
A Long Journey of CICA-17 Quinoa Variety to Salinity Conditions in Egypt: Mineral Concentration in the Seeds


 ,
,  , , , , , , ,
, , , , , , ,
Abstract
:1. Introduction
2. Results
2.1. Soil and Irrigation Water Analysis
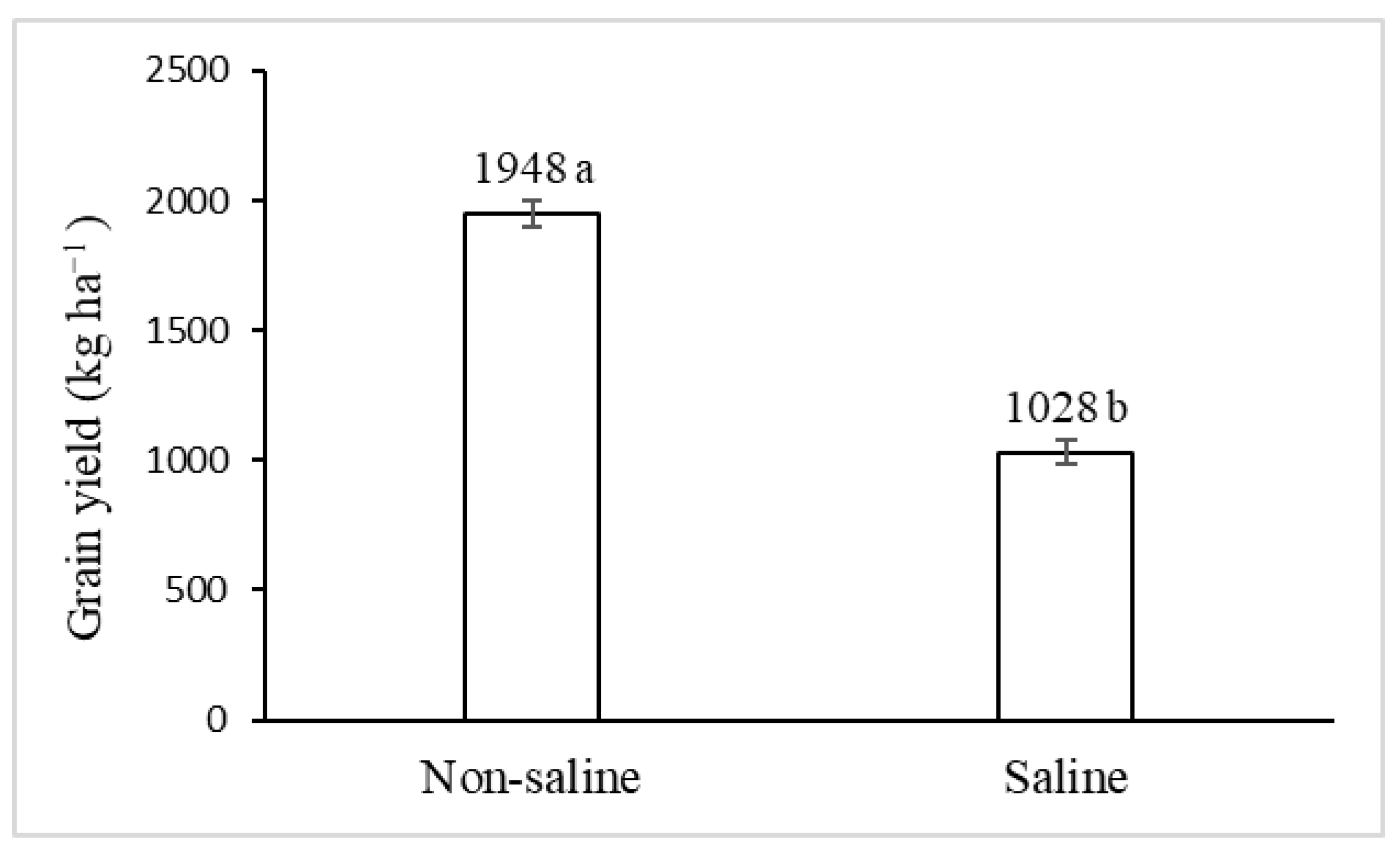
2.2. Grain Yield
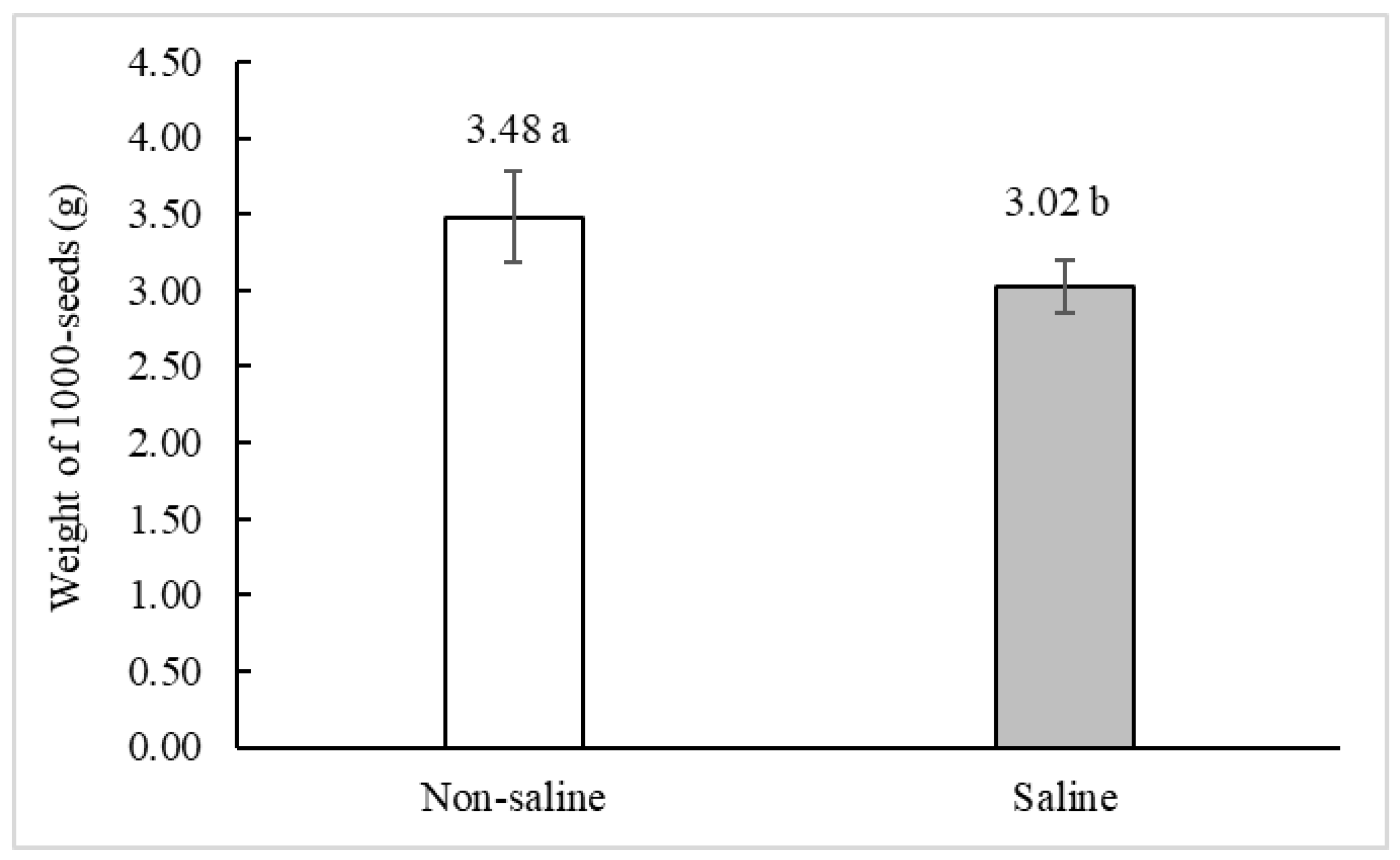
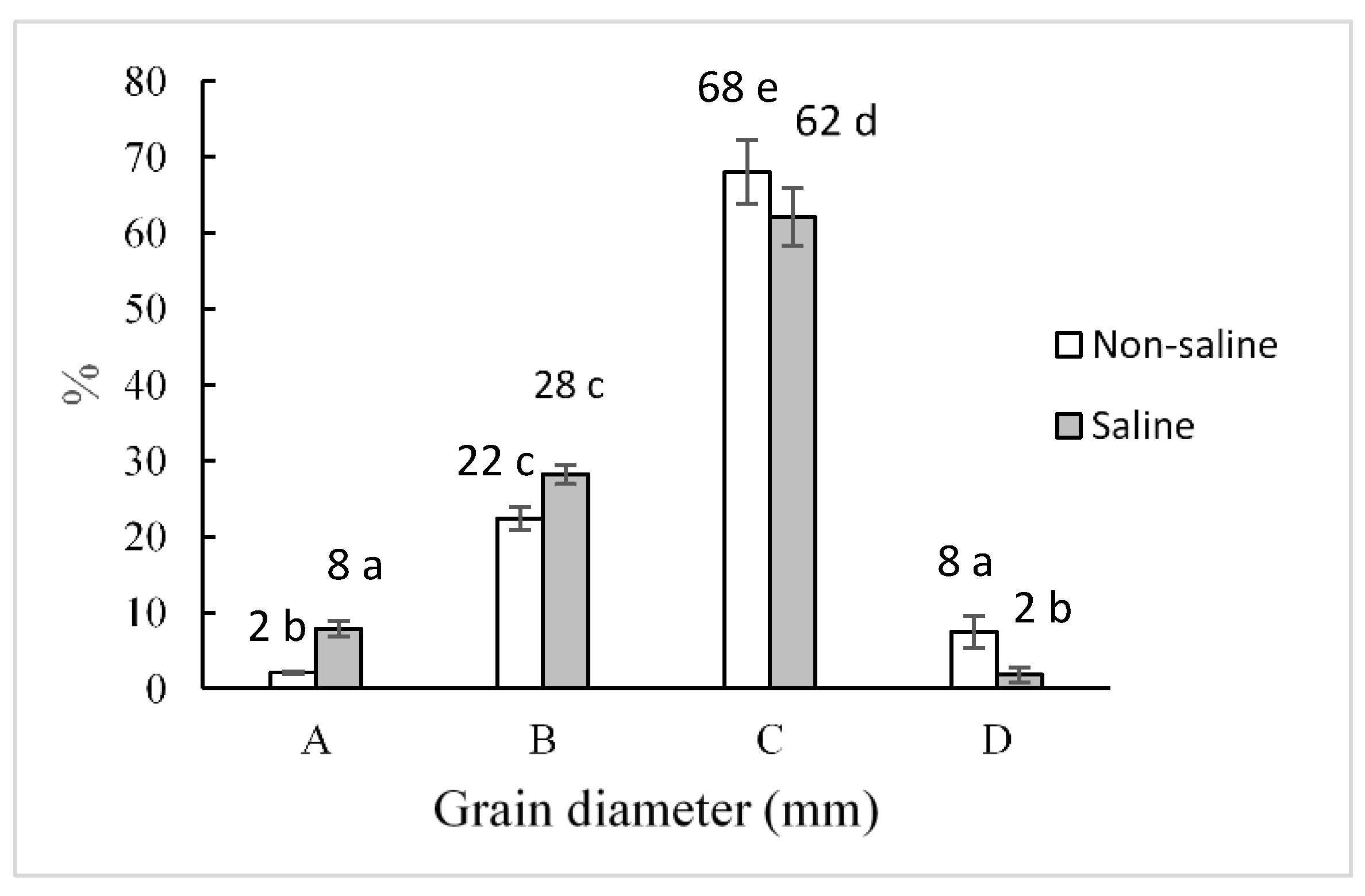
2.3. Grain Weight and Sizes
2.4. Mineral Content in Quinoa Grains
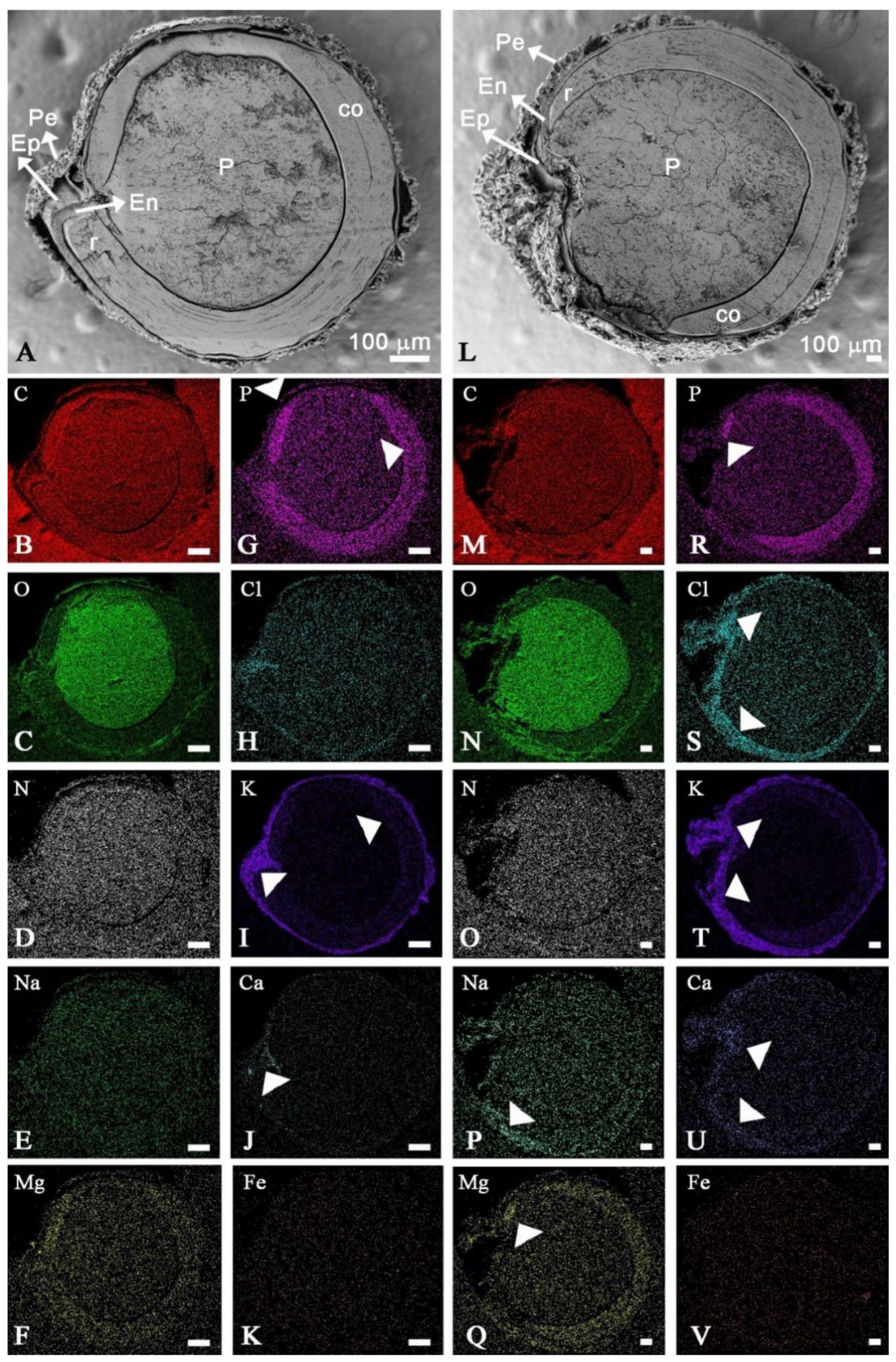
2.5. Mineral Spatial Distribution on Quinoa Grains
3. Discussion
4. Materials and Methods
4.1. Plant Material, Site Description, and Experimental Design
4.2. Yield Components
4.3. Mineral Content
4.4. Mineral Spatial Distribution
4.5. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- IPCC. Climate Change 2014—Impacts, Adaptation, and Vulnerability: Regional Aspects; Cambridge University Press: Cambridge, UK; New York, NY, USA, 2014; ISBN 978-1-107-05816-3. [Google Scholar]
- Rasool, S.; Hameed, A.; Azooz, M.M.; Muneeb-u-Rehman; Siddiqi, T.O.; Ahmad, P. Salt stress: Causes, types and responses of plants. In Ecophysiology and Responses of Plants under Salt Stress; Ahmad, P., Azooz, M.M., Prasad, M.N.V., Eds.; Springer: New York, NY, USA, 2013; pp. 1–24. ISBN 978-1-4614-4746-73. [Google Scholar]
- Tester, M.; Davenport, R. Na+ tolerance and Na+ transport in higher plants. Ann. Bot. 2003, 91, 503–527. [Google Scholar] [CrossRef] [PubMed]
- Dillehay, T.D.; Rossen, J.; Andres, T.C.; Williams, D.E. Preceramic adoption of peanut, squash, and cotton in Northern Peru. Science 2007, 316, 1890–1893. [Google Scholar] [CrossRef] [Green Version]
- Verena, I.A.; Alanoca, C.; Adriana, A.; Ricardo, A.; Alberto, J.A.; Fabiana, A.; Genaro, A.; Pilar, B.; Andrea, B.; Flora, B.; et al. State of the Art Report on Quinoa around the World 2013; Bazile, D., Bertero, H.D., Nieto, C., Eds.; FAO: Santiago, Chile; CIRAD: Montpellier, France, 2015; pp. 1–7. [Google Scholar]
- Fuentes, F.; Bazile, D.; Bhargava, A.; Martinez, E.A. Implications of farmers’ seed exchanges for on-farm conservation of quinoa, as revealed by its genetic diversity in Chile. J. Agric. Sci. 2012, 150, 702–716. [Google Scholar] [CrossRef] [Green Version]
