
Liquid Crystal Nanoparticle Conjugates for Scavenging Reactive Oxygen Species in Live Cells

 , and
, and 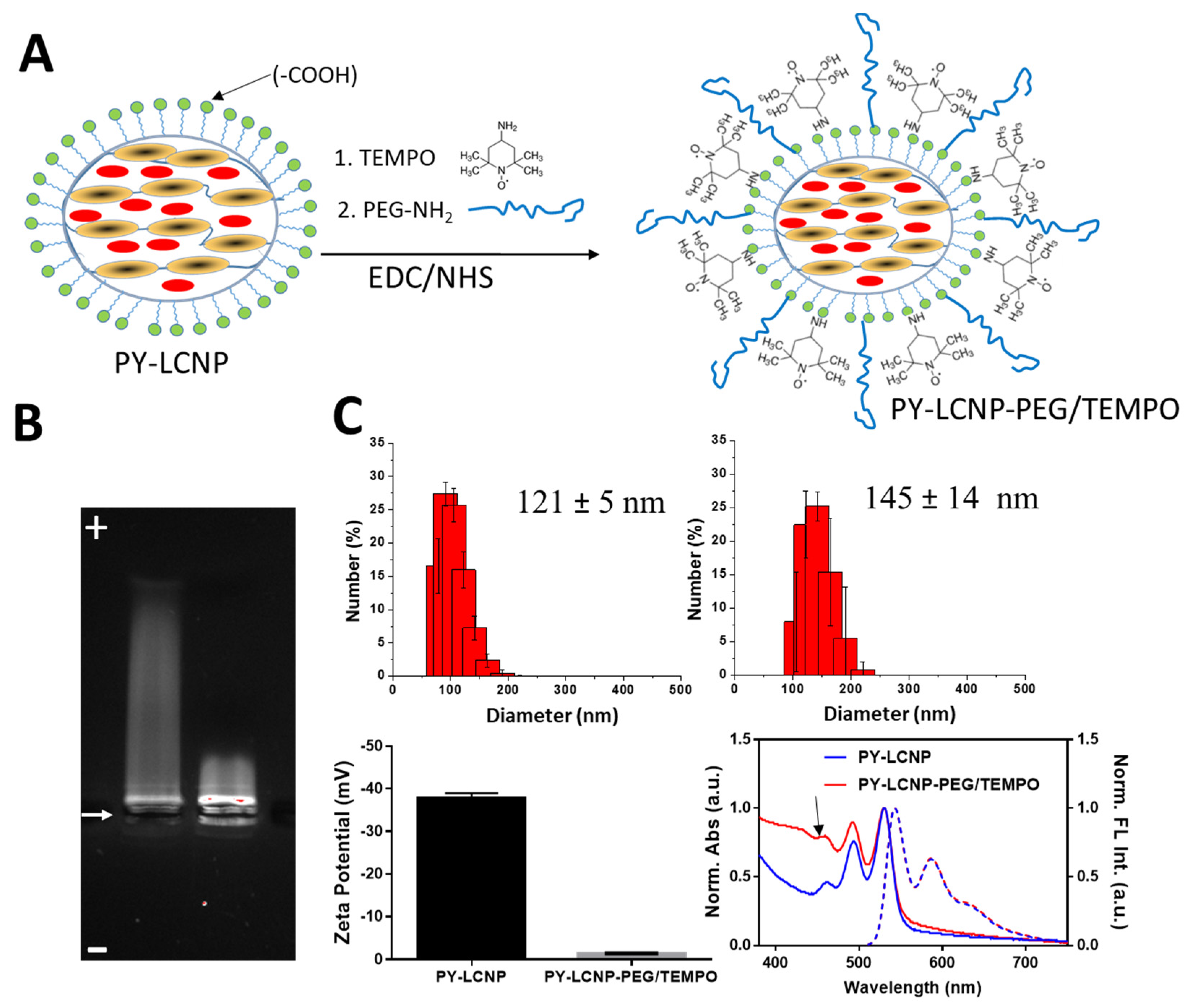{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results and Discussion
2.1. Rationale for the Synthesis of PY-LCNP-PEG/TEMPO
2.2. Synthesis and Characterization of PY-LCNP-PEG/TEMPO
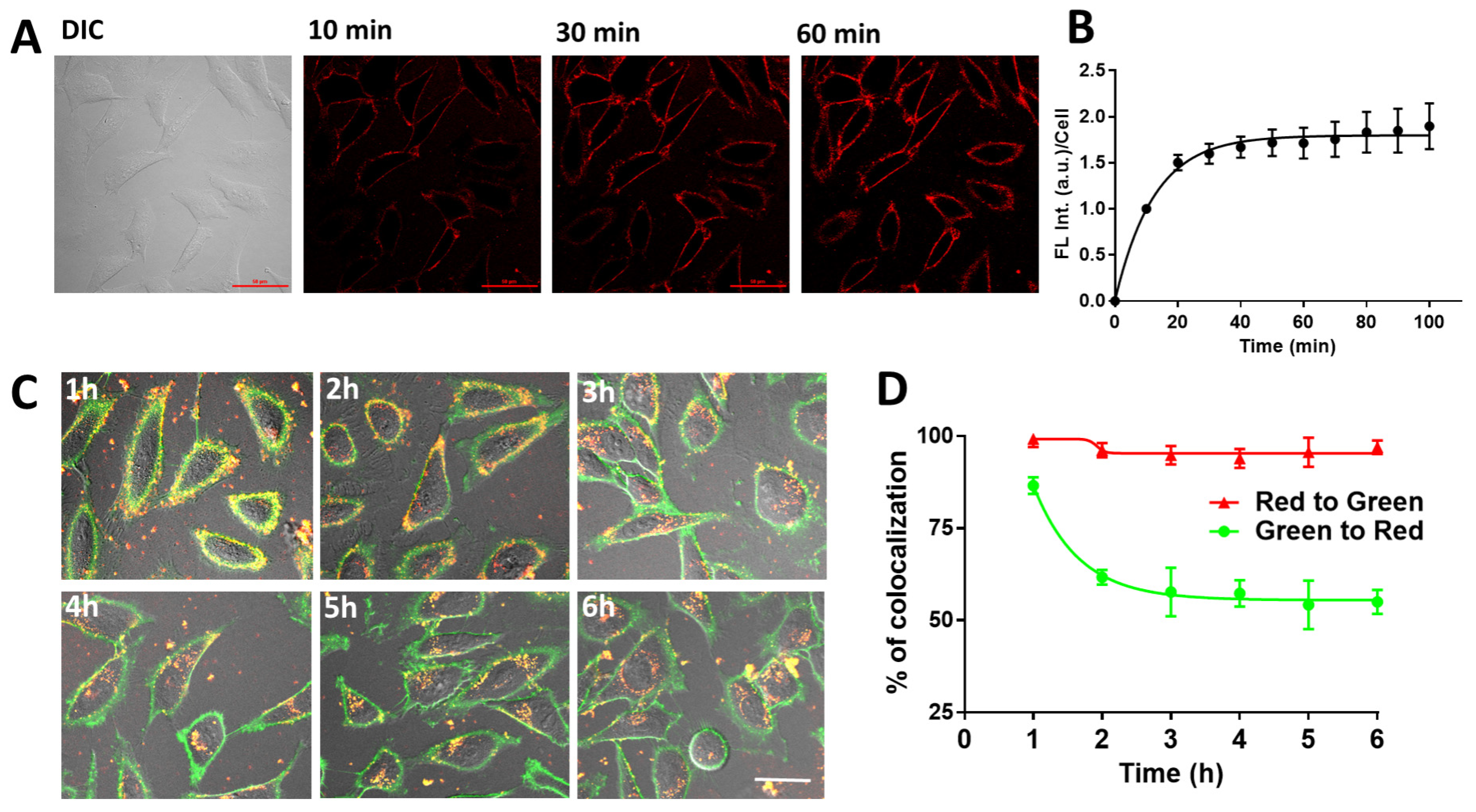
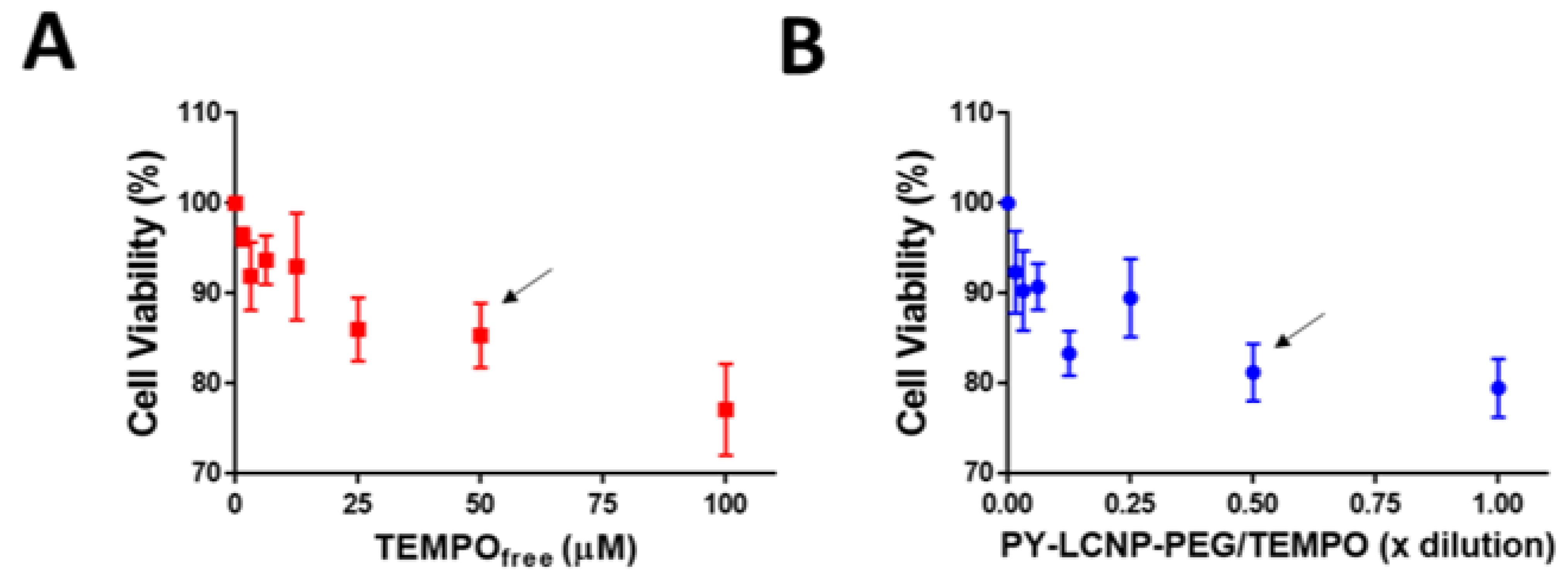
2.3. Cellular Uptake of PY-LCNP-PEG/TEMPO
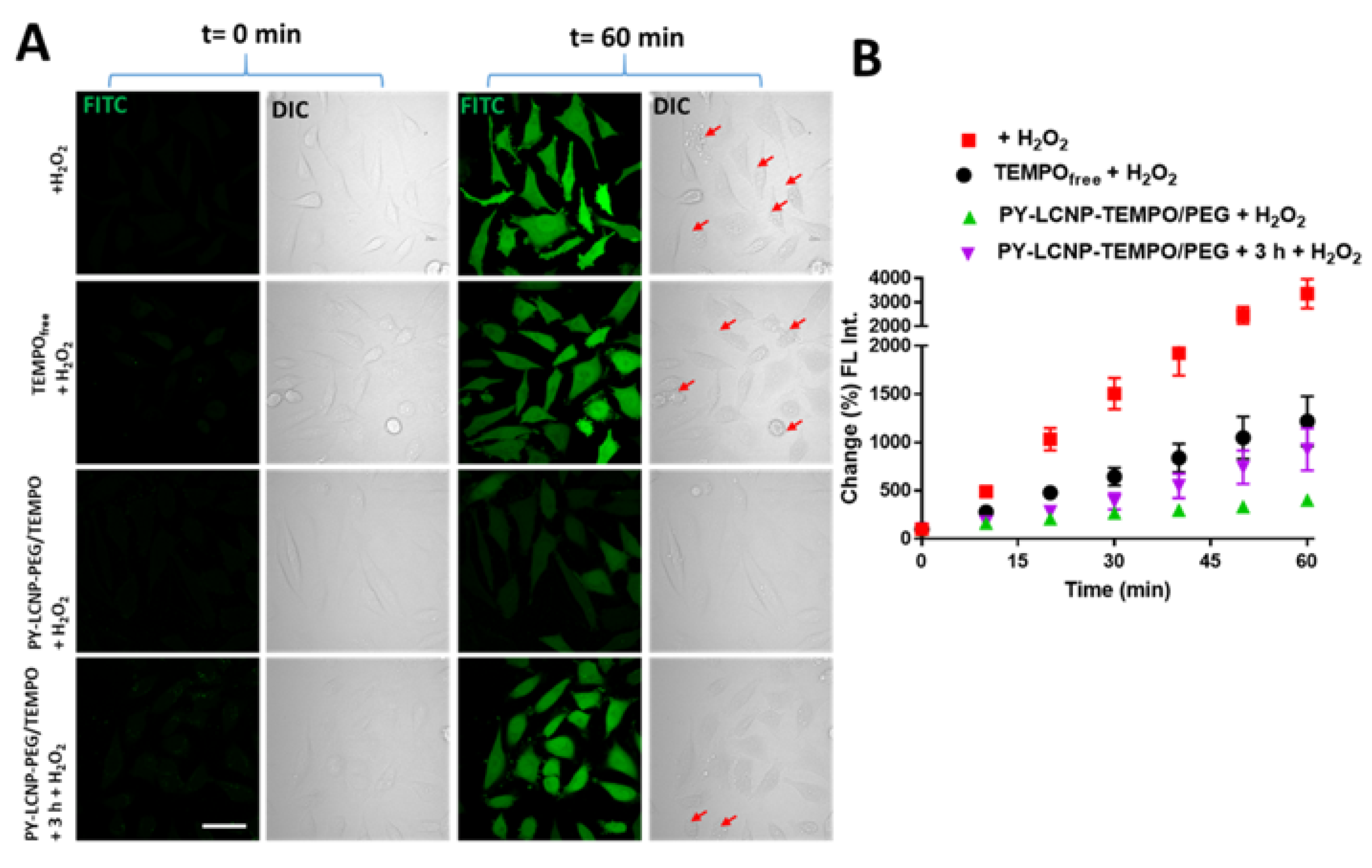
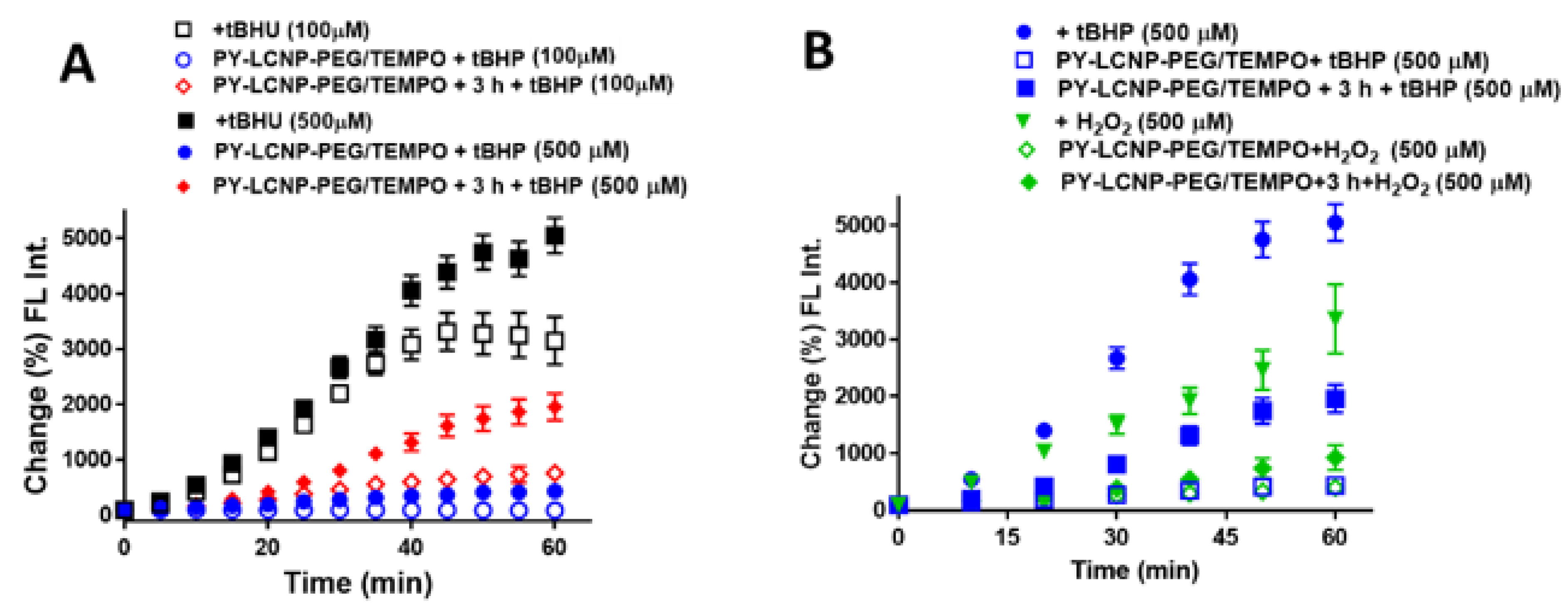
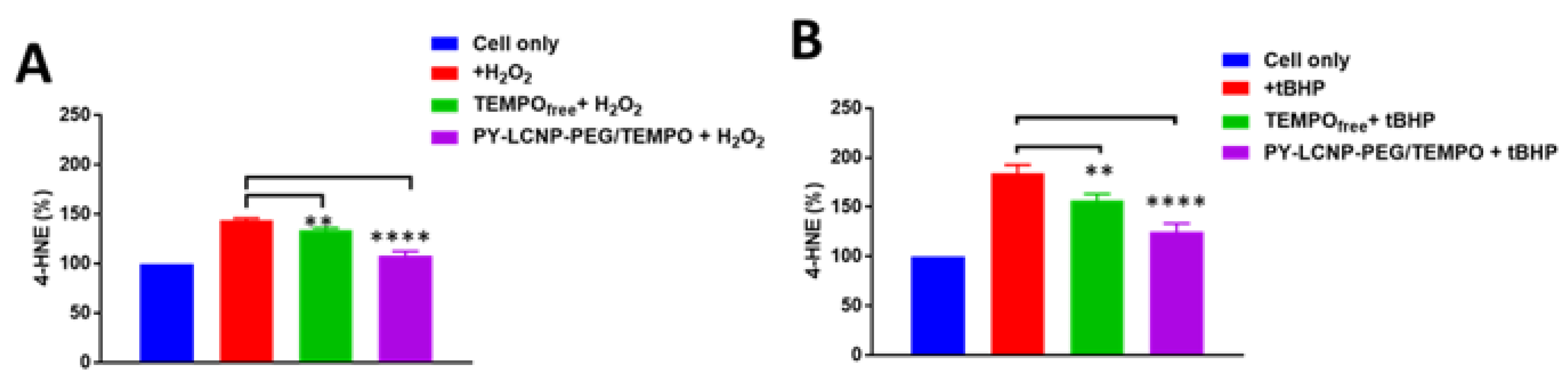
2.4. ROS-Scavenging and Cellular Protection Efficiency of PY-LCNP-PEG/TEMPO
3. Materials and Methods
3.1. Materials
3.2. Synthesis of PY-LCNP-PEG/TEMPO Nanoparticle Conjugates
3.3. Physicochemical Properties of PY-LCNP-PEG/TEMPO
3.4. Interaction of PY-LCNP-PEG/TEMPO Conjugates with HeLa Cells
3.5. Cellular ROS Scavenging and Protection of PY-LCNP-PEG/TEMPO
3.6. Lipid Peroxidation Assay
3.7. Cell Viability Assay
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Zhang, J.; Wang, X.; Vikash, V.; Ye, Q.; Wu, D.; Liu, Y.; Dong, W. ROS and ROS-mediated cellular signaling. Med. Cell. Longev. 2016, 2016, 4350965. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Abdal Dayem, A.; Hossain, M.K.; Lee, S.B.; Kim, K.; Saha, S.K.; Yang, G.-M.; Choi, H.Y.; Cho, S.