1. Introduction
Metabolic syndrome, an epidemic disorder of a global nature, is a cluster of risk factors (i.e., abdominal adiposity, dyslipidemia, high blood pressure, insulin resistance and proinflammatory states) that increase the likelihood of developing diabetes mellitus and cardiovascular disorders. Excessive fructose consumption has been associated with the pathogenesis of metabolic syndrome, as it leads to abdominal obesity and intrahepatic fat accumulation, which in turn may evolve into the non-alcoholic fatty liver disease (NAFLD), a hepatic manifestation of metabolic syndrome. In fact, abdominal obesity is regarded as the major cause of insulin resistance [
1,
2,
3,
4,
5,
6]. Adipose tissue releases free fatty acids that contribute to the development of insulin resistance in liver and muscles. Moreover, it is responsible for the persistent inflammation subsequently augmenting insulin resistance. Additionally, low-grade chronic inflammation affects local adipose physiology and exerts systemic detrimental effects on other tissues [
1,
4,
7]. This is primarily due to macrophage infiltration as well as increased secretion of a large number of pro-inflammatory and decreased secretion of anti-inflammatory substances, like adipokines (adiponectin, resistin), hormones (leptin) and cyto- and chemokines (i.e., monocyte chemoattractant protein 1 (MCP-1), tumor necrosis factor α (TNF-α) and interleukin-6 (IL-6) [
3,
4,
8,
9,
10,
11]. This inflammatory condition may alter the insulin signaling cascade in adipose tissue that includes insulin receptor substrate 1 (IRS-1) phosphorylation at the serine307 residue and, consequently, the decrease of protein kinase B (PKB/Akt) activity and Glucose transporter type 4 (GLUT-4) translocation and glucose transport [
12].
Obesity and insulin resistance together with compensatory hyperinsulinemia exert sympatho-stimulatory effect, sympathetic over-activity and α
1-adrenoceptor activation, which may lead to vasoconstriction and play a role, among others factors, in pathogenesis of hypertension [
3,
7,
13]. Quinazoline-based α
1-adrenoceptors antagonists such as prazosin or doxazosin have been used for years in the treatment of hypertension and they have been shown, in addition to their antihypertensive effect, to moderately improve the metabolic profile of the hypertensive patients, mainly through vasodilatation of blood vessels and improvement of regional blood flow and tissue glucose delivery [
14,
15,
16]. However, in the Antihypertensive and Lipid-Lowering Treatment to Prevent Heart Attack Trial (ALLHAT) study, doxazosin was associated with higher risk of combined cardiovascular disease events, and since then α
1-adrenoceptors antagonists are no longer recommended as a first line therapy of hypertension [
13,
17]. Recent studies have shown that quinazoline-based α
1-adrenoceptors antagonists induce the apoptosis and necrosis of cardiomyocytes and other cell types, which may be responsible, at least in part, for ALLHAT outcomes; however, apoptosis induced by quinazoline-based α
1-adrenergic antagonists was proven to be independent of α
1-adrenoceptor blockade and is associated with the presence of quinazoline moiety. Moreover, apoptosis induction was never shown for non-quinazoline-based α
1- adrenoceptors antagonists such as tamsulosin or urapidil [
17,
18,
19,
20,
21].
MH-76 (1-[3-(2,6-dimethylphenoxy)propyl]-4-(2-methoxyphenyl)piperazine hydrochloride) (
Figure 1) does not contain quinazoline moiety in its structure and may be regarded as urapidil analogue. It is a reversible and competitive antagonist of α
1-adrenoceptors, with no selectivity for a specific α
1-adrenoceptor subtype. We have showed previously that MH-76 has a potent hypotensive activity [
22]. Moreover, it exerts some additional pleiotropic features, such as activation of the endothelial NO—cGMP signaling pathway [
23].
In our previous study we showed that MH-76, in fructose-fed rats with metabolic syndrome, exerted antihypertensive effect, reversed endothelial dysfunction, decreased hyperglycemia and hypertriglyceridemia and reduced insulin resistance and abdominal adiposity. Moreover, MH-76 decreased TNF-α concentration and lipid peroxidation in adipose tissue. Prazosin treatment also exerted an antihypertensive effect and reduced hyperglycemia; however, it had no other beneficial effects characteristic for MH-76 such as improvement of endothelial dysfunction, reduction of insulin resistance and the abdominal adiposity [
24].
The present study was conducted to compare the effects of MH-76 and prazosin on some of the critical points associated with pro-inflammatory state and insulin signaling and their ability to affect adipocytokines in adipose tissue of fructose-fed rats, a well validated model of metabolic syndrome [
25,
26].
3. Discussion
Increasing evidence indicates that extensive fructose consumption, obesity and metabolic syndrome are strongly associated with pro-inflammatory signaling in many tissues including adipose tissue. A high fructose diet triggers the production of inflammatory cytokines, followed by an inflammatory response in adipose tissue, manifested with morphological alterations of adipocytes. These changes further increase visceral adiposity and fat accumulation and cause impairment in insulin signaling in adipose tissue of humans or rats fed a high fructose diet [
4,
11,
27].
MH-76 is a potent antagonist of α
1-adrenoceptors that was confirmed in rat α
1A-, α
1B- and α
1D- [
22] as well as human α
1A-adrenoceptors, and it did not show partial agonist properties. In our previous studies, we showed that MH-76, in addition to its antihypertensive effect, decreased hyperglycemia and hypertriglyceridemia in fructose-fed rats. Moreover MH-76 reduced abdominal adiposity and insulin resistance, as well as decreased TNF-α concentration and lipid peroxidation in adipose tissue. Prazosin treatment although exerted an antihypertensive effect and reduced hyperglycemia did not reduce insulin resistance or abdominal adiposity [
24]. As a continuation of our previous research, in this study we looked deeper and further investigated the positive effect of MH-76 on inflamed adipose tissue in metabolic syndrome since this particular tissue is considered to be the major source of the pro-inflammatory cytokines and a key player in obesity linked disorders [
28,
29].
