2 pathway. Am. J. Physiol. Gastrointest. Liver Physiol. 2014, 307, G732–G740. [Google Scholar] [CrossRef] [Green Version]
- McCrink, K.A.; Maning, J.; Vu, A.; Jafferjee, M.; Marrero, C.; Brill, A.; Bathgate-Siryk, A.; Dabul, S.; Koch, W.J.; Lymperopoulos, A. β-Arrestin2 improves post-myocardial infarction heart failure via sarco(endo)plasmic reticulum Ca2C-ATPase dependent positive inotropy in cardiac myocytes. Hypertension 2017, 70, 972–981. [Google Scholar] [CrossRef] [PubMed]
- Takase, O.; Yoshikawa, M.; Idei, M.; Hirahashi, J.; Fujita, T.; Takato, T.; Isagawa, T.; Nagae, G.; Suemori, H.; Aburatani, H.; et al. The role of NF-κB signaling in the maintenanve of pluripotency of human induced pluripotent stem cells. PLoS ONE 2013, 8, e56399. [Google Scholar] [CrossRef] [PubMed]
- Michael, S.; Achilleos, C.; Panayiotou, T.; Strati, K. Inflammation shapes stem cells and stemness during infection and beyond. Front. Cell Dev. Biol. 2016, 4, 118. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tsukamoto, S.; Shibasaki, A.; Naka, A.; Saito, H.; Iida, K. Lactate promotes myoblast differentiation and myotube hypertrophy via a pathway involving MyoD in vitro and enhances muscle regeneration in vivo. Int. J. Mol. Sci. 2018, 19, 3649. [Google Scholar] [CrossRef] [Green Version]
- Wu, B.U.; Hwang, J.Q.; Gardner, T.H.; Repas, K.; Delee, R.; Yu, S.; Smith, B.; Banks, P.A.; Conwell, D.L. Lactated Ringer’s solution reduces systemic inflammation compared with saline in patients with acute pancreatitis. Clin. Gastroenterol. Hepatol. 2011, 9, 710–717. [Google Scholar] [CrossRef] [PubMed]
- Westerfield, M. A Guide for the Laboratory Use of Zebrafish (Danio rerio). In The Zebrafish Book, 5th ed.; University of Oregon Press: Eugene, OR, USA, 2007. [Google Scholar]
- Huang, W.; Zhang, R.; Xu, X. Myofibrillogenesis in the developing zebrafish heart: A functional study of tnnt2. Dev. Biol. 2009, 331, 237–249. [Google Scholar] [CrossRef] [PubMed] [Green Version]







Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Haege, E.R.; Huang, H.-C.; Huang, C.-c. Identification of Lactate as a Cardiac Protectant by Inhibiting Inflammation and Cardiac Hypertrophy Using a Zebrafish Acute Heart Failure Model. Pharmaceuticals 2021, 14, 261. https://0-doi-org.brum.beds.ac.uk/10.3390/ph14030261
Haege ER, Huang H-C, Huang C-c. Identification of Lactate as a Cardiac Protectant by Inhibiting Inflammation and Cardiac Hypertrophy Using a Zebrafish Acute Heart Failure Model. Pharmaceuticals. 2021; 14(3):261. https://0-doi-org.brum.beds.ac.uk/10.3390/ph14030261
Chicago/Turabian StyleHaege, Elijah R., Hui-Chi Huang, and Cheng-chen Huang. 2021. "Identification of Lactate as a Cardiac Protectant by Inhibiting Inflammation and Cardiac Hypertrophy Using a Zebrafish Acute Heart Failure Model" Pharmaceuticals 14, no. 3: 261. https://0-doi-org.brum.beds.ac.uk/10.3390/ph14030261