-G. The role of reactive oxygen species (ROS) in the biological activities of metallic nanoparticles. Int. J. Mol. Sci. 2017, 18, 120. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Karihtala, P.; Soini, Y. Reactive oxygen species and antioxidant mechanisms in human tissues and their relation to malignancies. APMIS 2007, 115, 81–103. [Google Scholar] [CrossRef] [PubMed]
- Schieber, M.; Chandel, N.S. ROS function in redox signaling and oxidative stress. Curr. Biol. 2014, 24, R453–R462. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Avery, S.V. Molecular targets of oxidative stress. Biochemical 2011, 434, 201–210. [Google Scholar] [CrossRef] [Green Version]
- Oberley, T.D. Oxidative damage and cancer. Am. J. Pathol. 2002, 160, 403–408. [Google Scholar] [CrossRef] [Green Version]
- Zhang, C.; Wang, X.; Du, J.; Gu, Z.; Zhao, Y. Reactive oxygen species-regulating strategies based on nanomaterials for disease treatment. Adv. Sci. 2021, 8, 2002797. [Google Scholar] [CrossRef]
- Liou, G.-Y.; Storz, P. Reactive oxygen species in cancer. Free Radic. Res. 2010, 44, 479–496. [Google Scholar] [CrossRef] [Green Version]
- Lewandowski, M.; Gwozdzinski, K. Nitroxides as antioxidants and anticancer drugs. Int. J. Mol. Sci. 2017, 18, 2490. [Google Scholar] [CrossRef] [Green Version]
- Shashni, B.; Nagasaki, Y. Newly developed self-assembling antioxidants as potential therapeutics for the cancers. J. Pers. Med. 2021, 11, 92. [Google Scholar] [CrossRef]