- González, J.A.; Bruno, M.; Valoy, M.; Prado, F.E. Genotypic variation of gas exchange parameters and leaf stable carbon and nitrogen isotopes in ten quinoa cultivars grown under drought. J. Agron. Crop Sci. 2010, 197, 81–93. [Google Scholar] [CrossRef]
- Hinojosa, L.; Matanguihan, J.B.; Murphy, K.M. Effect of high temperature on pollen morphology, plant growth and seed yield in quinoa (Chenopodium quinoa Willd.). J. Agron. Crop Sci. 2018, 205, 33–45. [Google Scholar] [CrossRef] [Green Version]
- González, J.A.; Eisa, S.S.S.; Hussin, S.A.E.S.; Prado, F.E. Quinoa: An incan crop to face global changes in agriculture. In Quinoa: Improvement and Sustainable Production; Murphy, K., Matanguihan, J., Eds.; John Wiley & Sons, Inc.: Hoboken, NJ, USA, 2015; pp. 1–18. ISBN 978-1-118-62804-1. [Google Scholar]
- Vilcacundo, R.; Hernández-Ledesma, B. Nutritional and biological value of quinoa (Chenopodium quinoa Willd.). Curr. Opin. Food Sci. 2017, 14, 1–6. [Google Scholar] [CrossRef] [Green Version]
- Nowak, V.; Du, J.; Charrondière, U.R. Assessment of the nutritional composition of quinoa (Chenopodium quinoa Willd.). Food Chem. 2016, 193, 47–54. [Google Scholar] [CrossRef]
- García-Parra, M.A.; Roa-Acosta, D.F.; Stechauner-Rohringer, R.; García-Molano, F.; Bazile, D.; Plazas Leguizamón, N. Effect of temperature on the growth and development of quinoa plants (Chenopodium quinoa Willd.): A review on a global scale. Sylwan 2020, 164, 411–423. [Google Scholar]
- González, J.A.; Rosa, M.; Parrado, M.F.; Hilal, M.; Prado, F.E. Morphological and physiological responses of two varieties of a highland species (Chenopodium quinoa Willd.) growing under near-ambient and strongly reduced solar UV–B in a lowland location. J. Photochem. Photobiol. Biol. 2009, 96, 144–151. [Google Scholar] [CrossRef]
- Hinojosa, L.; González, J.A.; Barrios-Masias, F.H.; Fuentes, F.; Murphy, K.M. Quinoa abiotic stress responses: A review. Plants 2018, 7, 106. [Google Scholar] [CrossRef] [Green Version]
