
Chromatin Alterations in Neurological Disorders and Strategies of (Epi)Genome Rescue


Dioscuri Centre for Chromatin Biology and Epigenomics, Nencki Institute of Experimental Biology, Polish Academy of Sciences, Pasteur Street, 02-093 Warsaw, Poland
*
Author to whom correspondence should be addressed.
Pharmaceuticals 2021, 14(8), 765; https://0-doi-org.brum.beds.ac.uk/10.3390/ph14080765
Submission received: 6 July 2021
/
Revised: 23 July 2021
/
Accepted: 24 July 2021
/
Published: 4 August 2021
(This article belongs to the Special Issue Epigenetic Drugs)
Abstract
:Neurological disorders (NDs) comprise a heterogeneous group of conditions that affect the function of the nervous system. Often incurable, NDs have profound and detrimental consequences on the affected individuals’ lives. NDs have complex etiologies but commonly feature altered gene expression and dysfunctions of the essential chromatin-modifying factors. Hence, compounds that target DNA and histone modification pathways, the so-called epidrugs, constitute promising tools to treat NDs. Yet, targeting the entire epigenome might reveal insufficient to modify a chosen gene expression or even unnecessary and detrimental to the patients’ health. New technologies hold a promise to expand the clinical toolkit in the fight against NDs. (Epi)genome engineering using designer nucleases, including CRISPR-Cas9 and TALENs, can potentially help restore the correct gene expression patterns by targeting a defined gene or pathway, both genetically and epigenetically, with minimal off-target activity. Here, we review the implication of epigenetic machinery in NDs. We outline syndromes caused by mutations in chromatin-modifying enzymes and discuss the functional consequences of mutations in regulatory DNA in NDs. We review the approaches that allow modifying the (epi)genome, including tools based on TALENs and CRISPR-Cas9 technologies, and we highlight how these new strategies could potentially change clinical practices in the treatment of NDs.
1. Epigenetic Mechanisms Driving Cell Identity and Disease
In a seminal paper from 1942, Conrad Hal Waddington suggested the term “epigenetics” to describe the ensemble of outcomes that result from the functional interactions between genes which produce a mature organism during development [1]. At that time, the mechanisms underlying epigenetics remaindered largely obscure [2]. Evidence accumulated ever since indicates that factors orchestrating chromatin activity, and thereby gene expression, are vital players making up the epigenome allowing to establish and maintain cell identity during development and adult life [3,4]. The more recent definition that we will consider throughout this manuscript describes epigenetic events as the structural adaptation of chromosomal regions so as to register, signal and perpetuate altered activity states [5].
In eukaryotes, DNA associates with a core octamer of histones to form a nucleosome and the array of nucleosomes (the 10nm fiber) constitutes the basic unit of chromatin organization in the interphase [6,7,8,9,10,11,12]. This structural arrangement of DNA has profound consequences from the transcriptional standpoint [13,14,15,16] and the modifications of nucleosomes play a pivotal role in the regulation of gene activity [17]. Multiple layers of regulation of chromatin structure have evolved to ensure that only the proper subset of genes is switched on in a particular cell type [18]. At the most basic level, transcriptional control relies on the binding of sequence specific transcription factors (TFs). Since only a fraction of TFs can bind nucleosomal DNA [19], TF binding frequently requires removal of nucleosomes occluding the TF’s target sequence. TFs recruit transcriptional co-activators or co-repressors [20] which modify the local chromatin structure, thereby allowing for control of the on and off states of genes. Eukaryotic cells are endowed with additional mechanisms, which act seemingly without the direct implication of TFs but involve factors that modify DNA and chromatin proteins, allowing for the induction and the maintenance of stable gene repression, these strategies include for instance position effect (see below [21,22,23,24]) and can operate across multiple generations [25]. Modifications of the epigenetic machinery often result in aberrant functions of the central nervous system (CNS) and contribute to numerous neurological disorders (NDs).
We outline the implication of chromatin modifications in transcriptional regulation. We focus on the implication of the epigenetic mechanisms in NDs. We review the therapeutic approaches that take advantage of specific changes in the epigenome of NDs and discuss the most recent data uncovering the possible mechanistic implication of the genetic risk variants in NDs. Finally, we highlight promising cutting-edge technologies, directed at (epi)genome engineering, which can potentially be harnessed for the design of the future therapeutic strategies to treat NDs.
1.1. Mammalian DNA Cis-Regulatory Elements Classification and Chromatin Signature
TFs can act as activators or repressors, and it is the expression of unique combinations of TFs in each cell type that allows for eliciting and maintaining a defined cell identity [26]. TF binding sites (TFBS) are frequently clustered in the genome and the sequences enriched in TFBS often act as cis-regulatory elements (CREs). CREs can activate gene expression (enhancers and promoters) or silence it (silencers and, indirectly, insulators). Moreover, CREs can be in various positions with respect to their cognate transcription start site (TSS), including in 3’ and 5’ end of the regulated gene, intron of either the target or of an unrelated gene. Enhancers, silencers and insulators are frequently distal (up to megabases (Mb) apart from their target gene); they can be found in either 3’ or 5’ direction with respect to the 5′ end of the promoter they regulate [27] while promoters flank the gene’s TSS. Inactive or poised promoters are enriched in methylated DNA [28,29,30] and heterochromatic histone marks including H3K9me3, H3K27me3 [31] and feature overall histone deacetylation [32,33]. In contrast, active regulatory elements display decreased DNA methylation [34], increased accessibility to nuclease digestion [35,36,37] and enrichment in specific histone modifications including H3K27ac, H3K4me3 and H3K4me1 and binding of TFs rendering these regions more sensitive to nuclease action (Figure 1) [31,38,39,40,41,42,43]. Taken together, genome-wide mapping of chromatin openness and enrichment of H3K4me1, me3 and H3K27ac and CTCF allows for identifying promoters, enhancers and insulators in an unbiased manner.
1.2. DNA Methylation and Hydroxy-Methylation
In mammals, the carbon 5 in the cytosine’s aromatic ring is the major target of methylation (5mCy) [44], even though adenine can also undergo this modification [45]. DNA methylation is primarily deposited at cytosines that precede guanines (CpGs); CpG dinucleotides frequently occur in stretches—CpG islands often located close to gene promoters [29,46,47]. Secluded CpGs are largely methylated across the genome, while the ones within the context of CpG islands are most frequently unmethylated. DNA methylation is implicated in stable gene silencing and is important in the context of X-chromosome inactivation [48,49,50], allele-specific gene expression (imprinting) [51] and transcriptional regulation during development [29,30,48,52]. The reaction of DNA methylation is catalyzed by DNA methyltransferases [53,54] and occurs either de novo [53,55,56,57,58] or during genome replication [54]. 5mCy is recognized by proteins containing the methyl binding domains (MBDs) and the protein MeCP2.
The removal of methyl groups from DNA correlates with transcriptional onset. DNA methylation can be lost passively, during cell division or actively via enzymatic oxidization of carbon 5 within the aromatic ring of 5mCy, a reaction catalyzed by ten-eleven translocation (TET) proteins 1–3 [59]. The resultant 5-hydroxymethyl cytosine (5hmCy) is stable in the cell, non-mutagenic and particularly abundant in the brain [60]. Further oxidation of 5hmCy by TET proteins results in the formation of 5-formylcytosine (5fmCy) and next 5-carboxycytosine (5cmCy) [61,62], which are recognized by dedicated factors [63,64] and rapidly processed in the cell [61,62]. Both 5fmCy and 5cmCy are then excised by thymine DNA glycosylase (TDG)-mediated base excision repair (BER) mechanism [65].
1.3. Histone Modifications
Histone proteins can undergo a plethora of post-translational modifications that are deposited or removed by specialized multicomponent complexes. Histone acetylation and methylation at the unique lysine residues are related to transcriptional regulation (Figure 1) and the presence of a set of modifications at promoters and enhancers is related to their activity (see below). Other histone modifications including phosphorylation, ubiquitination, crotonylation, and sumoylation play diverse roles in chromatin biology [66,67] and will not be reviewed in the context of this manuscript.
Histone methylation is recognized by the royal family domains (including MBT, PWWP, Tudor and chromodomain), plant homeodomain (PhD) finger domains and zinc-finder and WD repeat modules. Acetyl-lysins are recognized by bromo- and YEATS-domains. Chromatin regulatory complexes are typically composed of multiple proteins with domains recognizing modified histones, thereby acting as nodes ensuring crosstalk between epigenetic pathways. Likewise, histone modifications alter three-dimensional (3D) chromatin arrangement [12,16,68,69] and, at a smaller scale, the structure of the nucleosome particle thereby impacting gene activity [67].
1.3.1. Histone Acetylation
Since the discovery of acetylated histones in vivo, it was postulated that this modification is implicated in the regulation of chromatin structure and activity [70]. The discoveries that histone acetyltransferases (HATs) constitute essential transcriptional co-activators [71,72,73] and that histone deacetylases are frequently implicated in the activity of co-repressor complexes [74,75,76,77,78,79,80] have further cemented this link. The acetylation of H3 and H4 is overwhelmingly associated with gene activation, genome-wide analyses of the distribution of acetylated histones revealed localized pattern of enrichment at active regulatory elements including enhancers [31,40,41,81], binding of p300 and CBP histone acetyltransferases is in fact amongst the most reliable markers of tissue-specific enhancer activity [82,83,84]. Histone acetylation is reversed by histone deacetylases (HDACs). As we will discuss below, several epigenetic drugs target HDACs to inhibit their activity, thereby restoring gene expression.
1.3.2. Histone Methylation
The transfer of methyl groups from S-adenosyl-methionine to lysins and arginines in histones is catalyzed by the SET domain in histone lysine methyltransferases (KMTs) including the trithorax-like proteins such as KMT2A and histone arginine methyltransferases (PRMTs). These enzymes deposit up to three methyl moieties on lysins and up to two methyl groups on arginine. Histone methylation is linked with both gene activation and silencing. For instance, histone 3 lysine 4 tri-methylation (H3K4me3) is enriched at promoters of active genes while H3K36me3 marks gene bodies of actively transcribed genes [31,85,86]. On the contrary, H3K27me3, deposited by the HMT EZH2 within that Polycomb Repressive Complex 2 (PRC2) is enriched at promoters of silenced loci. H3K9me2 and me3 are deposited by HMT SUVH1. They are implicated in the silencing of the repetitive sequences and are present at silenced loci within facultative heterochomatin.
Methylation competes with acetylation for lysine residues in histones and the replacement of one kind of modification by another can lead to a fundamentally different functional outcome. For instance, methylation of lysine 27 in histone 3 (H3K27me3) silences enhancers while the acetylation of the same residue is linked with their activation [40].
1.4. Nucleosome Positioning
The formation of nucleosomes can inhibit the binding of TFs and consequently transcription from a promoter [13,14,15]. Several multiprotein complexes are present in the cells and act to displace or evict nucleosomes in an ATP-dependent manner [87]. Based on the composition, chromatin remodelers are divided into four major families: SWI/SNF (switch/sucrose non-fermentable, also called BAF complexes) [88,89,90,91], ISWI (imitation SWI [92,93,94,95,96,97]), INO80 (SWI2/SNF2-related (SWR)), and CHD (chromodomain helicase DNA-binding). What unites these families of complexes is the presence of ATP-dependent DNA helicase from the SNF2 family. All these complexes display tissue specific composition. There are also several orphan families of chromatin remodelers which include the ATRX factor. Chromatin remodeling complexes are critical for proper activity of promoters [98,99], enhancers [100,101,102,103,104,105,106,107,108], and insulators [109,110,111,112]. Their action is related to both repression of CREs [113,114] and cell type specific gene activation. Chromatin remodelers are frequent target of mutations in cancer [115] and the deficiencies in their activity are related to multiple NDs (Table 1).
1.5. Three-Dimensional Structure of Chromatin–The Role of CTCF, Cohesins and Phase Separation in the Buildup of Genome Topology
Cognate pairs of CREs can be located at considerable genomic distances and a substantial fraction of enhancer-promoter pairs are separated by more than 10kb of DNA [220,221]. For instance, limb specific enhancer of the sonic hedgehog gene (SHH), an important morphogen implicated in the development of the nervous system, is located 850 kb away from the SHH promoter [222]. The current model posits that the functional interaction between CREs relies on a physical contact between these elements and one of the open questions in the field is how the cognate CREs are tethered to each other to maintain gene expression patterns. The factors that control associations between CREs in the 3D space are most likely of central importance in health and disease.
Over the past decade, there has been a considerable progress in the understanding of the mechanisms that guide the formation of contacts between genomic regions in the 3D space of the cell nucleus [223]. High resolution, genome wide chromatin conformation capture experiments including Hi-C [224], revealed that at the genomic distances of up to several megabases, genomes fold into domains of high self-contact which were deemed topologically associating domains (TADs [225,226,227], Figure 1). The fact that the cognate CREs frequently reside within the same TAD [34,228,229,230], led to a supposition that these structures constitute functional units of genome organization. Zooming out to larger genomic length-scales, Hi-C revealed that chromosomes are organized into two major compartments the euchromatic compartment A and a more heterochromatic compartment B [224,231], which can be further subdivided into sub-compartments [232]
Nuclear topology arises as a consequence of the interplay between the formation of compartments that bridge regions of similar activity to each other, and the cohesin-mediated loop extrusion [233,234,235,236,237] responsible for the formation of TADs [238,239,240,241,242] in an energy-dependent fashion [243,244,245,246] (Figure 1). These two activities counteract each other [238,239,240,241,242] and the compartmental interactions are disrupted by the loop extrusion machinery. Cohesins bind to DNA and their movement produces a growing chromatin loop (Figure 1). The process stops when cohesin complex encounters a CTCF protein bound to DNA motif that faces them (Figure 1). A stable loop assembles when both loop anchors are occupied by CTCF and the motifs of these two binding sites face each other [247].
Loop extrusion dismantles homotypic interactions that underlie compartments [238,239,240,241] but how compartments assemble remains unclear. Recently, the process phase demixing or liquid phase–liquid phase separation (LLPS) has been implicated in the regulation of nuclear activity and chromatin topology. LLPS occurs when two liquids characterized by distinct physicochemical properties separate. LLPS results in the emergence of droplets or condensates where high concentrations of the separated molecules can be attained [248,249]. LLPS contributes to genome arrangement in various organisms across phyla [250]. Nucleoli [251] and heterochromatin foci [252] for instance form as a consequence of LLPS. Numerous factors that regulate enhancer activity and enhancer-promoter dialogue including mediator complex, TFs, mRNA polymerase harbor intrinsically disordered regions (IDRs) and form condensates in the nucleus [253,254,255,256,257,258]. A disruption of the LLPS affects promoter activity [253,255]. LLPS emerges as an essential property of chromatin-modifying proteins and transcriptional machinery.
2. Genome Activity in Neurological Disorders
Neurological disorders (NDs) constitute a diverse group of conditions that affect the functions of the central and peripheral nervous systems. Depending on the condition, NDs can manifest themselves with mental retardation, intellectual disability, seizures, and as in the case of neurodegenerative diseases (NDGDs) a gradual and progressive decline in cognitive or motor functions. The most widespread and well described NDGDs include Alzheimer’s disease (AD), Parkinson’s disease (PD), Huntington’s disease (HD) as well as amyotrophic lateral sclerosis (ALS), spinal muscular atrophy (SMA), and Friedrich’s ataxia (FRDA). Neuropsychiatric diseases (NPDs) feature alterations in brain physiology and functions. NPDs including schizophrenia (SZ), bipolar disorder (BD), major depressive disorder (MDD), attention deficit hyperactivity disorder (ADHD), specifically affect emotions, and in the context of SZ the feeling and the definition of the self.
NDs can be related to dysfunction of a single gene (Table 1). However, frequently, the genetic underpinnings of NDs are more complex. For instance, there is a strong hereditary component to NPDs [259,260] and many variants have been associated with an increased risk of developing these diseases. However, the mechanisms implying the genetic variants linked to NPDs are frequently multilayered and complex and hence not fully understood. Likewise, the fact that the genetic alterations linked with NPDs are typically of low prevalence amongst patients [113,261,262,263] constitutes a major challenge for the classification and efficient treatment of these conditions.
2.1. NDs Frequently Feature Defective Machinery Establishing and Reading Histone and DNA Modifications
Mis-regulation of gene expression likely constitutes a pivotal event leading to the onset of NDs and their exacerbation [264]. Mutations in chromatin-related factors are significantly over-represented in disorders featuring altered brain physiology and functions [183,262,265,266,267,268,269,270] (Table 1 and references therein). At the same token, genes mutated in NDs, frequently encode factors implicated in the normal development of the nervous system [271,272,273]. Therefore, the understanding of how the chromatin-related factors, which are also linked to NDs, contribute to the normal development of the brain will likely help refine clinical strategies to diagnose and treat these conditions (Table 2) [271,274]. For instance, Rett syndrome, one of the most common causes of mental retardation in women, is caused by a mutation in the X-linked MeCP2, encoding a transcriptional regulator that binds to methylated DNA, and together with HDACs establishes repressive chromatin environment at CpG methylated promoters. Nicolaides–Baraitser and Coffin–Siris syndromes are caused by mutations in SMARCA2 and ARID1B respectively, the components of the SWI/SNF chromatin remodeling complex which orchestrates gene expression (Table 1).
Tightly established protein level of chromatin-related factors is a prerequisite to the normal functioning of the CNS. Erroneous protein dosage of even the same chromatin regulator can lead to distinct phenotypic manifestations. For instance, loss of one of the copies of the genes encoding in CBP or P300 histone acetyltransferases is the underlying cause of the Rubinstein–Taybi syndrome while the duplications in the region encoding CBP lead to a characteristic 16p13.3 syndrome [275,276] featuring unique characteristics including mild mental retardation. Likewise, multiple NDs are not directly caused by mutations in the coding sequence of chromatin factors, but instead feature their transcriptional mis-regulation and distorted nuclear activity. For instance, HDAC1, which catalyzes the removal of acetylation from histone proteins, is significantly overexpressed in the brain of SZ patients [277,278,279]. PD, a late-onset progressive neurodegenerative disorder caused by mutations in α-synuclein which leads to its sequestration and the assembly of the Lewy bodies [280]. Lewy bodies are toxic to neurons, causing their decline. Consequently, PD is characterized by a loss of coordination and decay of motor skills. The formation of Lewy bodies also correlates with a sequestration of DNMT1 to the cytoplasm, subsequent decrease in nuclear DNMT1 and global hypomethylation of the genome in neurons [281]. More recently, genome-wide histone acetylation analysis revealed a general promoter hyperacetylation in the prefrontal cortex of patients suffering from PD [282]. HD is another example of a late-onset progressive NDGD. HD is caused by an expanded polyglutamine repeat sequence in the huntingtin protein (HTT). The mutant HTT, unlike its wild-type counterpart, enters the nucleus where it binds and inhibits HATs, leading to a decrease in H3 and H4 histone acetylation, and thus to a global transcriptional mis-regulation including the silencing of neuronal genes [283,284]. HD features changes in DNA methylation; in comparison to wild type cells, polyglutamine-expanded HTT (STHdhQ111/Q111) cell line displays an increased DNA methylation at more that 21 thousand sites including promoters of essential to neuron differentiation, neurogenesis and transcription [285]. DNA demetylation in this HD model is much less pronounced and affects around 10,000 sites [285].
AD is one of the most common NDs and features a progressive impairment of cognitive functions. The formation of amyloid plaques that contain beta-amyloid protein is one of the hallmarks of AD and is mediated by the enzymatic cleavage of the amyloid precursor protein (APP) by β-secretase (BACE1). The overexpression of APP might be caused by a hypomethylation of its promoter’s DNA [286]. APP can also be elevated due to a less direct, epigenetic effect. For instance, promoter hypomethylation of transmembrane Protein 59 (TMEM59) in AD leads to its upregulation. As a consequence, APP cleavage by α- and β-secretase is inhibited, and APP deposition enhanced [287]. Remarkably, AD features a global activation of chromatin. Multi-omics analysis of AD and healthy brain samples revealed upregulation of chromatin factors including HATs (CBP and p300) and TRRAP (SAGA–ATAC complex subunit), HDACs (SIRT1 and HDAC4), histone methyltransferases (CXXC32) and histone demethylases (JMJD6) and gain in H3K27ac and H3K9ac at the promoter of CBP and CTCF [288,289,290,291]. Likewise, fine mapping of regulatory element chromatin landscape in individuals at different stages of AD revealed frequent DNA hypomethylation at enhancer elements linked with AD and potential mechanisms driving AD including the upregulation of β-secretase though transcriptional gain of DSCAML1 [292]. Altogether, deregulation of gene expression and chromatin signature are most likely essential for the development and progression of NDs.
2.2. Chromatin Topology Is Related to Neuronal Function and Can Be Affected in NDs
Mutations in CTCF and in the components of the loop extrusion machinery including cohesins and auxiliary factors that regulate the loading and the stability of the cohesin complex on chromatin, manifest themselves with multiple congenital defects including but not limited to mental retardation (Table 1). Likewise, as we saw, genetic alterations of factors impacting promoter-enhancer dialogue including mediator and histone acetyltransferase P300 result in neurological syndromes (Table 1). To harness the potential of these observations in clinical practice, we need to deepen our understanding of the interplay between the topology of the genome and transcriptional regulation. One important aspect of this question is to define how chromatin arrangement contributes to the action of enhancers and promoters and how insulators impact these transactions [223,293]. One of the ways to impact the 3D distance between genomic regions is to establish an extrusion-mediated CTCF-CTCF loop. Indeed, CTCF-mediated loops have been related to the control of promoter-enhancer contacts in various contexts [243,294,295,296,297,298] including the control of protocadherin gene expression during neural development [299]. Furthermore, CTCF binds to numerous genes underlying neuronal functions; CTCF knockdown in the hippocampus leads to transcriptional mis-regulation of over 700 loci correlating with an impairment in learning and memory in the mouse model [300]. Whether CTCF acts to impact promoter-enhancer loops at these loci remains open. In murine models, neuronal activity elicits the formation of promoter-enhancer loops at a subset of regulated loci [301,302], and leads to multiple changes in compartmental interactions. Genetic ablation of cohesin affects this process and leads to a reduction of promoter enhancer loops [301] and the removal of cohesins correlates with impaired motor learning [301]. These observations indicate that loop-extrusion-mediated chromatin topology plays an active role in the regulation of higher-level brain functions.
CTCF is the only known mammalian insulator to date and the insulatory functions of CTCF rely on the presence of cohesins [303,304]. Consistent with a convergent role of CTCF and cohesins in the formation of TADs, a disruption of either of the two mechanisms leads to loss of TAD insulation [238,239,240,305,306,307]. CTCF loss can cause a misalignment of transcriptional enhancers and an aberrant activation of nearby genes [294,308]. Alteration of CTCF binding can be caused by the mutation in the CTCF gene, changes in the level of CTCF expression level or by the modification of its binding site architecture. The latter can intervene at loci implicated in NDs. Modeling of the neuronal and astrocytic lineage function using induced pluripotent stem-cell models revealed an overrepresentation of genetic variants related to NPDs at the anchors of loops assembled specifically in iPS-derived neurons suggesting an important role of chromatin architecture in neuronal functions [309]. Whether the same phenomenon occurs in the brain of diseased individuals remains to be assessed but recent observations strongly link CTCF function to brain development and NDs [310]. At the 7p21 locus associated with neurodegenerative disease frontotemporal lobar degeneration, a mutation that precludes the binding of CTCF results in an altered chromatin topology featuring an increase in the regulatory interactions of the promoter of TMEM106B likely leading to an enhanced expression of TMEM106B and cytotoxicity [311]. The fact that disease risk variants related to NDGNs are enriched with CTCF binding events [311] leads to an assumption that there will be many more examples of loci that are transcriptionally mis-regulated as a consequence of aberrant recruitment of CTCF.
2.3. Next Generation Brain Regulome Maps to Link Genetic Variants to Essential Loci and Identify New Genes Underlying ND
There are several strategies that allow to uncover genes and CREs that are likely to contribute to NDs. Genetic linkage mapping can give hints as to the mechanisms of the NDs, help early diagnosis, and possibly reveal the most suitable intervention points for clinical practice. Genome wide association studies (GWAS) allow the discovery of common genetic variants, including single nucleotide polymorphisms, insertions, deletions, or duplications, which underlie traits, including disease susceptibility. GWAS relies on sequencing of the genome of the diseased and healthy individuals and a subsequent computational identification of the regions in the genome that segregate with a trait or a disease. Remarkably, while numerous GWAS hits affect protein-coding regions [312], the majority of disease risk variants, including loci linked with NDs, are often non-coding and frequently feature regulatory element chromatin signature and activity [313,314,315,316,317,318,319] further highlighting the contribution of transcriptional regulatory mechanisms to the etiology of NDs. Regulatory mechanisms that drive disease-related changes in gene expression can in turn be identified through the analysis of the expression quantitative trait loci (eQTLs) in which the transcriptomes of healthy individuals and patients are compared in the function of the genetic changes in the intergenic CREs.
Having established the cartography of CREs affected in NDs, the next step is to functionally link these elements to genes and pathways that can be targeted clinically. This task is challenging and addressing it constitutes one of the most outstanding goals in the field [320]. Several international consortia, including the Psychencode (https://www.nimhgenetics.org/resources/psychencode, accessed on 23 July 2021), ENCODE and GTEx [321] have been established to determine the implication of the regulatory element mutations and transcriptional deregulation in NDs. Multiple exciting examples of how functional genomics, by combining chromatin activity and architecture mapping, can help unravel the mechanisms of action of genetic hits related to chosen NDs highlight the great power of multiomics to address ND and suggest possible clinical intervention points.
2.3.1. Comparison of Wild Type and ND Chromatin Can Help Identifying Essential Pathways Related to NDs
Essential genes and regulatory elements underlying NDs can be identified via a direct comparison of healthy and disease brain tissue. For instance, AD-specific H3K27ac peaks frequently overlap AD GWAS hits with prominent examples, including putative enhancers mapping in the vicinity of MAPT, PSEN2, genes controlling amyloid-beta and tau biology [289], as well as genes related to synaptic transmission and immune response [322]. Likewise, single nucleotide polymorphisms associated with PD overlap enhancer elements at the α-synuclein (SNCA) and PARK16 genes, both which are implicated in the neuropathology of PD [323]. Genetic replacement of a wild-type SNCA enhancer, by its version displaying a significant association with PD, results in an increased level of SNCA expression in neurons derived from the genetically modified iPS cells [324]. Sequence analysis of the risk variant suggests that the up-regulation of SNCA in PD might arise as a consequence of a loss of binding of the EMX2 and NKX6-1 TFs that repress SNCA expression in neurons [324].
Due to its heterogeneity, SZ is an extremely challenging and not well understood ND; hence, there is an urgent need to determine which genes are essential to this syndrome. Like other NDs, SZ features significant changes in gene expression [325], which in part could be explained by an aberrant activation of key enhancers [326,327]. For instance, a risk allele within the first exon of the NDUFA6 gene, disrupts the production of the NDUFA6 protein by inducting nonsense-mediated decay of its messenger RNA. Remarkably, apart from acting on the NDUF6 transcript, this mutation also hampers the binding of YY1 transcriptional regulator, which in turn has leads to a reduced expression of NAGA, a gene located 32 kb upstream of the enhancer [328]. Interestingly, NAGA itself is a SZ risk gene, uncovered through eQTL analysis. Hence, the analysis of 3D chromatin arrangement helped identifying the possible mechanism that drives NAGA deregulation in SZ [328]. Altogether, the comparison of healthy and diseased tissue can illuminate on the mechanisms driving NDs. However, this approach cannot unveil the longitudinal effects of generic mutations that intervene at a restrained time window during brain development. Hence, it will miss relevant molecular events and markers for diagnostic purposes allowing to manage the disease at its earliest stages. One can address this question by functional analysis of the impact of GWAS on brain biology.
2.3.2. From GWAS to the Causal Gene
Having only the information about healthy tissue, how do we navigate the ocean of GWAS to identify the most interesting targets for follow up studies and clinical practice? There are several approaches that we can harness to accelerate the discovery in this domain. These include computational approaches that allow to prioritize variants for functional follow-up studies based on Bayesian fine-mapping [329] and regression analysis [330]. Likewise, summary association statistics inferred through GWAS and transcriptomic effects from eQTL analysis can be considered jointly, which enhances the power to detect interesting candidate loci [331,332]. Another approach calls to prioritize GWAS hits by assessing their impact on TFBS, thereby greatly facilitating the identification of most likely candidates that orchestrate transcriptional mis regulation in ND [333]. Comparative sequence analysis can also reveal fundamental mechanisms underlying NDs. Exemplifying this, a relatively recent global analysis of risk variants of SZ that specifically affect TFBS, revealed 132 loci whereby the sequence motif recognized by a TF is disrupted by the disease-linked mutation. Interestingly, a substantial fraction of these sequence changes affects the binding sites of CTCF and cohesins [326] strongly suggesting an essential role of chromatin topology in SZ. The classification and the identification of the most promising variants of CRE can be substantially aided experimentally. The impact of a mutation in a CRE can be then tested by assessing how it affects the binding of a TF in for instance Electromobility Shift Assay (EMSA). The impact of the genetic mutation on the CRE activity can be tested directly by reporter assays whereby the CREs are tested for their capacity to regulate the expression of a reporter gene (luciferase or a fluorescent protein) controlled by a weak promoter. Reporter assays can be used to test individual sequences or thousands of sites simultaneously like for instance in the massively parallel reporter assays (MPRA) [334,335,336].
Enhancers are typically at vast genomic distances from promoters and do not necessarily regulate their closest gene. When active, enhancer-promoter pairs can display an increased spatial proximity and the chromatin conformation assays can help to link them to each other [34,220,337,338,339,340,341]. Using PLAC-seq, which allows to identify the interactions between genomic sequences at high regulation, Nott et al., uncovered the most likely enhancer-promoter pairs in several major cell types in the human brain [342]. One of the essential conclusions of this study, somewhat in agreement with expectations [220,343], is that enhancers frequently do not simply act upon the closest gene. For instance, an intronic GWAS AD risk variant within the gene SLC24A4 that overlaps a putative enhancer tends to interact with several genes located >200 kb in the 5′ direction (CPSF2, ATXN3 and TRIP11). Strikingly, it does not form contacts with the promoter of SLC24A4 which was assigned to it by GWAS analysis.
The analysis of the 3D conformation of chromatin in developing neurons can highlight new functions of regulatory elements linked to NDs at the early stages of brain development. In SZ, chromatin conformation capture connected regulatory elements mutated in patients to new genes including acetylcholine receptors. This analysis revealed the association of SZ risk loci with essential neuronal TFs such as FOXG1 intervening in the cortical development [343]. Similarly, Golgi phosphoprotein 3-like (GOLPH3L), gene implicated in trafficking of key neuronal signaling receptors to the cell membrane has recently been linked with SZ through the discovery of a mutation in its putative enhancer in SZ patients [344]. The possible implication of GOLPH3L in SZ would have otherwise been overlooked, illustrating once more the power of epigenomics to uncover new factors related to NDs. The integrative analysis of chromatin activity and gene expression during the normal brain development is enhancing our mechanistic understanding of numerous other NPDs including bipolar disorder [345] and autism [346], further strengthening the association between aberrant transcriptional control during brain development and NPDs.
2.3.3. Towards High Resolution Functional Genomics of NDs
Multiple cell types, including but not limited to neurons, micro and macroglia (including astrocytes and oligodendrocytes) make up the CNS. Yet, the precise number and the molecular identity of the distinct cell populations in the human brain is not clear. To understand the implication of the risk variants in NDs, it will be essential to disentangle to impact of the disease-related genes and cis-acting elements in high resolution with respect to the cell types present in the CNS. Towards addressing this need, Sullivan et al., have performed in-vitro experiments and assessed how genes linked to AD affect the production of the extracellular β-amyloid by neurons and astrocytes [347]. The authors uncovered astrocyte- and neuron-specific role of the tested genes in regulating the levels of β-amyloid highlighting unique contribution of these cell lines to AD [347]. In a genome-wide study, Nott et al., have addressed the possible mechanistic implications of distinct cell types in NDGDs. Based on surface marker expression, the authors separated brain cortical cells into the major lineages including neurons, astrocytes, oligodendrocytes and microglia. Then, using ATAC-seq to identify open chromatin regions and ChIP-seq to assess the profiles of H3K4me3 and H3K27ac, Nott et al., mapped active enhancers and promoters in these lineages. Remarkably, GWAS loci related to NPDs preferentially overlapped enhancers active specifically in neuronal cells, while GWAS hits related to AD were primarily intersecting regulatory elements active in microglia [342] further cementing the link between microglial disfunction and AD.
Brain is highly regionalized and cells from its different parts display unique transcriptional programs which likely leads to unequal susceptibility to succumb to the functional impact of the genetic alterations causing ND. Therefore, to understand the genetics of ND, we need not only cell-type, but also region-specific regulome maps. Creyghton and colleagues have provided enhancer maps of 136 regions of the adult human brain [323]. Brain regions cluster according to putative enhancer activity and the segregation corresponds grossly to anatomical structures. While most of the enhancers are shared between regions, a substantial number of interesting CREs is region-specific. Focusing on enhancers overlapping GWAS hits related to PD, the authors uncovered new α-synuclein enhancer, active in the majority of the sampled brain regions, which overlaps a genetic variant related to PD. This element also interacts with GPRIN3 a subunit of the NMDA glutamate receptor which has been considered as a potential target in PD treatment [323]. Interestingly, the activity pattern of the putative enhancer of the PARK16 gene, the most significant PD-associated polymorphism within CREs uncovered in this study, was more restricted to the cerebellum. Through chromosome conformation capture in cerebellum, the enhancer at PARK16 was further connected to other genes including RAB7L1 and MK2 which are both related to neurodegeneration [323]. Hence, detailed regulome maps of enhancer-promoter dialogue will likely help us to understand the implication of disease linked loci in NDs and the intersection of data obtained from different angles constitutes the most powerful tool to determine the genetic variants that are the most susceptible to underlie a trait including the development of NDs [348].
Anticipating the existence of a substantially larger spectrum of cellular states and identities in the CNS that are in fact known to date, can we go even further and propose an unbiased classification of risk loci to cell types? New technologies including single cell/nuclei RNA-sequencing, single cell assay for activity by transposase coupled with sequencing (ATAC-seq) [349] or spatial transcriptomics allow to assess the transcriptomic and epigenomic signatures of the individual cells [350,351], revealing the molecular identity of cell types in the CNS [352,353,354,355,356,357], and the spatial architecture of the CNS tissues [358,359]. Single cell approaches can highlight alterations of cellular function in diseased conditions [360,361,362,363,364,365] and reveal the importance of otherwise overlooked cellular fates in NDs [366] in part due to their superior power to detect differences in gene expression as compared to bulk approaches [360]. In the future, single-cell approaches allowing to map spatial conformation of the genome will contribute to further clarify the dialogue between enhancers and promoters in challenging cell populations in the brain [367,368]. Moreover, the joint information about the genotype and expression in single cell/nuclei RNA-sequencing experiments will help identify genes impacted by a disease-linked genetic variant [353,369] and hopefully will lead to a better understating of the regulatory landscape of genes that remain difficult to assess using the current technologies and require large scale sequencing approaches [329]. The far-reaching potential of such analyses to functionally annotate GWAS hits will likely allow us to paint a comprehensive picture of the genetic underpinnings of NDs. Combined with the unique capabilities of new technologies of genome and epigenome engineering, this knowledge will be instrumental to design new targets for clinical intervention strategies [320].
2.4. Targetting Global Epigenetic Signature in NDs
Drugs that target various epigenetic pathways (epidrugs) can display beneficial impact on brain functions in the context of several NDs (Table 2). The inhibitors of HDACs (HDACi), including sodium butyrate (SB, which blocks HDAC1 and 2), MS-275 (which blocks HDAC1 and 3), suberoylanilide hydroxamic acid (SAHA, Vorinostat, non-specific HDACi) and valproate acid (VA, which inhibits many pathways, including HDACs) can readily cross the blood brain barrier (BBB) and display beneficial impact on the CNS [370]. Despite a global character of their action, epidrugs can have a beneficial impact in the animal models of NDs and perhaps alleviate the syndromes in ND patients. Multiple clinical trials have been initiated to date aiming to determine the usefulness of epidrugs in ND treatments (Table 2).
Numerous examples support the favorable impact of epidrugs on ND progression. For instance, experiments performed on a transgenic mouse model of AD, showed that in comparison to the vehicle-treated animals, SB can help restore learning and memory, decrease tau protein phosphorylation, increase expression of associative learning-associated genes and restore histone acetylation in the hippocampus [371,372,373]. Likewise, SB and MS-275 can alleviate depressive behavior in AD and act to increase sucrose preference following repeated stress. The beneficial effect of HDACi is, at least in part, mediated by an enhancement of the acetylation of histone 4 at the promotor region of Transthyretin (Ttr) and a subsequent increase in Ttr leading to a disruption of beta-amyloid plaques [399,400].
In the mouse model of HD, treatment with SAHA or VA leads to an improved survival and motor performance if the treated mice compared to the control littermates [384,401]. In HD model mice (N171-82Q) phenylbutyrate treatment reduces neuronal atrophy, increases histone 3 and 4 acetylation and mice survival [374]. Likewise, sodium butyrate treatment can alleviate PD symptoms in the mouse and rat models and improve in cognitive deficits in pre-motor stage of the disease [402].
Depression has been linked with changes in DNA methylation [403]. The inhibition of DNA methylation by 5-aza-2-deoxycytidine leads to improvement of disease symptoms but the precise mode of action is unclear. Brain-derived neurotrophic factor (BDNF) is one of the essential growth factors that orchestrate the functions and viability of neurons. The antidepressant effect in global DNA-demethylation in behavioral tests might be due to an increase in the expression of BDNF in the hippocampus upon loss of methylation [396,404]. Altogether, epidrugs constitute a potentially beneficial strategy to treat NDs. However, their relatively unspecific character together with the fact that epidrugs impact the expression of only a fraction of genes, calls for more tailored approaches allowing to impact the expression of precisely chosen sets of genes in defined cell types.