- Radak, Z.; Zhao, Z.; Koltai, E.; Ohno, H.; Atalay, M. Oxygen consumption and usage during physical exercise: The balance between oxidative stress and ros-dependent adaptive signaling. Antioxid. Redox Signal. 2013, 18, 1208–1246. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bjelakovic, G.; Nikolova, D.; Gluud, L.L.; Simonetti, R.G.; Gluud, C. Mortality in randomized trials of antioxidant supplements for primary and secondary prevention: Systematic review and meta-analysis. J. Am. Med. Assoc. 2007, 297, 842–857. [Google Scholar] [CrossRef] [PubMed]
- Sledziński, Z.; Woźniak, M.; Antosiewicz, J.; Lezoche, E.; Familiari, M.; Bertoli, E.; Greci, L.; Brunelli, A.; Mazera, N.; Wajda, Z. Protective effect of 4-hydroxy-tempo, a low molecular weight superoxide dismutase mimic, on free radical toxicity in experimental pancreatitis. Int. J. Pancreatol. 1995, 18, 153–160. [Google Scholar] [CrossRef] [PubMed]
- Griesser, M.; Shah, R.; Van Kessel, A.T.; Zilka, O.; Haidasz, E.A.; Pratt, D.A. The catalytic reaction of nitroxides with peroxyl radicals and its relevance to their cytoprotective properties. J. Am. Chem. Soc. 2018, 140, 3798–3808. [Google Scholar] [CrossRef]
- Yoshitomi, T.; Hirayama, A.; Nagasaki, Y. The ros scavenging and renal protective effects of pH-responsive nitroxide radical-containing nanoparticles. Biomaterials 2011, 32, 8021–8028. [Google Scholar] [CrossRef] [Green Version]