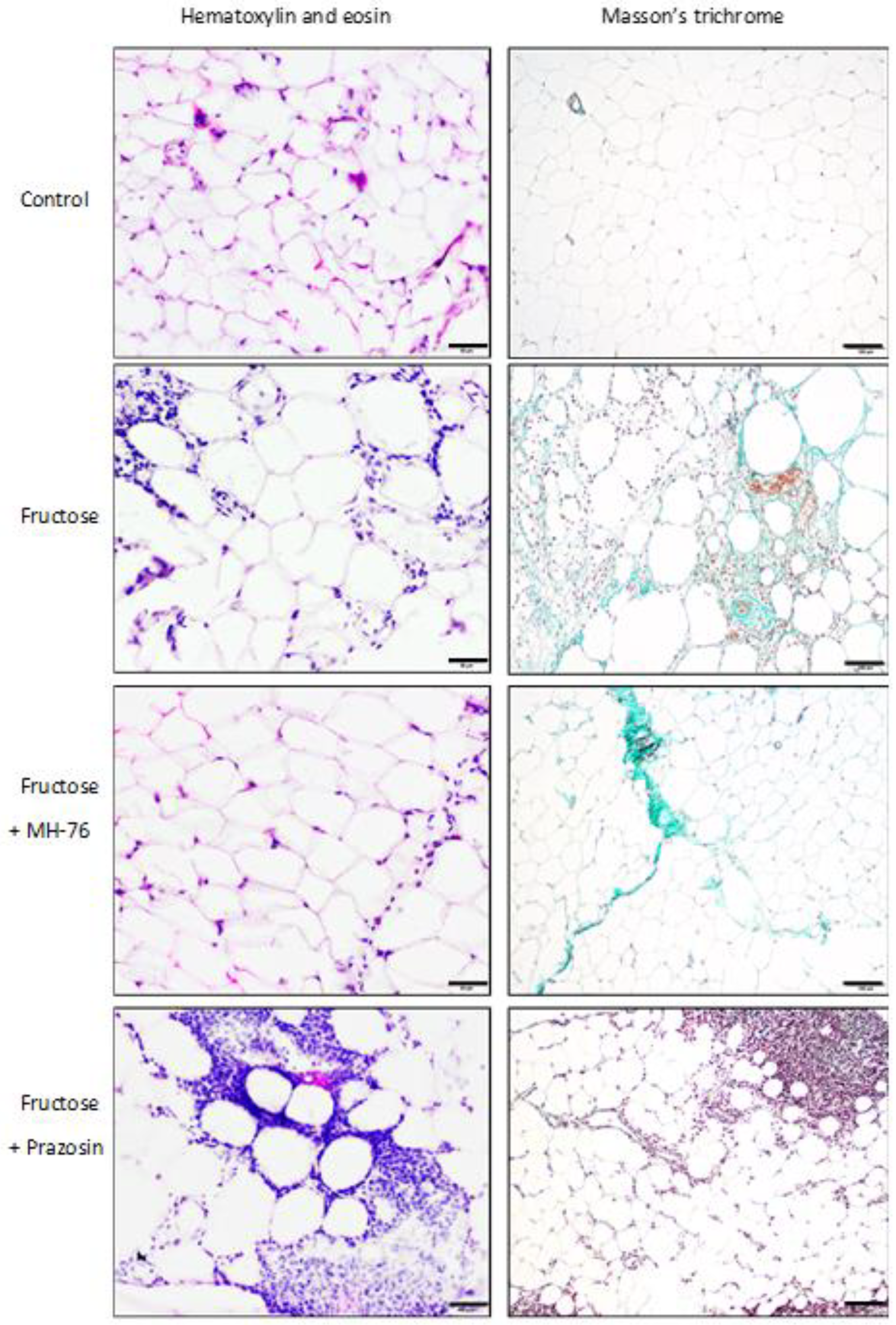
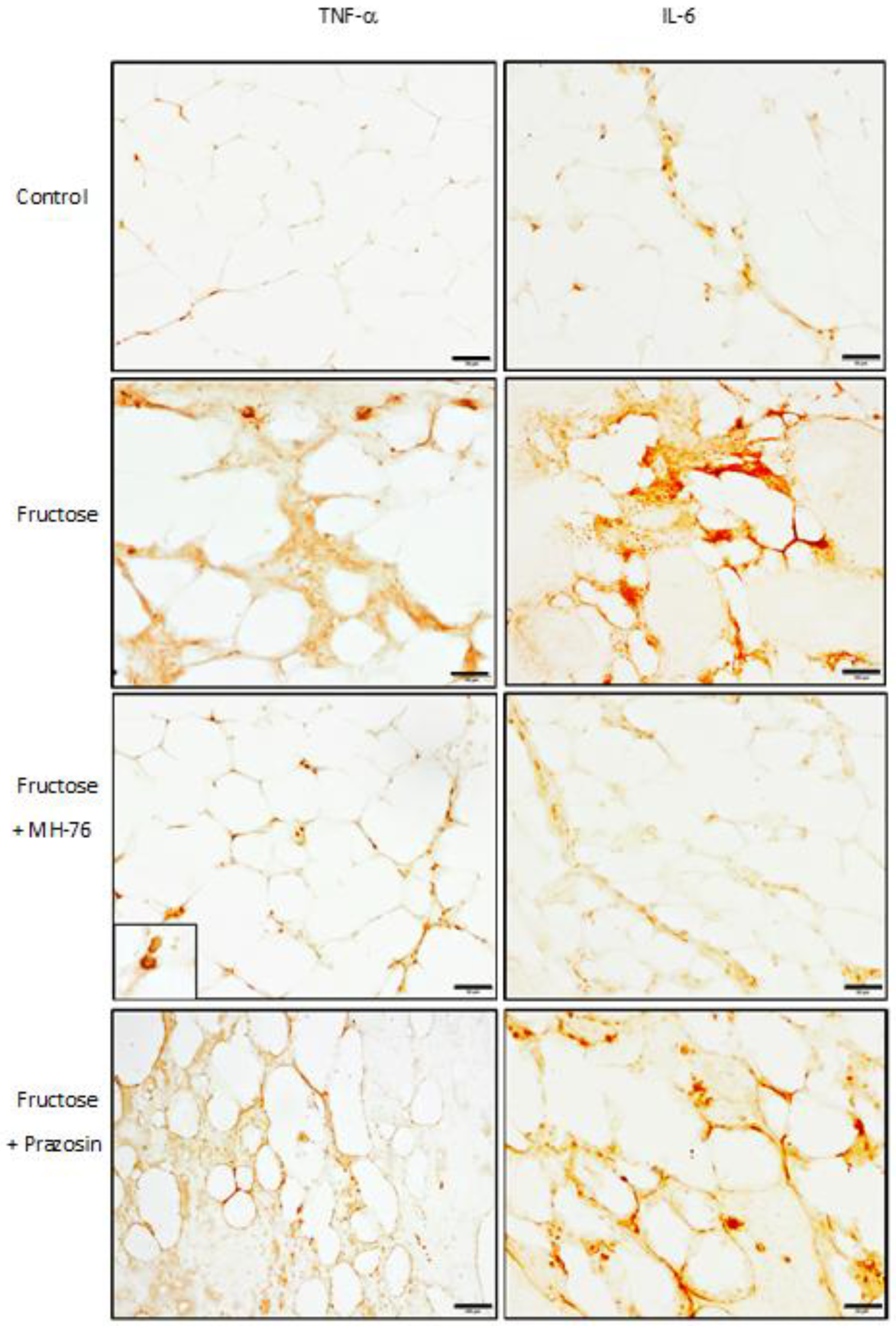
The results from our both previous and current biochemical and histopathological assays clearly indicate that excessive consumption of fructose leads to an increase in both mass and size of adipocytes as well as inflammation and endocrine dysfunction of adipose tissue. In particular, in fructose-fed rats we observed massive infiltration of different inflammatory cells, decreased adiponectine and increased leptine concentrations and increased levels of pro-inflammatory cyto/chemokines (MCP-1, IL-6, TNF-α). Treatment with MH-76 but not with prazosin proved to alleviate some of these conditions.