3. Precision Medicine for (Epi)genome Engineering and Their Possible Application to Treat NDs
Development of new compounds that target epigenetic modifiers more specifically will lead to better defined clinical strategies. However, the genetic data indicates that NDs constitute a broad spectrum of syndromes featuring highly patient-specific genetic alterations. Therefore, it will be challenging to target such a vast repertoire of diseases with a single compound. One way out is to define the essential genes that are deregulated in a disease and act specifically at these loci to restore their expression level.
Specificity of the regulation of gene expression in the cell relies on the recognition of a unique DNA sequence by a transcription factor. Hence, one way to modify the activity of a particular locus, for instance a regulatory element inactivated by a mutation, would be to engineer a DNA-binding domain capable of recognizing this mutated sequence, which would bring along a desired enzymatic activity and restore a proper activity of the locus. Inspiration on how to develop such tools came from the bacterial world. Plant pathogenic bacteria of the Xanthomonas genus secrete transcription activator like (TAL) effectors that recognize the promoter sequences of host genes that encode factors that help the bacteria enter the cells [405]. The unique alignment between a repeated amino acid sequence in TAL and bases in the DNA sequence renders TAL amenable for encoding any sequence specificity and targeting TALE to a locus of choice [406]. Similarly, repetitive zinc finger (ZF) domains from the TFIIIA Xenopus oocyte extracts [407] can be engineered to recognize a desired sequence in the genome [408,409]. Fusion with FokI nuclease domain produces zinc finger nucleases (ZFN) [410] and TAL-Like Effectors (TALE) [411] and allows to perform highly specific genome modification including gene knockouts in various contexts [412,413,414]. Alternatively, fusion between ZFs and activation domains such as VP64 yields a tool for a specific up-regulation of desired loci [415]. However, despite advances in cloning strategies [411,416], TALEs remain experimentally cumbersome, requiring extensive and often challenging cloning.
More recently the discovery of the prokaryotic acquired immune-like system relying on the Clustered Regularly Interspaced Short Palindromic Repeats (CRISPR) and CRISPR-associated protein (Cas) referred to as CRISPR-Cas machinery allowed the development of a more straightforward and versatile genome editing system (Figure 2).
3.1. CRISPR-Cas-Based Genome Engineering Principles and Considerations
Prokaryotes safeguard pieces of the genomic DNA (spacers) of their invaders within the CRISPR loci [417,418,419,420]. Transcription of CRISPR genes produces non-coding RNA molecules (crRNA) which are complementary to the genetic material of the intruder (protospacers) [421]. The crRNA, together with the transactivating CRISPR RNA (tracrRNA) [422], allows to specifically target the Cas endonuclease to the invader’s genome [423]. There are several families of Cas proteins, here we will focus on factors from the Class 2 CRISPR–Cas systems such as the Cas9 from Staphylococcus aureus or Cas12a (Cpf1) [424] from, for example, Lachnospiraceae bacterium which are currently most widely used tor (epi)genome modifications. Due to its small size (<1000amino acids), CasX [425] constitutes a great alternative to Cas9 and Cas12a and will probably be extensively developed in the future.
As we saw above, Cas proteins are tethered to the foreign genome by direct Watson-Crick base pairing between the crRNA and protospacers. The crRNA is 20 or 24 base pair long in Cas9 and Cas12a systems, respectively [422]. Such complementarity is, however, not sufficient to induce nucleolytic reaction and DNA can be cleaved only at sites where the protospacer sequence is flanked downstream by a specific protospacer adjacent motif (PAM) [426,427]. PAMs for distinct Cas proteins differ in composition Cas9 and Cas12a require a G- and T-rich PAMs respectively [428]. PAM is not present at the CRISPR genes, which avoids their destruction and autoimmunity.
Cas9 protein is composed of six domains including REC I, which binds the RNA sequence, REC II, Bridge Helix, PAM Interacting, and HNH and RuvC domains which cleave the single-stranded DNA in a specific manner: HNH attacks the DNA complementary to the crRNA while RuvC cuts the non-target strand [426,429,430], Cas9 produces blunt end double stranded DNA brakes (DSB) [428]. Cas12a lacks an HNH domain and contains the RuvC and a Nuc endonuclease domains, which introduce staggered DNA breaks into the two strands of the target sequence leaving cohesive ends with 4-5 nt. 5′ overhangs [428]. Single molecule approaches, using naked DNA as template, revealed that the Cas9 protein constantly scans the genome; yet only upon the recognition of the PAM sequence does the Cas9 initiate the unwinding of DNA and the hybridization between DNA and the spacer sequence containing RNA [427].
The adoption of the CRISPR-Cas system to mammalian cells rendered it possible to engineer the genome sequence by removing or inserting desired fragments of DNA in an efficient manner [431,432,433,434,435,436,437,438,439,440,441,442]. In the Cas9-based system, the procedure relies on a sequence-specific, 20bp-long guide RNA (gRNA) which is covalently modified to include tracrRNA moiety. This construct is sufficient to specifically target the Cas9 endonuclease to the genome and induce double strand breaks [433,434,443]. Cas12a requires solely the gRNA sequence to recognize and cleave DNA [428,440,442]. The number of sites that are targetable by CRISPR-Cas9 system is intimately linked with the the frequency of the PAM and in the human genome the 5′NGG3′ site required for the Cas9 occurs every 8-12 bp [426,427,444].
The genetic damage caused by CRISPR-Cas activates DNA repair mechanism(s) including the non-homologous end joining (NHEJ), microhomology-end joining (MHEJ), as well as homology-directed repair pathway (HDR) that requires an external template to mend the damage [434,445]. The error prone NHEJ pathway may lead to mutational insertions and deletions (indels) of short sequences at the targeted site. Indels can result in gene disruption by for instance frameshift mutation leading to a premature termination of translation and functional disruption of the gene. This property is widely used to silence loci, including in high-throughput genetic screens [446,447,448,449]. However, caution must be taken when designing and interpreting the results as NHEJ can lead to an emergence of gene isoforms with unclear functions [450].
Until recently, NHEJ was thought to prevail the homology-assisted mechanisms in resolving DSBs induced by programmable nucleases [451]. However, several reports indicate that MHEJ could in fact constitute the major pathway repairing DSB induced by Cas [452,453]. Moreover, MHEJ would allow us to achieve a remarkable precision in DNA repair [452]. For instance, at loci where frameshift mutations arise as a consequence of microduplications, MHEJ can lead to an efficient removal of the alteration and restoration of gene expression [454].
One can tilt the balance of how Cas9-induced DSB are resolved towards homology-based mechanisms including MHEJ. This can be attained by choosing the most optimal cut site at the locus of interest that would lead to MHEJ [452,454]. The most straightforward approach to control the genetic engineering process, however, implicates HDR pathway and is applicable to cells that cycle actively. HDR relies on the external provision of DNA donor sequence with homology to the target site. The HDR pathway mediates the insertion of the desired sequence, provided it is flanked by sequences homologous to the DNA surrounding the cut site. Thus, HDR mediates precise and controllable modification of the genome including a replacement of the mutated nucleotides, addition of an entire domain to an otherwise mutated locus or insertion of a new regulatory sequence in a vicinity of a silenced gene. It allows the inclusion of a transgene into either the site of interest or into the so-called safe-harbor locus which, by mitigating epigenetic silencing, robustly drives gene expression. The induction of the HDR machinery at the CRISPR-Cas-cut sites appears as the most optimal approach: it allows to precisely control the genetic modification and insert a desired sequence at the chosen location. Yet, it requires the cells to pass though the G2/S phase [455] which limits its applicability to NDs. (There are approaches that help increase the efficiency of HDR the reader may consider a recent excellent review on this topic by Yeh and colleagues [445].)
The intricacy of the genome engineering systems and reduced applicability in post-mitotic cells advocates for, where possible, somewhat simpler systems to orchestrate gene expression without the need to modify the DNA sequence. These include programmable nuclease—based approaches that allow to tether a desired enzymatic activity to a precisely defined site in the genome (Figure 2).
3.2. From Genome to (Epi)genome Edition
The binding of the Cas9 protein to the genome is stabilized at sites complementary to the gRNA. Like we saw in the case of TALENs and ZFNs, where TALs or ZFs brought the covalently linked FokI nuclease to a specific site in the genome, the DNA specificity of the CRISPR-Cas system can be harnessed to tether a desired activity provided its own endonuclease function is blocked. In Streptococcus pyogenes Cas9, this can be achieved by a simultaneous replacement of the aspartic acid at position 10 (RuvC domain) and histidine at position 840 (HNH domain) by alanine, which yields the so called catalytically defective Cas9 (dCas9Sp) [426,456]. In Staphylococcus aureus the inactivation of the endonuclease activity of Cas9 (dCas9Sa) requires the mutations of both D10 and aspargine at position 580 (HNH domain). Depending on the fused activity, dCas9 becomes a site-specific modifier of the genome from a single base pair editor to chromatin modification inducer or 3D chromatin structure moderator. As we will see below, it might also be favorable to retain part of Cas9 activity, allowing us to induce a single stranded DNA break (SSB, “nick”) in the instead of DSB; it can be achieved by mutating D10 to A leaving the H840 position unchanged.
3.2.1. dCas9-Orchestrated Base Edition in the Genome
Point mutations are a common cause of genetic disorders and the best approach to correct these alterations would, in principle, be to change the single mutated base pair without the need of larger scale replacement of the DNA sequence as in the HDR. Such modification can be achieved by fusing dCas9 or Cas9 with nick activity (nCas9) with cytosine or adenine deaminases. Cytosine deamination converts it to uracil which can be then converted to thymine when mismatch repair systems (MMR) are favored. The latter can be achieved using nCas9 [457,458,459,460]. Adenine base editors inherit from tRNA adenine deaminase (TadA) of E. coli which switches adenine to inosine, which is next converted to guanine in the cell [457]. Base edition constitutes a promising tool for precise base exchange at will, especially since it can successfully be applied to postmitotic cells in vivo [461].
3.2.2. Site-Specific Epigenome Rewiring to Enhance Gene Expression
Highly locus-specific up-regulation of gene expression has been developed over the past three decades. Initially, the approach was to take advantage of potent transcriptional activator domains (ActDs), such as virion protein 16 (VP16; from the herpes simplex virus) or the p65 (component of the NFκB pathway) and target them to the chosen sites in the genome with a factor that is able to specifically recognize DNA sequence such as yeast GAL4 [462] or later ZFs [463,464,465]. Binding of such a construct to its cognate location results in tethering of the transcriptional machinery and gene onset. More recently, ActDs were fused to TALE effectors and dCas9 rendering this tool much easier to use [411,412,466,467]. Multiple recent improvements of the locus-specific activation render this approach more robust, versatile, technically simple, and controllable.
Loci may appear difficult to modulate using ActD and several strategies allow to alleviate this problem. For instance, the inclusion of ActD arrays as in the VP48, VP64, VP160 or VP192 constructs, built by multiplying the VP16 domain can help increasing the transactivation effect [411,466,468]. In SunTag technology multiple peptides, each of which is recognized by ActD, are fused to dCas9 and altogether enhance gene expression at the locus complementary to the gRNA [469]. Another strategy consists in combining several unique and potent ActDs, as for example in the tripartite activator: VP64-p65-Rta (VPR) [470]. Likewise, an increase in the number of the tethered TALE [467] or dCas9- activators per locus, by for instance inclusion of multiple distinct TALEs or sgRNAs, respectively, and can be used to boost the transcription of the target gene [466].
To further increase the versatility of the approach, ActDs may also be replaced with domains displaying enzymatic activities allowing to directly remodel the local chromatin environment at a regulatory stie. Fusion of dCas9 with histone-modifying domains, for instance the HAT domain of p300 [471,472,473] or CBP [474], yields a potent gene expression activator that functions when tethered to both promoters and enhancers. This expands the possibility to address the deficiencies in gene activation due to mutations in regulatory elements including promoters and enhancers [471]. This remarkable tool can be also used to screen for functional regulatory elements in a cell type of choice ([475] for review see [476]).
Local (epi)genome rewiring may also be achieved by appending histone methyltransferase domains to dCas9. Bringing SET domain of the MLL3/4, which establishes H3K4me1 at enhancers, allows to attract cohesin complex thereby inducing the remodeling of the 3D chromatin arrangement at the locus correlating with the onset of the target gene expression [477]. Furthermore, the fusion of dCas9 with PR/SET Domain 9 that deposits H3K4me3 mark correlating with promoter activity, allows to establish and maintain gene expression [478].
Aberrant DNA methylation can lead to erroneous gene silencing with profound molecular consequences. Fusion between site-specific ZFs or dCas9 with either Ten-Eleven Translocation methylcytosine dioxygenase (Tet) or Thymine DNA glycosylase (TDG), enzymes that have been linked to demethylation of methylcytosine, can lead to reactivation of silenced genes [479,480] and disease rescue [481,482,483].
Transcriptional regulation relies on a concert action of TFs, enzymes and proteins that recognize modified chromatin and signal the appropriate conditions the transcriptional apparatus. As we saw above, nonselective HDAC inhibition my exert neuroprotective effects which can be accompanied by memory and learning enhancement. This effect is, in part, mediated by the onset of the expression of BDNF due to the increased histone acetylation arising as a consequence of HDAC inhibition [484,485]. Tethering BRD4, a BET bromodomain containing reader of acetylated histones, to the BDNF promoter further enhances the effect of HDAC inhibition and promotes learning [486]. Thus, the combination of epi-drug treatment with tailored dCas9-mediated epi-targeting might potentially constitute a beneficial clinical avenue (Figure 3).
3.2.3. Approaches for Locus-Specific Gene Silencing
There are multiple domains that can exert an inhibitory impact on chromatin and thereby gene expression. These include the Krüppel-associated box (KRAB) [487], ERF repressor domain (ERD) [488], mSIN3 interaction domain (SID) [489], friend of GATA1 (FOG1) [490], DNM3A [490], EZH2 and G9A methyltransferases [490,491] along with MeCP2 [492] and HDAC [493]. Echoing designer activators, multiplying repressor domains (RDs) [494] or combining several distinct RDs may help intensify the inhibitory effect on transcription of the target gene [492,495,496]. Mechanistically, repression depends on KAP1/TIF1beta/HP1a pathway in the case of KRAB repressors [497] and additional not yet fully understood mechanisms implicating H3K9me3 and H3K27me3 and DNA methylation [490,493].
Like in the case of site-specific ActDs, the first tailored repressors were recruited to the locus of interest by GCN4 or ZFs [465,487,498]. Later, TALE- [495,499,500] and CRISPR-dCas9-based approaches emerged [490,491,501,502,503]. Due to experimental ease, dCas9 based repressors are currently most widely used in the field. One important difference between ActD and RepD is the stability of the effect. In part, this difference might arise as a consequence of the intrinsic mechanisms that retain DNA methylation during the S-phase of the cell cycle, RD may induce long term effect on gene expression [493] even in cycling cells [498].
3.2.4. Complex Designs to Simultaneously Activate and Repress Distinct Sets of Genes
Transcriptional deregulation in disease is often a complex phenomenon featuring aberrant up- and down-regulation of genes. Thus, simultaneous activation of a subset of loci and inhibition of other targets appears as an attractive and desirable clinical design. It is possible to attain it in a single cell by combining distinct Cas orthologs (dCas9Sp and dCas9Sa for instance) fused to ActD or RepDs [504]. Another approach consists in combining CRISPR-dCas9 system with viral aptamers which recognize specific RNA sequences encoded within the gRNA construct (PP7, MS2, Com or PBS). The aptamers are fused with distinct effector domains: ActDs or RepDs [505,506].
3.2.5. Engineering Genome Topology to Orchestrate Gene Expression
One way to orchestrate gene expression is to create a chromatin loop that connects promoter to an enhancer. Artificial loop can be built for instance, via an incorporation of transcription-factor binding sites (TFBS) close to the two sequences that are desired to be in proximity. In Nolis et al. the insertion of TFBS for λ repressor, a prokaryotic TF, close to enhancer and promoter of INF1 gene, activates its transcription upon expression of the λ repressor [507]. Similar effect was achieved by engineering fusion protein between ZFs recognizing β-globin enhancer and the LIM Domain Binding 1 (LDB1), a factor that interacts with a broad array of TFs that contain LIM domain and mediates interactions between them. Expression of this fusion protein leads to a formation of a loop bridging the β-globin enhancer and promoter leading to a transcriptional up-regulation of β-globin [508].
An artificial loop can be also built using dCas9 heteromonomers or dCas9 monomers fused with a dimerization domain [509]. In this system, heterodimerization between the two fused dCas9 proteins is obtained by adding a chemical compound such as abscisic acid which stabilises an interaction between them. Alternatively, dimerization of dCas9-CIBN and dCas9- CRY2 proteins can be induced by blue light [510].
3.3. From Mutation to Precise Target CRISPR-Cas-Mediated In Vivo (Epi)genome Editing to Target NDs
Adeno-associated virus (AAV) delivery-based systems deliver Cas9 into post-mitotic neurons in the brain and has been used to for example study the functional consequences of the removal of DNA-methylation related machinery [511]. This result paves the way to the future possibility to engineer genomes in ND patients. (Epi)genome engineering of essential loci critically related to NDs have been initiated and show a great potential as new therapeutic agents (Table 3).
For instance, in the Fragile X-syndrome (FXS), FMR1 gene is silenced due to a local expansion of a repeated sequence leading to DNA methylation. Bringing Tet1 to the FMR1 promoter, in post-mitotic neurons derived from FXS patient’s induced pluripotent stem cells, can rescue neuronal functions [527]. Several other essential examples highlight a tremendous potential of (epi)genome rescue tools to address NDs (Table 3). Undoubtably, the strongest arguments in favor of Cas9-mediated treatment approaches are the robustness, specificity, versality and relatively high efficiency of the system Arguments against is the difficulty is the potential for off-target effects and the danger of some yet unknown irreversible consequences of the therapy. The latter being particularly difficult to address, unless sequencing of the whole patient’s genome becomes a routine.
The delivery of the machinery to the CNS might reveal to be challenging. Lentiviruses (LVs), adenoviruses, adeno-associated viruses (AAVs), as well as non-viral vehicles including extracellular vesicles (EVs), nanoparticles or hybrid formulations have successfully been used as carriers of (epi)genome editing tools in rodents [483,528,529,530]. Viral-based strategies rely on vectors lacking genes essential to viral propagation including rep and cap. Due to that, viral genome remains episomal for a long period of time supporting a continuous gene expression especially in non-dividing cells [530]. LVs are particularly suitable for CRISPR/Cas9 delivery [531], owing to their capacity to carry large DNA inserts including (d)Cas9Sp. Moreover, LVs can transduce a broad range of both dividing and nondividing cells, with minimal cytotoxic and immunogenic response [483]. However, LV are limited in the range of potential target cells (compared to AAVs), tissue penetration and efficiency at doses relevant in vivo [532]. When targeting cells in the CNS, these issues are more challenging.
A variety of AAVs have also been considered as means of Cas9 delivery to the CNS [533,534,535]. More specifically, AAV2 and AAV9 serotypes appear as the most suitable tools in the context of NDs. As compared to LVs, they possess high brain tissue tropism, desirable infection rate, and lower toxicity, immuno-stimulation, and decreased risk of causing disease in human. Yet, it is unclear how to set the precise level of expression of a transgene using AAVs [536]. In this respect, a transient activation in dividing cells might reveal to be the right approach.
Despite positive results obtained using AAVs in rodents, there is still a need to enhance the efficiency and robustness of the delivery system. Combination of viral and non-viral carriers could potentially constitute an interesting alternative to the non-hybrid strategies [537,538]. Likewise, more recently, EVs have gained attention in the field as EVs readily pass the BBB and can deliver genome editing tools like Cas9 RNPs and dCas9 RNPs to brain tissue [539] albeit with still relatively low efficiency. The future tools will need to feature larger stability, scalability, BBB and brain tissue permeability, together with the capacity to target chosen cell types in the bran while maintaining safety and optimal (epi)genome-editing efficiency.
Essential Considerations When Designing Epigenetic Engineering Strategies
We have entered an unprecedented era in precision biology, the strategies allowing site-specific modulation of the genome are changing both the fundamental and applied medicine. Yet, several essential points need to be taken into consideration when using ZF/TALE and (d)Cas tools. The efficiency of the reaction is not uniform across the genome and the engineering effect is often markedly affected by the position to which one tethers the effector; this effect impacts both TALE-activators [466,467] as well as dCas9-based factors [466,475]. As we saw above, activators able to recognize stem loops in the gRNA constructs may help diversify the list of genes that could be regulated in a particular cell type simultaneously [475,540] and with lower toxicity [541]. Such systems should also enhance the effector domain activity at challenging sites thereby allowing to bypass the local effects that block ActD or RepD.
Sustained presence of high concentrations of effector domains may distort cell’s physiology, ActD for instance may hijack transcriptional co-activators thereby impacting on other loci expressed in the cell. To address this limitation, inducible dCas9 systems are being developed for more precise temporal control of gene activation [483] which can be attained with both chemical compounds and light [542]. The core concept of these strategies lies in the capacity to control the heterodimerization of dCas9 and an activator or a repressor protein externally, using tailored extrinsic inducers of interactions between pairs of domains [483,542,543,544,545,546,547]. Alternatively, one can consider quick removal of Cas9 right after induction, KamiCas9 is an interesting approach in which cells are transfected by sgRNA that targets the sequence of interest in addition to a vector encoding sgRNA targeting Cas9 itself. The Cas9 specific sgRNA is expressed under a weak promoter thereby favoring the edition of the target sequence while sustaining the self-inactivating potential [520]. Research in this area is developing extremely rapidly, therefore, we anticipate the development of improved strategies that will be amenable for clinical practice in the nearest future.
Despite the, in principle, high specificity of ZFs/TALE and (d)Cas, these factors might nonetheless display non-negligeable off-target activity and a potential to introduce modifications at unwanted sites. The off-targets can be predicted using computational tools and directly identified using whole-genome sequencing (WGS) and ChIP-seq experiments allowing to map the regions bound by the dCas9 [548]. Remarkably, WGS-based identification of Cas9 off-target sites revealed an overall low mutation burden (2-2,5% of WGS reads display mutation) [549] and the Cas9 induced off-target indels are produced at a relatively low frequency (below 1-0.1%), primarily within sequences with high homology to the target [436,518,549,550,551]. The design of gRNA can be further optimized using on-line tools which allow to enhance the control over the (epi)genome editing reactions [552,553,554,555].
Altogether, it is essential to experimentally determine which site at a locus of interest is most optimal to attain a desired effect and which approach is best suited to enhance gene expression while keeping cytotoxicity at a minimal level. Likewise, a routine of high throughput sequencing to assess the level of off target edition appears as essential when developing new tools for (epi)genome engineering.
4. Discussion
The remarkable complexity of cell types and cellular interactions in the brain makes it challenging to propose new drugs that could target NDs. Furthermore, very limited regenerative potential of the brain tissue constitutes an additional difficulty in the treatment of NDs featuring neuronal loss such as AD or PD. Finally, the enhanced damming capacity of the BBB needs to be accounted for when designing treatment schemes. Thus, there is a need to develop tailored and specific agents to address NDs at the earliest possible stage.
Epigenetic drugs that affect the global level of histone modifications in the cell have been used to treat NDs (Table 2, Figure 3). However, this the treatment is unspecific and general and relies on differences in the responses of genes to the global change in epigenetic landscape of the cell. Another approach would be to restore the correct transcriptional pattern of genes mis-regulated in the ND (Figure 4).
Classical genetic therapies for cancer and other diseases are gaining momentum. CD19-directed chimeric antigen receptor (CAR) T cells are engineered to express a T-cell receptor specifically recognizing CD19 on relapsed or refractory large B cell lymphoma (YESCARTA®, NOVARTIS). Other formulations include Zolgensma® which brings a functional copy of the SMN1 gene to treat patients with spinal muscular atrophy. NDs have not yet been targeted with genetic therapies.
(Epi)genome engineering to restore the correct gene activity or remove a mutated copy of a gene is likely the most attractive future direction in clinical approaches. Can such strategy be envisioned in vivo? CRISPR-Cas9- and TALEN-mediated genome engineering has experimentally been proven feasible in murine model (Table 3) [556] and in the non-human primates [557,558]. The AAV-mediated delivery of the Cas9Sa to mammalian brain appears optimal enough to modulate gene expression and introduce DNA sequence changes. Consequently, there is a number of clinical trials worldwide to address the usefulness of CRISPR-Cas9-mediated genome modification [559]. A study published in June 2021, introduced a lipid nanoparticle-delivered CRISPR-Cas9 genome editing tool to treat individuals affected by transthyretin amyloidosis (ATTR amyloidosis), a life-threatening disease caused by progressive accumulation of misfolded transthyretin (TTR) [560]. This breakthrough finding will likely change the clinical management of monogenic diseases in the future, opening the possibility to edit genomes of patients attained by otherwise untreatable conditions.
Multitude of TALE/ZF and Cas9-based tools enable the editing of either the DNA sequence or directly gene activity. How can we approach (epi)genome editing? To this end, we need to cartography the regulomes of brain cells and understand how they are distorted in NDs. First step towards this goal is to find and annotate differentially expressed genes and identify their enhancers to promoters in distinct cell types in the CNS. One essential take-home message from the data generated so far is that a single enhancer can regulate multiple genes at a time and that this intricate network of regulatory transactions can be targeted by disease linked GWAS hit [323,342]. Thus, a strategy to restore the correct activity of the GWAS variants might potentially be more fruitful and less toxic to the patient than targeting the signaling pathway affected by the transcriptional mis-regulation of the cis-linked genes (Figure 2).
5. Conclusions
The understanding of the functional consequences of the mis-regulation of chromatin modifiers will help to determine the most suitable treatment strategies. One thing of course, is to ascertain whether NDs feature a general loss or gain of epigenetic modifications in the genome. In such cases, it seems reasonable to assume that the restoration of the correct global level of chromatin modification represents a potentially advantageous strategy to treat NDs. Yet, one important question is whether such a systemic action is the most suitable strategy or whether a tailored and highly precise approach allowing to restore the correct expression of the chromatin modifier or its essential targets in a particular cellular context will be preferential for the treatment of NDs. The answer will most likely be disease- and patient-dependent.
Funding
Work in AP lab is founded by the Dioscuri Grant, (Dioscuri is a program initiated by the Max Planck Society, managed jointly with the National Science Centre (NCN, Poland) and mutually funded by the Polish Ministry of Education and Science and the German Federal Ministry of Education and Research), by the OPUS17 grant (UMO-2019/33/B/NZ2/02437) from the NCN and by the EMBO Installation Grant.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Not applicable.