- Eisa, S.; Hussin, S.; Geissler, N.; Koyro, H.W. Effect of NaCl salinity on water relations, photosynthesis and chemical composition of Quinoa (Chenopodium quinoa Willd.) as a potential cash crop halophyte. Aust. J. Crop Sci. 2012, 6, 357–368. [Google Scholar]
- Eisa, S.S.; Eid, M.; El-Samad, E.H.A.; Hussin, S.; Abdel-Ati, A.; El-Bordeny, N.; Ali, S.; Al-Sayed, H.M.; Lotfy, M.; Masoud, A. Chenopodium quinoa Willd. A new cash crop halophyte for saline regions of Egypt. Aust. J. Crop Sci. 2017, 11, 343–351. [Google Scholar] [CrossRef]
- Orsini, F.; Accorsi, M.; Gianquinto, G.; Dinelli, G.; Antognoni, F.; Carrasco, K.B.R.; Martinez, E.A.; Alnayef, M.; Marotti, I.; Bosi, S.; et al. Beyond the ionic and osmotic response to salinity in Chenopodium quinoa: Functional elements of successful halophytism. Funct. Plant Biol. 2011, 38, 818–831. [Google Scholar] [CrossRef] [PubMed]
- Rojas, W.; Pinto, M.; Alanoca, C.; Gómez Pando, L.; León-Lobos, P.; Alercia, A.; Diulgheroff, S.; Padulosi, S.; Bazile, D. Quinoa genetic resources and ex situ conservation. In State of the Art Report on Quinoa around the World in 2013; Bazile, D., Bertero, H.D., Nieto, C., Eds.; FAO & CIRAD: Rome, Italy, 2015; pp. 56–82. [Google Scholar]
- Bazile, D.; Jacobsen, S.-E.; Verniau, A. The global expansion of quinoa: Trends and limits. Front. Plant Sci. 2016, 7, 622. [Google Scholar] [CrossRef] [Green Version]
- Ruiz, K.B.; Biondi, S.; Oses, R.; Acuña-Rodríguez, I.S.; Antognoni, F.; Martinez-Mosqueira, E.A.; Coulibaly, A.; Canahua-Murillo, A.; Pinto, M.; Zurita, A.; et al. Quinoa biodiversity and sustainability for food security under climate change. A review. Agron. Sustain. Dev. 2013, 34, 349–359. [Google Scholar] [CrossRef] [Green Version]
- Tapia, M. The Long Journey of Quinoa: Who wrote its history. In State of the Art Report on Quinoa around the World in 2013; Bazile, D., Bertero, H.D., Nieto, C., Eds.; FAO and CIRAD: Rome, Italy, 2015; Volume 1, pp. 1–7. ISBN 978-92-5-108558-5. [Google Scholar]