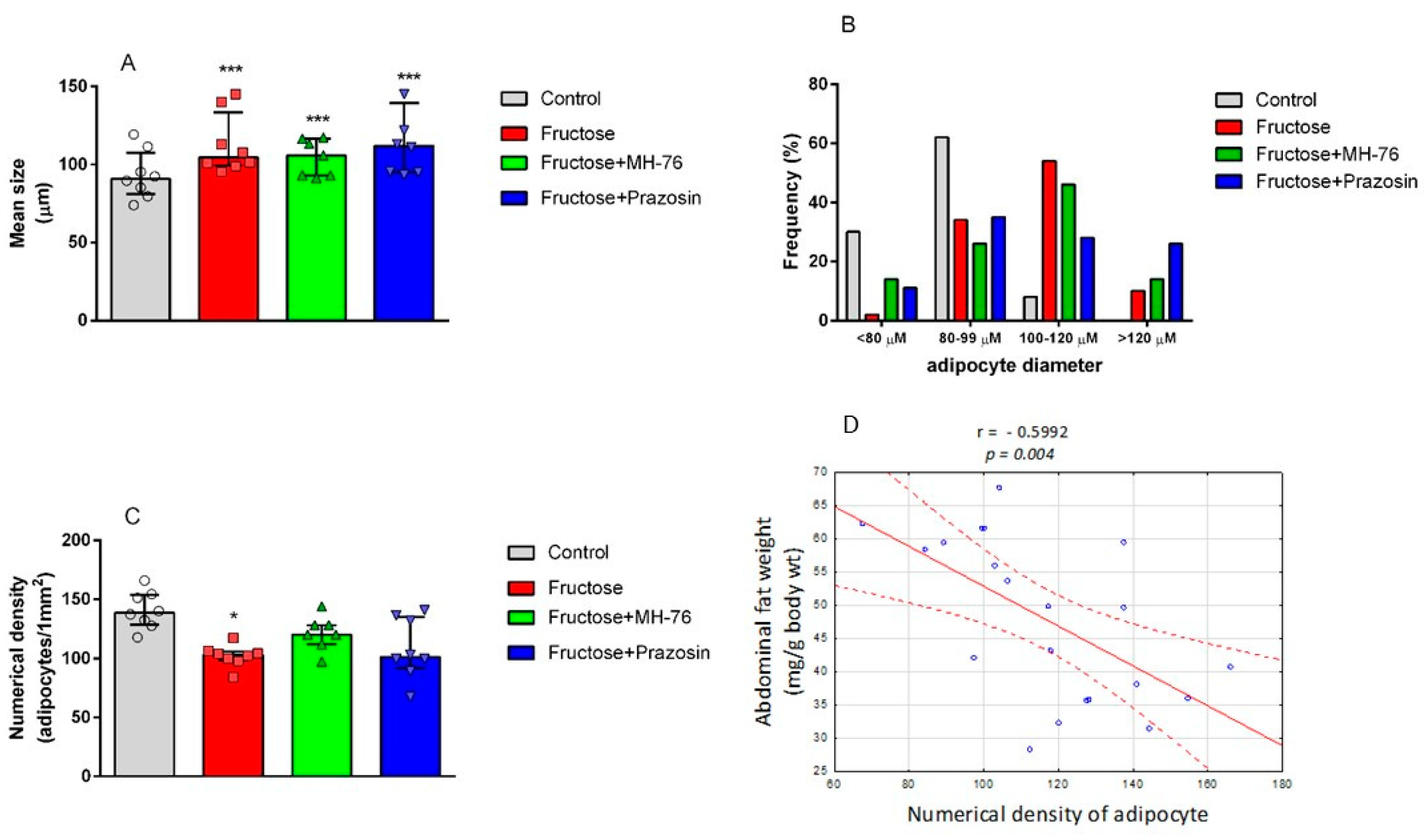
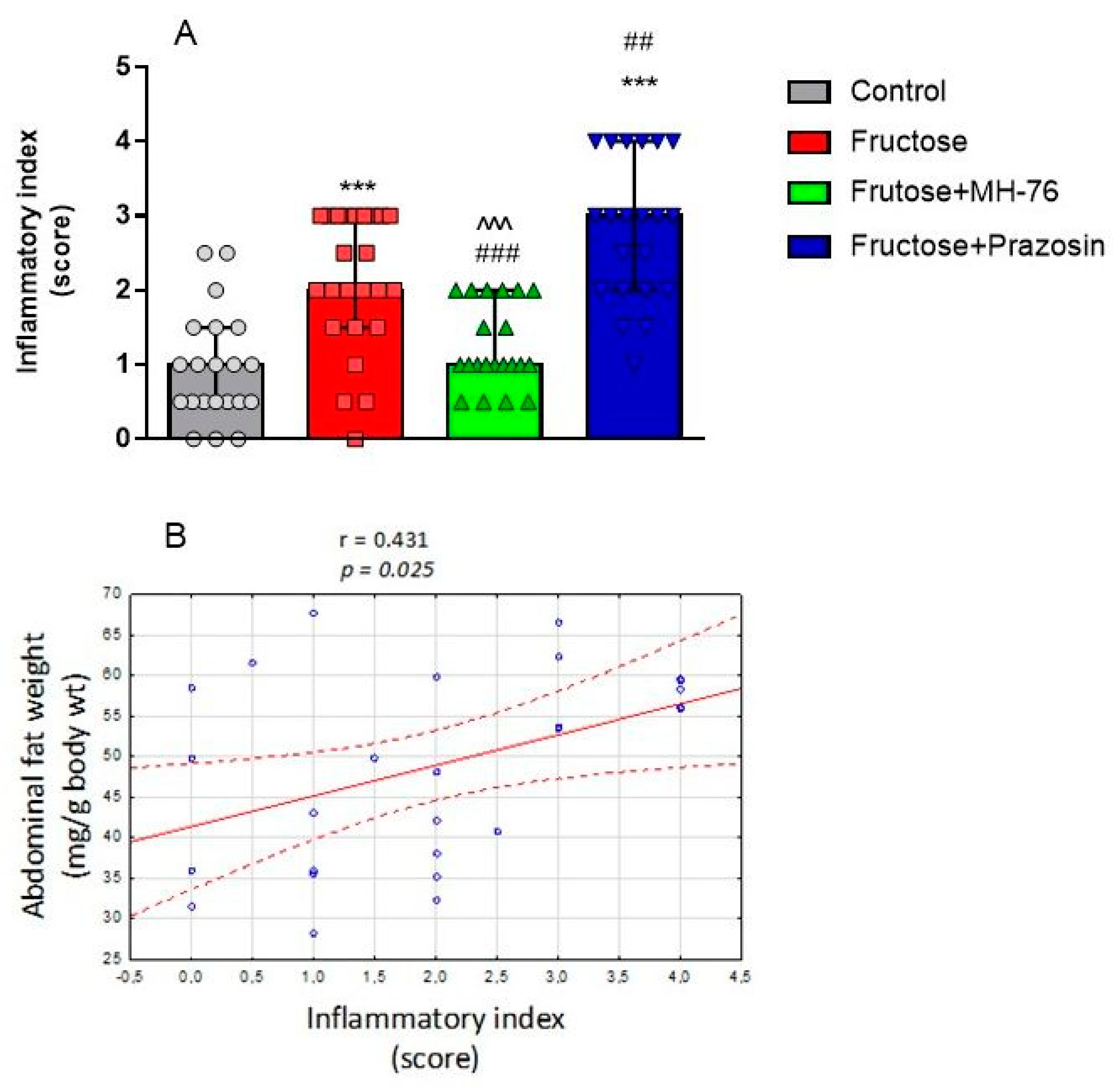
A major characteristic of abdominal obesity is the accumulation of fat deposition and an increase in adipocyte size. Increased fat cell size has been considered to be a marker or even a driver of metabolic disease and adipose tissue dysfunction [
30]. Since treatment with MH-76 reduced abdominal adiposity, we were wondering if that effect could be associated with the reduction in the number of large adipocytes. In MH-76-treated rats, abdominal fat loss could have been associated with the reduction of the number of large adipocytes that, subsequently, caused an increase of numerical density of adipocytes. However, the mean size of adipocytes in rats from MH-76 treated and from the Fructose group was similar. Interestingly, the highest number of very large adipocytes was observed in the Fructose + Prazosin group, which correlated with high expression of pro-inflammatory cytokines. Moreover, we have observed a strong negative correlation between abdominal fat weight [
24] and the numerical density of adipocytes.