Acknowledgments
We thank the members of the Dioscuri Centre for Chromatin Biology and Epigenomics for helpful discussions.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Waddington, C.H. The Epigenotype. Int. J. Epidemiol. 2012, 41, 10–13. [Google Scholar] [CrossRef] [Green Version]
- Felsenfeld, G. A Brief History of Epigenetics. Cold Spring Harb. Perspect. Biol. 2014, 6, a018200. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Allis, D.C.; Caparros, M.-L.; Jenuwein, T.; Reinberg, D.; Lachlan, M. (Eds.) Epigenetics, 2nd ed.; Cold Spring Harbor Laboratory: Cold Spring Harbor, NY, USA, 2015; ISBN 978-1-936113-59-0. [Google Scholar]
- Roeder, R.G. 50+ years of eukaryotic transcription: An expanding universe of factors and mechanisms. Nat. Struct. Mol. Biol. 2019, 26, 783–791. [Google Scholar] [CrossRef] [PubMed]
- Bird, A. Perceptions of epigenetics. Nature 2007, 447, 396–398. [Google Scholar] [CrossRef] [PubMed]
- Kornberg, R.D. Chromatin Structure: A Repeating Unit of Histones and DNA. Science 1974, 184, 868–871. [Google Scholar] [CrossRef]
- Olins, A.L.; Olins, D.E. Spheroid Chromatin Units (ngr Bodies). Science 1974, 183, 330–332. [Google Scholar] [CrossRef]
- Woodcock, C.; Safer, J.; Stanchfield, J. Structural repeating units in chromatin: I. Evidence for their general occurrence. Exp. Cell Res. 1976, 97, 101–110. [Google Scholar] [CrossRef]
- Ou, H.D.; Phan, S.; Deerinck, T.J.; Thor, A.; Ellisman, M.H.; O’Shea, C.C. ChromEMT: Visualizing 3D chromatin structure and compaction in interphase and mitotic cells. Science 2017, 357, eaag0025. [Google Scholar] [CrossRef] [Green Version]
- Fussner, E.; Strauss, M.; Djuric, U.; Li, R.; Ahmed, K.; Hart, M.; Ellis, J.; Bazett-Jones, D.P. Open and closed domains in the mouse genome are configured as 10-nm chromatin fibres. EMBO Rep. 2012, 13, 992–996. [Google Scholar] [CrossRef] [Green Version]
- Luger, K.; Mader, A.W.; Richmond, R.K.; Sargent, D.F.; Richmond, T.J. Crystal Structure of the Nucleosome Resolution Core Particle at 2.8 A Resolution. Nature 1997, 389, 251–259. [Google Scholar] [CrossRef]
- Ricci, M.A.; Manzo, C.; Garcia-Parajo, M.F.; Lakadamyali, M.; Cosma, M.P. Chromatin Fibers Are Formed by Heterogeneous Groups of Nucleosomes In vivo. Cell 2015, 160, 1145–1158. [Google Scholar] [CrossRef] [Green Version]
- Knezetic, J.A.; Luse, D.S. The presence of nucleosomes on a DNA template prevents initiation by RNA polymerase II in vitro. Cell 1986, 45, 95–104. [Google Scholar] [CrossRef]
- Lorch, Y.; Lapointe, J.W.; Kornberg, R.D. Nucleosomes inhibit the initiation of transcription but allow chain elongation with the displacement of histones. Cell 1987, 49, 203–210. [Google Scholar] [CrossRef]
- Han, M.; Grunstein, M. Nucleosome loss activates yeast downstream promoters in vivo. Cell 1988, 55, 1137–1145. [Google Scholar] [CrossRef]
- Kieffer-Kwon, K.-R.; Nimura, K.; Rao, S.S.; Xu, J.; Jung, S.; Pekowska, A.; Dose, M.; Stevens, E.; Mathe, E.; Dong, P.; et al. Myc Regulates Chromatin Decompaction and Nuclear Architecture during B Cell Activation. Mol. Cell 2017, 67, 566–578.e10. [Google Scholar] [CrossRef] [PubMed]
- Lawrence, M.; Daujat, S.; Schneider, R. Lateral Thinking: How Histone Modifications Regulate Gene Expression. Trends Genet. 2016, 32, 42–56. [Google Scholar] [CrossRef] [Green Version]
- Chen, K.; Rajewsky, N. The evolution of gene regulation by transcription factors and microRNAs. Nat. Rev. Genet. 2007, 8, 93–103. [Google Scholar] [CrossRef] [PubMed]
- Zaret, K.S. Pioneer Transcription Factors Initiating Gene Network Changes. Annu. Rev. Genet. 2020, 54, 367–385. [Google Scholar] [CrossRef] [PubMed]
- Cramer, P. Organization and regulation of gene transcription. Nat. Cell Biol. 2019, 573, 45–54. [Google Scholar] [CrossRef] [PubMed]
- Hannah, A. Localization and Function of Heterochromatin in Drosophila Melanogaster. Adv. Genet. 1951, 4, 87–125. [Google Scholar] [CrossRef] [PubMed]
- Muller, H.J. Types of visible variations induced by X-rays inDrosophila. J. Genet. 1930, 22, 299–334. [Google Scholar] [CrossRef]
- Dorer, D.R. Expansions of transgene repeats cause heterochromatin formation and gene silencing in Drosophila. Cell 1994, 77, 993–1002. [Google Scholar] [CrossRef]
- Schultz, J. Variegation in Drosophila and the Intert Chromosome Regions. Genetics 1936, 22, 27–33. [Google Scholar]
- Klosin, A.; Casas, E.; Hidalgo-Carcedo, C.; Vavouri, T.; Lehner, B. Transgenerational transmission of environmental information inC. elegans. Science 2017, 356, 320–323. [Google Scholar] [CrossRef] [Green Version]
- Spitz, F.; Furlong, E. Transcription factors: From enhancer binding to developmental control. Nat. Rev. Genet. 2012, 13, 613–626. [Google Scholar] [CrossRef] [PubMed]
- Mifsud, B.; Tavares-Cadete, F.; Young, A.N.; Sugar, R.; Schoenfelder, S.; Ferreira, L.; Wingett, S.; Andrews, S.; Grey, W.; Ewels, P.A.; et al. Mapping long-range promoter contacts in human cells with high-resolution capture Hi-C. Nat. Genet. 2015, 47, 598–606. [Google Scholar] [CrossRef]
- Meissner, A.; Mikkelsen, T.S.; Gu, H.; Wernig, M.; Hanna, J.H.; Sivachenko, A.; Zhang, X.; Bernstein, B.E.; Nusbaum, C.; Jaffe, D.B.; et al. Genome-scale DNA methylation maps of pluripotent and differentiated cells. Nature 2008, 454, 766–770. [Google Scholar] [CrossRef] [Green Version]
- Weber, M.; Davies, J.J.; Wittig, D.; Oakeley, E.J.; Haase, M.; Lam, W.L.; Schübeler, D. Chromosome-wide and promoter-specific analyses identify sites of differential DNA methylation in normal and transformed human cells. Nat. Genet. 2005, 37, 853–862. [Google Scholar] [CrossRef]
- Mikkelsen, T.S.; Ku, M.; Jaffe, D.B.; Issac, B.; Lieberman, E.; Giannoukos, G.; Alvarez, P.; Brockman, W.; Kim, T.-K.; Koche, R.P.; et al. Genome-wide maps of chromatin state in pluripotent and lineage-committed cells. Nature 2007, 448, 553–560. [Google Scholar] [CrossRef] [PubMed]
- Barski, A.; Cuddapah, S.; Cui, K.; Roh, T.-Y.; Schones, D.E.; Wang, Z.; Wei, G.; Chepelev, I.; Zhao, K. High-Resolution Profiling of Histone Methylations in the Human Genome. Cell 2007, 129, 823–837. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tie, F.; Banerjee, R.; Stratton, C.A.; Prasad-Sinha, J.; Stepanik, V.; Zlobin, A.; Diaz, M.O.; Scacheri, P.C.; Harte, P.J. CBP-mediated acetylation of histone H3 lysine 27 antagonizes Drosophila Polycomb silencing. Development 2009, 136, 3131–3141. [Google Scholar] [CrossRef] [Green Version]
- Wang, Z.; Zang, C.; Cui, K.; Schones, D.E.; Barski, A.; Peng, W.; Zhao, K. Genome-wide Mapping of HATs and HDACs Reveals Distinct Functions in Active and Inactive Genes. Cell 2009, 138, 1019–1031. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kieffer-Kwon, K.-R.; Tang, Z.; Mathe, E.; Qian, J.; Sung, M.-H.; Li, G.; Resch, W.; Baek, S.; Pruett, N.; Grøntved, L.; et al. Interactome Maps of Mouse Gene Regulatory Domains Reveal Basic Principles of Transcriptional Regulation. Cell 2013, 155, 1507–1520. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Varshavsky, A.J.; Sundin, O.; Bohn, M. A stretch of “late” SV40 viral DNA about 400 bp long which includes the origin of replication is specifically exposed in SV40 minichromosomes. Cell 1979, 16, 453–466. [Google Scholar] [CrossRef]
- Wu, C. The 5′ ends of Drosophila heat shock genes in chromatin are hypersensitive to DNase I. Nat. Cell Biol. 1980, 286, 854–860. [Google Scholar] [CrossRef] [PubMed]
- Almer, A.; Hörz, W. Nuclease Hypersensitive Regions with Adjacent Positioned Nucleosomes Mark the Gene Boundaries of the PHO5/PHO3 Locus in Yeast. EMBO J. 1986, 5, 2681–2687. [Google Scholar] [CrossRef]
- Heintzman, N.D.; Hon, G.C.; Hawkins, R.D.; Kheradpour, P.; Stark, A.; Harp, L.F.; Ye, Z.; Lee, L.K.; Stuart, R.; Ching, C.W.; et al. Histone modifications at human enhancers reflect global cell-type-specific gene expression. Nature 2009, 459, 108–112. [Google Scholar] [CrossRef] [Green Version]
- Schones, D.E.; Cui, K.; Cuddapah, S.; Roh, T.-Y.; Barski, A.; Wang, Z.; Wei, G.; Zhao, K. Dynamic Regulation of Nucleosome Positioning in the Human Genome. Cell 2008, 132, 887–898. [Google Scholar] [CrossRef] [Green Version]
- Rada-Iglesias, A.; Bajpai, R.; Swigut, T.; Brugmann, S.A.; Flynn, R.A.; Wysocka, J. A unique chromatin signature uncovers early developmental enhancers in humans. Nature 2010, 470, 279–283. [Google Scholar] [CrossRef] [Green Version]
- Creyghton, M.P.; Cheng, A.W.; Welstead, G.G.; Kooistra, T.; Carey, B.W.; Steine, E.J.; Hanna, J.; Lodato, M.A.; Frampton, G.M.; Sharp, P.A.; et al. Histone H3K27ac separates active from poised enhancers and predicts developmental state. Proc. Natl. Acad. Sci. USA 2010, 107, 21931–21936. [Google Scholar] [CrossRef] [Green Version]
- Pekowska, A.; Benoukraf, T.; Cabeza, J.Z.; Belhocine, M.; Koch, F.; Holota, H.; Imbert, J.; Andrau, J.-C.; Ferrier, P.; Spicuglia, S. H3K4 tri-methylation provides an epigenetic signature of active enhancers. EMBO J. 2011, 30, 4198–4210. [Google Scholar] [CrossRef] [PubMed]
- Bell, A.C.; West, A.G.; Felsenfeld, G. The Protein CTCF Is Required for the Enhancer Blocking Activity of Vertebrate Insulators. Cell 1999, 98, 387–396. [Google Scholar] [CrossRef] [Green Version]
- Doskočil, J.; Šorm, F. Distribution of 5-methylcytosine in pyrimidine sequences of deoxyribonucleic acids. Biochim. Biophys. Acta (BBA) Bioenerg. 1962, 55, 953–959. [Google Scholar] [CrossRef]
- Wu, T.P.; Wang, T.; Seetin, M.G.; Lai, Y.; Zhu, S.; Lin, K.; Liu, Y.; Byrum, S.D.; Mackintosh, S.G.; Zhong, M.; et al. DNA methylation on N6-adenine in mammalian embryonic stem cells. Nature 2016, 532, 329–333. [Google Scholar] [CrossRef] [Green Version]
- Bird, A.; Taggart, M.; Frommer, M.; Miller, O.J.; Macleod, D. A fraction of the mouse genome that is derived from islands of nonmethylated, CpG-rich DNA. Cell 1985, 40, 91–99. [Google Scholar] [CrossRef]
- Gardiner-Garden, M.F.M. CpG Islands in Vertebrate Genomes. J. Mol. Biol. 1987, 196, 261–282. [Google Scholar] [CrossRef]
- Holliday, R.; Pugh, J.E. DNA modification mechanisms and gene activity during development. Science 1975, 187, 226–232. [Google Scholar] [CrossRef]
- Iglesias, A.D. Epigenetic defects, assisted reproductive technology, and clinical practice: A call for clinicians and genetic counselors. Clin. Genet. 2004, 66, 481–482. [Google Scholar] [CrossRef]
- Riggs, A. X inactivation, differentiation, and DNA methylation. Cytogenet. Genome Res. 1975, 14, 9–25. [Google Scholar] [CrossRef]
- Cattanach, B.M.; Kirk, M. Differential activity of maternally and paternally derived chromosome regions in mice. Nat. Cell Biol. 1985, 315, 496–498. [Google Scholar] [CrossRef] [PubMed]
- Compere, S.J.; Palmiter, R.D. DNA methylation controls the inducibility of the mouse metallothionein-I gene in lymphoid cells. Cell 1981, 25, 233–240. [Google Scholar] [CrossRef]
- Yoder, J.; Soman, N.S.; Verdine, G.L.; Bestor, T.H. DNA (cytosine-5)-methyltransferases in mouse cells and tissues. studies with a mechanism-based probe. J. Mol. Biol. 1997, 270, 385–395. [Google Scholar] [CrossRef]
- Pradhan, S.; Chin, H.G.; Jacobsen, S.E. SET7/9 Mediated Methylation of Non-Histone Proteins in Mammalian Cells. Proteins 2009, 4, 383–387. [Google Scholar] [CrossRef]
- Hata, K.; Okano, M.; Lei, H.; Li, E. Dnmt3L Cooperates with the Dnmt3 Family of de Novo DNA Methyltransferases to Establish Maternal Imprints in Mice. Development 2002, 129, 1983–1993. [Google Scholar] [CrossRef]
- Ooi, S.K.T.; Qiu, C.; Bernstein, E.; Li, K.; Jia, D.; Yang, Z.; Erdjument-Bromage, H.; Tempst, P.; Lin, S.-P.; Allis, C.D.; et al. DNMT3L connects unmethylated lysine 4 of histone H3 to de novo methylation of DNA. Nature 2007, 448, 714–717. [Google Scholar] [CrossRef] [Green Version]
- Fatemi, M.; Hermann, A.; Gowher, H.; Jeltsch, A. Dnmt3a and Dnmt1 functionally cooperate during de novo methylation of DNA. JBIC J. Biol. Inorg. Chem. 2002, 269, 4981–4984. [Google Scholar] [CrossRef] [PubMed]
- Okano, M.; Bell, D.W.; Haber, D.A.; Li, E. DNA Methyltransferases Dnmt3a and Dnmt3b Are Essential for De Novo Methylation and Mammalian Development. Cell 1999, 99, 247–257. [Google Scholar] [CrossRef] [Green Version]
- Tahiliani, M.; Koh, K.P.; Shen, Y.; Pastor, W.A.; Bandukwala, H.; Brudno, Y.; Agarwal, S.; Iyer, L.M.; Liu, D.R.; Aravind, L.; et al. Conversion of 5-Methylcytosine to 5-Hydroxymethylcytosine in Mammalian DNA by MLL Partner TET1. Science 2009, 324, 930–935. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kriaucionis, S.; Heintz, N. The Nuclear DNA Base 5-Hydroxymethylcytosine Is Present in Purkinje Neurons and the Brain. Science 2009, 324, 929–930. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- He, Y.-F.; Li, B.-Z.; Li, Z.; Liu, P.; Wang, Y.; Tang, Q.; Ding, J.; Jia, Y.; Chen, Z.; Li, L.; et al. Tet-Mediated Formation of 5-Carboxylcytosine and Its Excision by TDG in Mammalian DNA. Science 2011, 333, 1303–1307. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ito, S.; Shen, L.; Dai, Q.; Wu, S.C.; Collins, L.B.; Swenberg, J.A.; He, C.; Zhang, Y. Tet Proteins Can Convert 5-Methylcytosine to 5-Formylcytosine and 5-Carboxylcytosine. Science 2011, 333, 1300–1303. [Google Scholar] [CrossRef] [Green Version]
- Iurlaro, M.; Ficz, G.; Oxley, D.; Raiber, E.-A.; Bachman, M.; Booth, M.J.; Andrews, S.; Balasubramanian, S.; Reik, W. A screen for hydroxymethylcytosine and formylcytosine binding proteins suggests functions in transcription and chromatin regulation. Genome Biol. 2013, 14, R119. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Spruijt, C.; Gnerlich, F.; Smits, A.H.; Pfaffeneder, T.; Jansen, P.W.; Bauer, C.; Muenzel, M.; Wagner, M.; Müller, M.; Khan, F.; et al. Dynamic Readers for 5-(Hydroxy)Methylcytosine and Its Oxidized Derivatives. Cell 2013, 152, 1146–1159. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Maiti, A.; Drohat, A.C. Thymine DNA Glycosylase Can Rapidly Excise 5-Formylcytosine and 5-Carboxylcytosine: POTENTIAL IMPLICATIONS FOR ACTIVE DEMETHYLATION OF CpG SITES. J. Biol. Chem. 2011, 286, 35334–35338. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bannister, A.J.; Kouzarides, T. Regulation of chromatin by histone modifications. Cell Res. 2011, 21, 381–395. [Google Scholar] [CrossRef] [PubMed]
- Morgan, M.A.J.; Shilatifard, A. Reevaluating the roles of histone-modifying enzymes and their associated chromatin modifications in transcriptional regulation. Nat. Genet. 2020, 52, 1271–1281. [Google Scholar] [CrossRef] [PubMed]
- Falk, M.; Feodorova, Y.; Naumova, N.; Imakaev, M.; Lajoie, B.R.; Leonhardt, H.; Joffe, B.; Dekker, J.; Fudenberg, G.; Solovei, I.; et al. Heterochromatin drives compartmentalization of inverted and conventional nuclei. Nat. Cell Biol. 2019, 570, 395–399. [Google Scholar] [CrossRef]
- Allfrey, V.G.; Mirsky, A.E. Evidence for The Complete Dna-Dependence of Rna Synthesis in Isolated Thymus Nuclei. Proc. Natl. Acad. Sci. USA 1962, 48, 1590–1596. [Google Scholar] [CrossRef] [Green Version]
- Allfrey, G.; Faulkner, R.; Mirsky, A.E. Acetylation and methylation of histones and their possible role in the regulation of rna synthesis. Proc. Natl. Acad. Sci. USA 1964, 51, 786–794. [Google Scholar] [CrossRef] [Green Version]
- Brownell, J.E.; Zhou, J.; Ranalli, T.; Kobayashi, R.; Edmondson, D.G.; Roth, S.Y.; Allis, C.D. Tetrahymena Histone Acetyltransferase A: A Homolog to Yeast Gcn5p Linking Histone Acetylation to Gene Activation. Cell 1996, 84, 843–851. [Google Scholar] [CrossRef] [Green Version]
- Kuo, M.H.; Zhou, J.; Jambeck, P.; Churchill, M.E.; Allis, C.D. Histone Acetyltransferase Activity of Yeast Gcn5p Is Required for the Activation of Target Genes in vivo. Genes Dev. 1998, 12, 627–639. [Google Scholar] [CrossRef] [Green Version]
- Ogryzko, V.; Schiltz, R.; Russanova, V.; Howard, B.H.; Nakatani, Y. The Transcriptional Coactivators p300 and CBP Are Histone Acetyltransferases. Cell 1996, 87, 953–959. [Google Scholar] [CrossRef] [Green Version]
- Taunton, J.; Hassig, C.A.; Schreiber, S.L. A Mammalian Histone Deacetylase Related to the Yeast Transcriptional Regulator Rpd3p. Science 1996, 272, 408–411. [Google Scholar] [CrossRef]
- Rundlett, S.E.; Carmen, A.A.; Kobayashi, R.; Bavykin, S.; Turner, B.M.; Grunstein, M. HDA1 and RPD3 are members of distinct yeast histone deacetylase complexes that regulate silencing and transcription. Proc. Natl. Acad. Sci. USA 1996, 93, 14503–14508. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vidal, M.; Gaber, R.F. RPD3 encodes a second factor required to achieve maximum positive and negative transcriptional states in Saccharomyces cerevisiae. Mol. Cell. Biol. 1991, 11, 6317–6327. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, W.-M.; Inouye, C.; Zeng, Y.; Bearss, D.; Seto, E. Transcriptional repression by YY1 is mediated by interaction with a mammalian homolog of the yeast global regulator RPD3. Proc. Natl. Acad. Sci. USA 1996, 93, 12845–12850. [Google Scholar] [CrossRef] [Green Version]
- Heinzel, T.; Lavinsky, R.M.; Mullen, T.-M.; Soderstrom, M.; Laherty, C.D.; Torchia, J.A.; Yang, W.-M.; Brard, G.; Ngo, S.D.; Davie, J.; et al. A complex containing N-CoR, mSln3 and histone deacetylase mediates transcriptional repression. Nat. Cell Biol. 1997, 387, 43–48. [Google Scholar] [CrossRef]
- Alland, L.; Muhle, R.A.; Hou, H.; Potes, J.; Chin, L.; Schreiber-Agus, N.; Depinho, R.A. Role for N-CoR and histone deacetylase in Sin3-mediated transcriptional repression. Nat. Cell Biol. 1997, 387, 49–55. [Google Scholar] [CrossRef] [PubMed]
- Nagy, L.; Kao, H.-Y.; Chakravarti, D.; Lin, R.J.; Hassig, C.A.; Ayer, D.; Schreiber, S.L.; Evans, R.M. Nuclear Receptor Repression Mediated by a Complex Containing SMRT, mSin3A, and Histone Deacetylase. Cell 1997, 89, 373–380. [Google Scholar] [CrossRef] [Green Version]
- Merika, M.; Williams, A.J.; Chen, G.; Collins, T.; Thanos, D. Recruitment of CBP/P300 by the IFNβ Enhanceosome Is Required for Synergistic Activation of Transcription. Mol. Cell 1998, 1, 277–287. [Google Scholar] [CrossRef]
- Visel, A.; Blow, M.; Li, Z.; Zhang, T.; Akiyama, J.A.; Holt, A.; Plajzer-Frick, I.; Shoukry, M.; Wright, C.; Chen, F.; et al. ChIP-seq accurately predicts tissue-specific activity of enhancers. Nature 2009, 457, 854–858. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Heintzman, N.D.; Stuart, R.K.; Hon, G.; Fu, Y.; Ching, C.W.; Hawkins, R.D.; Barrera, L.O.; Van Calcar, S.; Qu, C.; Ching, K.A.; et al. Distinct and predictive chromatin signatures of transcriptional promoters and enhancers in the human genome. Nat. Genet. 2007, 39, 311–318. [Google Scholar] [CrossRef]
- Chen, X.; Xu, H.; Yuan, P.; Fang, F.; Huss, M.; Vega, V.B.; Wong, E.; Orlov, Y.; Zhang, W.; Jiang, J.; et al. Integration of External Signaling Pathways with the Core Transcriptional Network in Embryonic Stem Cells. Cell 2008, 133, 1106–1117. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, Z.; Zang, C.; Rosenfeld, J.A.; Schones, D.E.; Barski, A.; Cuddapah, S.; Cui, K.; Roh, T.-Y.; Peng, W.; Zhang, M.Q.; et al. Combinatorial patterns of histone acetylations and methylations in the human genome. Nat. Genet. 2008, 40, 897–903. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Roh, T.-Y.; Cuddapah, S.; Cui, K.; Zhao, K. The genomic landscape of histone modifications in human T cells. Proc. Natl. Acad. Sci. USA 2006, 103, 15782–15787. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Clapier, C.R.; Iwasa, J.; Cairns, B.R.; Peterson, C.L. Mechanisms of action and regulation of ATP-dependent chromatin-remodelling complexes. Nat. Rev. Mol. Cell Biol. 2017, 18, 407–422. [Google Scholar] [CrossRef] [PubMed]
- Peterson, C.L.; Herskowitz, I. Characterization of the yeast SWI1, SWI2, and SWI3 genes, which encode a global activator of transcription. Cell 1992, 68, 573–583. [Google Scholar] [CrossRef]
- Chiba, H.; Muramatsu, M.; Nomoto, A.; Kato, H. Two human homologues ofSaccharomyces cerevisiae SWI2/SNF2andDrosophila brahmaare transcriptional coactivators cooperating with the estrogen receptor and the retinoic acid receptor. Nucleic Acids Res. 1994, 22, 1815–1820. [Google Scholar] [CrossRef] [Green Version]
- Kwon, H.; Imbalzano, A.N.; Khavari, P.A.; Kingston, R.E.; Green, M.R. Nucleosome disruption and enhancement of activator binding by a human SW1/SNF complex. Nat. Cell Biol. 1994, 370, 477–481. [Google Scholar] [CrossRef]
- Wang, W.; Côté, J.; Xue, Y.; Zhou, S.; Khavari, P.A.; Biggar, S.R.; Muchardt, C.; Kalpana, G.V.; Goff, S.P.; Yaniv, M.; et al. Pu-rification and Biochemical Heterogeneity of the Mammalian SWI-SNF Complex. EMBO J. 1996, 15, 5370–5382. [Google Scholar] [CrossRef]
- Tsukiyama, T.; Daniel, C.; Tamkun, J.; Wu, C. ISWI, a member of the SWl2/SNF2 ATPase family, encodes the 140 kDa subunit of the nucleosome remodeling factor. Cell 1995, 83, 1021–1026. [Google Scholar] [CrossRef] [Green Version]
- Tsukiyama, T.; Wu, C. Purification and properties of an ATP-dependent nucleosome remodeling factor. Cell 1995, 83, 1011–1020. [Google Scholar] [CrossRef] [Green Version]
- Varga-Weisz, P.D.; Wilm, M.; Bonte, E.; Dumas, K.; Mann, M.; Becker, P. Chromatin-remodelling factor CHRAC contains the ATPases ISWI and topoisomerase II. Nat. Cell Biol. 1997, 388, 598–602. [Google Scholar] [CrossRef]
- Ito, T.; Bulger, M.; Pazin, M.; Kobayashi, R.; Kadonaga, J.T. ACF, an ISWI-Containing and ATP-Utilizing Chromatin Assembly and Remodeling Factor. Cell 1997, 90, 145–155. [Google Scholar] [CrossRef] [Green Version]
- Aihara, T.; Miyoshi, Y.; Koyama, K.; Suzuki, M.; Takahashi, E.; Monden, M.; Nakamura, Y. Cloning and mapping of SMARCA5 encoding hSNF2H, a novel human homologue of Drosophila ISWI. Cytogenet. Cell Genet. 1998, 81, 191–193. [Google Scholar] [CrossRef]
- Lazzaro, M.A.; Picketts, D.J. Cloning and characterization of the murine Imitation Switch (ISWI) genes: Differential expression patterns suggest distinct developmental roles for Snf2h and Snf2l. J. Neurochem. 2001, 77, 1145–1156. [Google Scholar] [CrossRef]
- Tolstorukov, M.Y.; Sansam, C.; Lu, P.; Koellhoffer, E.; Helming, K.C.; Alver, B.; Tillman, E.; Evans, J.A.; Wilson, B.G.; Park, P.J.; et al. Swi/Snf chromatin remodeling/tumor suppressor complex establishes nucleosome occupancy at target promoters. Proc. Natl. Acad. Sci. USA 2013, 110, 10165–10170. [Google Scholar] [CrossRef] [Green Version]
- Yen, K.; Vinayachandran, V.; Batta, K.; Koerber, R.T.; Pugh, B.F. Genome-wide Nucleosome Specificity and Directionality of Chromatin Remodelers. Cell 2012, 149, 1461–1473. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hu, G.; Schones, D.E.; Cui, K.; Ybarra, R.; Northrup, D.; Tang, Q.; Gattinoni, L.; Restifo, N.P.; Huang, S.; Zhao, K. Regulation of nucleosome landscape and transcription factor targeting at tissue-specific enhancers by BRG1. Genome Res. 2011, 21, 1650–1658. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wood, C.D.; Veenstra, H.; Khasnis, S.; Gunnell, A.; Webb, H.M.; Shannon-Lowe, C.; Andrews, S.; Osborne, C.; West, M.J. MYC activation and BCL2L11 silencing by a tumour virus through the large-scale reconfiguration of enhancer-promoter hubs. eLife 2016, 5, e18270. [Google Scholar] [CrossRef] [PubMed]
- Bossen, C.; Murre, C.S.; Chang, A.N.; Mansson, R.; Rodewald, H.-R.; Murre, C. The chromatin remodeler Brg1 activates enhancer repertoires to establish B cell identity and modulate cell growth. Nat. Immunol. 2015, 16, 775–784. [Google Scholar] [CrossRef] [Green Version]
- Alexander, J.; Hota, S.; He, D.; Thomas, S.; Ho, L.; Pennacchio, L.A.; Bruneau, B.G. Brg1 modulates enhancer activation in mesoderm lineage commitment. Development 2015, 142, 1418–1430. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ni, Z.; El Hassan, M.A.; Xu, Z.; Yu, T.; Bremner, R. The chromatin-remodeling enzyme BRG1 coordinates CIITA induction through many interdependent distal enhancers. Nat. Immunol. 2008, 9, 785–793. [Google Scholar] [CrossRef]
- Shi, J.; Whyte, W.A.; Zepeda-Mendoza, C.J.; Milazzo, J.P.; Shen, C.; Roe, J.-S.; Minder, J.L.; Mercan, F.; Wang, E.; Eckersley-Maslin, M.; et al. Role of SWI/SNF in acute leukemia maintenance and enhancer-mediated Myc regulation. Genes Dev. 2013, 27, 2648–2662. [Google Scholar] [CrossRef] [Green Version]
- Yu, Y.; Chen, Y.; Kim, B.; Wang, H.; Zhao, C.; He, X.; Liu, L.; Liu, W.; Wu, L.M.N.; Mao, M.; et al. Olig2 Targets Chromatin Remodelers to Enhancers to Initiate Oligodendrocyte Differentiation. Cell 2013, 152, 248–261. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alver, B.; Kim, K.H.; Lu, P.; Wang, X.; Manchester, H.E.; Wang, W.; Haswell, J.R.; Park, P.J.; Roberts, C.W.M. The SWI/SNF chromatin remodelling complex is required for maintenance of lineage specific enhancers. Nat. Commun. 2017, 8, 14648. [Google Scholar] [CrossRef] [PubMed]
- Nakayama, R.T.; Pulice, J.L.; Valencia, A.; McBride, M.; McKenzie, Z.M.; Gillespie, M.A.; Ku, W.L.; Teng, M.; Cui, K.; Williams, R.; et al. SMARCB1 is required for widespread BAF complex–mediated activation of enhancers and bivalent promoters. Nat. Genet. 2017, 49, 1613–1623. [Google Scholar] [CrossRef] [PubMed]
- Kwon, S.Y.; Grisan, V.; Jang, B.; Herbert, J.; Badenhorst, P. Genome-Wide Mapping Targets of the Metazoan Chromatin Remodeling Factor NURF Reveals Nucleosome Remodeling at Enhancers, Core Promoters and Gene Insulators. PLoS Genet. 2016, 12, e1005969. [Google Scholar] [CrossRef] [Green Version]
- Barisic, D.; Stadler, M.B.; Iurlaro, M.; Schübeler, D. Mammalian ISWI and SWI/SNF selectively mediate binding of distinct transcription factors. Nat. Cell Biol. 2019, 569, 136–140. [Google Scholar] [CrossRef]
- Qiu, Z.; Song, C.; Malakouti, N.; Murray, D.; Hariz, A.; Zimmerman, M.; Gygax, D.; Alhazmi, A.; Landry, J.W. Functional Interactions between NURF and Ctcf Regulate Gene Expression. Mol. Cell. Biol. 2015, 35, 224–237. [Google Scholar] [CrossRef] [Green Version]
- Wiechens, N.; Singh, V.; Gkikopoulos, T.; Schofield, P.; Rocha, S.; Owen-Hughes, T. The Chromatin Remodelling Enzymes SNF2H and SNF2L Position Nucleosomes adjacent to CTCF and Other Transcription Factors. PLoS Genet. 2016, 12, e1005940. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Langer, L.F.; Ward, J.M.; Archer, T.K. Tumor suppressor SMARCB1 suppresses super-enhancers to govern hESC lineage determination. eLife 2019, 8, e45672. [Google Scholar] [CrossRef] [PubMed]
- Whitehouse, I.; Rando, O.; Delrow, J.; Tsukiyama, T. Chromatin remodelling at promoters suppresses antisense transcription. Nature 2007, 450, 1031–1035. [Google Scholar] [CrossRef] [PubMed]
- Kadoch, C.; Hargreaves, D.C.; Hodges, H.C.; Elias, L.; Ho, L.; Ranish, J.; Crabtree, G.R. Proteomic and bioinformatic analysis of mammalian SWI/SNF complexes identifies extensive roles in human malignancy. Nat. Genet. 2013, 45, 592–601. [Google Scholar] [CrossRef]
- Geerdink, N.; Rotteveel, J.J.; Lammens, M.; Sistermans, E.; Heikens, G.T.; Gabreëls, F.J.M.; Mullaart, R.A.; Hamel, B.C.J. MECP2 Mutation in a Boy with Severe Neonatal Encephalopathy: Clinical, Neuropathological and Molecular Findings. Neuropediatrics 2002, 33, 33–36. [Google Scholar] [CrossRef]
- Bourdon, V.; Philippe, C.; Labrune, O.; Amsallem, D.; Arnould, C.; Jonveaux, P. A detailed analysis of the MECP2 gene: Prevalence of recurrent mutations and gross DNA rearrangements in Rett syndrome patients. Qual. Life Res. 2001, 108, 43–50. [Google Scholar] [CrossRef]
- Xiang, F.; Buervenich, S.; Nicolao, P.; Bailey, M.E.S.; Zhang, Z.; Anvret, M. Mutation screening in Rett syndrome patients. J. Med. Genet. 2000, 37, 250–255. [Google Scholar] [CrossRef]
- Bienvenu, T.; Carrié, A.; de Roux, N.; Vinet, M.-C.; Jonveaux, P.; Couvert, P.; Villard, L.; Arzimanoglou, A.; Beldjord, C.; Fontes, M.; et al. MECP2 mutations account for most cases of typical forms of Rett syndrome. Hum. Mol. Genet. 2000, 9, 1377–1384. [Google Scholar] [CrossRef]
- Klein, C.J.; Botuyan, M.-V.; Wu, Y.; Ward, C.J.; Nicholson, G.A.; Hammans, S.; Hojo, K.; Yamanishi, H.; Karpf, A.R.; Wallace, D.C.; et al. Mutations in DNMT1 cause hereditary sensory neuropathy with dementia and hearing loss. Nat. Genet. 2011, 43, 595–600. [Google Scholar] [CrossRef] [PubMed]
- Klein, C.J.; Bird, T.; Ertekin-Taner, N.; Lincoln, S.; Hjorth, R.; Wu, Y.; Kwok, J.; Mer, G.; Dyck, P.J.; Nicholson, G.A. DNMT1 mutation hot spot causes varied phenotypes of HSAN1 with dementia and hearing loss. Neurology 2013, 80, 824–828. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Winkelmann, J.; Lin, L.; Schormair, B.; Kornum, B.R.; Faraco, J.; Plazzi, G.; Melberg, A.; Cornelio, F.; Urban, A.E.; Pizza, F.; et al. Mutations in DNMT1 cause autosomal dominant cerebellar ataxia, deafness and narcolepsy. Hum. Mol. Genet. 2012, 21, 2205–2210. [Google Scholar] [CrossRef] [Green Version]
- Tatton-Brown, K.; Childhood Overgrowth Consortium; Seal, S.; Ruark, E.; Harmer, J.; Ramsay, E.; Duarte, S.D.V.; Zachariou, A.; Hanks, S.; O’Brien, E.; et al. Mutations in the DNA methyltransferase gene DNMT3A cause an overgrowth syndrome with intellectual disability. Nat. Genet. 2014, 46, 385–388. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shen, W.; Heeley, J.M.; Carlston, C.M.; Acuna-Hidalgo, R.; Nillesen, W.M.; Dent, K.M.; Douglas, G.V.; Levine, K.L.; Bayrak-Toydemir, P.; Marcelis, C.L.; et al. The spectrum of DNMT3A variants in Tatton-Brown-Rahman syndrome overlaps with that in hematologic malignancies. Am. J. Med. Genet. 2017, 173, 3022–3028. [Google Scholar] [CrossRef]
- Kosaki, R.; Terashima, H.; Kubota, M.; Kosaki, K. Acute myeloid leukemia-associatedDNMT3Ap.Arg882His mutation in a patient with Tatton-Brown-Rahman overgrowth syndrome as a constitutional mutation. Am. J. Med. Genet. 2017, 173, 250–253. [Google Scholar] [CrossRef] [PubMed]
- Heyn, P.; Logan, C.V.; Fluteau, A.; Challis, R.C.; Auchynnikava, T.; Martin, C.-A.; Marsh, J.A.; Taglini, F.; Kilanowski, F.; Parry, D.A.; et al. Gain-of-function DNMT3A mutations cause microcephalic dwarfism and hypermethylation of Polycomb-regulated regions. Nat. Genet. 2019, 51, 96–105. [Google Scholar] [CrossRef] [PubMed]
- Roelfsema, J.H.; White, S.J.; Ariyürek, Y.; Bartholdi, D.; Niedrist, D.; Papadia, F.; Bacino, C.A.; Dunnen, J.T.D.; van Ommen, G.-J.B.; Breuning, M.H.; et al. Genetic Heterogeneity in Rubinstein-Taybi Syndrome: Mutations in Both the CBP and EP300 Genes Cause Disease. Am. J. Hum. Genet. 2005, 76, 572–580. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hamilton, M.J.; Newbury-Ecob, R.; Holder-Espinasse, M.; Yau, S.; Lillis, S.; Hurst, J.A.; Clement, E.; Reardon, W.; Joss, S.; Hobson, E.; et al. Rubinstein–Taybi syndrome type 2: Report of nine new cases that extend the phenotypic and genotypic spectrum. Clin. Dysmorphol. 2016, 25, 135–145. [Google Scholar] [CrossRef] [PubMed]
- Foley, P.; Bunyan, D.; Stratton, J.; Dillon, M.; Lynch, S.A. Further case of Rubinstein-Taybi syndrome due to a deletion in EP300. Am. J. Med. Genet. 2009, 149, 997–1000. [Google Scholar] [CrossRef]
- Menke, L.A.; Gardeitchik, T.; Hammond, P.; Heimdal, K.R.; Houge, G.; Hufnagel, S.B.; Ji, J.; Johansson, S.; Kant, S.G.; Kinning, E.; et al. Further delineation of an entity caused by CREBBP and EP300 mutations but not resembling Rubinstein-Taybi syndrome. Am. J. Med. Genet. 2018, 176, 862–876. [Google Scholar] [CrossRef] [PubMed]