- Yoshitomi, T.; Suzuki, R.; Mamiya, T.; Matsui, H.; Hirayama, A.; Nagasaki, Y. pH-sensitive radical-containing-nanoparticle (RNP) for the l-band-epr imaging of low pH circumstances. Bioconjugate Chem. 2009, 20, 1792–1798. [Google Scholar] [CrossRef]
- DeJulius, C.R.; Dollinger, B.R.; Kavanaugh, T.E.; Dailing, E.; Yu, F.; Gulati, S.; Miskalis, A.; Zhang, C.; Uddin, J.; Dikalov, S.; et al. Optimizing an antioxidant tempo copolymer for reactive oxygen species scavenging and anti-inflammatory effects in vivo. Bioconjugate Chem. 2021, 32, 928–941. [Google Scholar] [CrossRef]
- Shashni, B.; Alshwimi, A.; Minami, K.; Furukawa, T.; Nagasaki, Y. Nitroxide radical-containing nanoparticles as potential candidates for overcoming drug resistance in epidermoid cancers. Polymer 2017, 116, 429–438. [Google Scholar] [CrossRef]
- Zhu, Y.; Matsumura, Y.; Velayutham, M.; Foley, L.M.; Hitchens, T.K.; Wagner, W.R. Reactive oxygen species scavenging with a biodegradable, thermally responsive hydrogel compatible with soft tissue injection. Biomaterials 2018, 177, 98–112. [Google Scholar] [CrossRef]
- Li, J.; Zhang, J.; Chen, Y.; Kawazoe, N.; Chen, G. Tempo-conjugated gold nanoparticles for reactive oxygen species scavenging and regulation of stem cell differentiation. ACS Appl. Mater. Interfaces 2017, 9, 35683–35692. [Google Scholar] [CrossRef]