- Tang, Y.; Li, X.; Chen, P.X.; Zhang, B.; Hernandez, M.; Zhang, H.; Marcone, M.F.; Liu, R.; Tsao, R. Lipids, tocopherols, and carotenoids in leaves of amaranth and quinoa cultivars and a new approach to overall evaluation of nutritional quality traits. J. Agric. Food Chem. 2014, 62, 12610–12619. [Google Scholar] [CrossRef]
- Stuardo, M.; Martín, R.S. Antifungal properties of quinoa (Chenopodium quinoa Willd) alkali treated saponins against botrytis cinerea. Ind. Crop. Prod. 2008, 27, 296–302. [Google Scholar] [CrossRef]
- Bhargava, A.; Shukla, S.; Srivastava, J.; Singh, N.; Ohri, D. Genetic diversity for mineral accumulation in the foliage of Chenopodium spp. Sci. Hortic. 2008, 118, 338–346. [Google Scholar] [CrossRef]
- Jacobsen, S.-E. The scope for adaptation of quinoa in northern latitudes of Europe. J. Agron. Crop Sci. 2017, 203, 603–613. [Google Scholar] [CrossRef]
- Peterson, A.; Murphy, K.M. Quinoa cultivation for temperate North America: Considerations and areas for investigation. In Quinoa: Improvement and Sustainable Production; Murphy, K., Matanguihan, J., Eds.; John Wiley & Sons, Inc.: Hoboken, NJ, USA, 2015; pp. 173–192. ISBN 978-1-118-62804-1. [Google Scholar]
- Hinojosa, L.; Kumar, N.; Gill, K.S.; Murphy, K.M. Spectral reflectance indices and physiological parameters in quinoa under contrasting irrigation regimes. Crop. Sci. 2019, 59, 1927–1944. [Google Scholar] [CrossRef] [Green Version]
- Bhargava, A.; Shukla, S.; Ohri, D. Chenopodium quinoa—An Indian perspective. Ind. Crop. Prod. 2006, 23, 73–87. [Google Scholar] [CrossRef]
- Pulvento, C.; Riccardi, M.; Lavini, A.; Iafelice, G.; Marconi, E.; d’Andria, R. Yield and quality characteristics of quinoa grown in open field under different saline and non-saline irrigation regimes. J. Agron. Crop. Sci. 2012, 198, 254–263. [Google Scholar] [CrossRef]
- Mujica, A.; Jacobsen, S.E.; Izquierdo, J.; Marathee, J. Resultados de la Prueba Americana y Europea de la Quinoa. FAO, UNA-Puno, CIP. 2001. Available online: http://www.fao.org/tempref/GI/Reserved/FTP_FaoRlc/old/prior/segalim/prodalim/prodveg/cdrom/contenido/libro03/home03.htm (accessed on 20 November 2020).