- Kalkhoven, E.; Roelfsema, J.H.; Teunissen, H.; den Boer, A.; Ariyurek, Y.; Zantema, A.; Breuning, M.H.; Hennekam, R.C.M.; Peters, D.J.M. Loss of CBP Acetyltransferase Activity by PHD Finger Mutations in Rubinstein-Taybi Syndrome. Human Mol. Genet. 2003, 12, 441–450. [Google Scholar] [CrossRef]
- Petrij, F.; Dauwerse, H.G.; Blough, R.I.; Giles, R.H.; Van Der Smagt, J.J.; Wallerstein, R.; Maaswinkel-Mooy, P.D.; van Karnebeek, C.; Van Ommen, G.-J.B.; Van Haeringen, A.; et al. Diagnostic analysis of the Rubinstein-Taybi syndrome: Five cosmids should be used for microdeletion detection and low number of protein truncating mutations. J. Med. Genet. 2000, 37, 168–176. [Google Scholar] [CrossRef] [Green Version]
- Coupry, I.; Roudaut, C.; Stef, M.; Delrue, M.A.; Marche, M.; Burgelin, I.; Taine, L.; Cruaud, C.; Lacombe, D.; Arveiler, B. Molecular Analysis of the CBP Gene in 60 Patients with Rubinstein-Taybi Syndrome. J. Med. Genet. 2002, 39, 415–421. [Google Scholar] [CrossRef] [Green Version]
- Menke, L.A.; van Belzen, M.J.; Alders, M.; Cristofoli, F.; Ehmke, N.; Fergelot, P.; Foster, A.; Gerkes, E.H.; Hoffer, M.J.V.; Horn, D.; et al. CREBBPmutations in individuals without Rubinstein-Taybi syndrome phenotype. Am. J. Med. Genet. 2016, 170, 2681–2693. [Google Scholar] [CrossRef] [PubMed]
- Angius, A.; Uva, P.; Oppo, M.; Persico, I.; Onano, S.; Olla, S.; Pes, V.; Perria, C.; Cuccuru, G.; Atzeni, R.; et al. Confirmation of a new phenotype in an individual with a variant in the last part of exon 30 of CREBBP. Am. J. Med. Genet. 2019, 179, 634–638. [Google Scholar] [CrossRef] [PubMed]
- Kennedy, J.; Goudie, D.; Blair, E.; Chandler, K.; Joss, S.; McKay, V.; Green, A.; Armstrong, R.; Lees, M.; Kamien, B.; et al. KAT6A Syndrome: Genotype–phenotype correlation in 76 patients with pathogenic KAT6A variants. Genet. Med. 2019, 21, 850–860. [Google Scholar] [CrossRef] [PubMed]
- Arboleda, V.; Lee, H.; Dorrani, N.; Zadeh, N.; Willis, M.; MacMurdo, C.F.; Manning, M.A.; Kwan, A.; Hudgins, L.; Barthelemy, F.; et al. De Novo Nonsense Mutations in KAT6A, a Lysine Acetyl-Transferase Gene, Cause a Syndrome Including Microcephaly and Global Developmental Delay. Am. J. Hum. Genet. 2015, 96, 498–506. [Google Scholar] [CrossRef] [Green Version]
- Millan, F.; Cho, M.T.; Retterer, K.; Monaghan, K.G.; Bai, R.; Vitazka, P.; Everman, D.B.; Smith, B.; Angle, B.; Roberts, V.; et al. Whole exome sequencing reveals de novo pathogenic variants inKAT6Aas a cause of a neurodevelopmental disorder. Am. J. Med. Genet. 2016, 170, 1791–1798. [Google Scholar] [CrossRef]
- Tham, E.; Lindstrand, A.; Santani, A.; Malmgren, H.; Nesbitt, A.; Dubbs, H.A.; Zackai, E.H.; Parker, M.J.; Millan, F.; Rosenbaum, K.; et al. Dominant Mutations in KAT6A Cause Intellectual Disability with Recognizable Syndromic Features. Am. J. Hum. Genet. 2015, 96, 507–513. [Google Scholar] [CrossRef] [Green Version]
- Szakszon, K.; Salpietro, C.; Kakar, N.; Knegt, A.C.; Oláh, É.; Dallapiccola, B.; Borck, G. De novo mutations of the gene encoding the histone acetyltransferase KAT6B in two patients with Say-Barber/Biesecker/Young-Simpson syndrome. Am. J. Med. Genet. Part A 2013, 161, 884–888. [Google Scholar] [CrossRef] [PubMed]
- Szakszon, K.; Berényi, E.; Jakab, A.; Bessenyei, B.; Balogh, E.; Köbling, T.; Szilvássy, J.; Knegt, A.C.; Oláh, É. Blepharophimosis mental retardation syndrome Say-Barber/Biesecker/Young-Simpson type—New findings with neuroimaging. Am. J. Med. Genet. Part A 2011, 155, 634–637. [Google Scholar] [CrossRef] [PubMed]
- Yates, T.M.; Langley, C.L.; Grozeva, D.; Raymond, F.L.; Johnson, D.S.; DDD Study. Novel KAT6B proximal familial variant expands genotypic and phenotypic spectrum. Clin. Genet. 2019, 95, 334–335. [Google Scholar] [CrossRef]
- Clayton-Smith, J.; O’Sullivan, J.; Daly, S.; Bhaskar, S.; Day, R.; Anderson, B.; Voss, A.K.; Thomas, T.; Biesecker, L.G.; Smith, P.; et al. Whole-Exome-Sequencing Identifies Mutations in Histone Acetyltransferase Gene KAT6B in Individuals with the Say-Barber-Biesecker Variant of Ohdo Syndrome. Am. J. Hum. Genet. 2011, 89, 675–681. [Google Scholar] [CrossRef] [Green Version]
- Campeau, P.M.; Kim, J.C.; Lu, J.T.; Schwartzentruber, J.A.; Abdul-Rahman, O.A.; Schlaubitz, S.; Murdock, D.M.; Jiang, M.-M.; Lammer, E.J.; Enns, G.M.; et al. Mutations in KAT6B, Encoding a Histone Acetyltransferase, Cause Genitopatellar Syndrome. Am. J. Hum. Genet. 2012, 90, 282–289. [Google Scholar] [CrossRef] [Green Version]
- Simpson, M.; Deshpande, C.; Dafou, D.; Vissers, L.; Woollard, W.J.; Holder, S.E.; Gillessen-Kaesbach, G.; Derks, R.; White, S.M.; Cohen-Snuijf, R.; et al. De Novo Mutations of the Gene Encoding the Histone Acetyltransferase KAT6B Cause Genitopatellar Syndrome. Am. J. Hum. Genet. 2012, 90, 290–294. [Google Scholar] [CrossRef] [Green Version]
- Li, Y.; Haarhuis, J.H.I.; Cacciatore, Á.S.; Oldenkamp, R.; Van Ruiten, M.S.; Willems, L.; Teunissen, H.; Muir, K.W.; De Wit, E.; Rowland, B.D.; et al. The structural basis for cohesin–CTCF-anchored loops. Nat. Cell Biol. 2020, 578, 472–476. [Google Scholar] [CrossRef] [PubMed]
- Simon, D.; Laloo, B.; Barillot, M.; Barnetche, T.; Blanchard, C.; Rooryck, C.; Marche, M.; Burgelin, I.; Coupry, I.; Chassaing, N.; et al. A mutation in the 3′-UTR of the HDAC6 gene abolishing the post-transcriptional regulation mediated by hsa-miR-433 is linked to a new form of dominant X-linked chondrodysplasia. Hum. Mol. Genet. 2010, 19, 2015–2027. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chassaing, N.; Siani, V.; Carles, D.; Delezoide, A.L.; Alberti, E.M.; Battin, J.; Chateil, J.-F.; Gilbert-Dussardier, B.; Coupry, I.; Arveiler, B.; et al. X-linked dominant chondrodysplasia with platyspondyly, distinctive brachydactyly, hydrocephaly, and microphthalmia. Am. J. Med. Genet. 2005, 136, 307–312. [Google Scholar] [CrossRef] [PubMed]
- Harakalova, M.; Boogaard, M.-J.V.D.; Sinke, R.; Van Lieshout, S.; Van Tuil, M.C.; Duran, K.; Renkens, I.; Terhal, P.A.; de Kovel, C.; Nijman, I.; et al. X-exome sequencing identifies aHDAC8variant in a large pedigree with X-linked intellectual disability, truncal obesity, gynaecomastia, hypogonadism and unusual face. J. Med. Genet. 2012, 49, 539–543. [Google Scholar] [CrossRef] [PubMed]
- Deardorff, M.A.; Bando, M.; Nakato, R.; Watrin, E.; Itoh, T.; Minamino, M.; Saitoh, K.; Komata, M.; Katou, Y.; Clark, D.; et al. HDAC8 mutations in Cornelia de Lange syndrome affect the cohesin acetylation cycle. Nature 2012, 489, 313–317. [Google Scholar] [CrossRef]
- Libert, S.; Pointer, K.; Bell, E.L.; Das, A.; Cohen, D.E.; Asara, J.M.; Kapur, K.; Bergmann, S.; Preisig, M.; Otowa, T.; et al. SIRT1 Activates MAO-A in the Brain to Mediate Anxiety and Exploratory Drive. Cell 2011, 147, 1459–1472. [Google Scholar] [CrossRef] [Green Version]
- Wolff, D.; Endele, S.; Azzarello-Burri, S.; Hoyer, J.; Zweier, M.; Schanze, I.; Schmitt, B.; Rauch, A.; Reis, A.; Zweier, C. In-Frame Deletion and Missense Mutations of the C-Terminal Helicase Domain of SMARCA2 in Three Patients with Nicolaides-Baraitser Syndrome. Mol. Syndr. 2012, 2, 237–244. [Google Scholar] [CrossRef] [Green Version]
- Van Houdt, J.K.J.; Nowakowska, B.A.; Sousa, S.B.; Van Schaik, B.D.C.; Seuntjens, E.; Avonce, N.; Sifrim, A.; Abdul-Rahman, O.A.; Boogaard, M.-J.H.V.D.; Bottani, A.; et al. Heterozygous missense mutations in SMARCA2 cause Nicolaides-Baraitser syndrome. Nat. Genet. 2012, 44, 445–449. [Google Scholar] [CrossRef] [PubMed]
- Koga, M.; Ishiguro, H.; Yazaki, S.; Horiuchi, Y.; Arai, M.; Niizato, K.; Iritani, S.; Itokawa, M.; Inada, T.; Iwata, N.; et al. Involvement of SMARCA2/BRM in the SWI/SNF chromatin-remodeling complex in schizophrenia. Hum. Mol. Genet. 2009, 18, 2483–2494. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tsurusaki, Y.; Okamoto, N.; Ohashi, H.; Kosho, T.; Imai, Y.; Hibi-Ko, Y.; Kaname, T.; Naritomi, K.; Kawame, H.; Wakui, K.; et al. Mutations affecting components of the SWI/SNF complex cause Coffin-Siris syndrome. Nat. Genet. 2012, 44, 376–378. [Google Scholar] [CrossRef]
- Sévenet, N.; Sheridan, E.; Amram, D.; Schneider, P.; Handgretinger, R.; Delattre, O. Constitutional Mutations of the hSNF5/INI1 Gene Predispose to a Variety of Cancers. Am. J. Hum. Genet. 1999, 65, 1342–1348. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Taylor, M.D.; Gokgoz, N.; Andrulis, I.L.; Mainprize, T.G.; Drake, J.M.; Rutka, J.T. Familial Posterior Fossa Brain Tumors of Infancy Secondary to Germline Mutation of the hSNF5 Gene. Am. J. Hum. Genet. 2000, 66, 1403–1406. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Christiaans, I.; Kenter, S.B.; Brink, H.C.; Van Os, T.A.M.; Baas, F.; Munckhof, P.V.D.; Kidd, A.M.J.; Hulsebos, T.J.M. Germline SMARCB1 mutation and somatic NF2 mutations in familial multiple meningiomas. J. Med. Genet. 2010, 48, 93–97. [Google Scholar] [CrossRef]
- Hulsebos, T.J.; Plomp, A.S.; Wolterman, R.A.; Robanus-Maandag, E.C.; Baas, F.; Wesseling, P. Germline Mutation of INI1/SMARCB1 in Familial Schwannomatosis. Am. J. Hum. Genet. 2007, 80, 805–810. [Google Scholar] [CrossRef] [Green Version]
- Sestini, R.; Bacci, C.; Provenzano, A.; Genuardi, M.; Papi, L. Evidence of a four-hit mechanism involvingSMARCB1andNF2in schwannomatosis-associated schwannomas. Hum. Mutat. 2008, 29, 227–231. [Google Scholar] [CrossRef]
- Hadfield, K.D.; Newman, W.; Bowers, N.L.; Wallace, A.; Bolger, C.; Colley, A.; McCann, E.; Trump, D.; Prescott, T.; Evans, G. Molecular characterisation of SMARCB1 and NF2 in familial and sporadic schwannomatosis. J. Med. Genet. 2008, 45, 332–339. [Google Scholar] [CrossRef]
- Machol, K.; Rousseau, J.; Ehresmann, S.; Garcia, T.; Nguyen, T.T.M.; Spillmann, R.C.; Sullivan, J.A.; Shashi, V.; Jiang, Y.-H.; Stong, N.; et al. Expanding the Spectrum of BAF-Related Disorders: De Novo Variants in SMARCC2 Cause a Syndrome with Intellectual Disability and Developmental Delay. Am. J. Hum. Genet. 2019, 104, 164–178. [Google Scholar] [CrossRef] [Green Version]
- Nixon, K.C.; Rousseau, J.; Stone, M.H.; Sarikahya, M.; Ehresmann, S.; Mizuno, S.; Matsumoto, N.; Miyake, N.; Baralle, D.; McKee, S.; et al. A Syndromic Neurodevelopmental Disorder Caused by Mutations in SMARCD1, a Core SWI/SNF Subunit Needed for Context-Dependent Neuronal Gene Regulation in Flies. Am. J. Hum. Genet. 2019, 104, 596–610. [Google Scholar] [CrossRef] [Green Version]
- Smith, M.; O’Sullivan, J.; Bhaskar, S.S.; Hadfield, K.D.; Poke, G.; Caird, J.; Sharif, S.; Eccles, D.M.; FitzPatrick, D.R.; Rawluk, D.; et al. Loss-of-function mutations in SMARCE1 cause an inherited disorder of multiple spinal meningiomas. Nat. Genet. 2013, 45, 295–298. [Google Scholar] [CrossRef]
- Wieczorek, D.; Bögershausen, N.; Beleggia, F.; Steiner-Haldenstätt, S.; Pohl, E.; Li, Y.; Milz, E.; Martin, M.; Thiele, H.; Altmüller, J.; et al. A comprehensive molecular study on Coffin–Siris and Nicolaides–Baraitser syndromes identifies a broad molecular and clinical spectrum converging on altered chromatin remodeling. Hum. Mol. Genet. 2013, 22, 5121–5135. [Google Scholar] [CrossRef]
- Karaca, E.; Harel, T.; Pehlivan, D.; Jhangiani, S.N.; Gambin, T.; Akdemir, Z.C.; Gonzaga-Jauregui, C.; Erdin, S.; Bayram, Y.; Campbell, I.; et al. Genes that Affect Brain Structure and Function Identified by Rare Variant Analyses of Mendelian Neurologic Disease. Neuron 2015, 88, 499–513. [Google Scholar] [CrossRef] [Green Version]
- Fichera, M.; Failla, P.; Saccuzzo, L.; Miceli, M.; Salvo, E.; Castiglia, L.; Galesi, O.; Grillo, L.; Cali, F.; Greco, D.; et al. Mutations in ACTL6B, coding for a subunit of the neuron-specific chromatin remodeling complex nBAF, cause early onset severe developmental and epileptic encephalopathy with brain hypomyelination and cerebellar atrophy. Qual. Life Res. 2019, 138, 187–198. [Google Scholar] [CrossRef]
- Maddirevula, S.; Bs, F.A.; Al-Owain, M.; Al Muhaizea, M.A.; Kayyali, H.R.; Alhashem, A.; Rahbeeni, Z.; Al-Otaibi, M.; Alzaidan, H.I.; Ms, A.B.; et al. Autozygome and high throughput confirmation of disease genes candidacy. Genet. Med. 2019, 21, 736–742. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bell, S.; Rousseau, J.; Peng, H.; Aouabed, Z.; Priam, P.; Theroux, J.-F.; Jefri, M.; Tanti, A.; Wu, H.; Kolobova, I.; et al. Mutations in ACTL6B Cause Neurodevelopmental Deficits and Epilepsy and Lead to Loss of Dendrites in Human Neurons. Am. J. Hum. Genet. 2019, 104, 815–834. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Santen, G.W.; Aten, E.; Silfhout, A.T.V.-V.; Pottinger, C.; Van Bon, B.W.; Van Minderhout, I.J.; Snowdowne, R.; Van Der Lans, C.A.; Boogaard, M.; Linssen, M.M.; et al. Coffin-Siris Syndrome and the BAF Complex: Genotype-Phenotype Study in 63 Patients. Hum. Mutat. 2013, 34, 1519–1528. [Google Scholar] [CrossRef] [PubMed]
- Zweier, M.; Peippo, M.M.; Pöyhönen, M.; Kääriäinen, H.; Begemann, A.; Joset, P.; Oneda, B.; Rauch, A. The HHID Syndrome of Hypertrichosis, Hyperkeratosis, Abnormal Corpus Callosum, Intellectual Disability, and Minor Anomalies Is Caused by Mutations in ARID1B. Am. J. Med. Genet. 2017, 173, 1440–1443. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hoyer, J.; Ekici, A.B.; Endele, S.; Popp, B.; Zweier, C.; Wiesener, A.; Wohlleber, E.; Dufke, A.; Rossier, E.; Petsch, C.; et al. Haploinsufficiency of ARID1B, a Member of the SWI/SNF-A Chromatin-Remodeling Complex, Is a Frequent Cause of Intellectual Disability. Am. J. Hum. Genet. 2012, 90, 565–572. [Google Scholar] [CrossRef] [Green Version]
- Van Paemel, R.; De Bruyne, P.; Van Der Straaten, S.; D’Hondt, M.; Fränkel, U.; Dheedene, A.; Menten, B.; Callewaert, B. Confirmation of an ARID2 defect in SWI/SNF-related intellectual disability. Am. J. Med. Genet. 2017, 173, 3104–3108. [Google Scholar] [CrossRef]
- Shang, L.; Cho, M.T.; Retterer, K.; Folk, L.; Humberson, J.; Rohena, L.; Sidhu, A.; Saliganan, S.; Iglesias, A.; Vitazka, P.; et al. Mutations in ARID2 are associated with intellectual disabilities. Neurogenetics 2015, 16, 307–314. [Google Scholar] [CrossRef]
- Bramswig, N.C.; Caluseriu, O.; Lüdecke, H.-J.; Bolduc, F.V.; Noel, N.C.L.; Wieland, T.; Surowy, H.; Christen, H.-J.; Engels, H.; Strom, T.M.; et al. Heterozygosity for ARID2 loss-of-function mutations in individuals with a Coffin–Siris syndrome-like phenotype. Qual. Life Res. 2017, 136, 297–305. [Google Scholar] [CrossRef] [PubMed]
- Gearing, M.; Juncos, J.L.; Procaccio, V.; Gutekunst, C.-A.; Marino-Rodriguez, E.M.; Gyure, K.A.; Ono, S.; Santoianni, R.; Krawiecki, N.S.; Wallace, D.C.; et al. Aggregation of actin and cofilin in identical twins with juvenile-onset dystonia. Ann. Neurol. 2002, 52, 465–476. [Google Scholar] [CrossRef] [Green Version]
- Procaccio, V.; Salazar, G.; Ono, S.; Styers, M.L.; Gearing, M.; Davila, A.; Jimenez, R.; Juncos, J.; Gutekunst, C.-A.; Meroni, G.; et al. A Mutation of β-Actin That Alters Depolymerization Dynamics Is Associated with Autosomal Dominant Developmental Malformations, Deafness, and Dystonia. Am. J. Hum. Genet. 2006, 78, 947–960. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Di Donato, N.; Rump, A.; Koenig, R.; Der Kaloustian, V.M.; Halal, F.; Sonntag, K.; Krause, C.; Hackmann, K.; Hahn, G.; Schrock, E.; et al. Severe forms of Baraitser–Winter syndrome are caused by ACTB mutations rather than ACTG1 mutations. Eur. J. Hum. Genet. 2013, 22, 179–183. [Google Scholar] [CrossRef] [Green Version]
- Rivière, J.-B.; Van Bon, B.W.M.; Hoischen, A.; Kholmanskikh, S.S.; O’Roak, B.; Gilissen, C.; Gijsen, S.; Sullivan, C.T.; Christian, S.L.; Abdul-Rahman, O.A.; et al. De novo mutations in the actin genes ACTB and ACTG1 cause Baraitser-Winter syndrome. Nat. Genet. 2012, 44, 440–444. [Google Scholar] [CrossRef] [Green Version]
- Vasileiou, G.; Vergarajauregui, S.; Endele, S.; Popp, B.; Büttner, C.; Ekici, A.B.; Gerard, M.; Bramswig, N.C.; Albrecht, B.; Clayton-Smith, J.; et al. Mutations in the BAF-Complex Subunit DPF2 Are Associated with Coffin-Siris Syndrome. Am. J. Hum. Genet. 2018, 102, 468–479. [Google Scholar] [CrossRef] [Green Version]
- Dias, C.; Estruch, S.B.; Graham, S.; McRae, J.; Sawiak, S.; Hurst, J.A.; Joss, S.K.; Holder, S.E.; Morton, J.E.; Turner, C.; et al. BCL11A Haploinsufficiency Causes an Intellectual Disability Syndrome and Dysregulates Transcription. Am. J. Hum. Genet. 2016, 99, 253–274. [Google Scholar] [CrossRef] [Green Version]
- Lessel, D.; Gehbauer, C.; Bramswig, N.C.; Schluth-Bolard, C.; Venkataramanappa, S.; van Gassen, K.L.I.; Hempel, M.; Haack, T.B.; Baresic, A.; Genetti, C.A.; et al. BCL11B mutations in patients affected by a neurodevelopmental disorder with reduced type 2 innate lymphoid cells. Brain 2018, 141, 2299–2311. [Google Scholar] [CrossRef] [Green Version]
- Neale, B.M.; Kou, Y.; Liu, L.; Ma’Ayan, A.; Samocha, K.E.; Sabo, A.; Lin, C.-F.; Stevens, C.; Wang, L.-S.; Makarov, V.; et al. Patterns and rates of exonic de novo mutations in autism spectrum disorders. Nature 2012, 485, 242–245. [Google Scholar] [CrossRef]
- Borck, G.; Hög, F.; Dentici, M.L.; Tan, P.L.; Sowada, N.; Medeira, A.; Gueneau, L.; Thiele, H.; Kousi, M.; Lepri, F.; et al. BRF1mutations alter RNA polymerase III–dependent transcription and cause neurodevelopmental anomalies. Genome Res. 2015, 25, 155–166. [Google Scholar] [CrossRef] [Green Version]
- Stankiewicz, P.; Khan, T.; Szafranski, P.; Slattery, L.; Streff, H.; Vetrini, F.; Bernstein, J.A.; Brown, C.W.; Rosenfeld, J.A.; Rednam, S.; et al. Haploinsufficiency of the Chromatin Remodeler BPTF Causes Syndromic Developmental and Speech Delay, Postnatal Microcephaly, and Dysmorphic Features. Am. J. Hum. Genet. 2017, 101, 503–515. [Google Scholar] [CrossRef] [Green Version]
- Homann, O.R.; Misura, K.; Lamas, E.; Sandrock, R.W.; Nelson, P.; McDonough, S.I.; DeLisi, L.E. Whole-genome sequencing in multiplex families with psychoses reveals mutations in the SHANK2 and SMARCA1 genes segregating with illness. Mol. Psychiatry 2016, 21, 1690–1695. [Google Scholar] [CrossRef] [Green Version]
- Lopes, F.; Barbosa, M.; Ameur, A.; Soares, G.; De Sá, J.; Dias, A.I.; Oliveira, G.; Cabral, P.; Temudo, T.; Calado, E.; et al. Identification of novel genetic causes of Rett syndrome-likephenotypes. J. Med. Genet. 2016, 53, 190–199. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zaghlool, A.; Halvardson, J.; Zhao, J.J.; Etemadikhah, M.; Kalushkova, A.; Konska, K.; Jernberg-Wiklund, H.; Thuresson, A.-C.; Feuk, L. A Role for the Chromatin-Remodeling Factor BAZ1A in Neurodevelopment. Hum. Mutat. 2016, 37, 964–975. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pilarowski, G.; Vernon, H.; Applegate, C.D.; Boukas, L.; Cho, M.T.; Gurnett, C.; Benke, P.J.; Beaver, E.; Heeley, J.M.; Medne, L.; et al. Missense variants in the chromatin remodeler CHD1 are associated with neurodevelopmental disability. J. Med. Genet. 2018, 55, 561–566. [Google Scholar] [CrossRef]
- Suls, A.; Jaehn, J.A.; Kecskés, A.; Weber, Y.; Weckhuysen, S.; Craiu, D.; Siekierska, A.; Djémié, T.; Afrikanova, T.; Gormley, P.; et al. De Novo Loss-of-Function Mutations in CHD2 Cause a Fever-Sensitive Myoclonic Epileptic Encephalopathy Sharing Features with Dravet Syndrome. Am. J. Hum. Genet. 2013, 93, 967–975. [Google Scholar] [CrossRef] [Green Version]
- Rauch, A.; Wieczorek, D.; Graf, E.; Wieland, T.; Endele, S.; Schwarzmayr, T.; Albrecht, B.; Bartholdi, D.; Beygo, J.; Di Donato, N.; et al. Range of genetic mutations associated with severe non-syndromic sporadic intellectual disability: An exome sequencing study. Lancet 2012, 380, 1674–1682. [Google Scholar] [CrossRef]
- Petersen, A.K.; Streff, H.; Tokita, M.; Bostwick, B.L. The first reported case of an inherited pathogenic CHD2 variant in a clinically affected mother and daughter. Am. J. Med. Genet. 2018, 176, 1667–1669. [Google Scholar] [CrossRef] [PubMed]
- Carvill, G.L.; Heavin, S.B.; Yendle, S.C.; McMahon, J.M.; O’Roak, B.J.; Cook, J.; Khan, A.; Dorschner, M.O.; Weaver, M.; Calvert, S.; et al. Targeted resequencing in epileptic encephalopathies identifies de novo mutations in CHD2 and SYNGAP1. Nat. Genet. 2013, 45, 825–830. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Eising, E.; Carrion-Castillo, A.; Vino, A.; Strand, E.A.; Jakielski, K.J.; Scerri, T.S.; Hildebrand, M.S.; Webster, R.; Ma, A.; Mazoyer, B.; et al. A set of regulatory genes co-expressed in embryonic human brain is implicated in disrupted speech development. Mol. Psychiatry 2019, 24, 1065–1078. [Google Scholar] [CrossRef] [Green Version]
- Weiss, K.; Terhal, P.A.; Cohen, L.; Bruccoleri, M.; Irving, M.; Martinez, A.F.; Rosenfeld, J.A.; Machol, K.; Yang, Y.; Liu, P.; et al. De Novo Mutations in CHD4, an ATP-Dependent Chromatin Remodeler Gene, Cause an Intellectual Disability Syndrome with Distinctive Dysmorphisms. Am. J. Hum. Genet. 2016, 99, 934–941. [Google Scholar] [CrossRef] [Green Version]
- Sifrim, A.; Hitz, M.-P.; Wilsdon, A.; Breckpot, J.; Al Turki, S.H.; Thienpont, B.; McRae, J.; Fitzgerald, T.W.; Singh, T.; Swaminathan, G.J.; et al. Distinct genetic architectures for syndromic and nonsyndromic congenital heart defects identified by exome sequencing. Nat. Genet. 2016, 48, 1060–1065. [Google Scholar] [CrossRef]
- Gabriele, M.; Silfhout, A.T.V.-V.; Germain, P.-L.; Vitriolo, A.; Kumar, R.; Douglas, E.; Haan, E.; Kosaki, K.; Takenouchi, T.; Rauch, A.; et al. YY1 Haploinsufficiency Causes an Intellectual Disability Syndrome Featuring Transcriptional and Chromatin Dysfunction. Am. J. Hum. Genet. 2017, 100, 907–925. [Google Scholar] [CrossRef] [Green Version]
- Alazami, A.M.; Patel, N.; Shamseldin, H.E.; Anazi, S.; Al-Dosari, M.S.; Alzahrani, F.; Hijazi, H.; Alshammari, M.; Aldahmesh, M.A.; Salih, M.; et al. Accelerating Novel Candidate Gene Discovery in Neurogenetic Disorders via Whole-Exome Sequencing of Prescreened Multiplex Consanguineous Families. Cell Rep. 2015, 10, 148–161. [Google Scholar] [CrossRef] [Green Version]
- Hood, R.L.; Lines, M.A.; Nikkel, S.M.; Schwartzentruber, J.; Beaulieu, C.; Nowaczyk, M.J.; Allanson, J.; Kim, C.; Wieczorek, D.; Moilanen, J.; et al. Mutations in SRCAP, Encoding SNF2-Related CREBBP Activator Protein, Cause Floating-Harbor Syndrome. Am. J. Hum. Genet. 2012, 90, 308–313. [Google Scholar] [CrossRef] [Green Version]
- Musio, A.; Selicorni, A.; Focarelli, M.L.; Gervasini, C.; Milani, D.; Russo, S.; Vezzoni, P.; Larizza, L. X-linked Cornelia de Lange syndrome owing to SMC1L1 mutations. Nat. Genet. 2006, 38, 528–530. [Google Scholar] [CrossRef]
- Deardorff, M.A.; Kaur, M.; Yaeger, D.; Rampuria, A.; Korolev, S.; Pie, J.; Gil-Rodríguez, C.; Arnedo, M.; Loeys, B.; Kline, A.D.; et al. Mutations in Cohesin Complex Members SMC3 and SMC1A Cause a Mild Variant of Cornelia de Lange Syndrome with Predominant Mental Retardation. Am. J. Hum. Genet. 2007, 80, 485–494. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kruszka, P.; Berger, S.; Casa, V.; Dekker, M.R.; Gaesser, J.; Weiss, K.; Martinez, A.F.; Murdock, D.R.; Louie, R.J.; Prijoles, E.J.; et al. Cohesin complex-associated holoprosencephaly. Brain 2019, 142, 2631–2643. [Google Scholar] [CrossRef] [PubMed]
- Symonds, J.; Joss, S.; Metcalfe, K.A.; Somarathi, S.; Cruden, J.; Devlin, A.M.; Donaldson, A.; Di Donato, N.; FitzPatrick, D.R.; Kaiser, F.J.; et al. Heterozygous truncation mutations of theSMC1Agene cause a severe early onset epilepsy with cluster seizures in females: Detailed phenotyping of 10 new cases. Epilepsia 2017, 58, 565–575. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Goldstein, J.H.; Tim-Aroon, T.; Shieh, J.; Merrill, M.; Deeb, K.K.; Zhang, S.; Bass, N.E.; Bedoyan, J.K. Novel SMC1A frameshift mutations in children with developmental delay and epilepsy. Eur. J. Med. Genet. 2015, 58, 562–568. [Google Scholar] [CrossRef]
- Lebrun, N.; Lebon, S.; Jeannet, P.-Y.; Jacquemont, S.; Billuart, P.; Bienvenu, T. Early-onset encephalopathy with epilepsy associated with a novel splice site mutation inSMC1A. Am. J. Med. Genet. 2015, 167, 3076–3081. [Google Scholar] [CrossRef]
- Jansen, S.; Kleefstra, T.; Willemsen, M.H.; De Vries, P.; Pfundt, R.; Hehir-Kwa, J.Y.; Gilissen, C.; Veltman, J.A.; De Vries, B.B.A.; Vissers, L.E.L.M. De novoloss-of-function mutations in X-linkedSMC1Acause severe ID and therapy-resistant epilepsy in females: Expanding the phenotypic spectrum. Clin. Genet. 2016, 90, 413–419. [Google Scholar] [CrossRef]
- Ansari, M.; Poke, G.; Ferry, Q.; Williamson, K.; Aldridge, R.; Meynert, A.M.; Bengani, H.; Chan, C.Y.; Kayserili, H.; Avci, Ş.; et al. Genetic heterogeneity in Cornelia de Lange syndrome (CdLS) and CdLS-like phenotypes with observed and predicted levels of mosaicism. J. Med. Genet. 2014, 51, 659–668. [Google Scholar] [CrossRef] [Green Version]
- Gil-Rodríguez, M.C.; Deardorff, M.A.; Ansari, M.; Tan, C.A.; Parenti, I.; Baquero-Montoya, C.; Ousager, L.B.; Puisac, B.; Hernández-Marcos, M.; Teresa-Rodrigo, M.E.; et al. De NovoHeterozygous Mutations inSMC3Cause a Range of Cornelia de Lange Syndrome-Overlapping Phenotypes. Hum. Mutat. 2015, 36, 454–462. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Boyle, M.; Jespersgaard, C.; Nazaryan, L.; Bisgaard, A.-M.; Tümer, Z. A novelRAD21variant associated with intrafamilial phenotypic variation in Cornelia de Lange syndrome—Review of the literature. Clin. Genet. 2016, 91, 647–649. [Google Scholar] [CrossRef] [PubMed]
- Deardorff, M.A.; Wilde, J.; Albrecht, M.; Dickinson, E.; Tennstedt, S.; Braunholz, D.; Mönnich, M.; Yan, Y.; Xu, W.; Gil-Rodríguez, M.C.; et al. RAD21 Mutations Cause a Human Cohesinopathy. Am. J. Hum. Genet. 2012, 90, 1014–1027. [Google Scholar] [CrossRef] [Green Version]
- Goel, H.; Parasivam, G. Another case of holoprosencephaly associated with RAD21 loss-of-function variant. Brain 2020, 143, e64. [Google Scholar] [CrossRef]
- Lehalle, D.; Mosca-Boidron, A.-L.; Begtrup, A.; Boute-Benejean, O.; Charles, P.; Cho, M.T.; Clarkson, A.; Devinsky, O.; Duffourd, Y.; Duplomb-Jego, L.; et al. STAG1mutations cause a novel cohesinopathy characterised by unspecific syndromic intellectual disability. J. Med. Genet. 2017, 54, 479–488. [Google Scholar] [CrossRef]
- Yuan, B.; DDD study; Neira, J.; Pehlivan, D.; Santiago-Sim, T.; Song, X.; Rosenfeld, J.; Posey, J.; Patel, V.; Jin, W.; et al. Clinical exome sequencing reveals locus heterogeneity and phenotypic variability of cohesinopathies. Genet. Med. 2018, 21, 663–675. [Google Scholar] [CrossRef]
- Soardi, F.C.; Machado-Silva, A.; Linhares, N.; Zheng, G.; Qu, Q.; Pena, H.B.; Martins, T.M.D.M.; Vieira, H.G.S.; Pereira, N.B.; Melo-Minardi, R.C.; et al. Familial STAG2 germline mutation defines a new human cohesinopathy. NPJ Genom. Med. 2017, 2, 7. [Google Scholar] [CrossRef] [PubMed]
- Mullegama, S.V.; Klein, S.D.; Mulatinho, M.V.; Senaratne, T.N.; Singh, K.; Nguyen, D.C.; Gallant, N.M.; Strom, S.P.; Ghahremani, S.; Rao, N.P.; et al. De novo loss-of-function variants inSTAG2are associated with developmental delay, microcephaly, and congenital anomalies. Am. J. Med. Genet. 2017, 173, 1319–1327. [Google Scholar] [CrossRef] [PubMed]
- Krantz, I.D.; McCallum, J.; DeScipio, C.; Kaur, M.; Gillis, L.A.; Yaeger, D.; Jukofsky, L.; Wasserman, N.; Bottani, A.; Morris, C.A.; et al. Cornelia de Lange syndrome is caused by mutations in NIPBL, the human homolog of Drosophila melanogaster Nipped-B. Nat. Genet. 2004, 36, 631–635. [Google Scholar] [CrossRef] [PubMed]
- Gillis, L.A.; McCallum, J.; Kaur, M.; DeScipio, C.; Yaeger, D.; Mariani, A.; Kline, A.D.; Li, H.-H.; Devoto, M.; Jackson, L.G.; et al. NIPBL Mutational Analysis in 120 Individuals with Cornelia de Lange Syndrome and Evaluation of Genotype-Phenotype Correlations. Am. J. Hum. Genet. 2004, 75, 610–623. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tonkin, E.; Wang, T.-J.; Lisgo, S.; Bamshad, M.J.; Strachan, T. NIPBL, encoding a homolog of fungal Scc2-type sister chromatid cohesion proteins and fly Nipped-B, is mutated in Cornelia de Lange syndrome. Nat. Genet. 2004, 36, 636–641. [Google Scholar] [CrossRef] [Green Version]
- Gregor, A.; Oti, M.; Kouwenhoven, E.N.; Hoyer, J.; Sticht, H.; Ekici, A.B.; Kjaergaard, S.; Rauch, A.; Stunnenberg, H.G.; Uebe, S.; et al. De Novo Mutations in the Genome Organizer CTCF Cause Intellectual Disability. Am. J. Hum. Genet. 2013, 93, 124–131. [Google Scholar] [CrossRef] [Green Version]
- Sanyal, A.; Lajoie, B.R.; Jain, G.; Dekker, J. The long-range interaction landscape of gene promoters. Nature 2012, 489, 109–113. [Google Scholar] [CrossRef]
- Jin, F.; Li, Y.; Dixon, J.R.; Selvaraj, S.; Ye, Z.; Lee, A.Y.; Yen, C.-A.; Schmitt, A.D.; Espinoza, C.A.; Ren, B. A high-resolution map of the three-dimensional chromatin interactome in human cells. Nature 2013, 503, 290–294. [Google Scholar] [CrossRef] [Green Version]
- Williamson, I.; Lettice, L.A.; Hill, R.E.; Bickmore, W.A. Shh and ZRS enhancer co-localisation is specific to the zone of polarizing activity. Development 2016, 143, 2994–3001. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Oudelaar, A.M.; Higgs, D.R. The relationship between genome structure and function. Nat. Rev. Genet. 2021, 22, 154–168. [Google Scholar] [CrossRef] [PubMed]
- Lieberman-Aiden, E.; Van Berkum, N.L.; Williams, L.; Imakaev, M.; Ragoczy, T.; Telling, A.; Amit, I.; Lajoie, B.R.; Sabo, P.J.; Dorschner, M.O.; et al. Comprehensive Mapping of Long-Range Interactions Reveals Folding Principles of the Human Genome. Science 2009, 326, 289–293. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dixon, J.R.; Selvaraj, S.; Yue, F.; Kim, A.; Li, Y.; Shen, Y.; Hu, M.; Liu, J.S.; Ren, B. Topological domains in mammalian genomes identified by analysis of chromatin interactions. Nature 2012, 485, 376–380. [Google Scholar] [CrossRef] [Green Version]
- Larochelle, S.; Batliner, J.; Gamble, M.J.; Barboza, N.M.; Kraybill, B.C.; Blethrow, J.D.; Shokat, K.M.; Fisher, R.P. Dichotomous but stringent substrate selection by the dual-function Cdk7 complex revealed by chemical genetics. Nat. Struct. Mol. Biol. 2005, 13, 55–62. [Google Scholar] [CrossRef]
- Sexton, T.; Yaffe, E.; Kenigsberg, E.; Bantignies, F.; Leblanc, B.; Hoichman, M.; Parrinello, H.; Tanay, A.; Cavalli, G. Three-Dimensional Folding and Functional Organization Principles of the Drosophila Genome. Cell 2012, 148, 458–472. [Google Scholar] [CrossRef] [Green Version]
- Symmons, O.; Pan, L.; Remeseiro, S.; Aktas, T.; Klein, F.; Huber, W.; Spitz, F. The Shh Topological Domain Facilitates the Action of Remote Enhancers by Reducing the Effects of Genomic Distances. Dev. Cell 2016, 39, 529–543. [Google Scholar] [CrossRef] [Green Version]
- Symmons, O.; Uslu, V.V.; Tsujimura, T.; Ruf, S.; Nassari, S.; Schwarzer, W.; Ettwiller, L.; Spitz, F. Functional and topological characteristics of mammalian regulatory domains. Genome Res. 2014, 24, 390–400. [Google Scholar] [CrossRef] [Green Version]
- De Laat, W.; Duboule, D. Topology of mammalian developmental enhancers and their regulatory landscapes. Nature 2013, 502, 499–506. [Google Scholar] [CrossRef]
- Kalhor, R.; Tjong, H.; Jayathilaka, N.; Alber, F.; Chen, L. Genome architectures revealed by tethered chromosome conformation capture and population-based modeling. Nat. Biotechnol. 2012, 30, 90–98. [Google Scholar] [CrossRef] [Green Version]