In healthy adipose tissue, there are only adipocytes and very few inflammatory leukocytes. Hypertrophy of adipose tissue is usually followed by increased infiltration of immune cells and cytokine secretion [
28]. Subsequently, many different leukocytes including macrophages, mast cells, lymphocytes and neutrophils accumulate in the adipose tissue, contributing to inflammation [
28,
29]. Indeed, histopathological evaluation of adipose tissue from fructose and fructose-fed, prazosin treated rats showed massive recruitment of inflammatory cells, fibrosis and adipocyte hypertrophy. In fact, the most severe inflammation was observed in adipose tissue from Fructose + Prazosin group. Contrary, slight inflammation involving mostly septa was seen in adipose tissue from Fructose + MH-76 rats, which suggested the anti-inflammatory effect of this compound. We have also found a significant positive correlation between abdominal fat weight and inflammatory index.
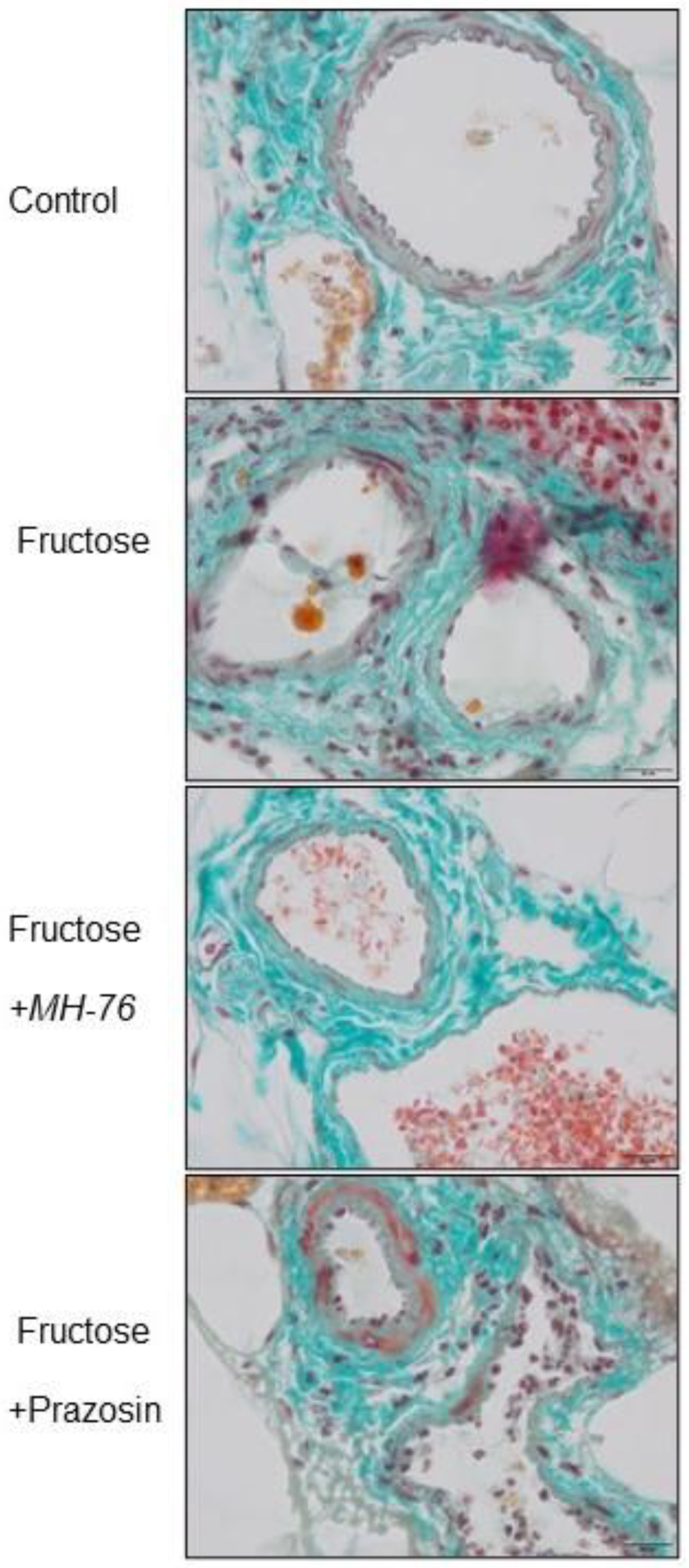
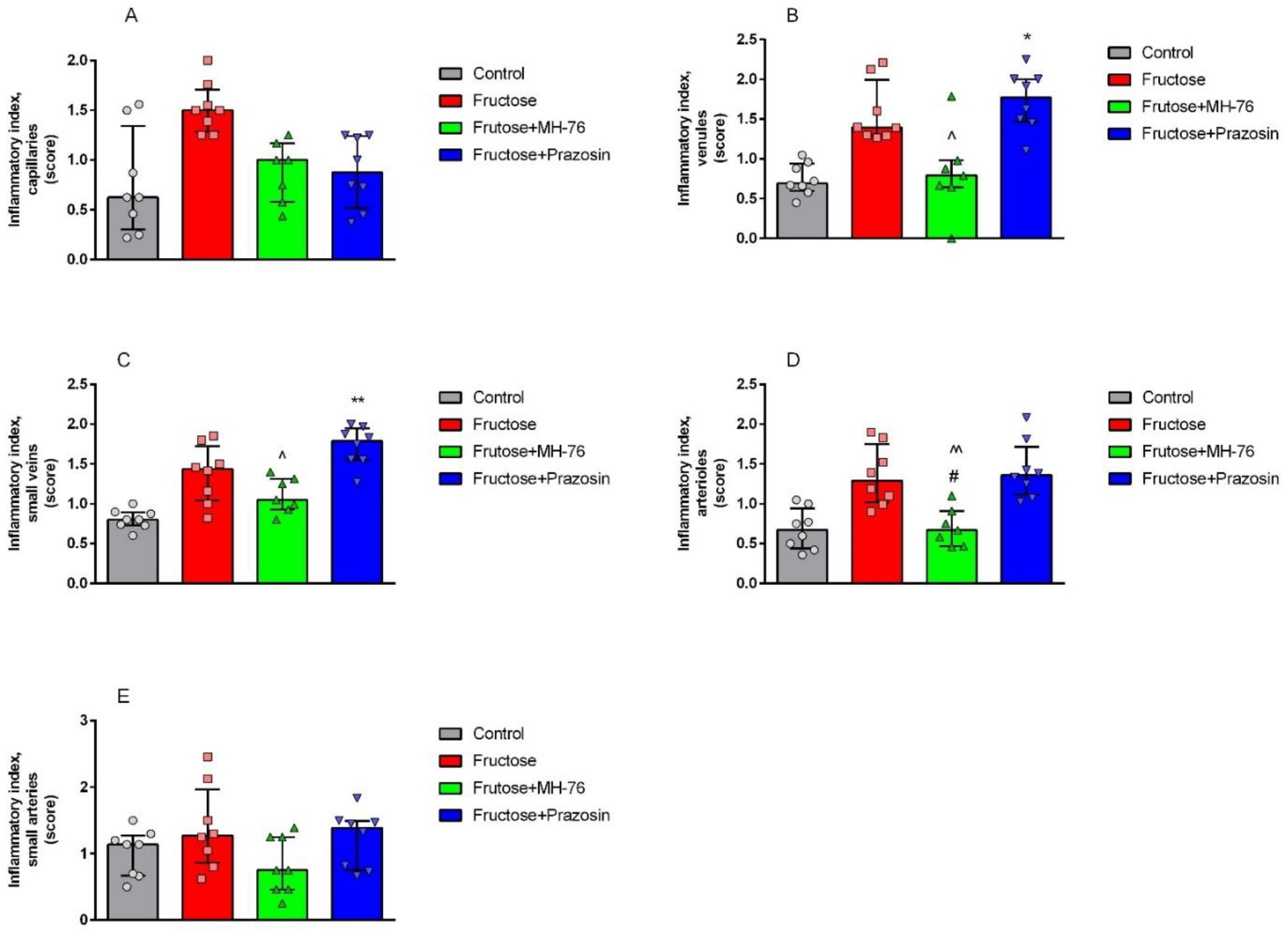
Inflammation is regulated by cytokines, which are produced and released by different inflammatory cells. In obesity, adipose tissue infiltrating leukocytes are probably the main source of locally-produced pro-inflammatory cytokines. However, adipocytes themselves also secrete them in substantial amounts. Cytokines further activate the inflammatory program in neighboring adipocytes, exacerbating inflammation [
29]. Mediators derived from an expanded pool of adipocytes and tissue macrophages are likely responsible for microvascular dysfunction, which further contributes to both the initiation and propagation of the inflammatory response [
31]. The microvasculature responds to these mediators by enhanced expression of the endothelial cell adhesion molecules, that leads to leukocytes peripheralization cascade, and consequently to impaired endothelial barrier function and increased vascular permeability. Such inflammatory phenotype in regard to endothelial damage symptoms, leukocyte adhesion and extravasation, as well as perivascular lymphocyte and macrophage aggregation, was shown in the Fructose group. This pathology was not only preserved, but markedly strengthened in the Fructose + Prazosin group especially in venules and smallest veins. Interestingly, Fructose + MH-76 treated animals showed a normalization of the inflammatory response. We reported previously that MH-76 but not prazosin reversed impaired endothelial vascular relaxation in fructose-fed rats [
24], and that MH-76 exerted endothelial activity through activating NO/sGC/cGMP signaling pathway [
23]. In addition to anti-inflammatory properties, these effects may also account for MH-76 beneficial influence on endothelial integrity in adipose tissue microvasculature found in our studies.