- Spillmann, C.M.; Naciri, J.; Algar, W.R.; Medintz, I.L.; Delehanty, J.B. Multifunctional liquid crystal nanoparticles for intracellular fluorescent imaging and drug delivery. ACS Nano 2014, 8, 6986–6997. [Google Scholar] [CrossRef] [PubMed]
- Nag, O.K.; Naciri, J.; Oh, E.; Spillmann, C.M.; Delehanty, J.B. Lipid raft-mediated membrane tethering and delivery of hydrophobic cargos from liquid crystal-based nanocarriers. Bioconjugate Chem. 2016, 27, 982–993. [Google Scholar] [CrossRef] [PubMed]
- Nag, O.K.; Naciri, J.; Erickson, J.S.; Oh, E.; Delehanty, J.B. Hybrid liquid crystal nanocarriers for enhanced zinc phthalocyanine-mediated photodynamic therapy. Bioconjugate Chem. 2018, 29, 2701–2714. [Google Scholar] [CrossRef] [PubMed]
- Tonga, G.Y.; Saha, K.; Rotello, V.M. 25th anniversary article: Interfacing nanoparticles and biology: New strategies for biomedicine. Adv. Mater. 2014, 26, 359–370. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nag, O.K.; Delehanty, J.B. Active cellular and subcellular targeting of nanoparticles for drug delivery. Pharmaceutics 2019, 11, 543. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Breger, J.; Delehanty, J.B.; Medintz, I.L. Continuing progress toward controlled intracellular delivery of semiconductor quantum dots. Wiely Interdiscip. Rev. Nanomed. 2015, 7, 131–151. [Google Scholar] [CrossRef]