- Rao, S.S.P.; Huntley, M.H.; Durand, N.C.; Stamenova, E.K.; Bochkov, I.D.; Robinson, J.T.; Sanborn, A.L.; Machol, I.; Omer, A.D.; Lander, E.S.; et al. A 3D Map of the Human Genome at Kilobase Resolution Reveals Principles of Chromatin Looping. Cell 2014, 159, 1665–1680. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nasmyth, K. Disseminating the Genome: Joining, Resolving, and Separating Sister Chromatids During Mitosis and Meiosis. Annu. Rev. Genet. 2001, 35, 673–745. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alipour, E.; Marko, J.F. Self-organization of domain structures by DNA-loop-extruding enzymes. Nucleic Acids Res. 2012, 40, 11202–11212. [Google Scholar] [CrossRef] [Green Version]
- Ganji, M.; Shaltiel, I.A.; Bisht, S.; Kim, E.; Kalichava, A.; Haering, C.H.; Dekker, C. Real-time imaging of DNA loop extrusion by condensin. Science 2018, 360, 102–105. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sanborn, A.L.; Rao, S.S.P.; Huang, S.-C.; Durand, N.C.; Huntley, M.H.; Jewett, A.I.; Bochkov, I.; Chinnappan, D.; Cutkosky, A.; Li, J.; et al. Chromatin extrusion explains key features of loop and domain formation in wild-type and engineered genomes. Proc. Natl. Acad. Sci. USA 2015, 112, E6456–E6465. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fudenberg, G.; Imakaev, M.; Lu, C.; Goloborodko, A.; Abdennur, N.; Mirny, L.A. Formation of Chromosomal Domains by Loop Extrusion. Cell Rep. 2016, 15, 2038–2049. [Google Scholar] [CrossRef] [Green Version]
- Schwarzer, W.; Abdennur, N.; Goloborodko, A.; Pekowska, A.; Fudenberg, G.; Loe-Mie, Y.; Fonseca, N.; Huber, W.; Haering, C.; Mirny, L.; et al. Two independent modes of chromatin organization revealed by cohesin removal. Nature 2017, 551, 51–56. [Google Scholar] [CrossRef] [Green Version]
- Rao, S.S.; Huang, S.-C.; Hilaire, B.G.S.; Engreitz, J.M.; Perez, E.; Kieffer-Kwon, K.-R.; Sanborn, A.L.; Johnstone, S.E.; Bascom, G.D.; Bochkov, I.; et al. Cohesin Loss Eliminates All Loop Domains. Cell 2017, 171, 305–320.e24. [Google Scholar] [CrossRef] [Green Version]
- Wutz, G.; Várnai, C.; Nagasaka, K.; Cisneros, D.A.; Stocsits, R.R.; Tang, W.; Schoenfelder, S.; Jessberger, G.; Muhar, M.; Hossain, M.J.; et al. Topologically associating domains and chromatin loops depend on cohesin and are regulated by CTCF, WAPL, and PDS5 proteins. EMBO J. 2017, 36, 3573–3599. [Google Scholar] [CrossRef]
- Haarhuis, J.H.; van der Weide, R.H.; Blomen, V.A.; Yáñez-Cuna, J.O.; Amendola, M.; Van Ruiten, M.S.; Krijger, P.; Teunissen, H.; Medema, R.; Van Steensel, B.; et al. The Cohesin Release Factor WAPL Restricts Chromatin Loop Extension. Cell 2017, 169, 693–707.e14. [Google Scholar] [CrossRef] [Green Version]
- Nuebler, J.; Fudenberg, G.; Imakaev, M.; Abdennur, N.; Mirny, L.A. Chromatin organization by an interplay of loop extrusion and compartmental segregation. Proc. Natl. Acad. Sci. USA 2018, 115, E6697–E6706. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vian, L.; Pekowska, A.; Rao, S.S.; Kieffer-Kwon, K.-R.; Jung, S.; Baranello, L.; Huang, S.-C.; el Khattabi, L.; Dose, M.; Pruett, N.; et al. The Energetics and Physiological Impact of Cohesin Extrusion. Cell 2018, 173, 1165–1178.e20. [Google Scholar] [CrossRef] [Green Version]
- Kim, Y.; Shi, Z.; Zhang, H.; Finkelstein, I.J.; Yu, H. Human cohesin compacts DNA by loop extrusion. Science 2019, 366, 1345–1349. [Google Scholar] [CrossRef] [PubMed]
- Davidson, I.F.; Bauer, B.; Goetz, D.; Tang, W.; Wutz, G.; Peters, J.-M. DNA loop extrusion by human cohesin. Science 2019, 366, 1338–1345. [Google Scholar] [CrossRef]
- Gutierrez-Escribano, P.; Newton, M.D.; Llauró, A.; Huber, J.; Tanasie, L.; Davy, J.; Aly, I.; Aramayo, R.; Montoya, A.; Kramer, H.; et al. A conserved ATP- and Scc2/4-dependent activity for cohesin in tethering DNA molecules. Sci. Adv. 2019, 5, eaay6804. [Google Scholar] [CrossRef] [Green Version]
- De Wit, E.; Vos, E.; Holwerda, S.J.; Valdes-Quezada, C.; Verstegen, M.J.; Teunissen, H.; Splinter, E.; Wijchers, P.J.; Krijger, P.; de Laat, W. CTCF Binding Polarity Determines Chromatin Looping. Mol. Cell 2015, 60, 676–684. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, J.; Choi, J.-M.; Holehouse, A.S.; Lee, H.O.; Zhang, X.; Jahnel, M.; Maharana, S.; Lemaitre, R.; Pozniakovsky, A.; Drechsel, D.; et al. A Molecular Grammar Governing the Driving Forces for Phase Separation of Prion-like RNA Binding Proteins. Cell 2018, 174, 688–699.e16. [Google Scholar] [CrossRef] [Green Version]
- Li, P.; Banjade, S.; Cheng, H.-C.; Kim, S.; Chen, B.; Guo, L.; Llaguno, M.; Hollingsworth, J.V.; King, D.S.; Banani, S.F.; et al. Phase transitions in the assembly of multivalent signalling proteins. Nature 2012, 483, 336–340. [Google Scholar] [CrossRef]
- Feric, M.; Misteli, T. Phase separation in genome organization across evolution. Trends Cell Biol. 2021. [Google Scholar] [CrossRef] [PubMed]
- Schmidt, H.B.; Rohatgi, R. In vivo Formation of Vacuolated Multi-phase Compartments Lacking Membranes. Cell Rep. 2016, 16, 1228–1236. [Google Scholar] [CrossRef] [Green Version]
- Strom, A.R.; Emelyanov, A.V.; Mir, M.; Fyodorov, D.V.; Darzacq, X.; Karpen, G.H. Phase separation drives heterochromatin domain formation. Nature 2017, 547, 241–245. [Google Scholar] [CrossRef]
- Sabari, B.R.; Dall’Agnese, A.; Boija, A.; Klein, I.A.; Coffey, E.L.; Shrinivas, K.; Abraham, B.J.; Hannett, N.M.; Zamudio, A.V.; Manteiga, J.C.; et al. Coactivator condensation at super-enhancers links phase separation and gene control. Science 2018, 361, eaar3958. [Google Scholar] [CrossRef] [Green Version]
- Cho, W.-K.; Spille, J.-H.; Hecht, M.; Lee, C.; Li, C.; Grube, V.; Cisse, I.I. Mediator and RNA polymerase II clusters associate in transcription-dependent condensates. Science 2018, 361, 412–415. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Boija, A.; Klein, I.A.; Sabari, B.R.; Dall’Agnese, A.; Coffey, E.L.; Zamudio, A.V.; Li, C.H.; Shrinivas, K.; Manteiga, J.C.; Hannett, N.M.; et al. Transcription Factors Activate Genes through the Phase-Separation Capacity of Their Activation Domains. Cell 2018, 175, 1842–1855.e16. [Google Scholar] [CrossRef] [Green Version]
- Lu, Y.; Wu, T.; Gutman, O.; Lu, H.; Zhou, Q.; Henis, Y.I.; Luo, K. Phase separation of TAZ compartmentalizes the transcription machinery to promote gene expression. Nat. Cell Biol. 2020, 22, 453–464. [Google Scholar] [CrossRef]
- Nair, S.J.; Yang, L.; Meluzzi, D.; Oh, S.; Yang, F.; Friedman, M.J.; Wang, S.; Suter, T.; Alshareedah, I.; Gamliel, A.; et al. Phase separation of ligand-activated enhancers licenses cooperative chromosomal enhancer assembly. Nat. Struct. Mol. Biol. 2019, 26, 193–203. [Google Scholar] [CrossRef] [PubMed]
- Henninger, J.E.; Oksuz, O.; Shrinivas, K.; Sagi, I.; LeRoy, G.; Zheng, M.M.; Andrews, J.O.; Zamudio, A.V.; Lazaris, C.; Hannett, N.M.; et al. RNA-Mediated Feedback Control of Transcriptional Condensates. Cell 2021, 184, 207–225.e24. [Google Scholar] [CrossRef] [PubMed]
- Bailey, A.; Le Couteur, A.; Gottesman, I.; Bolton, P.; Simonoff, E.; Yuzda, E.; Rutter, M.L. Autism as a strongly genetic disorder: Evidence from a British twin study. Psychol. Med. 1995, 25, 63–77. [Google Scholar] [CrossRef]
- Hallmayer, J.; Cleveland, S.; Torres, A.; Phillips, J.; Cohen, B.; Torigoe, T.; Miller, J.; Fedele, A.; Collins, J.; Smith, K.; et al. Genetic Heritability and Shared Environmental Factors Among Twin Pairs With Autism. Arch. Gen. Psychiatry 2011, 68, 1095–1102. [Google Scholar] [CrossRef]
- The Brainstorm Consortium; Anttila, V.; Bulik-Sullivan, B.; Finucane, H.K.; Walters, R.K.; Bras, J.; Duncan, L.; Escott-Price, V.; Falcone, G.J.; Gormley, P.; et al. Analysis of shared heritability in common disorders of the brain. Science 2018, 360, eaap8757. [Google Scholar] [CrossRef] [Green Version]
- Iossifov, I.; O’Roak, B.J.; Sanders, S.J.; Ronemus, M.; Krumm, N.; Levy, D.; Stessman, H.A.; Witherspoon, K.T.; Vives, L.; Patterson, K.E.; et al. The contribution of de novo coding mutations to autism spectrum disorder. Nature 2014, 515, 216–221. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, P.H.; Anttila, V.; Won, H.; Feng, Y.-C.A.; Rosenthal, J.; Zhu, Z.; Tucker-Drob, E.M.; Nivard, M.; Grotzinger, A.D.; Posthuma, D.; et al. Genomic Relationships, Novel Loci, and Pleiotropic Mechanisms across Eight Psychiatric Disorders. Cell 2019, 179, 1469–1482.e11. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, T.I.; Young, R.A. Transcriptional Regulation and Its Misregulation in Disease. Cell 2013, 152, 1237–1251. [Google Scholar] [CrossRef] [Green Version]
- De Rubeis, S.; He, X.; Goldberg, A.P.; Poultney, C.S.; Samocha, K.; Cicek, A.E.; Kou, Y.; Liu, L.; Fromer, M.; Walker, S.; et al. Synaptic, transcriptional and chromatin genes disrupted in autism. Nature 2014, 515, 209–215. [Google Scholar] [CrossRef] [PubMed]
- Fahrner, J.A.; Bjornsson, H.T. Mendelian Disorders of the Epigenetic Machinery: Tipping the Balance of Chromatin States. Annu. Rev. Genom. Hum. Genet. 2014, 15, 269–293. [Google Scholar] [CrossRef] [Green Version]
- Kleefstra, T.; Schenck, A.; Kramer, J.; van Bokhoven, H. The genetics of cognitive epigenetics. Neuropharmacology 2014, 80, 83–94. [Google Scholar] [CrossRef]
- Garcia-Dominguez, M.; Reyes, J.C. SUMO association with repressor complexes, emerging routes for transcriptional control. Biochim. Biophys. Acta (BBA) Bioenerg. 2009, 1789, 451–459. [Google Scholar] [CrossRef]
- Vitriolo, A.; Gabriele, M.; Testa, G. From enhanceropathies to the epigenetic manifold underlying human cognition. Hum. Mol. Genet. 2019, 28, R226–R234. [Google Scholar] [CrossRef]
- Pinto, D.; Delaby, E.; Merico, D.; Barbosa, M.; Merikangas, A.; Klei, L.; Thiruvahindrapuram, B.; Xu, X.; Ziman, R.; Wang, Z.; et al. Convergence of Genes and Cellular Pathways Dysregulated in Autism Spectrum Disorders. Am. J. Hum. Genet. 2014, 94, 677–694. [Google Scholar] [CrossRef] [Green Version]
- Gabriele, M.; Tobon, A.L.; D’Agostino, G.A.; Testa, G. The chromatin basis of neurodevelopmental disorders: Rethinking dysfunction along the molecular and temporal axes. Prog. Neuro-Psychopharmacol. Biol. Psychiatry 2018, 84, 306–327. [Google Scholar] [CrossRef]
- Pohodich, A.E.; Zoghbi, H.Y. Rett syndrome: Disruption of epigenetic control of postnatal neurological functions. Hum. Mol. Genet. 2015, 24, R10–R16. [Google Scholar] [CrossRef]
- Berson, A.; Sartoris, A.; Nativio, R.; Van Deerlin, V.; Toledo, J.; Porta, S.; Liu, S.; Chung, C.-Y.; Garcia, B.A.; Lee, V.M.-Y.; et al. TDP-43 Promotes Neurodegeneration by Impairing Chromatin Remodeling. Curr. Biol. 2017, 27, 3579–3590.e6. [Google Scholar] [CrossRef] [Green Version]
- Ernst, C. Proliferation and Differentiation Deficits are a Major Convergence Point for Neurodevelopmental Disorders. Trends Neurosci. 2016, 39, 290–299. [Google Scholar] [CrossRef] [PubMed]
- Thienpont, B.; Béna, F.; Breckpot, J.; Philip, N.; Menten, B.; Van Esch, H.; Scalais, E.; Salamone, J.M.; Fong, C.-T.; Kussmann, J.L.; et al. Duplications of the critical Rubinstein-Taybi deletion region on chromosome 16p13.3 cause a novel recognisable syndrome. J. Med. Genet. 2009, 47, 155–161. [Google Scholar] [CrossRef] [Green Version]
- Marangi, G.; Leuzzi, V.; Orteschi, D.; Grimaldi, M.E.; Lecce, R.; Neri, G.; Zollino, M. Duplication of the Rubinstein-Taybi region on 16p13.3 is associated with a distinctive phenotype. Am. J. Med. Genet. 2008, 146, 2313–2317. [Google Scholar] [CrossRef] [PubMed]
- Bahari-Javan, S.; Varbanov, H.; Halder, R.; Benito, E.; Kaurani, L.; Burkhardt, S.; Anderson-Schmidt, H.; Anghelescu, I.; Budde, M.; Stilling, R.; et al. HDAC1 links early life stress to schizophrenia-like phenotypes. Proc. Natl. Acad. Sci. USA 2017, 114, E4686–E4694. [Google Scholar] [CrossRef] [Green Version]
- Benes, F.M.; Lim, B.; Matzilevich, D.; Walsh, J.P.; Subburaju, S.; Minns, M. Regulation of the GABA cell phenotype in hippocampus of schizophrenics and bipolars. Proc. Natl. Acad. Sci. USA 2007, 104, 10164–10169. [Google Scholar] [CrossRef] [Green Version]
- Sharma, R.P.; Grayson, D.R.; Gavin, D.P. Histone deactylase 1 expression is increased in the prefrontal cortex of schizophrenia subjects: Analysis of the National Brain Databank microarray collection. Schizophr. Res. 2008, 98, 111–117. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stefanis, L. α-Synuclein in Parkinson’s Disease. Cold Spring Harb. Perspect. Med. 2011, 2, a009399. [Google Scholar] [CrossRef] [Green Version]
- Desplats, P.; Spencer, B.; Coffee, E.; Patel, P.; Michael, S.; Patrick, C.; Adame, A.; Rockenstein, E.; Masliah, E. α-Synuclein Sequesters Dnmt1 from the Nucleus. J. Biol. Chem. 2011, 286, 9031–9037. [Google Scholar] [CrossRef] [Green Version]
- Toker, L.; Tran, G.T.; Sundaresan, J.; Tysnes, O.-B.; Alves, G.; Haugarvoll, K.; Nido, G.S.; Dölle, C.; Tzoulis, C. Genome-wide histone acetylation analysis reveals altered transcriptional regulation in the Parkinson’s disease brain. Mol. Neurodegener. 2021, 16, 1–20. [Google Scholar] [CrossRef]
- Corpión, I.H.; Guiretti, D.; Alcaraz-Iborra, M.; Olivares, R.; Campos-Caro, A.; Barco, A.; Valor, L.M. Early alteration of epigenetic-related transcription in Huntington’s disease mouse models. Sci. Rep. 2018, 8, 1–14. [Google Scholar] [CrossRef]
- Steffan, J.S.; Kazantsev, A.; Spasic-Boskovic, O.; Greenwald, M.; Zhu, Y.-Z.; Gohler, H.; Wanker, E.; Bates, G.; Housman, D.E.; Thompson, L.M. The Huntington’s disease protein interacts with p53 and CREB-binding protein and represses transcription. Proc. Natl. Acad. Sci. USA 2000, 97, 6763–6768. [Google Scholar] [CrossRef] [Green Version]
- Ng, C.W.; Yildirim, F.; Yap, Y.S.; Dalin, S.; Matthews, B.J.; Velez, P.J.; Labadorf, A.; Housman, D.E.; Fraenkel, E. Extensive changes in DNA methylation are associated with expression of mutant huntingtin. Proc. Natl. Acad. Sci. USA 2013, 110, 2354–2359. [Google Scholar] [CrossRef] [Green Version]
- West, R.L.; Lee, J.M.; Maroun, L.E. Hypomethylation of the amyloid precursor protein gene in the brain of an alzheimer’s disease patient. J. Mol. Neurosci. 1995, 6, 141–146. [Google Scholar] [CrossRef]
- Bakulski, K.M.; Dolinoy, D.C.; Sartor, M.A.; Paulson, H.L.; Konen, J.R.; Lieberman, A.P.; Albin, R.L.; Hu, H.; Rozek, L.S. Genome-Wide DNA Methylation Differences Between Late-Onset Alzheimer’s Disease and Cognitively Normal Controls in Human Frontal Cortex. J. Alzheimer’s Dis. 2012, 29, 571–588. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Klein, H.-U.; McCabe, C.; Gjoneska, E.; Sullivan, S.E.; Kaskow, B.; Tang, A.; Smith, R.V.; Xu, J.; Pfenning, A.R.; Bernstein, B.E.; et al. Epigenome-wide study uncovers large-scale changes in histone acetylation driven by tau pathology in aging and Alzheimer’s human brains. Nat. Neurosci. 2019, 22, 37–46. [Google Scholar] [CrossRef]
- Marzi, S.; Leung, S.K.; Ribarska, T.; Hannon, E.; Smith, A.R.; Pishva, E.; Poschmann, J.; Moore, K.; Troakes, C.; Al-Sarraj, S.; et al. A histone acetylome-wide association study of Alzheimer’s disease identifies disease-associated H3K27ac differences in the entorhinal cortex. Nat. Neurosci. 2018, 21, 1618–1627. [Google Scholar] [CrossRef] [Green Version]
- Rao, J.S.; Keleshian, V.L.; Klein, S.; Rapoport, S.I. Epigenetic modifications in frontal cortex from Alzheimer’s disease and bipolar disorder patients. Transl. Psychiatry 2012, 2, e132. [Google Scholar] [CrossRef]
- Zhang, K.; Schrag, M.; Crofton, A.; Trivedi, R.; Vinters, H.; Kirsch, W. Targeted proteomics for quantification of histone acetylation in Alzheimer’s disease. Proteomics 2012, 12, 1261–1268. [Google Scholar] [CrossRef]
- Li, P.; Marshall, L.; Oh, G.; Jakubowski, J.L.; Groot, D.; He, Y.; Wang, T.; Petronis, A.; Labrie, V. Epigenetic dysregulation of enhancers in neurons is associated with Alzheimer’s disease pathology and cognitive symptoms. Nat. Commun. 2019, 10, 2246. [Google Scholar] [CrossRef] [Green Version]
- Furlong, E.E.M.; Levine, M. Developmental enhancers and chromosome topology. Science 2018, 361, 1341–1345. [Google Scholar] [CrossRef] [Green Version]
- Lupiáñez, D.G.; Kraft, K.; Heinrich, V.; Krawitz, P.; Brancati, F.; Klopocki, E.; Horn, D.; Kayserili, H.; Opitz, J.M.; Laxova, R.; et al. Disruptions of Topological Chromatin Domains Cause Pathogenic Rewiring of Gene-Enhancer Interactions. Cell 2015, 161, 1012–1025. [Google Scholar] [CrossRef] [Green Version]
- Tang, Z.; Luo, O.J.; Li, X.; Zheng, M.; Zhu, J.J.; Szalaj, P.; Trzaskoma, P.; Magalska, A.; Wlodarczyk, J.; Ruszczycki, B.; et al. CTCF-Mediated Human 3D Genome Architecture Reveals Chromatin Topology for Transcription. Cell 2015, 163, 1611–1627. [Google Scholar] [CrossRef] [Green Version]
- Hyle, J.; Zhang, Y.; Wright, S.; Xu, B.; Shao, Y.; Easton, J.; Tian, L.; Feng, R.; Xu, P.; Li, C. Acute depletion of CTCF directly affects MYC regulation through loss of enhancer–promoter looping. Nucleic Acids Res. 2019, 47, 6699–6713. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schuijers, J.; Manteiga, J.C.; Weintraub, A.S.; Day, D.S.; Zamudio, A.V.; Hnisz, D.; Lee, T.I.; Young, R.A. Transcriptional Dysregulation of MYC Reveals Common Enhancer-Docking Mechanism. Cell Rep. 2018, 23, 349–360. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ushiki, A.; University of Washington Center for Mendelian Genomics; Zhang, Y.; Xiong, C.; Zhao, J.; Georgakopoulos-Soares, I.; Kane, L.; Jamieson, K.; Bamshad, M.J.; Nickerson, D.A.; et al. Deletion of CTCF sites in the SHH locus alters enhancer–promoter interactions and leads to acheiropodia. Nat. Commun. 2021, 12, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Hirayama, T.; Tarusawa, E.; Yoshimura, Y.; Galjart, N.; Yagi, T. CTCF Is Required for Neural Development and Stochastic Expression of Clustered Pcdh Genes in Neurons. Cell Rep. 2012, 2, 345–357. [Google Scholar] [CrossRef] [Green Version]
- Sams, D.S.; Nardone, S.; Getselter, D.; Raz, D.; Tal, M.; Rayi, P.R.; Kaphzan, H.; Hakim, O.; Elliott, E. Neuronal CTCF Is Necessary for Basal and Experience-Dependent Gene Regulation, Memory Formation, and Genomic Structure of BDNF and Arc. Cell Rep. 2016, 17, 2418–2430. [Google Scholar] [CrossRef] [Green Version]
- Yamada, T.; Yang, Y.; Valnegri, P.; Juric, I.; Abnousi, A.; Markwalter, K.H.; Guthrie, A.N.; Godec, A.; Oldenborg, A.; Hu, M.; et al. Sensory experience remodels genome architecture in neural circuit to drive motor learning. Nature 2019, 569, 708–713. [Google Scholar] [CrossRef]
- Beagan, J.A.; Pastuzyn, E.D.; Fernandez, L.R.; Guo, M.H.; Feng, K.; Titus, K.; Chandrashekar, H.; Shepherd, J.D.; Phillips-Cremins, J.E. Three-dimensional genome restructuring across timescales of activity-induced neuronal gene expression. Nat. Neurosci. 2020, 23, 707–717. [Google Scholar] [CrossRef]
- Sofueva, S.; Yaffe, E.; Chan, W.-C.; Georgopoulou, D.; Rudan, M.V.; Mira-Bontenbal, H.; Pollard, S.; Schroth, G.P.; Tanay, A.; Hadjur, S. Cohesin-mediated interactions organize chromosomal domain architecture. EMBO J. 2013, 32, 3119–3129. [Google Scholar] [CrossRef] [Green Version]
- Seitan, V.; Faure, A.; Zhan, Y.; McCord, R.; Lajoie, B.R.; Ing-Simmons, E.; Lenhard, B.; Giorgetti, L.; Heard, E.; Fisher, A.G.; et al. Cohesin-based chromatin interactions enable regulated gene expression within preexisting architectural compartments. Genome Res. 2013, 23, 2066–2077. [Google Scholar] [CrossRef] [Green Version]
- Nora, E.P.; Goloborodko, A.; Valton, A.-L.; Gibcus, J.H.; Uebersohn, A.; Abdennur, N.; Dekker, J.; Mirny, L.A.; Bruneau, B.G. Targeted Degradation of CTCF Decouples Local Insulation of Chromosome Domains from Genomic Compartmentalization. Cell 2017, 169, 930–944.e22. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wendt, K.S.; Yoshida, K.; Itoh, T.; Bando, M.; Koch, B.; Schirghuber, E.; Tsutsumi, S.; Nagae, G.; Ishihara, K.; Mishiro, T.; et al. Cohesin mediates transcriptional insulation by CCCTC-binding factor. Nature 2008, 451, 796–801. [Google Scholar] [CrossRef] [PubMed]
- Zuin, J.; Dixon, J.R.; van der Reijden, M.I.J.A.; Ye, Z.; Kolovos, P.; Brouwer, R.W.W.; van de Corput, M.P.C.; van de Werken, H.J.G.; Knoch, T.A.; van Ijcken, W.F.J.; et al. Cohesin and CTCF differentially affect chromatin architecture and gene expression in human cells. Proc. Natl. Acad. Sci. USA 2014, 111, 996–1001. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Franke, M.; Ibrahim, D.M.; Andrey, G.; Schwarzer, W.; Heinrich, V.; Schöpflin, R.; Kraft, K.; Kempfer, R.; Jerković, I.; Chan, W.-L.; et al. Formation of new chromatin domains determines pathogenicity of genomic duplications. Nature 2016, 538, 265–269. [Google Scholar] [CrossRef] [PubMed]
- Rajarajan, P.; Borrman, T.; Liao, W.; Schrode, N.; Flaherty, E.; Casiño, C.; Powell, S.; Yashaswini, C.; Lamarca, E.A.; Kassim, B.; et al. Neuron-specific signatures in the chromosomal connectome associated with schizophrenia risk. Science 2018, 362, eaat4311. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lu, L.; Liu, X.; Huang, W.-K.; Giusti-Rodríguez, P.; Cui, J.; Zhang, S.; Xu, W.; Wen, Z.; Ma, S.; Rosen, J.D.; et al. Robust Hi-C Maps of Enhancer-Promoter Interactions Reveal the Function of Non-coding Genome in Neural Development and Diseases. Mol. Cell 2020, 79, 521–534.e15. [Google Scholar] [CrossRef]
- Gallagher, M.D.; Posavi, M.; Huang, P.; Unger, T.L.; Berlyand, Y.; Gruenewald, A.L.; Chesi, A.; Manduchi, E.; Wells, A.D.; Grant, S.F.; et al. A Dementia-Associated Risk Variant near TMEM106B Alters Chromatin Architecture and Gene Expression. Am. J. Hum. Genet. 2017, 101, 643–663. [Google Scholar] [CrossRef] [Green Version]
- Hillary, R.F.; McCartney, D.L.; Harris, S.E.; Stevenson, A.; Seeboth, A.; Zhang, Q.; Liewald, D.C.; Evans, K.L.; Ritchie, C.W.; Tucker-Drob, E.M.; et al. Genome and epigenome wide studies of neurological protein biomarkers in the Lothian Birth Cohort 1936. Nat. Commun. 2019, 10, 1–9. [Google Scholar] [CrossRef] [Green Version]
- Maurano, M.T.; Humbert, R.; Rynes, E.; Thurman, R.E.; Haugen, E.; Wang, H.; Reynolds, A.P.; Sandstrom, R.; Qu, H.; Brody, J.; et al. Systematic Localization of Common Disease-Associated Variation in Regulatory DNA. Science 2012, 337, 1190–1195. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Maurano, M.T.; Haugen, E.; Sandstrom, R.; Vierstra, J.; Shafer, A.; Kaul, R.; Stamatoyannopoulos, J.A. Large-scale identification of sequence variants influencing human transcription factor occupancy in vivo. Nat. Genet. 2015, 47, 1393–1401. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ernst, J.; Kheradpour, P.; Mikkelsen, T.S.; Shoresh, N.; Ward, L.; Epstein, C.B.; Zhang, X.; Wang, L.; Issner, R.; Coyne, M.; et al. Mapping and analysis of chromatin state dynamics in nine human cell types. Nature 2011, 473, 43–49. [Google Scholar] [CrossRef] [PubMed]
- Chang, D.; Nalls, M.A.; Hallgrímsdóttir, I.B.; Hunkapiller, J.; Van Der Brug, M.; Cai, F.; Kerchner, G.A.; Ayalon, G.; Bingol, B.; Sheng, M.; et al. A meta-analysis of genome-wide association studies identifies 17 new Parkinson’s disease risk loci. Nat. Genet. 2017, 49, 1511–1516. [Google Scholar] [CrossRef] [PubMed]
- Jansen, I.E.; Savage, J.E.; Watanabe, K.; Bryois, J.; Williams, D.; Steinberg, S.; Sealock, J.; Karlsson, I.; Hägg, S.; Athanasiu, L.; et al. Genome-wide meta-analysis identifies new loci and functional pathways influencing Alzheimer’s disease risk. Nat. Genet. 2019, 51, 404–413. [Google Scholar] [CrossRef]
- Hannon, E.; Marzi, S.J.; Schalkwyk, L.S.; Mill, J. Genetic risk variants for brain disorders are enriched in cortical H3K27ac domains. Mol. Brain 2019, 12, 1–6. [Google Scholar] [CrossRef]
- Adolfsson, R. Biological insights from 108 schizophrenia-associated genetic loci. Nature 2014, 511, 421–427. [Google Scholar] [CrossRef] [Green Version]
- Gallagher, M.D.; Chen-Plotkin, A.S. The Post-GWAS Era: From Association to Function. Am. J. Hum. Genet. 2018, 102, 717–730. [Google Scholar] [CrossRef] [Green Version]
- Ki Sung GTEx Consortium Genetic effects on gene expression across human tissues. Nature 2017, 550, 204–213. [CrossRef] [PubMed]
- Kikuchi, M.; Hara, N.; Hasegawa, M.; Miyashita, A.; Kuwano, R.; Ikeuchi, T.; Nakaya, A. Enhancer variants associated with Alzheimer’s disease affect gene expression via chromatin looping. BMC Med. Genom. 2019, 12, 128. [Google Scholar] [CrossRef] [Green Version]
- Vermunt, M.W.; Reinink, P.; Korving, J.; De Bruijn, E.; Creyghton, P.M.; Basak, O.; Geeven, G.; Toonen, P.W.; Lansu, N.; Meunier, C.; et al. Large-Scale Identification of Coregulated Enhancer Networks in the Adult Human Brain. Cell Rep. 2014, 9, 767–779. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Soldner, F.; Stelzer, Y.; Shivalila, C.S.; Abraham, B.; Latourelle, J.C.; Barrasa, M.I.; Goldmann, J.; Myers, R.H.; Young, R.A.; Jaenisch, R. Parkinson-associated risk variant in distal enhancer of α-synuclein modulates target gene expression. Nature 2016, 533, 95–99. [Google Scholar] [CrossRef] [Green Version]
- Fromer, M.; Roussos, P.; Sieberts, S.K.; Johnson, J.; Kavanagh, D.H.; Perumal, T.M.; Ruderfer, D.M.; Oh, E.C.; Topol, A.; Shah, H.R.; et al. Gene expression elucidates functional impact of polygenic risk for schizophrenia. Nat. Neurosci. 2016, 19, 1442–1453. [Google Scholar] [CrossRef] [Green Version]
- Huo, Y.; Li, S.; Liu, J.; Li, X.; Luo, X.-J. Functional genomics reveal gene regulatory mechanisms underlying schizophrenia risk. Nat. Commun. 2019, 10, 1–19. [Google Scholar] [CrossRef] [PubMed]
- Bryois, J.; Garrett, M.E.; Song, L.; Safi, A.; Giusti-Rodriguez, P.; Johnson, G.D.; Shieh, A.W.; Buil, A.; Fullard, J.F.; Roussos, P.; et al. Evaluation of chromatin accessibility in prefrontal cortex of individuals with schizophrenia. Nat. Commun. 2018, 9, 1–15. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Ma, C.; Li, W.; Yang, Y.; Li, X.; Liu, J.; Wang, J.; Li, S.; Liu, Y.; Li, K.; et al. A missense variant in NDUFA6 confers schizophrenia risk by affecting YY1 binding and NAGA expression. Mol. Psychiatry 2021, 1–16. [Google Scholar] [CrossRef]
- Kumasaka, N.; Knights, A.J.; Gaffney, D.J. High-resolution genetic mapping of putative causal interactions between regions of open chromatin. Nat. Genet. 2019, 51, 128–137. [Google Scholar] [CrossRef]
- Cano-Gamez, E.; Trynka, G. From GWAS to Function: Using Functional Genomics to Identify the Mechanisms Underlying Complex Diseases. Front. Genet. 2020, 11, 424. [Google Scholar] [CrossRef]
- Gusev, A.; Ko, A.; Shi, H.; Bhatia, G.; Chung, W.; Penninx, B.W.J.H.; Jansen, R.; De Geus, E.J.C.; Boomsma, D.I.; Wright, F.A.; et al. Integrative approaches for large-scale transcriptome-wide association studies. Nat. Genet. 2016, 48, 245–252. [Google Scholar] [CrossRef] [Green Version]
- Zhu, Z.; Zhang, F.; Hu, H.; Bakshi, A.; Robinson, M.R.; Powell, J.E.; Montgomery, G.W.; Goddard, M.E.; Wray, N.R.; Visscher, P.M.; et al. Integration of summary data from GWAS and eQTL studies predicts complex trait gene targets. Nat. Genet. 2016, 48, 481–487. [Google Scholar] [CrossRef] [PubMed]
- Forrest, M.; Zhang, H.; Moy, W.; McGowan, H.; Leites, C.; Dionisio, L.; Xu, Z.; Shi, J.; Sanders, A.R.; Greenleaf, W.J.; et al. Open Chromatin Profiling in hiPSC-Derived Neurons Prioritizes Functional Noncoding Psychiatric Risk Variants and Highlights Neurodevelopmental Loci. Cell Stem Cell 2017, 21, 305–318.e8. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Melnikov, A.; Murugan, A.; Zhang, X.; Tesileanu, T.; Wang, L.; Rogov, P.; Feizi, S.; Gnirke, A.; Jr, C.G.C.; Kinney, J.; et al. Systematic dissection and optimization of inducible enhancers in human cells using a massively parallel reporter assay. Nat. Biotechnol. 2012, 30, 271–277. [Google Scholar] [CrossRef] [Green Version]
- Van Arensbergen, J.; Pagie, L.; Fitzpatrick, V.D.; De Haas, M.; Baltissen, M.P.; Comoglio, F.; Van Der Weide, R.H.; Teunissen, H.; Võsa, U.; Franke, L.; et al. High-throughput identification of human SNPs affecting regulatory element activity. Nat. Genet. 2019, 51, 1160–1169. [Google Scholar] [CrossRef] [PubMed]
- Tewhey, R.; Kotliar, D.; Park, D.; Liu, B.; Winnicki, S.; Reilly, S.; Andersen, K.G.; Mikkelsen, T.S.; Lander, E.S.; Schaffner, S.F.; et al. Direct Identification of Hundreds of Expression-Modulating Variants using a Multiplexed Reporter Assay. Cell 2016, 165, 1519–1529. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Noordermeer, D.; de Wit, E.; Klous, P.; Van De Werken, H.; Simonis, M.; Lopez-Jones, M.; Eussen, B.; De Klein, A.; Singer, R.H.; de Laat, W. Variegated gene expression caused by cell-specific long-range DNA interactions. Nat. Cell Biol. 2011, 13, 944–951. [Google Scholar] [CrossRef] [Green Version]
- Vernimmen, D.; De Gobbi, M.; Sloane-Stanley, J.A.; Wood, W.G.; Higgs, D.R. Long-range chromosomal interactions regulate the timing of the transition between poised and active gene expression. EMBO J. 2007, 26, 2041–2051. [Google Scholar] [CrossRef]
- Simonis, M.; Klous, P.; Splinter, E.; Moshkin, Y.; Willemsen, R.; de Wit, E.; Van Steensel, B.; de Laat, W. Nuclear organization of active and inactive chromatin domains uncovered by chromosome conformation capture–on-chip (4C). Nat. Genet. 2006, 38, 1348–1354. [Google Scholar] [CrossRef]
- Baù, D.; Sanyal, A.; Lajoie, B.R.; Capriotti, E.; Byron, M.; Lawrence, J.B.; Dekker, J.; Marti-Renom, M.A. The three-dimensional folding of the α-globin gene domain reveals formation of chromatin globules. Nat. Struct. Mol. Biol. 2010, 18, 107–114. [Google Scholar] [CrossRef] [Green Version]
- Palstra, R.-J.; Tolhuis, B.; Splinter, E.; Nijmeijer, R.; Grosveld, F.; de Laat, W. The β-globin nuclear compartment in development and erythroid differentiation. Nat. Genet. 2003, 35, 190–194. [Google Scholar] [CrossRef]
- Nott, A.; Holtman, I.R.; Coufal, N.G.; Schlachetzki, J.C.M.; Yu, M.; Hu, R.; Han, C.Z.; Pena, M.; Xiao, J.; Wu, Y.; et al. Brain cell type–specific enhancer–promoter interactome maps and disease-risk association. Science 2019, 366, 1134–1139. [Google Scholar] [CrossRef] [PubMed]
- Won, H.; De La Torre-Ubieta, L.; Stein, J.; Parikshak, N.; Huang, J.; Opland, C.K.; Gandal, M.; Sutton, G.J.; Hormozdiari, F.; Lu, D.; et al. Chromosome conformation elucidates regulatory relationships in developing human brain. Nature 2016, 538, 523–527. [Google Scholar] [CrossRef] [Green Version]
- Hauberg, M.E.; The CommonMind Consortium; Fullard, J.F.; Zhu, L.; Cohain, A.T.; Giambartolomei, C.; Misir, R.; Reach, S.; Johnson, J.; Wang, M.; et al. Differential activity of transcribed enhancers in the prefrontal cortex of 537 cases with schizophrenia and controls. Mol. Psychiatry 2019, 24, 1685–1695. [Google Scholar] [CrossRef] [PubMed]
- Ruderfer, D.M.; Ripke, S.; McQuillin, A.; Boocock, J.; Stahl, E.A.; Pavlides, J.M.W.; Mullins, N.; Charney, A.W.; Ori, A.P.; Loohuis, L.M.O.; et al. Genomic Dissection of Bipolar Disorder and Schizophrenia, Including 28 Subphenotypes. Cell 2018, 173, 1705–1715.e16. [Google Scholar] [CrossRef] [Green Version]
- Parikshak, N.; Luo, R.; Zhang, A.; Won, H.; Lowe, J.K.; Chandran, V.; Horvath, S.; Geschwind, D.H. Integrative Functional Genomic Analyses Implicate Specific Molecular Pathways and Circuits in Autism. Cell 2013, 155, 1008–1021. [Google Scholar] [CrossRef] [Green Version]
- Sullivan, S.E.; Liao, M.; Smith, R.V.; White, C.; Lagomarsino, V.N.; Xu, J.; Taga, M.; Bennett, D.A.; De Jager, P.L.; Young-Pearse, T. Candidate-based screening via gene modulation in human neurons and astrocytes implicates FERMT2 in Aβ and TAU proteostasis. Hum. Mol. Genet. 2018, 28, 718–735. [Google Scholar] [CrossRef]