Several works [
28,
32,
33] demonstrated a beneficial effect of eosinophils on the course of inflammation in adipose tissue in animal studies. It has been shown that eosinophils migrating to the septa between adipocytes are the main IL-4 producing cells. IL-4 is involved in the activation of M2 macrophages, which in turn are strongly engaged in tissue repair and reconstruction. Moreover, eosinophils influence the morphology of adipose tissue by modulating the size of adipocytes and intercellular spaces. The presence of eosinophils is therefore essential in the healing process of inflammatory cell infiltrates and also necessary for the proper morphology of adipose tissue. It has been shown previously that in obesity the quantity of adipose tissue eosinophils decreases [
9,
28,
29,
34]. Indeed, in our study we have found that the density of eosinophils was markedly lowered in adipose tissue from Fructose and Fructose + Prazosin animals, whereas the amount of eosinophils in adipose tissue of Fructose + MH-76 rats was markedly higher than in other fructose-fed animals. We have also found that the higher density of eosinophils was accompanied with a higher concentration of IL-4, the phenomenon observed only in adipose tissue of control and fructose-fed, MH-76 treated rats. Data from animal studies show that eosinophils may also exert a beneficial effect on insulin sensitivity in adipose tissue [
32,
33,
34]. It is in line with our studies, where the number of eosinophils was associated with the healing of inflamed tissue and the reduction of insulin resistance. However, it must be pointed out that some human studies do not support a protective role of eosinophils in metabolic syndrome [
35] and the relationship between eosinophils and adipose tissue requires further studies.
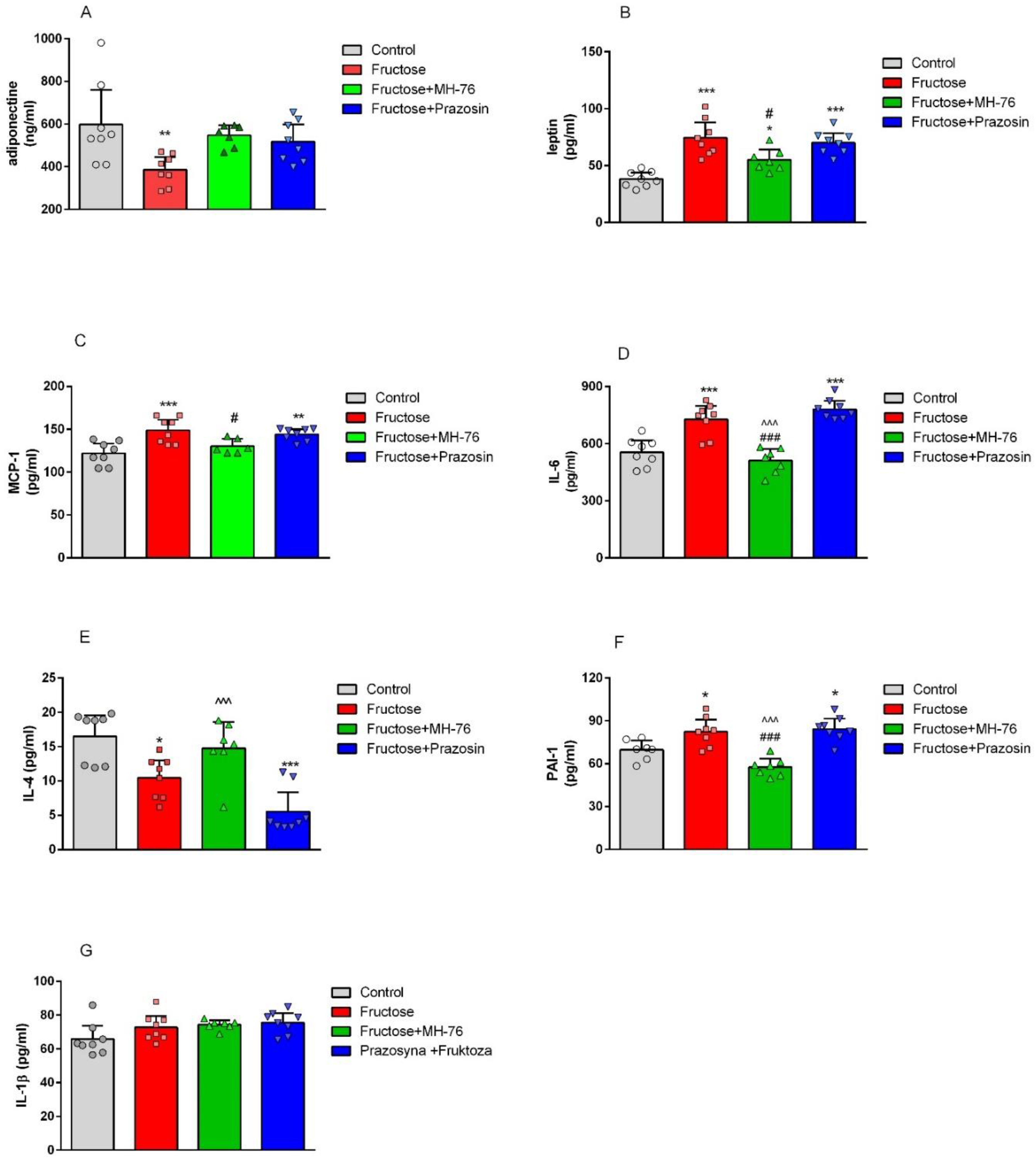
A vital role in the maintenance of insulin sensitivity plays adiponectin, the most abundant adipose-derived protein, that also exerts a pronounced anti-inflammatory effect [
1,
3,
29]. Many studies reported that in abdominal adiposity caused by fructose feeding adiponectin secretion was reduced [
11]. Indeed, in our studies fructose consumption caused a decrease of adiponectin concentration in adipose tissue, which was restored by treatment with both α
1-adrenoceptors antagonists, which what may indicate that this beneficial effect is α
1-adrenoceptor dependent.