- González, J.A.; Konishi, Y.; Bruno, M.; Valoy, M.; Prado, F.E. Interrelationships among seed yield, total protein and amino acid composition of ten quinoa (Chenopodium quinoa) cultivars from two different agroecological regions. J. Sci. Food Agric. 2011, 92, 1222–1229. [Google Scholar] [CrossRef] [PubMed]
- Al-Naggar, A.M.M.; El-Salam, R.M.A.; Badran, A.E.E.; El-Moghazi, M.M.A. Genotype and drought effects on morphological, physiological and yield traits of quinoa (Chenopodium quinoa Willd.). Asian J. Adv. Agric. Res. 2017, 3, 1–15. [Google Scholar] [CrossRef]
- Eisa, S.; Abd El-Samad, E.H.; Hussin, S.; Ali, E.A.; Ebrahim, M.; González, J.A.; Ordano, M.; Erazzú, L.E.; El-Bordeny, N.E.; Abdel-Ati, A.A. Quinoa in Egypt-Plant Density Effects on Seed Yield and Nutritional Quality under Marginal Regions. Middle East J. Appl. Sci. 2018, 8, 512–522. [Google Scholar]
- Ebrahim, M.; Abdel-Ati, A.A.; Hussin, S.A.; Ali, S.H.; Eisa, S.S. Evaluation of some Chenopodium quinoa cultivars under saline soil conditions in Egypt. Arab. Univ. J. Agric. Sci. 2018, 26, 337–347. [Google Scholar] [CrossRef]
- Adolf, V.I.; Jacobsen, E.; Shabala, S. Salt tolerance mechanisms in quinoa (Chenopodium quinoa Willd.). Environ. Exp. Bot. 2013, 92, 43–54. [Google Scholar] [CrossRef]
- Konishi, Y.; Hirano, S.; Tsuboi, H.; Wada, M. Distribution of minerals in quinoa (Chenopodium quinoa Willd) seeds. Biosci. Biotechnol. Biochem. 2004, 68, 231–234. [Google Scholar] [CrossRef] [Green Version]
- Prado, F.E.; Fernández-Turiel, J.L.; Tsarouchi, M.; Psaras, G.K.; González, J.A. Variation of seed mineral concentrations in seven quinoa cultivars grown in two agroecological sites. Cereal Chem. J. 2014, 91, 453–459. [Google Scholar] [CrossRef] [Green Version]
- Ruiz, K.B.; Biondi, S.; Martínez, E.A.; Orsini, F.; Antognoni, F.; Jacobsen, S.-E. Quinoa—A model crop for understanding salt-tolerance mechanisms in halophytes. Plant Biosyst.—Int. J. Deal. Asp. Plant Biol. 2016, 150, 357–371. [Google Scholar] [CrossRef]