- Delaneau, O.; Zazhytska, M.; Borel, C.; Giannuzzi, G.; Rey, G.; Howald, C.; Kumar, S.; Ongen, H.; Popadin, K.; Marbach, D.; et al. Chromatin three-dimensional interactions mediate genetic effects on gene expression. Science 2019, 364, eaat8266. [Google Scholar] [CrossRef] [PubMed]
- Cusanovich, D.; Daza, R.; Adey, A.; Pliner, H.; Christiansen, L.; Gunderson, K.L.; Steemers, F.J.; Trapnell, C.; Shendure, J. Multiplex single-cell profiling of chromatin accessibility by combinatorial cellular indexing. Science 2015, 348, 910–914. [Google Scholar] [CrossRef] [Green Version]
- Armand, E.J.; Li, J.; Xie, F.; Luo, C.; Mukamel, E.A. Single-Cell Sequencing of Brain Cell Transcriptomes and Epigenomes. Neuron 2021, 109, 11–26. [Google Scholar] [CrossRef] [PubMed]
- Zeng, H.; Sanes, J.R. Neuronal cell-type classification: Challenges, opportunities and the path forward. Nat. Rev. Neurosci. 2017, 18, 530–546. [Google Scholar] [CrossRef] [PubMed]
- Lake, B.B.; Ai, R.; Kaeser, G.E.; Salathia, N.S.; Yung, Y.C.; Liu, R.; Wildberg, A.; Gao, D.; Fung, H.-L.; Chen, S.; et al. Neuronal subtypes and diversity revealed by single-nucleus RNA sequencing of the human brain. Science 2016, 352, 1586–1590. [Google Scholar] [CrossRef] [Green Version]
- Lake, B.B.; Chen, S.; Sos, B.C.; Fan, J.; Kaeser, G.E.; Yung, Y.C.; Duong, T.E.; Gao, D.; Chun, J.; Kharchenko, P.V.; et al. Integrative single-cell analysis of transcriptional and epigenetic states in the human adult brain. Nat. Biotechnol. 2018, 36, 70–80. [Google Scholar] [CrossRef]
- Darmanis, S.; Sloan, S.A.; Zhang, Y.; Enge, M.; Caneda, C.; Shuer, L.M.; Gephart, M.G.H.; Barres, B.A.; Quake, S.R. A survey of human brain transcriptome diversity at the single cell level. Proc. Natl. Acad. Sci. USA 2015, 112, 7285–7290. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Romanov, R.; Zeisel, A.; Bakker, J.; Girach, F.; Hellysaz, A.; Tomer, R.; Alpár, A.; Mulder, J.; Clotman, F.; Keimpema, E.; et al. Molecular interrogation of hypothalamic organization reveals distinct dopamine neuronal subtypes. Nat. Neurosci. 2017, 20, 176–188. [Google Scholar] [CrossRef] [PubMed]
- Zeisel, A.; Muñoz-Manchado, A.B.; Codeluppi, S.; Lönnerberg, P.; La Manno, G.; Juréus, A.; Marques, S.; Munguba, H.; He, L.; Betsholtz, C.; et al. Cell types in the mouse cortex and hippocampus revealed by single-cell RNA-seq. Science 2015, 347, 1138–1142. [Google Scholar] [CrossRef] [PubMed]
- Tasic, B.; Yao, Z.; Graybuck, L.T.; Smith, K.A.; Nguyen, T.N.; Bertagnolli, D.; Goldy, J.; Garren, E.; Economo, M.N.; Viswanathan, S.; et al. Shared and distinct transcriptomic cell types across neocortical areas. Nat. Cell Biol. 2018, 563, 72–78. [Google Scholar] [CrossRef] [PubMed]
- Maynard, K.R.; Collado-Torres, L.; Weber, L.M.; Uytingco, C.; Barry, B.K.; Williams, S.R.; Ii, J.L.C.; Tran, M.N.; Besich, Z.; Tippani, M.; et al. Transcriptome-scale spatial gene expression in the human dorsolateral prefrontal cortex. Nat. Neurosci. 2021, 24, 425–436. [Google Scholar] [CrossRef]
- Ortiz, C.; Navarro, J.F.; Jurek, A.; Märtin, A.; Lundeberg, J.; Meletis, K. Molecular atlas of the adult mouse brain. Sci. Adv. 2020, 6, eabb3446. [Google Scholar] [CrossRef]
- Mathys, H.; Davila-Velderrain, J.; Peng, Z.; Gao, F.; Mohammadi, S.; Young, J.Z.; Menon, M.; He, L.; Abdurrob, F.; Jiang, X.; et al. Single-cell transcriptomic analysis of Alzheimer’s disease. Nature 2019, 570, 332–337. [Google Scholar] [CrossRef]
- Chen, W.-T.; Lu, A.; Craessaerts, K.; Pavie, B.; Frigerio, C.S.; Corthout, N.; Qian, X.; Laláková, J.; Kühnemund, M.; Voytyuk, I.; et al. Spatial Transcriptomics and In Situ Sequencing to Study Alzheimer’s Disease. Cell 2020, 182, 976–991.e19. [Google Scholar] [CrossRef]
- Grubman, A.; Chew, G.; Ouyang, J.F.; Sun, G.; Choo, X.Y.; McLean, C.; Simmons, R.; Buckberry, S.; Vargas-Landin, D.B.; Poppe, D.; et al. A single-cell atlas of entorhinal cortex from individuals with Alzheimer’s disease reveals cell-type-specific gene expression regulation. Nat. Neurosci. 2019, 22, 2087–2097. [Google Scholar] [CrossRef]
- Zhou, Y.; Song, W.M.; Andhey, P.S.; Swain, A.; Levy, T.; Miller, K.R.; Poliani, P.L.; Cominelli, M.; Grover, S.; Gilfillan, S.; et al. Human and mouse single-nucleus transcriptomics reveal TREM2-dependent and TREM2-independent cellular responses in Alzheimer’s disease. Nat. Med. 2020, 26, 131–142. [Google Scholar] [CrossRef]
- Velmeshev, D.; Schirmer, L.; Jung, D.; Haeussler, M.; Perez, Y.; Mayer, S.; Bhaduri, A.; Goyal, N.; Rowitch, D.H.; Kriegstein, A.R. Single-cell genomics identifies cell type–specific molecular changes in autism. Science 2019, 364, 685–689. [Google Scholar] [CrossRef]
- Renthal, W.; Boxer, L.; Hrvatin, S.; Li, E.; Silberfeld, A.; Nagy, M.A.; Griffith, E.; Vierbuchen, T.; Greenberg, M.E. Characterization of human mosaic Rett syndrome brain tissue by single-nucleus RNA sequencing. Nat. Neurosci. 2018, 21, 1670–1679. [Google Scholar] [CrossRef]
- Nagy, C.; Maitra, M.; Tanti, A.; Suderman, M.; Théroux, J.-F.; Davoli, M.A.; Perlman, K.; Yerko, V.; Wang, Y.C.; Tripathy, S.J.; et al. Single-nucleus transcriptomics of the prefrontal cortex in major depressive disorder implicates oligodendrocyte precursor cells and excitatory neurons. Nat. Neurosci. 2020, 23, 771–781. [Google Scholar] [CrossRef]
- Lee, D.-S.; Luo, C.; Zhou, J.; Chandran, S.; Rivkin, A.; Bartlett, A.; Nery, J.R.; Fitzpatrick, C.; O’Connor, C.; Dixon, J.R.; et al. Simultaneous profiling of 3D genome structure and DNA methylation in single human cells. Nat. Methods 2019, 16, 999–1006. [Google Scholar] [CrossRef] [PubMed]
- Li, G.; Liu, Y.; Zhang, Y.; Kubo, N.; Yu, M.; Fang, R.; Kellis, M.; Ren, B. Joint profiling of DNA methylation and chromatin architecture in single cells. Nat. Methods 2019, 16, 991–993. [Google Scholar] [CrossRef]
- Luo, C.; Liu, H.; Xie, F.; Armand, E.J.; Siletti, K.; Bakken, T.E.; Fang, R.; Doyle, W.I.; Hodge, R.D.; Hu, L.; et al. Single Nucleus Multi-Omics Links Human Cortical Cell Regulatory Genome Diversity to Disease Risk Variants. bioRxiv 2019. [Google Scholar] [CrossRef] [Green Version]
- Butler, R.; Bates, G. Histone deacetylase inhibitors as therapeutics for polyglutamine disorders. Nat. Rev. Neurosci. 2006, 7, 784–796. [Google Scholar] [CrossRef] [PubMed]
- Fischer, A.; Sananbenesi, F.; Wang, X.; Dobbin, M.; Tsai, L.-H. Recovery of learning and memory is associated with chromatin remodelling. Nature 2007, 447, 178–182. [Google Scholar] [CrossRef] [PubMed]
- Govindarajan, N.; Agis-Balboa, R.C.; Walter, J.; Sananbenesi, F.; Fischer, A. Sodium Butyrate Improves Memory Function in an Alzheimer’s Disease Mouse Model When Administered at an Advanced Stage of Disease Progression. J. Alzheimer’s Dis. 2011, 26, 187–197. [Google Scholar] [CrossRef]
- Ricobaraza, A.; Cuadrado-Tejedor, M.; Marco, S.; Pérez-Otaño, I.; García-Osta, A. Phenylbutyrate rescues dendritic spine loss associated with memory deficits in a mouse model of Alzheimer disease. Hippocampus 2012, 22, 1040–1050. [Google Scholar] [CrossRef]
- Gardian, G.; Browne, S.E.; Choi, D.-K.; Klivenyi, P.; Gregorio, J.; Kubilus, J.K.; Ryu, H.; Langley, B.; Ratan, R.R.; Ferrante, R.J.; et al. Neuroprotective Effects of Phenylbutyrate in the N171-82Q Transgenic Mouse Model of Huntington’s Disease. J. Biol. Chem. 2005, 280, 556–563. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Z.-Y.; Schluesener, H.J. Oral Administration of Histone Deacetylase Inhibitor MS-275 Ameliorates Neuroinflammation and Cerebral Amyloidosis and Improves Behavior in a Mouse Model. J. Neuropathol. Exp. Neurol. 2013, 72, 178–185. [Google Scholar] [CrossRef]
- Sung, Y.M.; Lee, T.; Yoon, H.; DiBattista, A.; Song, J.M.; Sohn, Y.; Moffat, E.I.; Turner, R.; Jung, M.; Kim, J.; et al. Mercaptoacetamide-based class II HDAC inhibitor lowers Aβ levels and improves learning and memory in a mouse model of Alzheimer’s disease. Exp. Neurol. 2013, 239, 192–201. [Google Scholar] [CrossRef] [Green Version]
- Cuadrado-Tejedor, M.; Garciabarroso, C.; Sanzhez-Arias, J.; Mederos, S.; Rabal, O.; Ugarte, A.; Franco, R.; Pascuallucas, M.; Segura, V.; Perea, G.; et al. Concomitant histone deacetylase and phosphodiesterase 5 inhibition synergistically prevents the disruption in synaptic plasticity and it reverses cognitive impairment in a mouse model of Alzheimer’s disease. Clin. Epigenet. 2015, 7, 1–11. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cuadrado-Tejedor, M.; Garcia-Barroso, C.; Sanchez-Arias, J.A.; Rabal, O.; González, M.P.; Mederos, S.; Ugarte, A.; Franco, R.; Segura, V.; Perea, G.; et al. A First-in-Class Small-Molecule that Acts as a Dual Inhibitor of HDAC and PDE5 and that Rescues Hippocampal Synaptic Impairment in Alzheimer’s Disease Mice. Neuropharmacology 2017, 42, 524–539. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rabal, O.; Sanchez-Arias, J.A.; Cuadrado-Tejedor, M.; de Miguel, I.; González, M.P.; García-Barroso, C.; Ugarte, A.; de Mendoza, A.E.-H.; Sáez, E.; Espelosin, M.; et al. Design, Synthesis, and Biological Evaluation of First-in-Class Dual Acting Histone Deacetylases (HDACs) and Phosphodiesterase 5 (PDE5) Inhibitors for the Treatment of Alzheimer’s Disease. J. Med. Chem. 2016, 59, 8967–9004. [Google Scholar] [CrossRef] [PubMed]
- Hu, J.-P.; Xie, J.-W.; Wang, C.-Y.; Wang, T.; Wang, X.; Wang, S.-L.; Teng, W.-P.; Wang, Z.-Y. Valproate reduces tau phosphorylation via cyclin-dependent kinase 5 and glycogen synthase kinase 3 signaling pathways. Brain Res. Bull. 2011, 85, 194–200. [Google Scholar] [CrossRef]
- Green, K.N.; Steffan, J.S.; Martínez-Coria, H.; Sun, X.; Schreiber, S.S.; Thompson, L.M.; LaFerla, F.M. Nicotinamide Restores Cognition in Alzheimer’s Disease Transgenic Mice via a Mechanism Involving Sirtuin Inhibition and Selective Reduction of Thr231-Phosphotau. J. Neurosci. 2008, 28, 11500–11510. [Google Scholar] [CrossRef] [PubMed]
- Gauthier, L.R.; Charrin, B.C.; Borrell, M.; Dompierre, J.P.; Rangone, H.; Cordelières, F.; De Mey, J.; MacDonald, M.E.; Leßmann, V.; Humbert, S.; et al. Huntingtin Controls Neurotrophic Support and Survival of Neurons by Enhancing BDNF Vesicular Transport along Microtubules. Cell 2004, 118, 127–138. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dompierre, J.P.; Godin, J.; Charrin, B.C.; Cordelières, F.; King, S.J.; Humbert, S.; Saudou, F. Histone Deacetylase 6 Inhibition Compensates for the Transport Deficit in Huntington’s Disease by Increasing Tubulin Acetylation. J. Neurosci. 2007, 27, 3571–3583. [Google Scholar] [CrossRef]
- Hockly, E.; Richon, V.M.; Woodman, B.; Smith, D.L.; Zhou, X.; Rosa, E.; Sathasivam, K.; Ghazi-Noori, S.; Mahal, A.; Lowden, P.A.S.; et al. Suberoylanilide hydroxamic acid, a histone deacetylase inhibitor, ameliorates motor deficits in a mouse model of Huntington’s disease. Proc. Natl. Acad. Sci. USA 2003, 100, 2041–2046. [Google Scholar] [CrossRef] [Green Version]
- Ferrante, R.J.; Kubilus, J.K.; Lee, J.; Ryu, H.; Beesen, A.; Zucker, B.; Smith, K.; Kowall, N.W.; Ratan, R.R.; Luthi-Carter, R.; et al. Histone Deacetylase Inhibition by Sodium Butyrate Chemotherapy Ameliorates the Neurodegenerative Phenotype in Huntington’s Disease Mice. J. Neurosci. 2003, 23, 9418–9427. [Google Scholar] [CrossRef]
- Zhou, W.; Bercury, K.; Cummiskey, J.; Luong, N.; Lebin, J.; Freed, C.R. Phenylbutyrate Up-regulates the DJ-1 Protein and Protects Neurons in Cell Culture and in Animal Models of Parkinson Disease. J. Biol. Chem. 2011, 286, 14941–14951. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Steffan, J.S.; Bodai, L.; Pallos, J.; Poelman, M.; McCampbell, A.; Apostol, B.L.; Kazantsev, A.; Schmidt, E.; Zhu, Y.-Z.; Greenwald, M.; et al. Histone deacetylase inhibitors arrest polyglutamine-dependent neurodegeneration in Drosophila. Nat. Cell Biol. 2001, 413, 739–743. [Google Scholar] [CrossRef] [PubMed]
- Saft, C.; Lauter, T.; Kraus, P.H.; Przuntek, H.; Andrich, J.E. Dose-dependent improvement of myoclonic hyperkinesia due to Valproic acid in eight Huntington’s Disease patients: A case series. BMC Neurol. 2006, 6, 11. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jia, H.; Kast, R.J.; Steffan, J.S.; Thomas, E.A. Selective histone deacetylase (HDAC) inhibition imparts beneficial effects in Huntington’s disease mice: Implications for the ubiquitin–proteasomal and autophagy systems. Hum. Mol. Genet. 2012, 21, 5280–5293. [Google Scholar] [CrossRef]
- Collins, L.; Adriaanse, L.J.; Theratile, S.D.; Hegarty, S.V.; Sullivan, A.; O’Keeffe, G.W. Class-IIa Histone Deacetylase Inhibition Promotes the Growth of Neural Processes and Protects Them Against Neurotoxic Insult. Mol. Neurobiol. 2014, 51, 1432–1442. [Google Scholar] [CrossRef]
- Pan, Y.; Daito, T.; Sasaki, Y.; Chung, Y.H.; Xing, X.; Pondugula, S.; Swamidass, S.J.; Wang, T.; Kim, A.H.; Yano, H. Inhibition of DNA Methyltransferases Blocks Mutant Huntingtin-Induced Neurotoxicity. Sci. Rep. 2016, 6, 31022. [Google Scholar] [CrossRef]
- Monti, B.; Gatta, V.; Piretti, F.; Raffaelli, S.S.; Virgili, M.; Contestabile, A. Valproic Acid is Neuroprotective in the Rotenone Rat Model of Parkinson’s Disease: Involvement of α-Synuclein. Neurotox. Res. 2010, 17, 130–141. [Google Scholar] [CrossRef] [PubMed]
- Ryu, H.; Smith, K.; Camelo, S.I.; Carreras, I.; Lee, J.; Iglesias, A.H.; Dangond, F.; Cormier, K.A.; Cudkowicz, M.E.; Jr, R.H.B.; et al. Sodium phenylbutyrate prolongs survival and regulates expression of anti-apoptotic genes in transgenic amyotrophic lateral sclerosis mice. J. Neurochem. 2005, 93, 1087–1098. [Google Scholar] [CrossRef] [PubMed]
- Camelo, S.; Iglesias, A.H.; Hwang, D.; Due, B.; Ryu, H.; Smith, K.; Gray, S.; Imitola, J.; Duran, G.; Assaf, B.; et al. Transcriptional therapy with the histone deacetylase inhibitor trichostatin A ameliorates experimental autoimmune encephalomyelitis. J. Neuroimmunol. 2005, 164, 10–21. [Google Scholar] [CrossRef] [PubMed]
- Ge, Z.; Da, Y.; Xue, Z.; Zhang, K.; Zhuang, H.; Peng, M.; Li, Y.; Li, W.; Simard, A.; Hao, J.; et al. Vorinostat, a histone deacetylase inhibitor, suppresses dendritic cell function and ameliorates experimental autoimmune encephalomyelitis. Exp. Neurol. 2013, 241, 56–66. [Google Scholar] [CrossRef]
- Sales, A.; Biojone, C.; Terceti, M.S.; Guimarães, F.S.; Gomes, M.V.M.; Joca, S.R.L. Antidepressant-like effect induced by systemic and intra-hippocampal administration of DNA methylation inhibitors. Br. J. Pharmacol. 2011, 164, 1711–1721. [Google Scholar] [CrossRef] [Green Version]
- Covington, H.E.; Vialou, V.; LaPlant, Q.; Ohnishi, Y.N.; Nestler, E.J. Hippocampal-dependent antidepressant-like activity of histone deacetylase inhibition. Neurosci. Lett. 2011, 493, 122–126. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lin, H.; Geng, X.; Dang, W.; Wu, B.; Dai, Z.; Li, Y.; Yang, Y.; Zhang, H.; Shi, J. Molecular mechanisms associated with the antidepressant effects of the class I histone deacetylase inhibitor MS-275 in the rat ventrolateral orbital cortex. Brain Res. 2012, 1447, 119–125. [Google Scholar] [CrossRef]
- Schroeder, F.A.; Lin, C.L.; Crusio, W.; Akbarian, S. Antidepressant-Like Effects of the Histone Deacetylase Inhibitor, Sodium Butyrate, in the Mouse. Biol. Psychiatry 2007, 62, 55–64. [Google Scholar] [CrossRef]
- Yamawaki, Y.; Fuchikami, M.; Morinobu, S.; Segawa, M.; Matsumoto, T.; Yamawaki, S. Antidepressant-like effect of sodium butyrate (HDAC inhibitor) and its molecular mechanism of action in the rat hippocampus. World J. Biol. Psychiatry 2011, 13, 458–467. [Google Scholar] [CrossRef]
- Zádori, D.; Geisz, A.; Vámos, E.; Vécsei, L.; Klivényi, P. Valproate ameliorates the survival and the motor performance in a transgenic mouse model of Huntington’s disease. Pharmacol. Biochem. Behav. 2009, 94, 148–153. [Google Scholar] [CrossRef] [Green Version]
- Rane, P.; Shields, J.; Heffernan, M.; Guo, Y.; Akbarian, S.; King, J.A. The histone deacetylase inhibitor, sodium butyrate, alleviates cognitive deficits in pre-motor stage PD. Neuropharmacology 2012, 62, 2409–2412. [Google Scholar] [CrossRef]
- Jovanova, O.S.; (Nedeljkovic), I.P.; Spieler, D.; Walker, R.; Liu, C.; Luciano, M.; Bressler, J.; Brody, J.; Drake, A.J.; Evans, K.; et al. DNA Methylation Signatures of Depressive Symptoms in Middle-aged and Elderly Persons. JAMA Psychiatry 2018, 75, 949–959. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tsankova, N.M.; Berton, O.; Renthal, W.; Kumar, A.; Neve, R.L.; Nestler, E.J. Sustained hippocampal chromatin regulation in a mouse model of depression and antidepressant action. Nat. Neurosci. 2006, 9, 519–525. [Google Scholar] [CrossRef] [PubMed]
- Li, L.; Atef, A.; Piatek, A.; Ali, Z.; Piatek, M.; Aouida, M.; Sharakuu, A.; Mahjoub, A.; Wang, G.; Khan, M.S.; et al. Characterization and DNA-Binding Specificities of Ralstonia TAL-Like Effectors. Mol. Plant 2013, 6, 1318–1330. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Morbitzer, R.; Römer, P.; Boch, J.; Lahaye, T. Regulation of selected genome loci using de novo-engineered transcription activator-like effector (TALE)-type transcription factors. Proc. Natl. Acad. Sci. USA 2010, 107, 21617–21622. [Google Scholar] [CrossRef] [Green Version]
- Miller, J.; McLachlan, A.; Klug, A. Repetitive zinc-binding domains in the protein transcription factor IIIA from Xenopus oocytes. EMBO J. 1985, 4, 1609–1614. [Google Scholar] [CrossRef] [PubMed]
- Moscou, M.J.; Bogdanove, A.J. A Simple Cipher Governs DNA Recognition by TAL Effectors. Science 2009, 326, 1501. [Google Scholar] [CrossRef]
- Boch, J.; Scholze, H.; Schornack, S.; Landgraf, A.; Hahn, S.; Kay, S.; Lahaye, T.; Nickstadt, A.; Bonas, U. Breaking the Code of DNA Binding Specificity of TAL-Type III Effectors. Science 2009, 326, 1509–1512. [Google Scholar] [CrossRef] [PubMed]
- Kim, Y.G.; Cha, J.; Chandrasegaran, S. Hybrid restriction enzymes: Zinc finger fusions to Fok I cleavage domain. Proc. Natl. Acad. Sci. USA 1996, 93, 1156–1160. [Google Scholar] [CrossRef] [Green Version]
- Zhang, F.; Cong, L.; Lodato, S.; Kosuri, S.; Church, G.; Arlotta, P. Efficient construction of sequence-specific TAL effectors for modulating mammalian transcription. Nat. Biotechnol. 2011, 29, 149–153. [Google Scholar] [CrossRef] [Green Version]
- Miller, J.C.; Tan, S.; Qiao, G.; Barlow, K.; Wang, J.; Xia, D.F.; Meng, X.; Paschon, D.E.; Leung, E.; Hinkley, S.J.; et al. A TALE nuclease architecture for efficient genome editing. Nat. Biotechnol. 2010, 29, 143–148. [Google Scholar] [CrossRef] [PubMed]
- Wood, A.J.; Lo, T.-W.; Zeitler, B.; Pickle, C.S.; Ralston, E.J.; Lee, A.H.; Amora, R.; Miller, J.C.; Leung, E.; Meng, X.; et al. Targeted Genome Editing Across Species Using ZFNs and TALENs. Science 2011, 333, 307. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hockemeyer, D.; Wang, H.; Kiani, S.; Lai, C.S.; Gao, Q.; Cassady, J.P.; Cost, G.; Zhang, L.; Santiago, Y.; Miller, J.C.; et al. Genetic engineering of human pluripotent cells using TALE nucleases. Nat. Biotechnol. 2011, 29, 731–734. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Eguchi, A.; Wleklinski, M.J.; Spurgat, M.C.; Heiderscheit, E.A.; Kropornicka, A.S.; Vu, C.K.; Bhimsaria, D.; Swanson, S.A.; Stewart, R.; Ramanathan, P.; et al. Reprogramming cell fate with a genome-scale library of artificial transcription factors. Proc. Natl. Acad. Sci. USA 2016, 113, E8257–E8266. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Briggs, A.W.; Rios, X.; Chari, R.; Yang, L.; Zhang, F.; Mali, P.; Church, G. Iterative capped assembly: Rapid and scalable synthesis of repeat-module DNA such as TAL effectors from individual monomers. Nucleic Acids Res. 2012, 40, e117. [Google Scholar] [CrossRef] [Green Version]
- Pourcel, C.; Salvignol, G.; Vergnaud, G. CRISPR elements in Yersinia pestis acquire new repeats by preferential uptake of bacteriophage DNA, and provide additional tools for evolutionary studies. Microbiology 2005, 151, 653–663. [Google Scholar] [CrossRef] [Green Version]
- Mojica, F.J.; Diez-Villasenor, C.; Garcia-Martinez, J.; Soria, E. Intervening Sequences of Regularly Spaced Prokaryotic Repeats Derive from Foreign Genetic Elements. J. Mol. Evol. 2005, 60, 174–182. [Google Scholar] [CrossRef] [PubMed]
- Bolotin, A.; Quinquis, B.; Sorokin, A.; Ehrlich, S.D. Clustered regularly interspaced short palindrome repeats (CRISPRs) have spacers of extrachromosomal origin. Microbiology 2005, 151, 2551–2561. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mojica, F.J.M.; Rodriguez-Valera, F. The discovery of CRISPR in archaea and bacteria. FEBS J. 2016, 283, 3162–3169. [Google Scholar] [CrossRef]
- Tang, T.-H.; Bachellerie, J.-P.; Rozhdestvensky, T.; Bortolin, M.-L.; Huber, H.; Drungowski, M.; Elge, T.; Brosius, J.; Hüttenhofer, A. Identification of 86 candidates for small non-messenger RNAs from the archaeon Archaeoglobus fulgidus. Proc. Natl. Acad. Sci. USA 2002, 99, 7536–7541. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Deltcheva, E.; Chylinski, K.; Sharma, C.M.; Gonzales, K.; Chao, Y.; Pirzada, Z.A.; Eckert, M.R.; Vogel, J.; Charpentier, E. CRISPR RNA maturation by trans-encoded small RNA and host factor RNase III. Nature 2011, 471, 602–607. [Google Scholar] [CrossRef] [Green Version]
- Jansen, R.; Van Embden, J.D.A.; Gaastra, W.; Schouls, L.M. Identification of genes that are associated with DNA repeats in prokaryotes. Mol. Microbiol. 2002, 43, 1565–1575. [Google Scholar] [CrossRef] [PubMed]
- Shmakov, S.; Smargon, A.; Scott, D.; Cox, D.; Pyzocha, N.; Yan, W.; Abudayyeh, O.O.; Gootenberg, J.S.; Makarova, K.S.; Wolf, Y.I.; et al. Diversity and evolution of class 2 CRISPR–Cas systems. Nat. Rev. Microbiol. 2017, 15, 169–182. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, J.-J.; Orlova, N.; Oakes, B.L.; Ma, E.; Spinner, H.B.; Baney, K.L.M.; Chuck, J.; Tan, D.; Knott, G.J.; Harrington, L.B.; et al. CasX enzymes comprise a distinct family of RNA-guided genome editors. Nat. Cell Biol. 2019, 566, 218–223. [Google Scholar] [CrossRef] [PubMed]
- Jinek, M.; Jiang, F.; Taylor, D.W.; Sternberg, S.H.; Kaya, E.; Ma, E.; Anders, C.; Hauer, M.; Zhou, K.; Lin, S.; et al. Structures of Cas9 Endonucleases Reveal RNA-Mediated Conformational Activation. Science 2014, 343, 1247997. [Google Scholar] [CrossRef] [Green Version]
- Sternberg, S.H.; Redding, S.; Jinek, M.; Greene, E.C.; Doudna, J.A. DNA interrogation by the CRISPR RNA-guided endonuclease Cas9. Nature 2014, 507, 62–67. [Google Scholar] [CrossRef] [Green Version]
- Zetsche, B.; Gootenberg, J.S.; Abudayyeh, O.O.; Slaymaker, I.M.; Makarova, K.S.; Essletzbichler, P.; Volz, S.E.; Joung, J.; van der Oost, J.; Regev, A.; et al. Cpf1 Is a Single RNA-Guided Endonuclease of a Class 2 CRISPR-Cas System. Cell 2015, 163, 759–771. [Google Scholar] [CrossRef] [Green Version]
- Nishimasu, H.; Ran, F.A.; Hsu, P.; Konermann, S.; Shehata, S.I.; Dohmae, N.; Ishitani, R.; Zhang, F.; Nureki, O. Crystal Structure of Cas9 in Complex with Guide RNA and Target DNA. Cell 2014, 156, 935–949. [Google Scholar] [CrossRef] [Green Version]
- Jore, M.M.; Lundgren, M.; Van Duijn, E.; Bultema, J.B.; Westra, E.R.; Waghmare, S.P.; Wiedenheft, B.; Pul, Ü.; Wurm, R.; Wagner, R.; et al. Structural basis for CRISPR RNA-guided DNA recognition by Cascade. Nat. Struct. Mol. Biol. 2011, 18, 529–536. [Google Scholar] [CrossRef] [Green Version]
- Hsu, P.D.; Lander, E.S.; Zhang, F. Development and Applications of CRISPR-Cas9 for Genome Engineering. Cell 2014, 157, 1262–1278. [Google Scholar] [CrossRef] [Green Version]
- Cong, L.; Ran, F.A.; Cox, D.; Lin, S.; Barretto, R.; Habib, N.; Hsu, P.D.; Wu, X.; Jiang, W.; Marraffini, L.A.; et al. Multiplex Genome Engineering Using CRISPR/Cas Systems. Science 2013, 339, 819–823. [Google Scholar] [CrossRef] [Green Version]
- Mali, P.; Yang, L.; Esvelt, K.M.; Aach, J.; Guell, M.; DiCarlo, J.E.; Norville, J.E.; Church, G.M. RNA-Guided Human Genome Engineering via Cas9. Science 2013, 339, 823–826. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Doudna, J.A.; Charpentier, E. The new frontier of genome engineering with CRISPR-Cas9. Science 2014, 346, 1258096. [Google Scholar] [CrossRef]
- Charpentier, E.; Marraffini, L.A. Harnessing CRISPR-Cas9 immunity for genetic engineering. Curr. Opin. Microbiol. 2014, 19, 114–119. [Google Scholar] [CrossRef] [Green Version]
- Cho, S.W.; Kim, S.; Kim, J.M.; Kim, J.-S. Targeted genome engineering in human cells with the Cas9 RNA-guided endonuclease. Nat. Biotechnol. 2013, 31, 230–232. [Google Scholar] [CrossRef]
- Hwang, W.Y.; Fu, Y.; Reyon, D.; Maeder, M.L.; Tsai, S.Q.; Sander, J.D.; Peterson, R.T.; Yeh, J.-R.J.; Joung, J.K. Efficient genome editing in zebrafish using a CRISPR-Cas system. Nat. Biotechnol. 2013, 31, 227–229. [Google Scholar] [CrossRef]
- Kim, Y.; Cheong, S.-A.; Lee, J.G.; Lee, S.-W.; Lee, M.S.; Baek, I.-J.; Sung, Y.H. Generation of knockout mice by Cpf1-mediated gene targeting. Nat. Biotechnol. 2016, 34, 808–810. [Google Scholar] [CrossRef] [PubMed]
- Hur, J.K.; Kim, K.; Been, K.W.; Baek, G.; Ye, S.; Hur, J.W.; Ryu, S.-M.; Lee, Y.S.; Kim, J.-S. Targeted mutagenesis in mice by electroporation of Cpf1 ribonucleoproteins. Nat. Biotechnol. 2016, 34, 807–808. [Google Scholar] [CrossRef] [PubMed]
- Kleinstiver, B.P.; Tsai, S.Q.; Prew, M.S.; Nguyen, N.T.; Welch, M.M.; Lopez, J.M.; McCaw, Z.; Aryee, Z.R.M.M.J.; Joung, B.P.K.S.Q.T.M.S.P.N.T.N.M.M.W.J.M.L.Z.R.M.M.J.A.J.K. Genome-wide specificities of CRISPR-Cas Cpf1 nucleases in human cells. Nat. Biotechnol. 2016, 34, 869–874. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, D.K.J.K.K.W.B.S.-H.Y.J.-S.; Kim, J.; Hur, J.K.; Been, K.W.; Yoon, S.-H.; Kim, J.-S. Genome-wide analysis reveals specificities of Cpf1 endonucleases in human cells. Nat. Biotechnol. 2016, 34, 863–868. [Google Scholar] [CrossRef] [PubMed]
- Tóth, E.; Weinhardt, N.; Bencsura, P.; Huszár, K.; Kulcsár, P.I.; Tálas, A.; Fodor, E.; Welker, E. Cpf1 nucleases demonstrate robust activity to induce DNA modification by exploiting homology directed repair pathways in mammalian cells. Biol. Direct 2016, 11, 46. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mali, P.; Esvelt, K.M.; Church, G. Cas9 as a versatile tool for engineering biology. Nat. Methods 2013, 10, 957–963. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ran, F.A.; Cong, L.; Yan, W.X.; Scott, D.A.; Gootenberg, J.; Kriz, A.J.; Zetsche, B.; Shalem, O.; Wu, X.; Makarova, K.S.; et al. In vivo genome editing using Staphylococcus aureus Cas9. Nature 2015, 520, 186–191. [Google Scholar] [CrossRef] [PubMed]
- Yeh, C.D.; Richardson, C.D.; Corn, J.E. Advances in genome editing through control of DNA repair pathways. Nat. Cell Biol. 2019, 21, 1468–1478. [Google Scholar] [CrossRef]
- Koike-Yusa, H.; Li, Y.; Tan, E.-P.; Velasco-Herrera, M.D.C.; Yusa, K. Genome-wide recessive genetic screening in mammalian cells with a lentiviral CRISPR-guide RNA library. Nat. Biotechnol. 2014, 32, 267–273. [Google Scholar] [CrossRef] [PubMed]
- Birsoy, K.; Wang, T.; Chen, W.W.; Freinkman, E.; Abu-Remaileh, M.; Sabatini, D.M. An Essential Role of the Mitochondrial Electron Transport Chain in Cell Proliferation Is to Enable Aspartate Synthesis. Cell 2015, 162, 540–551. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, T.; Wei, J.J.; Sabatini, D.M.; Lander, E.S. Genetic Screens in Human Cells Using the CRISPR-Cas9 System. Science 2014, 343, 80–84. [Google Scholar] [CrossRef] [Green Version]
- Chen, S.; Sanjana, N.; Zheng, K.; Shalem, O.; Lee, K.; Shi, X.; Scott, D.A.; Song, J.; Pan, J.Q.; Weissleder, R.; et al. Genome-wide CRISPR Screen in a Mouse Model of Tumor Growth and Metastasis. Cell 2015, 160, 1246–1260. [Google Scholar] [CrossRef] [Green Version]
- Smits, A.H.; Ziebell, F.; Joberty, G.; Zinn, N.; Mueller, W.F.; Clauder-Münster, S.; Eberhard, D.; Savitski, M.F.; Grandi, P.; Jakob, P.; et al. Biological plasticity rescues target activity in CRISPR knock outs. Nat. Methods 2019, 16, 1087–1093. [Google Scholar] [CrossRef]
- Bae, S.; Kweon, J.; Kim, H.S.; Kim, J.-S. Microhomology-based choice of Cas9 nuclease target sites. Nat. Methods 2014, 11, 705–706. [Google Scholar] [CrossRef]
- Shen, M.W.; Arbab, M.; Hsu, J.Y.; Worstell, D.; Culbertson, S.J.; Krabbe, O.; Cassa, C.A.; Liu, D.R.; Gifford, D.K.; Sherwood, R.I. Predictable and precise template-free CRISPR editing of pathogenic variants. Nat. Cell Biol. 2018, 563, 646–651. [Google Scholar] [CrossRef] [PubMed]
- Allen, F.; Crepaldi, L.; Alsinet, C.; Strong, A.J.; Kleshchevnikov, V.; De Angeli, P.; Páleníková, P.; Khodak, A.; Kiselev, V.; Kosicki, M.; et al. Predicting the mutations generated by repair of Cas9-induced double-strand breaks. Nat. Biotechnol. 2019, 37, 64–72. [Google Scholar] [CrossRef] [PubMed]
- Iyer, S.; Suresh, S.; Guo, D.; Daman, K.; Chen, J.C.J.; Liu, P.; Zieger, M.; Luk, K.; Roscoe, B.P.; Mueller, C.; et al. Precise therapeutic gene correction by a simple nuclease-induced double-stranded break. Nat. Cell Biol. 2019, 568, 561–565. [Google Scholar] [CrossRef] [PubMed]
- Lin, S.; Staahl, B.T.; Alla, R.K.; Doudna, J.A. Enhanced homology-directed human genome engineering by controlled timing of CRISPR/Cas9 delivery. eLife 2014, 3, e04766. [Google Scholar] [CrossRef] [PubMed]
- Gasiunas, G.; Barrangou, R.; Horvath, P.; Siksnys, V. Cas9-crRNA ribonucleoprotein complex mediates specific DNA cleavage for adaptive immunity in bacteria. Proc. Natl. Acad. Sci. USA 2012, 109, E2579–E2586. [Google Scholar] [CrossRef] [Green Version]
- Gaudelli, N.M.; Komor, A.C.; Rees, H.A.; Packer, M.S.; Badran, A.H.; Bryson, D.I.; Liu, D.R. Programmable base editing of A•T to G•C in genomic DNA without DNA cleavage. Nature 2017, 551, 464–471. [Google Scholar] [CrossRef]
- Kim, Y.B.; Komor, A.; Levy, J.M.; Packer, M.S.; Zhao, K.; Liu, Y.B.K.A.C.K.J.M.L.M.S.P.K.T.Z.D.R. Increasing the genome-targeting scope and precision of base editing with engineered Cas9-cytidine deaminase fusions. Nat. Biotechnol. 2017, 35, 371–376. [Google Scholar] [CrossRef]
- Rees, H.A.; Komor, A.; Yeh, W.-H.; Caetano-Lopes, J.; Warman, M.; Edge, A.S.B.; Liu, D.R. Improving the DNA specificity and applicability of base editing through protein engineering and protein delivery. Nat. Commun. 2017, 8, 15790. [Google Scholar] [CrossRef] [Green Version]
- Komor, A.C.; Kim, Y.B.; Packer, M.S.; Zuris, J.A.; Liu, D.R. Programmable editing of a target base in genomic DNA without double-stranded DNA cleavage. Nature 2016, 533, 420–424. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yeh, W.-H.; Chiang, H.; Rees, H.A.; Edge, A.S.B.; Liu, D.R. In vivo base editing of post-mitotic sensory cells. Nat. Commun. 2018, 9, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Sadowski, I.; Ma, J.; Triezenberg, S.; Ptashne, M. GAL4-VP16 is an unusually potent transcriptional activator. Nature 1988, 335, 563–564. [Google Scholar] [CrossRef]
- Liu, P.-Q.; Rebar, E.J.; Zhang, L.; Liu, Q.; Jamieson, A.C.; Liang, Y.; Qi, H.; Li, P.-X.; Chen, B.; Mendel, M.C.; et al. Regulation of an Endogenous Locus Using a Panel of Designed Zinc Finger Proteins Targeted to Accessible Chromatin Regions. J. Biol. Chem. 2001, 276, 11323–11334. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, L.; Spratt, S.K.; Liu, Q.; Johnstone, B.; Qi, H.; Raschke, E.E.; Jamieson, A.C.; Rebar, E.J.; Wolffe, A.P.; Case, C.C. Synthetic Zinc Finger Transcription Factor Action at an Endogenous Chromosomal Site. J. Biol. Chem. 2000, 275, 33850–33860. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Beerli, R.R.; Segal, D.; Dreier, B.; Barbas, C.F. Toward controlling gene expression at will: Specific regulation of the erbB-2/HER-2 promoter by using polydactyl zinc finger proteins constructed from modular building blocks. Proc. Natl. Acad. Sci. USA 1998, 95, 14628–14633. [Google Scholar] [CrossRef] [Green Version]