Adipocytes are also the source of leptin, a hormone involved in controlling food intake, feeding behavior and energy homeostasis [
11,
28]. High blood levels of leptin play a major role in insulin resistance by inducing the production of IL-6 and TNF-α [
10]. Leptin concentrations are markedly increased in obesity, including fructose feeding model, and positively correlated with adipose mass, indicating leptin resistance [
11,
36]. This again is in line with our studies; in adipose tissue from fructose-fed rats we found higher leptin concentration, which was reduced by treatment with MH-76 but not with prazosin. Leptin receptors are also expressed on vascular endothelial cells and their activation induce the proinflammatory phenotype resulting in an increased capacity to recruit immune cells [
37]. In our studies, MH-76 but not prazosin decreased leukocyte adhesion and extravasation in adipose tissue.
Treatment with MH-76, contrary to prazosin, also reduced the levels of some proinflammatory cytokines such as TNF-α [
24], MCP-1 or IL-6. TNF-α is overexpressed in adipose tissue of obese rodents and humans, which is related to increased infiltration of inflammatory cells, mainly macrophages. Their recruitment involves the synthesis of chemokines such as MCP-1 [
2,
29]. As a response to a high concentration of MCP-1, there is an increased infiltration of monocytes to adipose tissue, where they are differentiated into macrophages responsible for further increase in pro-inflammatory cytokines, resulting in highly inflammatory environment [
28]. Lowering concentrations of TNF-α and MCP-1 in adipose tissue of fructose-fed rats suggests the strong anti-inflammatory activity of MH-76. Adipocytes and macrophages produce also IL-6, which expression is enhanced in obesity [
9,
11]. Indeed, increased tissue expressions of IL-6 and TNF-α were found in macrophages, in some endothelial cells and also in adipocytes; however, they were elevated only in adipose tissue from Fructose and Fructose + Prazosin animals, confirming the anti-inflammatory effect of MH-76.
At the molecular and cellular levels, several signaling pathways have been proposed to link inflammation and insulin resistance [
1,
2]. Typically, as insulin binds to its receptor, it stimulates autophosphorylation of tyrosine residues. Activated insulin receptor (IR) phosphorylates selective tyrosine residues on IRS, and the phosphorylated proteins are able to activate different signaling pathways. The activation of phosphatidylinositol 3-kinase (PI3K), mainly through IRS-1, is preferentially involved in the metabolic effects of insulin.
One mechanism mediating insulin resistance involves phosphorylation of serine residues in IRS-1, inducing its conformational change that in turn can lead to disruption of the interaction between IR and IRS-1 [
8,
34]. There are several serine phosphorylation sites on IRS-1 and their phosphorylation is stimulated by cytokines [
38]. Various kinases, which can be stimulated by TNF-α and IL-6, such as Jun NH2-terminal kinase (JNK), an inhibitor of NF-κB kinase (IKK) and protein kinase C (PKC) are candidates that can mediate insulin resistance [
1,
39]. TNF-α is known to be involved in the disturbance of insulin/IR-initiated signals by activating JNK-1 responsible for phosphorylation of Ser307 in IRS-1 [
1]. Thus, TNF-α induces phosphorylation of IRS-1 at the serine 307 residue, decreasing insulin sensitivity in adipose tissue [
11,
29,
38,
39]. This leads to progressive accumulation of IRS-1 molecules phosphorylated at Ser307, which in turn results in reduced interaction between IRS-1 and the IR or PI3K, and inhibition of insulin signaling [
10,
38,
39]. It also has been shown that IKK-β by phosphorylating inhibitory serine residue on IRS-1 or activating NF-κB, a transcription factor, promotes further inflammatory gene expression and stimulates the expression of inflammatory mediators such as TNF-α or IL-6 [
1]. Like TNF-α, also IL-6 alters insulin sensitivity and impairs insulin signaling through serine phosphorylation of IRS-1 [
1,
29,
40]. All these factors impair insulin signal transduction through IRS and PI3K kinase pathway [
28].
Consistent with these studies, we observed elevated levels of TNF-α and IL-6 as well as enhanced Ser307 phosphorylation of IRS-1 in fructose-fed rats. Treatment with MH-76 markedly reduced the phosphorylation of IRS-1 at Ser307, which may explain the reduction of insulin resistance and hyperinsulinemia observed in our previous study [
24]. The observed modulation of insulin signaling by MH-76 might be attributed to both its anti-inflammatory effect and the reduction of increased levels of cytokines such as TNF-α and IL-6.
Finally, we have found the increased PAI-1 concentration in adipose tissue from fructose-fed animals, which was effectively lowered by treatment with MH-76 but not with prazosin. PAI-1, a serine protease inhibitor, is a marker of impaired fibrinolysis and atherothrombosis. PAI-1 over-expression is associated with atherosclerosis in humans, especially in patients with the metabolic syndrome [
8,
41]. It has been suggested that abdominal adipose tissue is the major source of plasma PAI-1, as it originates from adipose tissue in response to chronically elevated levels of TNF-α and insulin in individuals with metabolic syndrome [
1,
3,
8].
An additional aspect, which also has to be taken into account, is that the doses of prazosin and the MH-76 compound administered to experimental animals were different (0.2 vs. 5 mg/kg b.w.). They were selected based on the affinity of the investigated compounds for α
1-receptor (Ki = 0.1 or 2 nM for prazosin and MH-76, respectively); therefore, the tested dose of MH-76 was 25 times higher than that of prazosin. However, due to the low bioavailability of MH-76 after i.p. administration, the total exposure to this compound was only 3.7 times higher than to prazosin [
24]. Thus, it seems that the compound MH-76 has more beneficial, as compared to prazosin, pharmacological properties at plasma concentrations much lower than expected. Therefore, we have directly measured the concentrations of prazosin and MH-76 in the target organs, namely liver and adipose tissue. We have determined that MH-76, preferably distributed into the abdominal fat, and its concentration in this tissue was almost 70 times higher than that of prazosin (1372.33 vs. 20.83 ng/g). Such pharmacokinetic property might also be involved in the overall beneficial effect of MH-76. On the other hand, studies performed on cells expressing human α
1-adrenoceptor showed that the antagonistic potency of prazosin towards α
1-adrenoceptor is two times higher compared to MH-76 (0.68 nM vs. 1.42 nM). The dose normalized concentration of MH-76 in fat tissue was ca. two times higher than prazosin (
Supplementary Table S1). Therefore, with high probability, we can say that these two drugs produce a comparable degree of α
1-adrenoceptor antagonism in adipocytes at chosen doses.
In the end, it should be emphasized that prazosin not only did not reduce but even augmented the inflammation in the adipose tissue, exacerbating the recruitment of inflammatory cells and fibrosis. Consequently, prazosin did not reduce the elevated concentrations of leptin, TNF-α, IL-6 and MCP-1, and did not improve insulin signaling. Similar results we found in our previous studies, in which in DOCA-salt hypertensive rats, treatment with prazosin, despite the strong blood-pressure-lowering activity, revealed detrimental effects on coronary and renal arteries [
42]. Probably, other factors than α
1-adrenoceptor blockade (i.e., anti-inflammatory effect together with favorable pharmacokinetic parameters) determine high efficacy of MH-76, leading to the effective reduction in inflammatory status in adipose tissue.