- Żamojć, K.; Zdrowowicz, M.; Wiczk, W.; Jacewicz, D.; Chmurzyński, L. Dihydroxycoumarins as highly selective fluorescent probes for the fast detection of 4-hydroxy-tempo in aqueous solution. RSC Adv. 2015, 5, 63807–63812. [Google Scholar] [CrossRef]
- Nita, M.; Grzybowski, A. The role of the reactive oxygen species and oxidative stress in the pathomechanism of the age-related ocular diseases and other pathologies of the anterior and posterior eye segments in adults. Oxid. Med. Cell Longev. 2016, 2016, 3164734. [Google Scholar] [CrossRef] [Green Version]
- Alía, M.; Ramos, S.; Mateos, R.; Bravo, L.; Goya, L. Response of the antioxidant defense system to tert-butyl hydroperoxide and hydrogen peroxide in a human hepatoma cell line (HEPG2). J. Biochem. Mol. Toxicol. 2005, 19, 119–128. [Google Scholar] [CrossRef] [Green Version]
- Kučera, O.; Endlicher, R.; Roušar, T.; Lotková, H.; Garnol, T.; Drahota, Z.; Červinková, Z. The effect of tert-butyl hydroperoxide-induced oxidative stress on lean and steatotic rat hepatocytes in vitro. Oxid. Med. Cell. Longev. 2014, 2014, 752506. [Google Scholar] [CrossRef] [Green Version]