- Maeder, M.L.; Linder, S.J.; Reyon, D.; Angstman, J.F.; Fu, Y.; Sander, J.D.; Joung, J.K. Robust, synergistic regulation of human gene expression using TALE activators. Nat. Chem. Biol. 2013, 10, 243–245. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Perez-Pinera, P.; Ousterout, D.G.; Brunger, J.M.; Farin, A.M.; Glass, K.A.; Guilak, F.; Crawford, G.E.; Hartemink, A.J.; Gersbach, C.A. Synergistic and tunable human gene activation by combinations of synthetic transcription factors. Nat. Methods 2013, 10, 239–242. [Google Scholar] [CrossRef] [PubMed]
- Beerli, R.R.; Dreier, B.; Barbas, C.F. Positive and negative regulation of endogenous genes by designed transcription factors. Proc. Natl. Acad. Sci. USA 2000, 97, 1495–1500. [Google Scholar] [CrossRef] [Green Version]
- Tanenbaum, M.E.; Gilbert, L.; Qi, L.S.; Weissman, J.S.; Vale, R.D. A Protein-Tagging System for Signal Amplification in Gene Expression and Fluorescence Imaging. Cell 2014, 159, 635–646. [Google Scholar] [CrossRef] [Green Version]
- Chavez, A.; Scheiman, J.; Vora, S.D.; Pruitt, B.; Tuttle, M.; Iyer, E.P.R.; Lin, S.; Kiani, S.; Guzman, C.; Wiegand, D.J.; et al. Highly efficient Cas9-mediated transcriptional programming. Nat. Methods 2015, 12, 326–328. [Google Scholar] [CrossRef] [Green Version]
- Hilton, I.B.; D’Ippolito, A.M.; Vockley, C.M.; Thakore, P.I.; Crawford, G.E.; Reddy, T.E.; Gersbach, C.A. Epigenome editing by a CRISPR-Cas9-based acetyltransferase activates genes from promoters and enhancers. Nat. Biotechnol. 2015, 33, 510–517. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Okada, M.; Kanamori, M.; Someya, K.; Nakatsukasa, H.; Yoshimura, A. Stabilization of Foxp3 expression by CRISPR-dCas9-based epigenome editing in mouse primary T cells. Epigenet. Chromatin 2017, 10, 1–17. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Wang, W.; Shan, L.; Han, L.; Ma, S.; Zhang, Y.; Hao, B.; Lin, Y.; Rong, Z. Gene activation in human cells using CRISPR/Cpf1-p300 and CRISPR/Cpf1-SunTag systems. Protein Cell 2018, 9, 380–383. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sajwan, S.; Mannervik, M. Gene activation by dCas9-CBP and the SAM system differ in target preference. Sci. Rep. 2019, 9, 1–11. [Google Scholar] [CrossRef] [Green Version]
- Konermann, S.; Brigham, M.; Trevino, A.E.; Joung, J.; Abudayyeh, O.O.; Barcena, C.; Hsu, P.; Habib, N.; Gootenberg, J.; Nishimasu, H.; et al. Genome-scale transcriptional activation by an engineered CRISPR-Cas9 complex. Nature 2015, 517, 583–588. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Klein, J.C.; Chen, W.; Gasperini, M.; Shendure, J. Identifying Novel Enhancer Elements with CRISPR-Based Screens. ACS Chem. Biol. 2018, 13, 326–332. [Google Scholar] [CrossRef]
- Yan, J.; Chen, S.-A.A.; Local, A.; Liu, T.; Qiu, Y.; Dorighi, K.M.; Preissl, S.; Rivera, C.M.; Wang, C.; Ye, Z.; et al. Histone H3 lysine 4 monomethylation modulates long-range chromatin interactions at enhancers. Cell Res. 2018, 28, 204–220. [Google Scholar] [CrossRef] [Green Version]
- Rodriguez, D.C.; Gjaltema, R.; Jilderda, L.J.; Jellema, P.; Dokter-Fokkens, J.; Ruiters, M.H.J.; Rots, D.C.-R.R.A.F.G.L.J.J.P.J.J.D.-F.M.H.J.R.M.G. Writing of H3K4Me3 overcomes epigenetic silencing in a sustained but context-dependent manner. Nat. Commun. 2016, 7, 12284. [Google Scholar] [CrossRef] [PubMed]
- Maeder, M.L.; Angstman, J.F.; Richardson, M.E.; Linder, S.J.; Cascio, V.M.; Tsai, S.Q.; Ho, Q.H.; Sander, J.D.; Reyon, D.; Bernstein, B.E.; et al. Targeted DNA demethylation and activation of endogenous genes using programmable TALE-TET1 fusion proteins. Nat. Biotechnol. 2013, 31, 1137–1142. [Google Scholar] [CrossRef] [Green Version]
- Gregory, D.J.; Zhang, Y.; Kobzik, L.; Fedulov, A.V. Specific transcriptional enhancement of inducible nitric oxide synthase by targeted promoter demethylation. Epigenetics 2013, 8, 1205–1212. [Google Scholar] [CrossRef] [Green Version]
- Huisman, C.; van der Wijst, M.; Schokker, M.; Blancafort, P.; Terpstra, M.M.; Kok, K.; van der Zee, A.G.J.; Schuuring, E.; Wisman, G.B.A.; Rots, M.G. Re-expression of Selected Epigenetically Silenced Candidate Tumor Suppressor Genes in Cervical Cancer by TET2-directed Demethylation. Mol. Ther. 2016, 24, 536–547. [Google Scholar] [CrossRef] [Green Version]
- Chen, H.; Kazemier, H.G.; De Groote, M.L.; Ruiters, M.H.J.; Xu, G.-L.; Rots, M.G. Induced DNA demethylation by targeting Ten-Eleven Translocation 2 to the human ICAM-1 promoter. Nucleic Acids Res. 2014, 42, 1563–1574. [Google Scholar] [CrossRef] [Green Version]
- Sgro, A.; Blancafort, P. Epigenome engineering: New technologies for precision medicine. Nucleic Acids Res. 2020, 48, 12453–12482. [Google Scholar] [CrossRef]
- Guan, J.-S.; Haggarty, S.J.; Giacometti, E.; Dannenberg, J.-H.; Joseph, N.; Gao, J.; Nieland, T.J.F.; Zhou, Y.; Wang, X.; Mazitschek, R.; et al. HDAC2 negatively regulates memory formation and synaptic plasticity. Nature 2009, 459, 55–60. [Google Scholar] [CrossRef] [PubMed]
- Intlekofer, K.A.; Berchtold, N.C.; Malvaez, M.; Carlos, A.J.; McQuown, S.C.; Cunningham, M.J.; Wood, M.A.; Cotman, C.W. Exercise and Sodium Butyrate Transform a Subthreshold Learning Event into Long-Term Memory via a Brain-Derived Neurotrophic factor-Dependent Mechanism. Neuropsychopharmacology 2013, 38, 2027–2034. [Google Scholar] [CrossRef]
- Sartor, G.C.; Malvezzi, A.M.; Kumar, A.; Andrade, N.S.; Wiedner, H.; Vilca, S.; Janczura, K.J.; Bagheri, A.; Al-Ali, H.; Powell, S.; et al. Enhancement of BDNF Expression and Memory by HDAC Inhibition Requires BET Bromodomain Reader Proteins. J. Neurosci. 2019, 39, 612–626. [Google Scholar] [CrossRef] [PubMed]
- Margolin, J.F.; Friedman, J.; Meyer, W.K.; Vissing, H.; Thiesen, H.J.; Rauscher, F.J. Kruppel-associated boxes are potent transcriptional repression domains. Proc. Natl. Acad. Sci. USA 1994, 91, 4509–4513. [Google Scholar] [CrossRef] [Green Version]
- Sgouras, D.N.; Athanasiou, M.A.; Beal, G.J.J.; Fisher, R.J.; Blair, D.G.; Mavrothalassitis, G.J. ERF: An ETS Domain Protein with Strong Transcriptional Repressor Activity, Can Suppress Ets-Associated Tumorigenesis and Is Regulated by Phosphorylation during Cell Cycle and Mitogenic Stimulation. EMBO J. 1995, 14, 4781–4793. [Google Scholar] [CrossRef]
- Ayer, D.E.; Laherty, C.D.; Lawrence, Q.A.; Armstrong, A.P.; Eisenman, R.N. Mad proteins contain a dominant transcription repression domain. Mol. Cell. Biol. 1996, 16, 5772–5781. [Google Scholar] [CrossRef] [Green Version]
- O’Geen, H.; Ren, C.; Nicolet, C.M.; Perez, A.; Halmai, J.; Le, V.M.; Mackay, J.; Farnham, P.J.; Segal, D.J. dCas9-based epigenome editing suggests acquisition of histone methylation is not sufficient for target gene repression. Nucleic Acids Res. 2017, 45, 9901–9916. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- O’Geen, H.; Bates, S.L.; Carter, S.S.; Nisson, K.A.; Halmai, J.; Fink, K.D.; Rhie, S.K.; Farnham, P.J.; Segal, D.J. Ezh2-dCas9 and KRAB-dCas9 enable engineering of epigenetic memory in a context-dependent manner. Epigenet. Chromatin 2019, 12, 1–20. [Google Scholar] [CrossRef]
- Yeo, N.C.; Chavez, A.; Lance-Byrne, A.; Chan, Y.; Menn, D.; Milanova, D.; Kuo, C.-C.; Guo, X.; Sharma, S.; Tung, A.; et al. An enhanced CRISPR repressor for targeted mammalian gene regulation. Nat. Methods 2018, 15, 611–616. [Google Scholar] [CrossRef] [PubMed]
- Bintu, L.; Yong, J.; Antebi, Y.; McCue, K.; Kazuki, Y.; Uno, N.; Oshimura, M.; Elowitz, M.B. Dynamics of epigenetic regulation at the single-cell level. Science 2016, 351, 720–724. [Google Scholar] [CrossRef] [Green Version]
- Huang, Y.-H.; Su, J.; Lei, Y.; Brunetti, L.; Gundry, M.; Zhang, X.; Jeong, M.; Li, W.; Goodell, M.A. DNA epigenome editing using CRISPR-Cas SunTag-directed DNMT3A. Genome Biol. 2017, 18, 1–11. [Google Scholar] [CrossRef] [Green Version]
- Amabile, A.; Migliara, A.; Capasso, P.; Biffi, M.; Cittaro, D.; Naldini, L.; Lombardo, A.L. Inheritable Silencing of Endogenous Genes by Hit-and-Run Targeted Epigenetic Editing. Cell 2016, 167, 219–232.e14. [Google Scholar] [CrossRef] [Green Version]
- Siddique, A.N.; Nunna, S.; Rajavelu, A.; Zhang, Y.; Jurkowska, R.Z.; Reinhardt, R.; Rots, M.G.; Ragozin, S.; Jurkowski, T.; Jeltsch, A. Targeted Methylation and Gene Silencing of VEGF-A in Human Cells by Using a Designed Dnmt3a–Dnmt3L Single-Chain Fusion Protein with Increased DNA Methylation Activity. J. Mol. Biol. 2013, 425, 479–491. [Google Scholar] [CrossRef]
- Sripathy, S.P.; Stevens, J.; Schultz, D.C. The KAP1 Corepressor Functions To Coordinate the Assembly of De Novo HP1-Demarcated Microenvironments of Heterochromatin Required for KRAB Zinc Finger Protein-Mediated Transcriptional Repression. Mol. Cell. Biol. 2006, 26, 8623–8638. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stolzenburg, S.; Rots, M.G.; Beltran, A.S.; Rivenbark, A.G.; Yuan, X.; Qian, H.; Strahl, B.D.; Blancafort, P. Targeted silencing of the oncogenic transcription factor SOX2 in breast cancer. Nucleic Acids Res. 2012, 40, 6725–6740. [Google Scholar] [CrossRef]
- Cong, L.; Zhou, R.; Kuo, Y.-C.; Cunniff, M.M.; Zhang, F. Comprehensive interrogation of natural TALE DNA-binding modules and transcriptional repressor domains. Nat. Commun. 2012, 3, 1–6. [Google Scholar] [CrossRef] [Green Version]
- Mlambo, T.; Nitsch, S.; Hildenbeutel, M.; Romito, M.; Müller, M.; Bossen, C.; Diederichs, S.; Cornu, T.I.; Cathomen, T.; Mussolino, C. Designer epigenome modifiers enable robust and sustained gene silencing in clinically relevant human cells. Nucleic Acids Res. 2018, 46, 4456–4468. [Google Scholar] [CrossRef]
- Qi, L.S.; Larson, M.H.; Gilbert, L.A.; Doudna, J.A.; Weissman, J.S.; Arkin, A.P.; Lim, W.A. Repurposing CRISPR as an RNA-Guided Platform for Sequence-Specific Control of Gene Expression. Cell 2013, 152, 1173–1183. [Google Scholar] [CrossRef] [Green Version]
- Gilbert, L.; Larson, M.H.; Morsut, L.; Liu, Z.; Brar, G.A.; Torres, S.E.; Stern-Ginossar, N.; Brandman, O.; Whitehead, E.H.; Doudna, J.A.; et al. CRISPR-Mediated Modular RNA-Guided Regulation of Transcription in Eukaryotes. Cell 2013, 154, 442–451. [Google Scholar] [CrossRef] [Green Version]
- Wang, H.; Guo, R.; Du, Z.; Bai, L.; Li, L.; Cui, J.; Li, W.; Hoffman, A.R.; Hu, J.-F. Epigenetic Targeting of Granulin in Hepatoma Cells by Synthetic CRISPR dCas9 Epi-suppressors. Mol. Ther. Nucleic Acids 2018, 11, 23–33. [Google Scholar] [CrossRef] [Green Version]
- Josipović, G.; Tadić, V.; Klasić, M.; Zanki, V.; Bečeheli, I.; Chung, F.; Ghantous, A.; Keser, T.; Madunić, J.; Bošković, M.; et al. Antagonistic and synergistic epigenetic modulation using orthologous CRISPR/dCas9-based modular system. Nucleic Acids Res. 2019, 47, 9637–9657. [Google Scholar] [CrossRef] [Green Version]
- Zalatan, J.; Lee, M.E.; Almeida, R.; Gilbert, L.; Whitehead, E.H.; La Russa, M.; Tsai, J.; Weissman, J.S.; Dueber, J.E.; Qi, L.S.; et al. Engineering Complex Synthetic Transcriptional Programs with CRISPR RNA Scaffolds. Cell 2015, 160, 339–350. [Google Scholar] [CrossRef] [Green Version]
- Cheng, A.W.; Jillette, N.; Lee, P.; Plaskon, D.; Fujiwara, Y.; Wang, W.; Taghbalout, A.; Wang, H. Casilio: A versatile CRISPR-Cas9-Pumilio hybrid for gene regulation and genomic labeling. Cell Res. 2016, 26, 254–257. [Google Scholar] [CrossRef] [Green Version]
- Nolis, I.K.; McKay, D.J.; Mantouvalou, E.; Lomvardas, S.; Merika, M.; Thanos, D. Transcription factors mediate long-range enhancer-promoter interactions. Proc. Natl. Acad. Sci. USA 2009, 106, 20222–20227. [Google Scholar] [CrossRef] [Green Version]
- Deng, W.; Rupon, J.W.; Krivega, I.; Breda, L.; Motta, I.; Jahn, K.S.; Reik, A.; Gregory, P.; Rivella, S.; Dean, A.; et al. Reactivation of Developmentally Silenced Globin Genes by Forced Chromatin Looping. Cell 2014, 158, 849–860. [Google Scholar] [CrossRef] [Green Version]
- Hao, N.; Shearwin, K.; Dodd, I. Programmable DNA looping using engineered bivalent dCas9 complexes. Nat. Commun. 2017, 8, 1–12. [Google Scholar] [CrossRef] [Green Version]
- Kim, J.H.; Rege, M.; Valeri, J.; Dunagin, M.C.; Metzger, A.; Titus, K.; Gilgenast, T.G.; Gong, W.; Beagan, J.A.; Raj, A.; et al. LADL: Light-activated dynamic looping for endogenous gene expression control. Nat. Methods 2019, 16, 633–639. [Google Scholar] [CrossRef]
- Swiech, L.; Heidenreich, M.; Banerjee, A.; Habib, N.; Li, Y.; Trombetta, J.J.; Sur, M.; Zhang, F. In vivo interrogation of gene function in the mammalian brain using CRISPR-Cas9. Nat. Biotechnol. 2015, 33, 102–106. [Google Scholar] [CrossRef]
- Kantor, B.; Tagliafierro, L.; Gu, J.; Zamora, M.E.; Ilich, E.; Grenier, C.; Huang, Z.Y.; Murphy, S.; Chiba-Falek, O. Downregulation of SNCA Expression by Targeted Editing of DNA Methylation: A Potential Strategy for Precision Therapy in PD. Mol. Ther. 2018, 26, 2638–2649. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Laganiere, J.; Kells, A.P.; Lai, J.T.; Guschin, D.; Paschon, D.E.; Meng, X.; Fong, L.K.; Yu, Q.; Rebar, E.J.; Gregory, P.; et al. An Engineered Zinc Finger Protein Activator of the Endogenous Glial Cell Line-Derived Neurotrophic Factor Gene Provides Functional Neuroprotection in a Rat Model of Parkinson’s Disease. J. Neurosci. 2010, 30, 16469–16474. [Google Scholar] [CrossRef] [Green Version]
- Baxter, P.S.; Márkus, N.M.; Dando, O.; He, X.; Al-Mubarak, B.R.; Qiu, J.; Hardingham, G.E. Targeted de-repression of neuronal Nrf2 inhibits α-synuclein accumulation. Cell Death Dis. 2021, 12, 1–15. [Google Scholar] [CrossRef]
- Bustos, F.; Ampuero, E.; Jury, N.; Aguilar, R.; Falahi, F.; Toledo, J.; Ahumada, J.; Lata, J.; Cubillos, P.; Henríquez, B.; et al. Epigenetic editing of the Dlg4/PSD95 gene improves cognition in aged and Alzheimer’s disease mice. Brain 2017, 140, 3252–3268. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- György, B.; Lööv, C.; Zaborowski, M.; Takeda, S.; Kleinstiver, B.; Commins, C.; Kastanenka, K.; Mu, D.; Volak, A.; Giedraitis, V.; et al. CRISPR/Cas9 Mediated Disruption of the Swedish APP Allele as a Therapeutic Approach for Early-Onset Alzheimer’s Disease. Mol. Ther. Nucleic Acids 2018, 11, 429–440. [Google Scholar] [CrossRef] [Green Version]
- Sun, J.; Carlson-Stevermer, J.; Das, U.; Shen, M.; Delenclos, M.; Snead, A.M.; Koo, S.Y.; Wang, L.; Qiao, D.; Loi, J.; et al. CRISPR/Cas9 editing of APP C-terminus attenuates β-cleavage and promotes α-cleavage. Nat. Commun. 2019, 10, 1–11. [Google Scholar] [CrossRef] [Green Version]
- Park, H.; Oh, J.; Shim, G.; Cho, B.; Chang, Y.; Kim, S.; Baek, S.; Kim, H.; Shin, J.; Choi, H.; et al. In vivo neuronal gene editing via CRISPR–Cas9 amphiphilic nanocomplexes alleviates deficits in mouse models of Alzheimer’s disease. Nat. Neurosci. 2019, 22, 524–528. [Google Scholar] [CrossRef]
- Garriga-Canut, M.; Agustín-Pavón, C.; Herrmann, F.; Sánchez, A.; Dierssen, M.; Fillat, C.; Isalan, M. Synthetic zinc finger repressors reduce mutant huntingtin expression in the brain of R6/2 mice. Proc. Natl. Acad. Sci. USA 2012, 109, E3136–E3145. [Google Scholar] [CrossRef] [Green Version]
- Merienne, N.; Vachey, G.; de Longprez, L.; Meunier, C.; Zimmer, V.; Perriard, G.; Canales, M.; Mathias, A.; Herrgott, L.; Beltraminelli, T.; et al. The Self-Inactivating KamiCas9 System for the Editing of CNS Disease Genes. Cell Rep. 2017, 20, 2980–2991. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Monteys, A.M.; Ebanks, S.A.; Keiser, M.S.; Davidson, B.L. CRISPR/Cas9 Editing of the Mutant Huntingtin Allele In vitro and In vivo. Mol. Ther. 2017, 25, 12–23. [Google Scholar] [CrossRef] [Green Version]
- Yang, S.; Chang, R.; Yang, H.; Zhao, T.; Hong, Y.; Kong, H.E.; Sun, X.; Qin, Z.; Jin, P.; Li, S.; et al. CRISPR/Cas9-mediated gene editing ameliorates neurotoxicity in mouse model of Huntington’s disease. J. Clin. Investig. 2017, 127, 2719–2724. [Google Scholar] [CrossRef] [Green Version]
- Zeitler, B.; Froelich, S.; Marlen, K.; Shivak, D.A.; Yu, Q.; Li, D.; Pearl, J.R.; Miller, J.C.; Zhang, L.; Paschon, D.E.; et al. Allele-selective transcriptional repression of mutant HTT for the treatment of Huntington’s disease. Nat. Med. 2019, 25, 1131–1142. [Google Scholar] [CrossRef]
- Yu, B.; Yuan, B.; Dai, J.-K.; Cheng, T.-L.; Xia, S.-N.; He, L.-J.; Yuan, Y.-T.; Zhang, Y.-F.; Xu, H.-T.; Xu, F.-Q.; et al. Reversal of Social Recognition Deficit in Adult Mice with MECP2 Duplication via Normalization of MeCP2 in the Medial Prefrontal Cortex. Neurosci. Bull. 2020, 36, 570–584. [Google Scholar] [CrossRef] [PubMed]
- Le, T.T.H.; Tran, N.T.; Dao, T.M.L.; Nguyen, D.D.; Do, H.D.; Ha, T.L.; Kühn, R.; Nguyen, T.L.; Rajewsky, K.; Chu, V.T. Efficient and Precise CRISPR/Cas9-Mediated MECP2 Modifications in Human-Induced Pluripotent Stem Cells. Front. Genet. 2019, 10, 625. [Google Scholar] [CrossRef] [PubMed]
- Croci, S.; Carriero, M.L.; Capitani, K.; Daga, S.; Donati, F.; Frullanti, E.; Lamacchia, V.; Tita, R.; Giliberti, A.; Valentino, F.; et al. High rate of HDR in gene editing of p.(Thr158Met) MECP2 mutational hotspot. Eur. J. Hum. Genet. 2020, 28, 1231–1242. [Google Scholar] [CrossRef]
- Liu, X.S.; Wu, H.; Krzisch, M.; Wu, X.; Graef, J.; Muffat, J.; Hnisz, D.; Li, C.; Yuan, B.; Xu, C.; et al. Rescue of Fragile X Syndrome Neurons by DNA Methylation Editing of the FMR1 Gene. Cell 2018, 172, 979–992.e6. [Google Scholar] [CrossRef] [Green Version]
- Nelson, C.E.; Gersbach, C.A. Engineering Delivery Vehicles for Genome Editing. Annu. Rev. Chem. Biomol. Eng. 2016, 7, 637–662. [Google Scholar] [CrossRef] [Green Version]
- Yin, H.; Kauffman, K.J.; Anderson, H.Y.K.J.K.D.G. Delivery technologies for genome editing. Nat. Rev. Drug Discov. 2017, 16, 387–399. [Google Scholar] [CrossRef] [PubMed]
- Ates, I.; Rathbone, T.; Stuart, C.; Bridges, P.H.; Cottle, R.N. Delivery Approaches for Therapeutic Genome Editing and Challenges. Genes 2020, 11, 1113. [Google Scholar] [CrossRef] [PubMed]
- Tagliafierro, L.; Ilich, E.; Moncalvo, M.; Gu, J.; Sriskanda, A.; Grenier, C.; Murphy, S.K.; Chiba-Falek, O.; Kantor, B. Lentiviral Vector Platform for the Efficient Delivery of Epigenome-editing Tools into Human Induced Pluripotent Stem Cell-derived Disease Models. J. Vis. Exp. 2019, 10, e59241. [Google Scholar] [CrossRef]
- Van Haasteren, J.; Li, J.; Scheideler, O.J.; Murthy, N.; Schaffer, D.V. The delivery challenge: Fulfilling the promise of therapeutic genome editing. Nat. Biotechnol. 2020, 38, 845–855. [Google Scholar] [CrossRef]
- Li, R.; Qu, Y.; Liu, Y.; Noor, A.F.; Tran, J. Characteristics and advantages of adeno-associated virus vector-mediated gene therapy for neurodegenerative diseases. Neural Regen. Res. 2019, 14, 931–938. [Google Scholar] [CrossRef]
- Fuentes, C.M.; Schaffer, D.V. Adeno-associated virus-mediated delivery of CRISPR-Cas9 for genome editing in the central nervous system. Curr. Opin. Biomed. Eng. 2018, 7, 33–41. [Google Scholar] [CrossRef]
- Hanafy, A.S.; Schoch, S.; Lamprecht, A. CRISPR/Cas9 Delivery Potentials in Alzheimer’s Disease Management: A Mini Review. Pharmaceutics 2020, 12, 801. [Google Scholar] [CrossRef] [PubMed]
- Van Alstyne, M.; Tattoli, I.; Delestrée, N.; Recinos, Y.; Workman, E.; Shihabuddin, L.S.; Zhang, C.; Mentis, G.Z.; Pellizzoni, L. Gain of toxic function by long-term AAV9-mediated SMN overexpression in the sensorimotor circuit. Nat. Neurosci. 2021, 24, 1–11. [Google Scholar] [CrossRef]
- Tong, S.; Moyo, B.; Lee, C.M.; Leong, K.; Bao, G. Engineered materials for in vivo delivery of genome-editing machinery. Nat. Rev. Mater. 2019, 4, 726–737. [Google Scholar] [CrossRef] [PubMed]
- Yin, H.; Song, C.-Q.; Dorkin, J.R.; Zhu, L.J.; Li, Y.; Wu, Q.; Park, A.; Yang, J.; Suresh, S.; Bizhanova, A.; et al. Therapeutic genome editing by combined viral and non-viral delivery of CRISPR system components in vivo. Nat. Biotechnol. 2016, 34, 328–333. [Google Scholar] [CrossRef] [PubMed]
- Kostyushev, D.; Kostyusheva, A.; Brezgin, S.; Smirnov, V.; Volchkova, E.; Lukashev, A.; Chulanov, V. Gene Editing by Extracellular Vesicles. Int. J. Mol. Sci. 2020, 21, 7362. [Google Scholar] [CrossRef]
- Huang, H.; Zou, X.; Zhong, L.; Hou, Y.; Zhou, J.; Zhang, Z.; Xing, X.; Sun, J. CRISPR/dCas9-mediated activation of multiple endogenous target genes directly converts human foreskin fibroblasts into Leydig-like cells. J. Cell. Mol. Med. 2019, 23, 6072–6084. [Google Scholar] [CrossRef] [Green Version]
- Fidanza, A.; Lopez-Yrigoyen, M.; Romanò, N.; Jones, R.; Taylor, A.H.; Forrester, L.M. An all-in-one UniSam vector system for efficient gene activation. Sci. Rep. 2017, 7, 6394. [Google Scholar] [CrossRef]
- Gao, Y.; Xiong, X.; Wong, S.; Charles, E.J.; Lim, W.A.; Qi, L.S. Complex transcriptional modulation with orthogonal and inducible dCas9 regulators. Nat. Methods 2016, 13, 1043–1049. [Google Scholar] [CrossRef] [PubMed]
- Liang, F.-S.; Ho, W.Q.; Crabtree, G.R. Engineering the ABA Plant Stress Pathway for Regulation of Induced Proximity. Sci. Signal. 2011, 4, rs2. [Google Scholar] [CrossRef] [Green Version]
- Miyamoto, T.; DeRose, R.; Suarez, A.; Ueno, T.; Chen, M.; Sun, T.-P.; Wolfgang, M.J.; Mukherjee, C.; Meyers, D.J.; Inoue, T. Rapid and orthogonal logic gating with a gibberellin-induced dimerization system. Nat. Chem. Biol. 2012, 8, 465–470. [Google Scholar] [CrossRef] [PubMed]
- Braun, S.; Kirkland, J.G.; Chory, E.J.; Husmann, D.; Calarco, J.P.; Crabtree, G.R. Rapid and reversible epigenome editing by endogenous chromatin regulators. Nat. Commun. 2017, 8, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Konermann, S.M.; Brigham, M.D.; Trevino, A.E.; Hsu, P.; Heidenreich, M.; Cong, L.; Platt, R.; Scott, D.A.; Church, G.; Zhang, F. Optical control of mammalian endogenous transcription and epigenetic states. Nature 2013, 500, 472–476. [Google Scholar] [CrossRef]
- Polstein, L.R.; Gersbach, C.A. Light-Inducible Spatiotemporal Control of Gene Activation by Customizable Zinc Finger Transcription Factors. J. Am. Chem. Soc. 2012, 134, 16480–16483. [Google Scholar] [CrossRef]
- Singh, R.; Kuscu, C.; Quinlan, A.; Qi, Y.; Adli, M. Cas9-chromatin binding information enables more accurate CRISPR off-target prediction. Nucleic Acids Res. 2015, 43, e118. [Google Scholar] [CrossRef]
- Hruscha, A.; Krawitz, P.; Rechenberg, A.; Heinrich, V.; Hecht, J.; Haass, C.; Schmid, B. Efficient CRISPR/Cas9 genome editing with low off-target effects in zebrafish. Development 2013, 140, 4982–4987. [Google Scholar] [CrossRef] [Green Version]
- Cho, S.W.; Kim, S.; Kim, Y.; Kweon, J.; Kim, H.S.; Bae, S.; Kim, J.-S. Analysis of off-target effects of CRISPR/Cas-derived RNA-guided endonucleases and nickases. Genome Res. 2014, 24, 132–141. [Google Scholar] [CrossRef] [Green Version]
- Cradick, T.; Fine, E.; Antico, C.J.; Bao, G. CRISPR/Cas9 systems targeting β-globin and CCR5 genes have substantial off-target activity. Nucleic Acids Res. 2013, 41, 9584–9592. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mali, P.; Aach, J.; Stranges, P.; Esvelt, K.; Moosburner, M.; Kosuri, S.; Yang, L.; Church, G.M. CAS9 transcriptional activators for target specificity screening and paired nickases for cooperative genome engineering. Nat. Biotechnol. 2013, 31, 833–838. [Google Scholar] [CrossRef]
- Heigwer, F.; Kerr, G.; Boutros, M. E-CRISP: Fast CRISPR target site identification. Nat. Methods 2014, 11, 122–123. [Google Scholar] [CrossRef] [PubMed]
- Ran, F.A.; Hsu, P.D.; Wright, J.; Agarwala, V.; Scott, D.A.; Zhang, F. Genome engineering using the CRISPR-Cas9 system. Nat. Protoc. 2013, 8, 2281–2308. [Google Scholar] [CrossRef] [Green Version]
- Doench, J.G.; Root, D.E. Optimized SgRNA Design to Maximize Activity and Minimize Off-Target Effects of CRISPR-Cas9 Synthesis of an Arrayed SgRNA Library Targeting the Human Genome. Nat. Biotechnol. 2016, 34, 184–191. [Google Scholar] [CrossRef] [Green Version]
- Senís, E.; Fatouros, C.; Große, S.; Wiedtke, E.; Niopek, D.; Mueller, A.-K.; Börner, K.; Grimm, D. CRISPR/Cas9-mediated genome engineering: An adeno-associated viral (AAV) vector toolbox. Biotechnol. J. 2014, 9, 1402–1412. [Google Scholar] [CrossRef] [PubMed]
- Niu, Y.; Shen, B.; Cui, Y.; Chen, Y.; Wang, J.; Wang, L.; Kang, Y.; Zhao, X.; Si, W.; Li, W.; et al. Generation of Gene-Modified Cynomolgus Monkey via Cas9/RNA-Mediated Gene Targeting in One-Cell Embryos. Cell 2014, 156, 836–843. [Google Scholar] [CrossRef] [Green Version]
- Sato, K.; Oiwa, R.; Kumita, W.; Henry, R.; Sakuma, T.; Ito, R.; Nozu, R.; Inoue, T.; Katano, I.; Sato, K.; et al. Generation of a Nonhuman Primate Model of Severe Combined Immunodeficiency Using Highly Efficient Genome Editing. Cell Stem Cell 2016, 19, 127–138. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schacker, M.; Seimetz, D. From fiction to science: Clinical potentials and regulatory considerations of gene editing. Clin. Transl. Med. 2019, 8, 1–16. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gillmore, J.D.; Gane, E.; Taubel, J.; Kao, J.; Fontana, M.; Maitland, M.L.; Seitzer, J.; O’Connell, D.; Walsh, K.R.; Wood, K.; et al. CRISPR-Cas9 In vivo Gene Editing for Transthyretin Amyloidosis. N. Engl. J. Med. 2021. [Google Scholar] [CrossRef]
Figure 1.
Chromatin 3D structure in the cell in relation to transcriptional regulation. Promoter-enhancer loops form within the context of the topologically associating domains (TADs), which arise as a consequence of the action of the cohesin complex. The binding of CTCF blocks the displacement of the cohesin ring thereby defining the location of the TADs boundaries. Active promoters and enhancers are devoid of DNA methylation. The two classes of elements display a specific histone modification signature featuring high levels of H3K27ac (both enhancer and promoters) and unique pattern of H3K4me3 modification (promoters being highly enriched and enhancers displaying non-detectable or low level of this modification). H3K4me1 is particularly enriched at active and poised enhancers. When active, promoters and enhancers can often display an increased spatial proximity in the cell nucleus.
Figure 1.
Chromatin 3D structure in the cell in relation to transcriptional regulation. Promoter-enhancer loops form within the context of the topologically associating domains (TADs), which arise as a consequence of the action of the cohesin complex. The binding of CTCF blocks the displacement of the cohesin ring thereby defining the location of the TADs boundaries. Active promoters and enhancers are devoid of DNA methylation. The two classes of elements display a specific histone modification signature featuring high levels of H3K27ac (both enhancer and promoters) and unique pattern of H3K4me3 modification (promoters being highly enriched and enhancers displaying non-detectable or low level of this modification). H3K4me1 is particularly enriched at active and poised enhancers. When active, promoters and enhancers can often display an increased spatial proximity in the cell nucleus.

Figure 2.
Mechanisms of action of TALE (upper panel) and CRISPR-(d)Cas9 (lower panel). In the TALE system, a designed protein chimera, consisting of transcription activator-like domains (TAL) each recognizing a specific base in the DNA molecule, binds to the target sequence. Fused with a chosen effector domain (ED), TAL effectors (TALEs) can tether a desired enzymatic activity to the target site. This way TALEs can be used to modify the DNA sequence (when endonuclease FokI is the ED) or alter the chromatin landscape. The later can be achieved by fusing TAL with for instance VP16 transactivation domain or the enzymatic domain of p300 HAT allowing to enhance gene expression. Fusion of TAL with KRAB and MeCP2 can induce locus silencing. In CRISPR-Cas9 system, a single guide RNA molecule (sgRNA: guide RNA fused with tracrRNA), harboring a region complementary to the locus of interest, stabilizes the binding of the Cas9 endonuclease and favors the introduction of a double- (Cas9) or a single-strand (nickase-Cas9) DNA break at the recognized site and mutation in the target DNA sequence. When using a catalytically inactive Cas9 (dCas9), this system can also be harnessed to tether an ED to a locus of interest. The dCas9-based epigenome engineering can also be used to modify genome topology and re-introduce promoter-enhancer loops.
Figure 2.
Mechanisms of action of TALE (upper panel) and CRISPR-(d)Cas9 (lower panel). In the TALE system, a designed protein chimera, consisting of transcription activator-like domains (TAL) each recognizing a specific base in the DNA molecule, binds to the target sequence. Fused with a chosen effector domain (ED), TAL effectors (TALEs) can tether a desired enzymatic activity to the target site. This way TALEs can be used to modify the DNA sequence (when endonuclease FokI is the ED) or alter the chromatin landscape. The later can be achieved by fusing TAL with for instance VP16 transactivation domain or the enzymatic domain of p300 HAT allowing to enhance gene expression. Fusion of TAL with KRAB and MeCP2 can induce locus silencing. In CRISPR-Cas9 system, a single guide RNA molecule (sgRNA: guide RNA fused with tracrRNA), harboring a region complementary to the locus of interest, stabilizes the binding of the Cas9 endonuclease and favors the introduction of a double- (Cas9) or a single-strand (nickase-Cas9) DNA break at the recognized site and mutation in the target DNA sequence. When using a catalytically inactive Cas9 (dCas9), this system can also be harnessed to tether an ED to a locus of interest. The dCas9-based epigenome engineering can also be used to modify genome topology and re-introduce promoter-enhancer loops.

Figure 3.
NDs frequently feature altered chromatin landscape including promoter silencing and gene repression. Global or locus-specific approaches can be envisioned to restore proper gene activity. Epidrugs, including histone deacetylase inhibitors (HDACi) or DNA methyltransferase inhibitors (DNMTi), exert a global impact on chromatin and can help reactivate a proportion of aberrantly silenced genes. Designer nuclease-based systems (TALEs and CRISPR-Cas9-based) modify chromatin structure locally and specifically impacting by and large only the chosen loci. Designer nuclease-based systems display minimal off-target effects and potentially extremely low toxicity.
Figure 3.
NDs frequently feature altered chromatin landscape including promoter silencing and gene repression. Global or locus-specific approaches can be envisioned to restore proper gene activity. Epidrugs, including histone deacetylase inhibitors (HDACi) or DNA methyltransferase inhibitors (DNMTi), exert a global impact on chromatin and can help reactivate a proportion of aberrantly silenced genes. Designer nuclease-based systems (TALEs and CRISPR-Cas9-based) modify chromatin structure locally and specifically impacting by and large only the chosen loci. Designer nuclease-based systems display minimal off-target effects and potentially extremely low toxicity.

Figure 4.