- Miranda-Apodaca, J.; Agirresarobe, A.; Martínez-Goñi, X.; Yoldi-Achalandabaso, A.; Pérez-López, U. N metabolism performance in Chenopodium quinoa subjected to drought or salt stress conditions. Plant Physiol. Biochem. 2020, 155, 725–734. [Google Scholar] [CrossRef]
- Koyro, H.-W.; Eisa, S.S. Effect of salinity on composition, viability and germination of seeds of Chenopodium quinoa Willd. Plant Soil 2008, 302, 79–90. [Google Scholar] [CrossRef]
- Hussain, M.I.; Dakheel, A.J.; Reigosa, M.J. Genotypic differences in agro-physiological, biochemical and isotopic responses to salinity stress in quinoa (Chenopodium quinoa Willd.) plants: Prospects for salinity tolerance and yield stability. Plant Physiol. Biochem. 2018, 129, 411–420. [Google Scholar] [CrossRef]
- Karyotis, T.; Iliadis, C.; Noulas, C.; Mitsibonas, T. Preliminary research on seed production and nutrient content for certain quinoa varieties in a saline-sodic soil. J. Agron. Crop Sci. 2003, 189, 402–408. [Google Scholar] [CrossRef]
- Kuljanabhagavad, T.; Thongphasuk, P.; Chamulitrat, W.; Wink, M. Triterpene saponins from Chenopodium quinoa Willd. Phytochemistry 2008, 69, 1919–1926. [Google Scholar] [CrossRef]
- Ando, H.; Chen, Y.-C.; Tang, H.; Shimizu, M.; Watanabe, K.; Mitsunaga, T. Food components in fractions of Quinoa Seed. Food Sci. Technol. Res. 2002, 8, 80–84. [Google Scholar] [CrossRef] [Green Version]
- González, J.A.; Lizárraga, E.F.; Erazzú, L.E.; Martínez-Calsina, L.; Buedo, S.E.; González, D.A. Goat manure fertilization effect on saponin and protein content in quinoa (Chenopodium quinoa Willd) grain of different origin. Middle East J. Agric. Res. 2020, 9, 434–443. [Google Scholar] [CrossRef]
- Bacigalupo, A.; Tapia, M. Potencial agroindustrial de los cultivos andinos subexplotados. In Cultivos andinos subexplotados y su aporte a la alimentación; Tapia, M., Ediciones Gegra, S.A., Eds.; FAO: Santiago, Chile, 1990; pp. 136–163. [Google Scholar]
- Vega-Gálvez, A.; Miranda, M.; Vergara, J.; Uribe, E.; Puente, L.; Martínez, E.A. Nutrition facts and functional potential of quinoa (Chenopodium quinoa Willd.), an ancient Andean grain: A review. J. Sci. Food Agric. 2010, 90, 2541–2547. [Google Scholar] [CrossRef]