- Kwon, H.J.; Kim, D.; Seo, K.; Kim, Y.G.; Han, S.I.; Kang, T.; Soh, M.; Hyeon, T. Ceria nanoparticle systems for selective scavenging of mitochondrial, intracellular, and extracellular reactive oxygen species in parkinson’s disease. Angew. Chem. Int. Ed. 2018, 57, 9408–9412. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Liu, Q.; Xu, Y.; Zhang, Y.; Lv, Y.; Tan, Y.; Jiang, N.; Cao, G.; Ma, X.; Wang, J.; et al. Ginsenoside Rg1 protects against oxidative stress-induced neuronal apoptosis through myosin iia-actin related cytoskeletal reorganization. Int. J. Biol. Sci. 2016, 12, 1341–1356. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Reddan, J.; Sevilla, M.; Giblin, F.; Padgaonkar, V.; Dziedzic, D.; Leverenz, V. T Tempol and deferoxamine protect cultured rabbit lens epithelial cells from H2O2 insult: Insight into the mechanism of H2O2-induced injury. Lens Eye Toxic. Res. 1992, 9, 385–393. [Google Scholar] [PubMed]
- Rodríguez-García, A.; García-Vicente, R.; Morales, M.L.; Ortiz-Ruiz, A.; Martínez-López, J.; Linares, M. Protein carbonylation and lipid peroxidation in hematological malignancies. Antioxidants 2020, 9, 1212. [Google Scholar] [CrossRef]
- Su, L.-J.; Zhang, J.-H.; Gomez, H.; Murugan, R.; Hong, X.; Xu, D.; Jiang, F.; Peng, Z.-Y. Reactive oxygen species-induced lipid peroxidation in apoptosis, autophagy, and ferroptosis. Oxid. Med. Cell. Longev. 2019, 2019, 5080843. [Google Scholar] [CrossRef] [Green Version]