Possible approaches to modulate the activity of the non-coding sequences affected in NDs, including regulatory elements identified in genome-wide association studies (GWAS). (A) Manhattan plot displaying an example of GWAS hits linked to a ND. (B) GWAS hits may intersect enhancers that can be aberrantly repressed (left panel) or activated (right panel) in NDs. (C) (Epi)genome editing strategies to activate an aberrantly silenced enhancer element in ND. (D) Like in (C) but displayed are strategies to repress an aberrantly activated enhancer. CRISPR-dCas9 system consists of DNA-binding domain (catalytically deficient Cas9 (dCas9)), and an effector domain which either activates (ActD) or represses (RepD) the activity of a regulatory sequence (here an enhancer). As a consequence, this leads to changes in the expression of the enhancer’s target gene. Gene re-activation is possible via tethering of the dCas9 fused with VP (16, 48, or 64) or the catalytic domain of the p300 HAT to a chosen sequence. Gene repression can be achieved by bringing a dCas9 fused with KRAB, KRAB-MECP2, or HDAC enzymatic domain to the aberrantly activated regulatory sequence. The alternative approach combines the CRISPR-dCas9 system with viral aptamers, recognizing specific RNA sequences encoded within the sgRNA construct (PP7, MS2, Com, or PBS). The aptamers are fused with distinct effector domains: ActD activation domain, RepD- repression domains with activatory or inhibitory impact. (E) Complex design allowing to simultaneously repress a set of chosen elements and activate another group of enhancers. (F) Gene expression can also be increased due to the induction of the formation of an enhancer-promoter contact. In the CLOuD9 approach, the addition of abscisic acid (ABA) stabilizes the interaction between two complementary dCas9 molecules (Cas9 S. pyogenes and Cas9 s aureus.) tethered to an enhancer and promoter respectively.
Figure 4.
Possible approaches to modulate the activity of the non-coding sequences affected in NDs, including regulatory elements identified in genome-wide association studies (GWAS). (A) Manhattan plot displaying an example of GWAS hits linked to a ND. (B) GWAS hits may intersect enhancers that can be aberrantly repressed (left panel) or activated (right panel) in NDs. (C) (Epi)genome editing strategies to activate an aberrantly silenced enhancer element in ND. (D) Like in (C) but displayed are strategies to repress an aberrantly activated enhancer. CRISPR-dCas9 system consists of DNA-binding domain (catalytically deficient Cas9 (dCas9)), and an effector domain which either activates (ActD) or represses (RepD) the activity of a regulatory sequence (here an enhancer). As a consequence, this leads to changes in the expression of the enhancer’s target gene. Gene re-activation is possible via tethering of the dCas9 fused with VP (16, 48, or 64) or the catalytic domain of the p300 HAT to a chosen sequence. Gene repression can be achieved by bringing a dCas9 fused with KRAB, KRAB-MECP2, or HDAC enzymatic domain to the aberrantly activated regulatory sequence. The alternative approach combines the CRISPR-dCas9 system with viral aptamers, recognizing specific RNA sequences encoded within the sgRNA construct (PP7, MS2, Com, or PBS). The aptamers are fused with distinct effector domains: ActD activation domain, RepD- repression domains with activatory or inhibitory impact. (E) Complex design allowing to simultaneously repress a set of chosen elements and activate another group of enhancers. (F) Gene expression can also be increased due to the induction of the formation of an enhancer-promoter contact. In the CLOuD9 approach, the addition of abscisic acid (ABA) stabilizes the interaction between two complementary dCas9 molecules (Cas9 S. pyogenes and Cas9 s aureus.) tethered to an enhancer and promoter respectively.


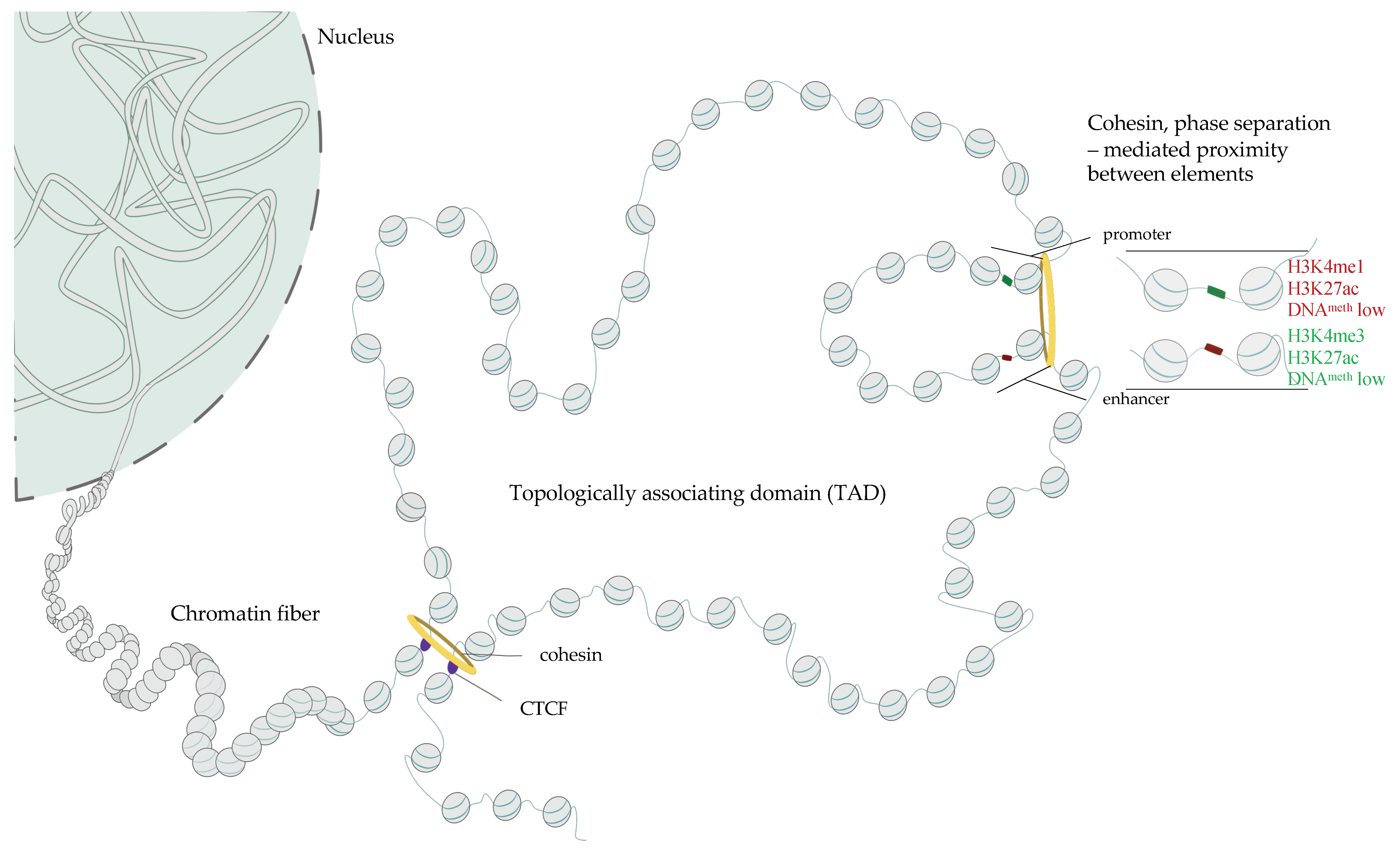{kind=link}
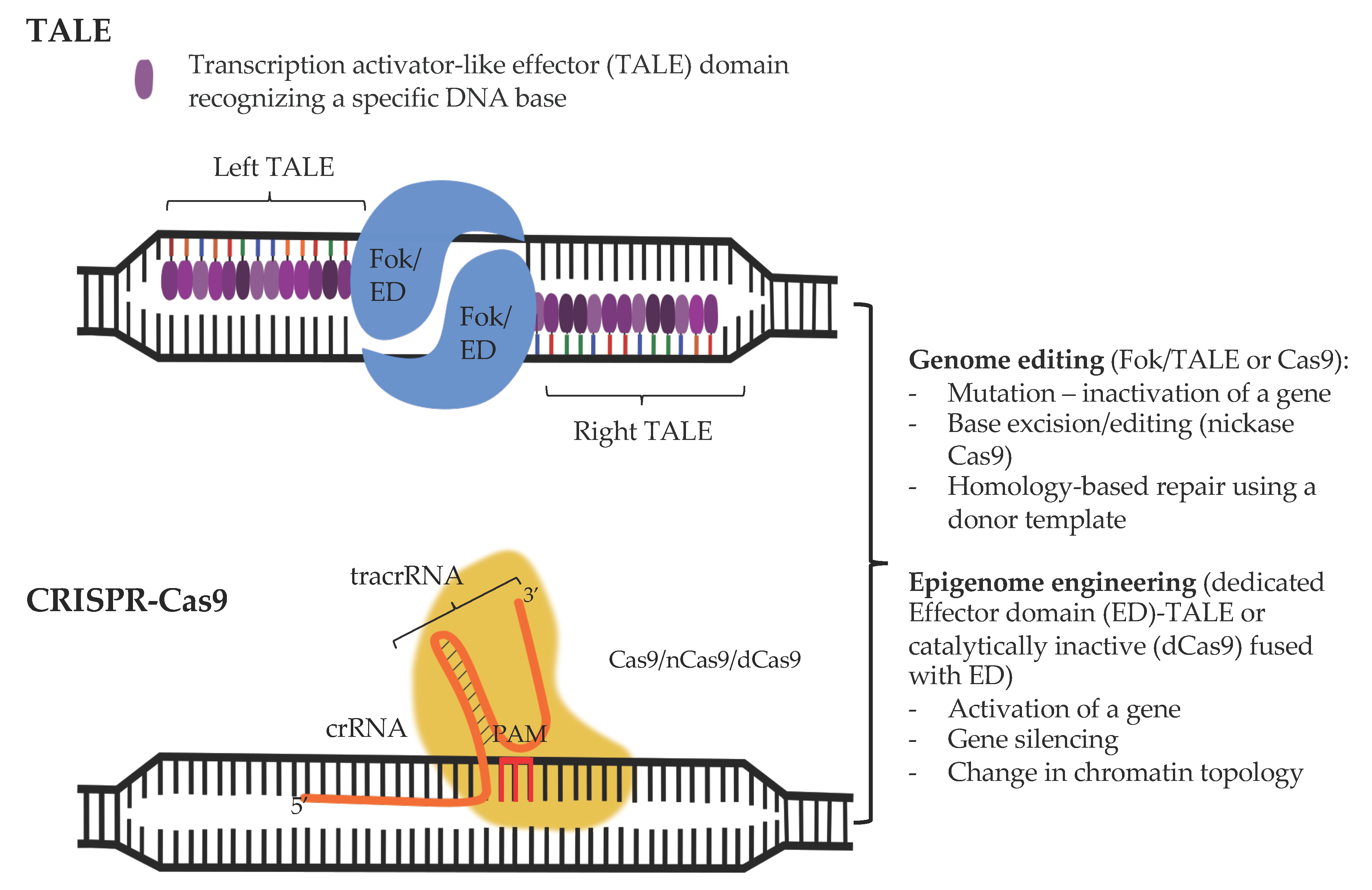{kind=link}
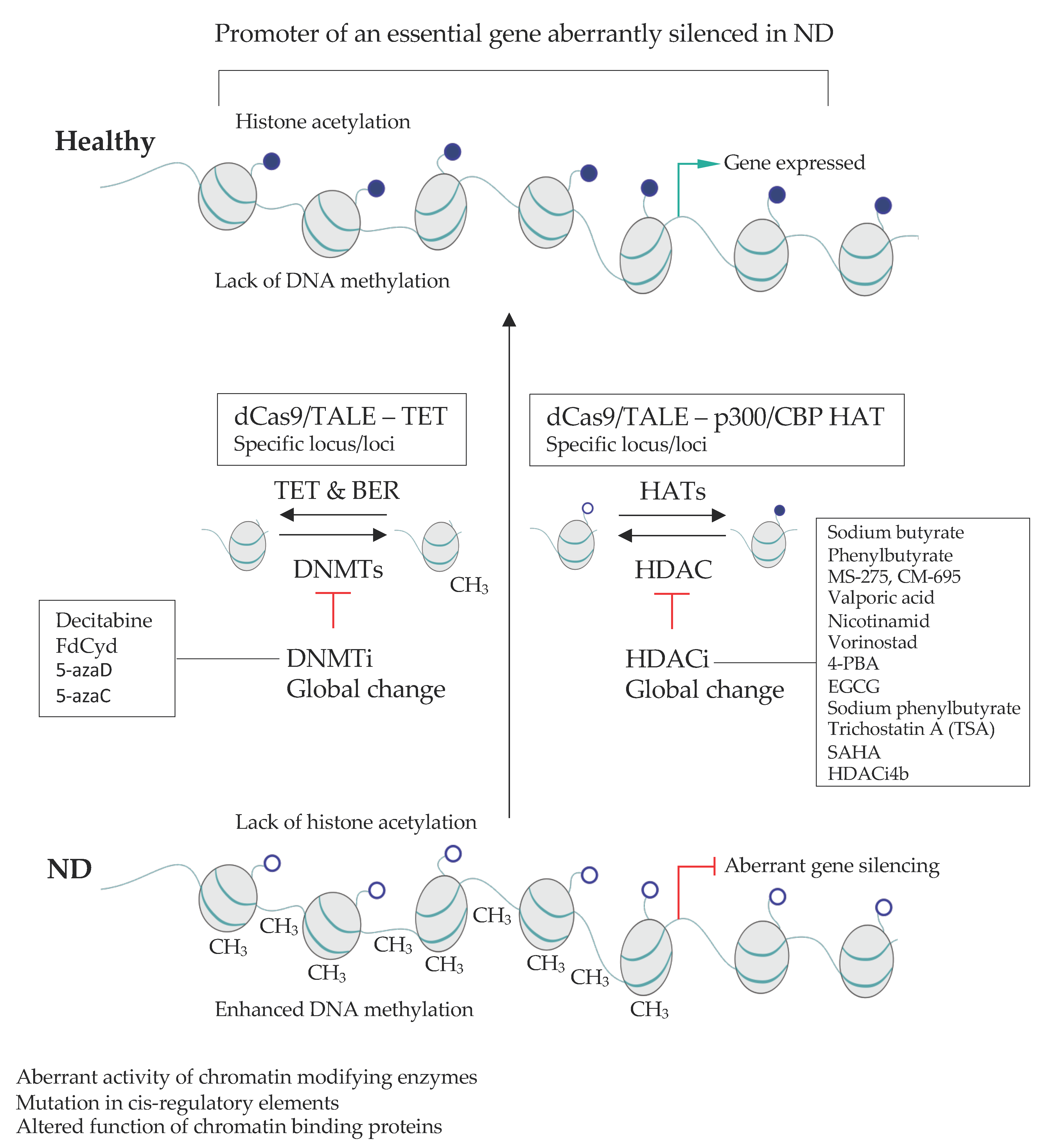{kind=link}
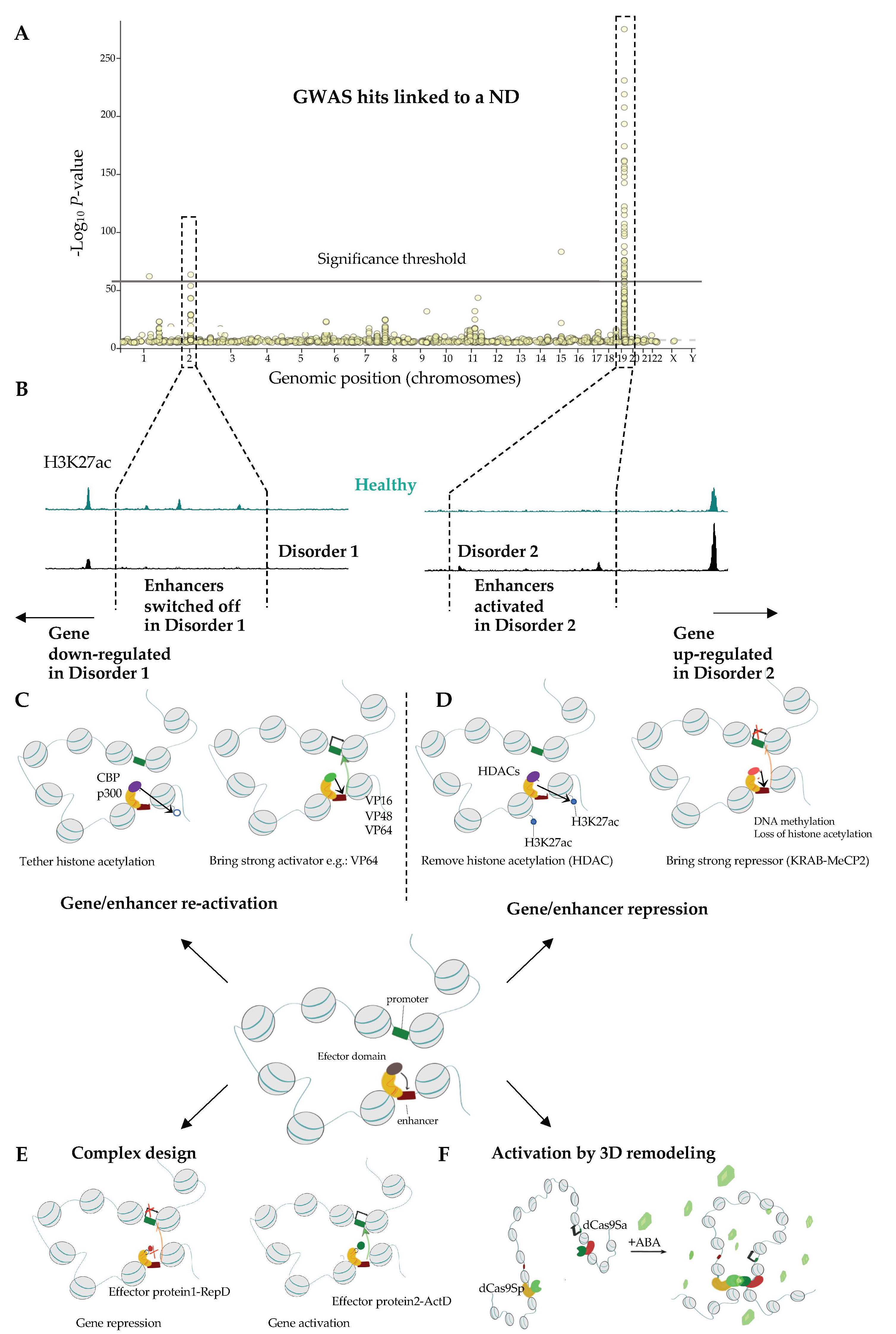{kind=link}
Table 1.
Chromatin-related factors implicated in NDs.
| Protein | Disease | Phenotype | Reference |
|---|---|---|---|
| DNA Methylases and DNA Methylation-Related Machinery | |||
| MECP2 | Rett syndrome | Encephalopathy, intellectual disability (ID), epilepsy | [116,117,118,119] |
| DNMT1 | Hereditary Sensory Neuropathy Type IE—heterozygous mutation | [120] | |
| autonomic neuropathies with dementia and hearing loss (HSANIE)—heterozygous mutation | [121] | ||
| cerebellar ataxia with deafness and narcolepsy | cerebellar ataxia, narcolepsy/cataplexy, sensorineural deafness, and dementia | [122] | |
| DNMT3a | Tatton-Brown-Rahman syndrome—heterozygous mutation | intellectual disability | [123,124,125] |
| Heyn-Sproul-Jackson syndrome—heterozygous mutation | microcephalic dwarfism | [126] | |
| Histone Acetylases | |||
| P300 | Rubinstein-Taybi syndrome—heterozygous mutation | microcephaly, mental retardation | [127,128,129] |
| Menke-Hennekam syndrome—heterozygous mutation | microcephaly, mental retardation, autistic behavior | [128,130] | |
| CBP | Rubinstein-Taybi syndrome | microcephaly, mental retardation, postnatal growth deficiency | [131,132,133] |
| Menke-Hennekam syndrome | microcephaly, Intellectual impairment, autistic behavior | [130,134,135] | |
| MOZ/MYST3/KAT6A | Arboleda-Tham syndrome—heterozygous mutation | microcephaly, impaired intellectual development | [136,137,138,139] |
| MORF/MYST4/KAT6B | SBBYSS syndrome | mental retardation | [140,141,142,143] |
| Genitopatellar syndrome | microcephaly, severe psychomotor retardation | [144,145] | |
| MOF/MYST1/KAT8 | Li-Ghorbani-Weisz-Hubshman syndrome—heterozygous mutation | developmental delay, impaired intellectual development | [146] |
| Histone deacetylases | |||
| HDAC6 | dominant X-linked chondrodyspasia | Chondrodysplasia with platyspondyly, distinctive brachydactyly, hydrocephaly, microphthalmia | [147,148] |
| HDAC8 | Cornelia de Lange syndrome | dysmorphic facial features, cleft palate, distal limb defects, growth retardation, developmental delay | [149,150] |
| SIRT1 | anxiety disorder, panic disorder and social phobia | feeling persistent fear, panic attacks, fear of social situations | [151] |
| Chromatin remodelers | |||
| SWI/SNF complex | |||
| BRM/SMARCA2 | Nicolaides-Baraitser syndrome—heterozygous mutation | severe mental retardation, early-onset seizures, short stature, dysmorphic facial features | [152,153] |
| Schizophrenia | delusions, disorganized thinking, hallucinations | [154] | |
| Coffin-Siris syndrome 4 | developmental delay, intellectual disability, coarse facial features, feeding difficulties | [155] | |
| BRG1/SMARCA4 | Rhabdoid Tumor Predisposition syndrome 2; RTPS1—heterozygous mutation | brain, spinal cord and kidney tumors | [156,157] |
| BAF47/SMARCB1 | Schwannomatosis 1—heterozygous mutation | multiple cutaneous neurilemmomas, spinal schwannomas | [158,159,160,161] |
| Multiple meningiomas | meningeal tumor | [158] | |
| Coffin-Siris syndrome | developmental delay, intellectual disability, coarse facial features, feeding difficulties | [155] | |
| BAF170/SMARCC2 | Coffin-Siris syndrome—heterozygous mutation | developmental delay, intellectual disability, coarse facial features, feeding difficulties | [162] |
| BAF60A/SMARCD1 | Coffin-Siris syndrome—heterozygous mutation | developmental delay, intellectual disability, coarse facial features, feeding difficulties | [163] |
| BAF57/SMARCE1 | Meningioma—heterozygous mutation | spinal tumors | [164] |
| Coffin-Siris syndrome—heterozygous mutation | developmental delay, intellectual disability, coarse facial features, feeding difficulties | [155,165] | |
| BAF53B/ACTL6B | Developmental and Epileptic Encephalopathy | delayed global development, hypotonia, peripheral spasticity, mainly cerebral atrophy and delayed myelination | [166,167,168,169] |
| Intellectual Developmental Disorder With Severe Speech And Ambulation Defects—heterozygous mutation | global developmental delay, impaired intellectual, dysmorphic features: prominent forehead and wide mouth | [169] | |
| BAF250A/ARID1A | Coffin-Siris syndrome—heterozygous mutation | developmental delay, intellectual disability, coarse facial features, feeding difficulties | [155,170] |
| BAF250B/ARID1B | Coffin-Siris syndrome—heterozygous mutation | developmental delay, intellectual disability, coarse facial features, feeding difficulties | [155,171,172] |
| BAF200/ARID2 | Coffin-Siris syndrome—heterozygous mutation | developmental delay, intellectual disability, coarse facial features, feeding difficulties | [173,174,175] |
| ACTB | Juvenile-Onset Dystonia—homozygous mutation | eosinophilic, rod-like cytoplasmic inclusions in neocortical and thalamic neurons; abundant eosinophilic spherical structures in the striatum that were strongly actin-and actin depolarizing factor/cofilin-positive | [176,177] |
| Baraitser-Winter syndrome—heterozygous mutation | microcephaly, rare lissencephaly or neuronal heterotopia | [178,179] | |
| BAF45D/DPF2 | Coffin-Siris syndrome—heterozygous mutation | developmental delay, intellectual disability, coarse facial features, feeding difficulties | [180] |
| BCL11A | Intellectual developmental disorder with persistence of fetal hemoglobin—heterozygous mutation | delayed psychomotor development, intellectual disability, microcephaly, downslanting palpebral fissures, strabismus, external ear abnormalities, and asymptomatic persistence of fetal hemoglobin (HbF) | [181] |
| BCL11B | Intellectual Developmental Disorder with Speech Delay, Dysmorphic Facies, and T-Cell Abnormalities—heterozygous mutation | delayed psychomotor development with intellectual disability and speech delay. included autistic features, attention deficit-hyperactivity disorder, anxiety, and other behavioral abnormalities | [182] |
| BAF180/PBRM1 | Autism spectrum disorders | difficulties with social interaction and communication, restricted and repetitive behavior | [183] |
| ISWI complex | |||
| BRF1 | cerebellofaciodental syndrome (CFDS)—homozygous mutation | delayed development, intellectual disability, abnormal facial and dental findings, cerebellar hypoplasia | [184] |
| BPTF | neurodevelopmental disorder with dysmorphic facies and distal limb anomalies (NEDDFL)—heterozygous mutation | delayed psychomotor development and intellectual disability | [185] |
| SNF2L/SMARCA1 | Schizophrenia syndrome disorders | delusions, disorganized thinking, hallucinations | [186] |
| Rett syndrome-like phenotype | Encephalopathy, severe intellectual disability, autistic features, epilepsy | [187] | |
| Coffin-Siris syndrome-like phenotype | developmental delay, intellectual disability, coarse facial features, feeding difficulties | [166] | |
| BAZ1A/ACF1 | Intelectual disability, epilepsy | [188] | |
| CHD protein family | |||
| CHD1 | Pilarowski-Bjornsson syndrome—heterozygous mutation | delayed development, intellectual disability, autistic features, speech apraxia, mild dysmorphic features. | [189] |
| CHD2 | childhood-onset epileptic encephalopathy (EEOC)—heterozygous mutation | severe intellectual disability, epilepsy characterized by onset of multiple seizure | [190,191,192,193] |
| CHD3 | Snijders Blok-Campeau syndrome (SNIBCPS)—heterozygous mutation | global developmental delay, impaired intellectual development. Macrocephaly, prominent forehead and hypertelorism, hypotonia, and joint laxity. | [194] |
| CHD4 | Sifrim-Hitz-Weiss syndrome | intellectual developmental disorder | [195,196] |
| CHD8 | Autism spectrum disorders | difficulties with social interaction and communication, restricted and repetitive behavior | [183] |
| INO80 complex | |||
| YY1 | Gabriele-de Vries syndrome—heterozygous mutation | delayed psychomotor development, variable cognitive impairment, behavioral problems, feeding problems | [197] |
| INO80E | microcephaly, intelectual disability | [198] | |
| SRCAP | Floating-harbor sydrome—heterozygous mutation | short stature, delayed bone age, delayed speech development, facial features | [199] |
| Cohesin complex | |||
| SMC1 | Cornelia de Lange syndrome | intelectual disability, microcephaly | [200,201] |
| Developmental and Epileptic Encephalopathy 85 with or without Midline Brain Defects—heterozygous mutation | severe seizures in the first year of life, global developmental delay, impaired intellectual development, poor/absent speech, dysmorphic facial features. | [202,203,204,205,206] | |
| SMC3 | Cornelia de Lange syndrome—heterozygous mutation | intelectual disability, microcephaly | [207,208] |
| RAD21 | Cornelia de Lange syndrome—heterozygous mutation | Intelectual disability, microcephaly | [202,209,210,211] |
| STAG1 | autosomal dominant mental retardation-47 (MRD47)—heterozygous mutation | delayed psychomotor development, mild to moderate intellectual disability | [212] |
| STAG2 | Mullegama-Klein-Martinez syndrome—heterozygous mutation | impaired intellectual development, speech delay, hypotonia, microcephaly | [213,214,215] |
| X-Linked Holoprosencephaly—heterozygous mutation | incomplete division of the embryonic forebrain | [202] | |
| NIPBL | Cornelia de Lange syndrome | Intelectual disability, microcephaly | [216,217,218] |
| CTCF | mental retardation-21 (MRD21) | mild intellectual disability, short stature, microcephaly | [219] |
Table 2.
Epidrugs in NDs treatment.
| Disease | Epidrug class | Active compound | Model Organism | Remarks | References |
|---|---|---|---|---|---|
| Alzeimer’s disease | HDACi | Sodium Butyrate | CK-p25 Tg mice | Improved learning and memory | [371] |
| APPPS1-21 mice | Increased histone acetylation, upregulation of neuronal plasticity-related genes, improved associative memory | [372] | |||
| Phenylbutyrate | Tg2576 mice | Improved memory, increased dendritic spines density, Increased histone acetylation | [373] | ||
| N171-82Q mice | Increased H3 and h4 acetylation, mice survival, reduces brain gross and neuronal atrophy | [374] | |||
| MS-275 | APPPS1-21 mice | Reduced behavioral impairment, reduced number of amyloid plaques in cortex, enhanced microglia activation in cortex | [375] | ||
| Mercaptoacetamide-based HDACi (W2 and I2) | 3xTg AD mice | Decreased alpha amyloid level in vitro, Alpha amyloid synthesis genes downregulation/degradation genes upregulation, decreased Tau protein phosphorylation, improved learning and memory | [376] | ||
| CM-695 | Tg2576 Mice | Improved memory, Decreased alpha amyloid level | [377,378,379] | ||
| Valproic acid | APP23 mice | Improved memory, Decreased tau and alpha amyloid | [380] | ||
| Clinical trials | ClinicalTrials.gov Identifier: NCT01729598; NCT00088387 | ||||
| Nicotinamide | Triple Knock-in mice: presenilin PS1M146V, Amyloid Precursor protein (APPKM670/671NL), Tau (tauP301L) | Prevented cognitive deficits, decreased phosphorylation of tau protein | [381] | ||
| Clinical trials | ClinicalTrials.gov Identifier: NTC00580931; TC03061474 | ||||
| Vorinostat | Clinical trials | ClinicalTrials.gov Identifier: NCT03056495 | |||
| 4-PBA | Clinical trials | ClinicalTrials.gov Identifier: NCT03533257 | |||
| epigallocatechin-3-gallate (EGCG) | Clinical trials | ClinicalTrials.gov Identifier: NCT00951834 | |||
| Huntington’s disease | HDACi | TSA/SAHA | Hdh-Q109 mice | Increased BDNF transport, improved neuron survival | [382,383] |
| SAHA | R6/2 HD mouse | improved motor skills | [384] | ||
| Sodium Butyrate | R6/2 HD mice | Improved survival of R6/2 HD mice | [385] | ||
| Sodium butyrate/phenylbutyrate | Human Y39C α-Synuclein Transgenic mouse | Increased expression of DJ1 gene, protection against α-Synuclein Induced Toxicity, reduce α-Synuclein level in mice brain, Prevents Age-related Motor and Cognitive Decline | [386] | ||
| Sodium butyrate, SAHA | D.melanogaster Q48 | Decreased neurodegeneration | [387] | ||
| valproic acid | PD patients | Improved myoclonic hyperkinesia | [388] | ||
| HDACi 4b | N171-82Q mice | Decreased Huntingtin aggregation in brain’s cells nuclei, ameliorated cognitive functions | [389] | ||
| MC1568 | SHSY5Y Cell line | Enhanced neurite growth of Dopaminergic and Sympathetic Neurons, Increased histone acetylation, Enhanced protection against MPP+ induced neurotoxicity | [390] | ||
| DMNTi | decitabine, FdCyd | In vitro cultured neurons | Reduced cytotoxicity, Increased BDNF expression | [391] | |
| Parkinson’s Disease | HDACi | Sodium phenylbutyrate | Clinical trials | ClinicalTrials.gov Identifier: NCT02046434 | |
| Valproic acid | Rotenone-treated rats | Protection against neurotoxicity | [392] | ||
| phenylbutyrate | SOD1 (G93A) mice | Increased histone acetylation, increased motoneuron survival | [132,393] | ||
| Amyotrophic lateral sclerosis | HDACi | trichostatin A | myelin oligodendrocyte glycoprotein peptide-treated WT mice | improved neuronal survival and enhance anti-inflammatory pathway action | [394] |
| Vorinostat | myelin oligodendrocyte glycoprotein peptide-treated WT mice | CNS inflammation and demyelination inhibition | [395] | ||
| Multiple sclerosis | HDACi | Sodium butyrate | WT rats | Histone 4 acetylation in promotor region of Transthyretin (Ttr), improved performance in FST, TST tests | [396] |
| MS-275 | WT mice | Increased H3K27 acetylation in accumbens nucleus, decreased expression of stress induced genes | [397] | ||
| Depressive disorders | HDACi | MS-275 | WT mice | Increased CREB and BDNF expression | [398] |
| DNMTi | 5-azaD 5-azaC | WT mice | BDNF expression, improved performance in FST test | [396] |
Abreviations: HDACi-histone deacetylase inhibitors; DNMTi-DNA methyltransferase (DNMT) inhibitors; 5-azaD-5-aza-2-deoxycytidine; 5-azaC-5-azacytidine.
Table 3.
(Epi)genome-targeting for treatment of NDs.
| Disease | (Epi)genome Modification Strategy and Delivery System | Remarks | References |
|---|---|---|---|
| Parkinson ‘s Disease (PD) | CRISPR-dCas9- DNMT3A targeting α-synuclein Lentiviral vector delivery. | PD patient iPS cell-derived neurons. Silencing of SNCA (α-synuclein) leads to a reduction of disease phenotype downmodulation of ROS * production and an increase in cellular viability. | [512] |
| ZF-p65 targeting glial cell line-derived neurotrophic factor (GDNF). AAV2 delivery. | Rat model of PD. Striatal GDNF activation increases neuronal survival. | [513] | |
| Lewy Body Dementia | Combined dCas9-VP64 and sgRNA-MS2-p65 targeting Nrf2 promoter. | In vitro mouse neurons Nrf2 overexpression indued by HDACi and dCas9 ActD helps to clear the toxic deposits of α-synuclein | [514] |
| Alzheimer’s disease (AD) | ZF fused to G9a, Suvdel76, SKD) or VP64 Dlg4/PSD95. | Mouse. Activation of PSD95 leads to memory rescue in aged and Alzheimer’s disease mice. | [515] |
| CRISPR-Cas9 deletion of the mutated allele coding the amyloid precursor protein (APP). | Patient-derived fibroblasts and mouse. Selective silencing of the mutated amyloid precursor protein (APP); 60% reduction in secreted Aβ | [516] | |
| CRISPR-Cas9-mediated truncation of the C-terminus of the APP AAV9–mediated delivery of CRISPR-Cas9 into mouse hippocampus. | iPS cell-derived neurons, cultured hippocampal neurons, mouse. Removal of the C-terminus of APP results in upregulation of the neuroprotective α-cleavage pathway. | [517] | |
| CRISPR-Cas9 nanocomplexes Disruption of Bace1 and Th genes in neurons. | 5XFAD and App knock-in mice mouse. Reduction in Aβ plaque accumulation and Aβ42 secretion. Reduction of Bace1 lead to an increased associative learning and improved spatial working memory and reduced deficit in alternation performance compared with 5XFAD controls. Significant reduction of Aβ42 plaque accumulation and secretion at 8- and 12-weeks post Cas9 injection. | [518] | |
| Huntington‘s Disease | ZFs to block mutant HTT expression AAV2/1 intra-striatal injection. | Cell lines and the R6/2 mouse model. Reduction of diseased clasping behavior, enhanced motor coordination. | [519] |
| KAMI-Cas9 targeting HTT gene lentiviral-mediated in vivo delivery. | Neuronal/glial cultures, striatum of mice, and patient-specific iPSC neuronal cells. Decreased HTT aggregation, reduced neuronal dysfunction both in vitro and in vivo. | [520] | |
| CRISPR-Cas9. rAAV delivery of Cas9. | HD-patient-derived fibroblasts and BacHD mice. Allele-specific removal of mutated HTT gene | [521] | |
| CRISPR-dCas9MeCP2 downregulation of human HTT AAV-mediated delivery. | Adult brain (striatum) of the mouse HD model (HD140Q-KI). Silencing of the HTT led to reduced neuropathology and increased performance in rotarod, balance beam, and grip strength tests. | [522] | |
| ZF-KRAB, AAV-mediated delivery. | Patient-derived fibroblasts and neurons, HD mouse models (intra-striatal injection). Improvement in neuronal functions and alleviation of motoneuron symptoms. | [523] | |
| MeCP2 Duplication syndrome | CRISPR-Cas9.AAV-mediated delivery. | Transgenic mouse expressing human MeCP2. Knockout of the human MeCP2. Reversal of the social aversion deficit, yet no impact on the reduced locomotor activity, the heightened anxiety-like behaviors, and the fear generalization phenotype. | [524] |
| Rett syndrome | CRISPR-Cas9 | HR in human iPS cells to correct disease variants. | [525,526] |
| Fragile X syndrome | CRISPR-dCas9Tet1 demethylation of the FMR1 gene Lentiviral delivery. | iPS cell model of the Fragile X syndrome Demethylation of the FMR1 promoter accompanied by a gain of H3K27ac and transcriptional onset in post mitotic neurons derived from the iPS cells | [527] |
* Abbreviations: ROS—reactive oxygen species; APP—amyloid precursor protein, HTT—huntingtin, HD—Huntington’s disease.
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Janowski, M.; Milewska, M.; Zare, P.; Pękowska, A. Chromatin Alterations in Neurological Disorders and Strategies of (Epi)Genome Rescue. Pharmaceuticals 2021, 14, 765. https://0-doi-org.brum.beds.ac.uk/10.3390/ph14080765
AMA Style
Janowski M, Milewska M, Zare P, Pękowska A. Chromatin Alterations in Neurological Disorders and Strategies of (Epi)Genome Rescue. Pharmaceuticals. 2021; 14(8):765. https://0-doi-org.brum.beds.ac.uk/10.3390/ph14080765
Chicago/Turabian StyleJanowski, Marcin, Małgorzata Milewska, Peyman Zare, and Aleksandra Pękowska. 2021. "Chromatin Alterations in Neurological Disorders and Strategies of (Epi)Genome Rescue" Pharmaceuticals 14, no. 8: 765. https://0-doi-org.brum.beds.ac.uk/10.3390/ph14080765
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.