- Isobe, K.; Nakajima, E.; Morita, N.; Kawakura, S.; Higo, M. Effects of NaCl on growth and cesium absorption in quinoa (Chenopodium quinoa Willd.). Water Air Soil Pollut. 2019, 230, 66. [Google Scholar] [CrossRef] [Green Version]
- Nielsen, F.H. Possibly essential trace elements. Nutrition and Health. In Clinical Nutrition of the Essential Trace Elements and Minerals: The Guide for Health Professionals; Bogden, J.D., Klevay, L.M., Eds.; Humana Press: Totowa, NJ, USA, 2000; pp. 11–36. ISBN 978-1-59259-040-7. [Google Scholar]
- Nielsen, F. Ultratrace elements. In Encyclopedia of Human Nutrition; Sadler, M.J., Strain, J.J., Caballero, B., Eds.; Academic Press: San Diego, CA, USA, 1999; pp. 1884–1887. [Google Scholar]
- Cefalu, W.T.; Hu, F.B. Role of chromium in human health and in diabetes. Diabetes Care 2004, 27, 2741–2751. [Google Scholar] [CrossRef] [Green Version]
- Chiu, C.-T.; Chuang, D.-M. Neuroprotective action of lithium in disorders of the central nervous system. Zhong Nan Da Xue Xue Bao Yi Xue Ban 2011, 36, 461–476. [Google Scholar] [CrossRef] [PubMed]
- Coelho, I.; Gueifão, S.; Nascimento, A.C.; Lobo, M.; Gimenez, G.; Samman, N.; Castanheira, I. Proximate and mineral profile of quinoa (Chenopodium quinoa), and kiwicha (Amaranthus caudatus) consumed in north of Argentina. Int. Food Data Conf. 2011, 9, 14–17. [Google Scholar]
- Chappell, H.F.; Jugdaohsingh, R.; Powell, J.J. Physiological silicon incorporation into bone mineral requires orthosilicic acid metabolism to SiO44−. J. R. Soc. Interface 2020, 17, 20200145. [Google Scholar] [CrossRef] [PubMed]
- Amaral, A.F.S.; Porta, M.; Silverman, D.T.; Milne, R.L.; Kogevinas, M.; Rothman, N.; Cantor, K.P.; Jackson, B.P.; Pumarega, J.A.; López, T.; et al. Pancreatic cancer risk and levels of trace elements. Gut 2012, 61, 1583–1588. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Uthus, E.O. Arsenic essentiality and factors affecting its importance. In Arsenic Exposure and Health; Chappell, W.R., Abernathy, C.O., Cothern, C.R., Eds.; Science and Technology Letters: Northwood, UK, 1994; pp. 199–208. [Google Scholar]