Our study has possible limitations; although the performed experiments showed that MH-76 is an antagonist of α1-adrenoceptors, we cannot exclude the possibility of biased agonism or inverse agonism in its molecular mechanism of action. Further studies are required to elucidate the exact cellular mechanisms of MH-76 pharmacological activity.
4. Materials and Methods
Abdominal adipose tissue was collected from fructose-fed rats with metabolic syndrome from our previous experiment. All experimental procedures were conducted in accordance with the ARRIVE guidelines and with the guidelines of the National Institutes of Health Animal Care and Use Committee and approved by the Local Ethics Committee on Animal Experimentation (resolutions no. 338/2017 and 187/2018) in Krakow, Poland [
24]. Male Wistar rats (Krf: (WI) WU) weighing 190–210 g, age 7 weeks, obtained from an accredited animal house at the Faculty of Pharmacy, Jagiellonian University Medical College, Krakow, Poland, were used.
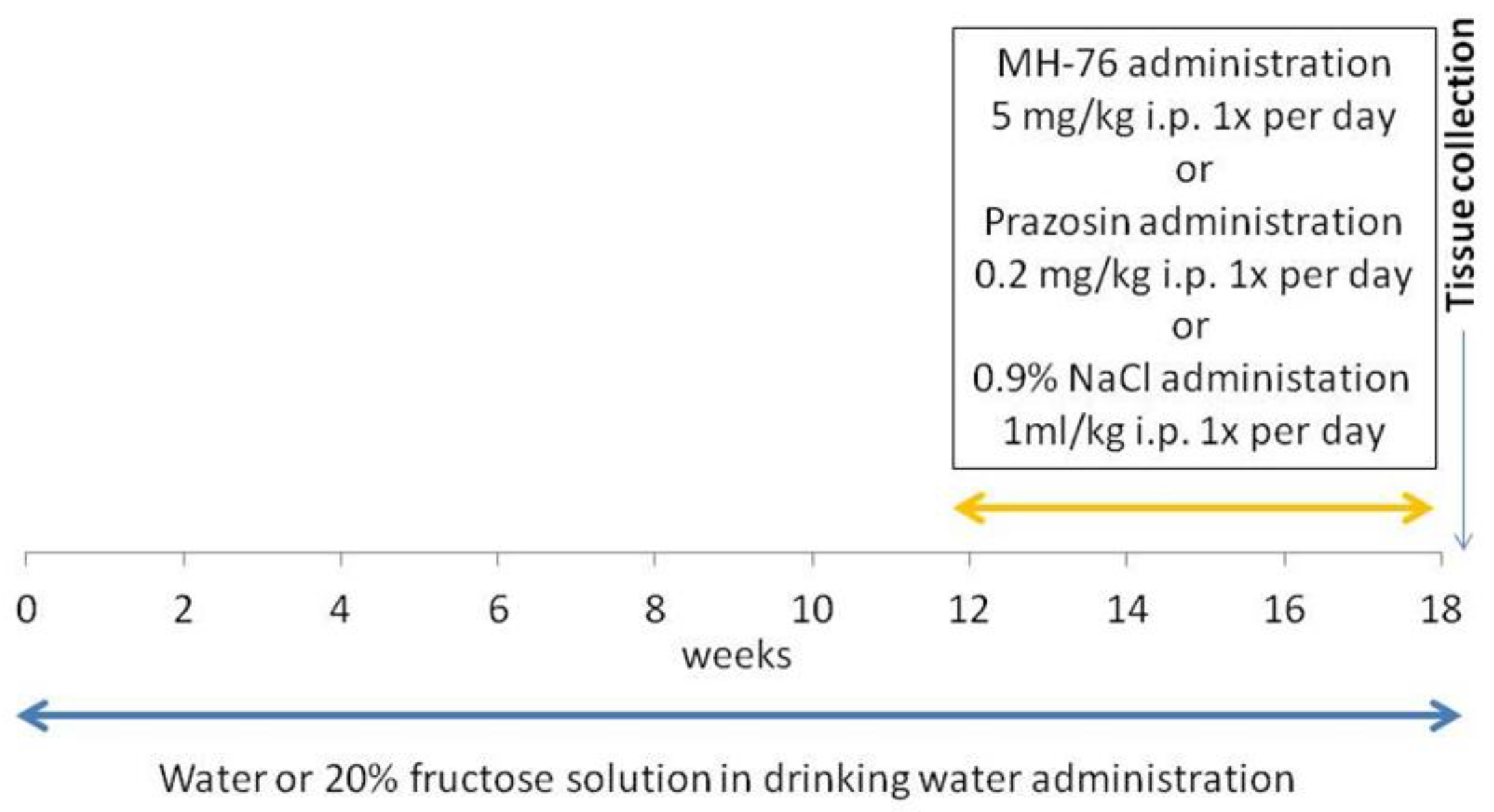
Rats were divided into 4 groups (n = 7–8) and studied for 18 weeks. The groups were as follows:
Control group. Animals received regular diet and water ad libitum for 18 weeks. After 12 weeks, this group received saline (1 mL/kg ip daily) during the last 6 weeks of the experiment.
Fructose: Animals received a regular diet and fructose was administered as 20% solution in drinking water for 18 weeks. After 12 weeks, this group received saline (1 mL/kg ip daily) during the last 6 weeks of the experiment.
Fructose + MH-76: Animals received a regular diet and fructose was administered as 20% solution in drinking water for 18 weeks. After 12 weeks, this group received MH-76 5 mg/kg/day ip during the last 6 weeks of the experiment.
Fructose + Prazosin: Animals received a regular diet and fructose was administered as 20% solution in drinking water for 18 weeks. After 12 weeks, this group received prazosin 0.2 mg/kg/day ip during the last 6 weeks of the experiment [
24].
At the end of experiment, after 16 h fasting but with free access to water, all rats were anesthetized with thiopental (75 mg/kg i.p.), decapitated and abdominal adipose tissue was dissected out and weighed by investigators unaware of the groups allocation. Abdominal adipose tissue was stored at −80 °C until assayed.
Table 2 shows the number of experimental groups, final body mass and abdominal fat index at the end of experimental period [
24].
4.1. Biochemical Assays
Preparation of Tissue Homogenates
The frozen abdominal adipose tissue was weighed and homogenates were prepared by homogenization of 1 g of the tissue in 4 mL of 0.1 M phosphate buffer, pH 7.4, using the IKA-ULTRA-TURRAX T8 homogenizer. Adipose tissue homogenates were next used for biochemical assays.
The concentrations of IRS-1 (E0919Ra, Bioassay Technology Laboratory), pIRS-1 (Ser307) (BC-ER140997, Biocodon Technologies) and IL-4 (E0133Ra, Bioassay Technology Laboratory) in adipose tissue were determined by the ELISA method with the use of commercially available kits. The concentrations of adiponectine, leptine, MCP-1, PAI-1 and IL-6 were determined using commercially available MilliplexTM MAP Kit (RADPCMAG-82K, Millipore, Billerica, MA, USA) according to the manufacturer’s protocol. The quantitative analysis was performed using MAGPIX Luminex analyzer with xPONENT software (Luminex Corporation, Austin, TX, USA).
Levels of analytes were calculated based on the standard curves using spline curve-fitting method and were expressed in pg/mL or ng/mL of homogenate.