- Ayala, A.; Muñoz, M.F.; Argüelles, S. Lipid peroxidation: Production, metabolism, and signaling mechanisms of malondialdehyde and 4-hydroxy-2-nonenal. Oxid. Med. Cell. Longev. 2014, 2014, 360438. [Google Scholar] [CrossRef]
- Spillmann, C.M.; Naciri, J.; Wahl, K.J.; Garner, Y.H.; Chen, M.-S.; Ratna, B.R. Role of surfactant in the stability of liquid crystal-based nanocolloids. Langmuir 2009, 25, 2419–2426. [Google Scholar] [CrossRef]
- Spillmann, C.M.; Naciri, J.; Anderson, G.P.; Chen, M.S.; Ratna, B.R. Spectral tuning of organic nanocolloids by controlled molecular interactions. ACS Nano 2009, 3, 3214–3220. [Google Scholar] [CrossRef]






Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Nag, O.K.; Naciri, J.; Lee, K.; Oh, E.; Almeida, B.; Delehanty, J.B. Liquid Crystal Nanoparticle Conjugates for Scavenging Reactive Oxygen Species in Live Cells. Pharmaceuticals 2022, 15, 604. https://0-doi-org.brum.beds.ac.uk/10.3390/ph15050604
Nag OK, Naciri J, Lee K, Oh E, Almeida B, Delehanty JB. Liquid Crystal Nanoparticle Conjugates for Scavenging Reactive Oxygen Species in Live Cells. Pharmaceuticals. 2022; 15(5):604. https://0-doi-org.brum.beds.ac.uk/10.3390/ph15050604
Chicago/Turabian StyleNag, Okhil K., Jawad Naciri, Kwahun Lee, Eunkeu Oh, Bethany Almeida, and James B. Delehanty. 2022. "Liquid Crystal Nanoparticle Conjugates for Scavenging Reactive Oxygen Species in Live Cells" Pharmaceuticals 15, no. 5: 604. https://0-doi-org.brum.beds.ac.uk/10.3390/ph15050604