4.2. Histopathological Analysis
4.2.1. Quantitative Evaluation of Adipose Tissue
The abdominal adipose tissue samples were fixed in 4% formaldehyde and embedded in paraffin. Sections from each sample were cut (5 μm) and stained using standard staining protocols for hematoxylin and eosin (H&E) and Masson’s trichrome. The histopathological studies were performed in six images per section under 100×, 40× and 20× magnification by a pathologist in blinded randomized sections of the tissues. Images were captured with a light microscope (Olympus BX41, Japan) and color camera (Olympus UC90, Olympus). The digital images were then analyzed using computerized imaging CellSensDimension (Olympus) software. The mean adipocyte size, the size distribution of the cell populations and numerical density (adipocytes/1 mm2 cross section area) were measured. Adipocyte size was determined by microscopic measurement of cell diameter. About 200 adipocytes per case were measured. The numerical density of eosinophils per 0.2 mm2 cross section area was also analyzed. The inflammatory reaction in adipose tissue was classified in terms of the presence of infiltrated immune cells, fibrosis and widening of the interstitial space and was graded on a semiquantitative 4-point scale ranging from 0 to 3+. The mean calculated from independently assessed values was presented as an inflammatory index. An analogous semi-quantitative scale (0—minimal, 1—mild, 2—moderate, 3—marked), and the averaged index was used to examine the inflammation in the microcirculation of the adipose tissue, covering capillaries, arterioles and small arteries, venules and small veins, including vascular endothelial damage, leukocyte recruitment, adhesion and extravasation as well as perivascular lymphocyte/macrophage aggregation. Only microvasculature in the parallel alignment of descending arterioles and ascending collecting venules were taken into account to ensure vessels identification.
4.2.2. Immunohistochemistry
The same paraffin slides of abdominal adipose tissue were immunohistochemically stained for TNF-α and IL-6 using the labelled streptavidin-biotinylated peroxidase system (IHC Select Immunoperoxidase Secondary Detection System, MILLIPORE, Burlington, MA, USA). Deparaffinization, rehydration and antigen retrieval were performed by placing slides in target retrieval solution (Declere, Cell Marque, Rocklin, CA, USA) under pressure cooker boiling for 30 min. After washing in TBS (Tris-buffered saline), endogenous peroxidase activity was blocked by incubating the sections in 3% H2O2/methanol for 15 min. Non-specific antibody binding sites were blocked with normal goat serum (MILLIPORE, Burlington, MA, USA) for 10 min at room temperature. Sections were incubated in a moist chamber at 4 °C overnight and then for 1 h at room temperature with 0.125 μg/mL of a mouse anti-rat monoclonal IL-6 antibody (ab9324, Abcam, Cambridge, UK), or with 1 μg/mL of a rabbit anti-rat polyclonal TNF-α antibody (ab6671, Abcam, Cambridge, UK). After several washes in TBS, the sections were incubated in a biotinylated secondary IgG and then with streptavidin-biotin horseradish peroxidase complex for 10 min at room temperature in accordance with the instructions from the producers. The binding of primary antibody was visualized using 3,3′-diaminobenzidine (DAB, MILLIPORE, Burlington, MA, USA) for 6 min. The slides were dehydrated, cleared with xylene and mounted in DPX. For each antibody tested, a negative control was performed by omitting the primary antibody. Histological slides with the expression of TNF-α and Il-6 were examined under the optical microscope connected to a digital camera under 40× magnification.
4.3. Tissue Distribution Studies
Concentrations of prazosin and MH-76 in adipose tissue and liver homogenates were determined using the LC-MS/MS method, validated according to the FDA guidelines [
43]. Accuracy (percent of recovery) was evaluated as [mean found concentration/theoretical concentration] × 100. Precision was given by the percent relative standard deviations (RSD, CV%). The criteria for acceptability of the data included accuracy within ±15% deviation from the nominal values and a precision within 15% RSD except for LLOQ, where it should not exceed ±20% of CV. Due to the small number of samples analyzed, no stability studies were performed. Liver samples were weighted and homogenized in water at the ratio of 1:4 (w:v). After protein precipitation with acetonitrile (ACN) and centrifugation, the supernatant (10 µL) was injected onto a LC-MS/MS system. Adipose tissue samples were homogenized in acetonitrile and centrifuged, then the supernatant was diluted 100 times with ACN and 10 µL was injected onto a LC-MS/MS system. Chromatography was performed on the XBridge C18 5 µm 3 × 100 mm (Waters, Ireland) analytical column with gradient elution using a mobile phase containing ACN and water with 0.1% of formic acid. Chromatographic system Exion LC AC (AB Sciex, Vaughan, OT, Canada) was connected to the QTRAP 4500 (AB Sciex, Vaughan, OT, Canada) mass spectrometer operated in the selected reactions monitoring mode (SRM), monitoring the transition of the protonated molecular ions to their specific fragments. Two pairs were used for each compound (for MH-76 = 355/205 m/z and 355/136 m/z; for prazosin = 384/247 m/z and 384/138 m/z). Data acquisition and processing were accomplished by the Analyst version 1.7 software.
4.4. Drugs and Chemicals
Prazosin (prazosin hydrochloride) was purchased from Tocris, UK. Compound MH-76 was synthesized in the Department of Bioorganic Chemistry, Chair of Organic Chemistry, Pharmaceutical Faculty, Jagiellonian University [
44],
Figure 1. Melting point of MH-76 was measured on Büchi SMP-560 apparatus (Büchi Labortechnik AG, Switzerland) and was given as uncorrected. The purity of MH-76 was estimated by LCMS system, that consisted of Waters Acquity UPLC coupled to Waters TQD mass spectrometer (Waters, Milford, MA, USA) with electrospray ionization mode, ESI-tandem quadrupole. Acquity BEH C18, 1.7 µm, 2.1 × 100 mm column was used. M.p. 214–216; the exact mass of MH-76 base—354,23. LC-MS [M+H]+ m/z—355.045, 100% purity (
Figures S1 and S2).
4.5. In Vitro Functional Bioassays at Cells Transfected with Human α1A-Adrenoceptor (Aequorin and Luminescence-Based Intracellular Calcium Assay)
Intrinsic activity assay was performed using an Aequoscreen assay according to the manufacturer of the ready to use cells with stable expression of the α1A-adrenoceptor (Perkin Elmer). The Aequoscreen technology uses the recombinant cell lines with stable co-expression of apoaequorin and a GPCR as a system to detect activation of the receptor, following addition of an agonist, via the measurement of light emission. For measurement, cells (frozen, ready to use) were thawed and resuspended in 10 mL of assay buffer containing 5 μM of coelenterazine h. Cells suspension was placed in a 10 mL Falcon tube, fixed onto a rotating wheel and incubated for overnight at RT° in the dark. Then cells were diluted with Assay Buffer to 2000 cells/50 µL. Agonistic ligands 2 times concentrated (50 μL/well), diluted in Assay Buffer, were prepared in 1/2 white polystyrene area plates, and the cell suspension was dispensed on the ligands in 50 μL volume using the injector. The light emission was recorded for 20 s. Cells with antagonist were incubated for 15 min at RT°. Then, 50 µL of agonist (3 times of EC80 final concentration—phenylephrine) was injected onto the mix of cells and antagonist and the light emission was recorded for 20 s.
4.6. Statistical Analysis
Results of this exploratory study are expressed as means ± Δ/2, where Δ is a width of the 95% confidence interval (CI). Statistically significant differences between groups were calculated using one-way ANOVA and the post-hoc Bonferroni multiple comparison test with all possible pairwise comparisons. For histopathological studies, results are expressed by median and inter-quartile range (IQR). For these data, the comparisons among groups were performed using Kruskal–Wallis test by ranks followed by Dunn’s post-hoc test with all pairwise comparisons. The Pearson correlation was used to assess the correlation values between the selected parameters. Differences were considered significant at p < 0.05.


 ,
, 












{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}