- Martin, R.B. The chemistry of aluminum as related to biology and medicine. Clin. Chem. 1986, 32, 1797–1806. [Google Scholar] [CrossRef]
- Thomas, E.C.; Lavkulich, L.M. Community considerations for quinoa production in the urban environment. Can. J. Plant Sci. 2014, 95, 397–404. [Google Scholar] [CrossRef] [Green Version]
- Page, A.L. Methods of Soil Analysis: Chemical and Microbiological Properties; American Society of Agronomy: Madison, WI, USA, 1982; ISBN 0-89118-072-9. [Google Scholar]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Physical Properties | ||||||||||
| depth Soil cm | pH | Sp 1 % | EC 2 dS m−1 | DB 3 g cm−3 | OM 4 % | CO3Ca % | Sand % | Silt % | Clay % | Textural class |
| 0–60 (non-saline) | 8.0 | 66 | 2.1 | 1.51 | 1.25 | 2.8 | 30.0 | 58.0 | 12.0 | silt loam |
| 0–60 (saline) | 8.2 | 75 | 26.0 | 1.92 | 0.58 | 0.3 | 33.3 | 53.2 | 3.5 | silt loam |
| Chemical Properties | ||||||||||
| Na | K | Ca | Mg | Cl | SO42 | HCO−3 | CO3−2 | Fe | Mn | |
| meq L−1 | ||||||||||
| 0–60 (non-saline) | 7.8 | 0.6 | 11.7 | 4.3 | 6.5 | 13.5 | 4.45 | 0 | 4.36 | 5.29 |
| 0–60 (saline) | 187.5 | 6.0 | 47.0 | 76.8 | 280.0 | 35 | 1.26 | 0 | 5.78 | 2.44 |
| Location | EC | pH | Na+ | K+ | Ca2+ | Mg2+ | Cl− | HCO−3 | CO3−2 | SO4−2 |
|---|---|---|---|---|---|---|---|---|---|---|
| dS m−1 | meq L−1 | |||||||||
| Non-saline | 0.43 | 7.16 | 1.04 | 0.21 | 1.40 | 1.22 | 1.02 | 0.62 | 0.00 | 1.29 |
| Saline | 1.70 | 7.23 | 6.37 | 0.35 | 3.40 | 4.50 | 8.50 | 4.60 | 0.00 | 6.12 |
| Element | Non Saline Soil | Saline Soil | Difference (%) | Element | Non Saline Soil | Saline Soil | Difference (%) |
|---|---|---|---|---|---|---|---|
| mg kg−1 dry weight | mg kg−1 dry weight | ||||||
| Major elements | Zr * | 0.50 a | 0.18 b | 64.6 | |||
| K | 9707.62 a | 8226.40 b | 15.3 | Ni | 0.48 b | 0.30 a | 37.3 |
| P | 3334.57 b | 3959.37 a | 18.7 | Pb * | 0.46 a | 0.11 b | 75.8 |
| Mg | 1443.81 b | 1690.27 a | 17.1 | V * | 0.29 a | 0.11 b | 61.1 |
| Ca | 678.22 a | 447.04 b | 34.1 | Ce * | 0.09 a | 0.05 b | 41.8 |
| Na | 44.17 b | 267.00 a | 504.5 | As | 0.06 a | 0.03 b | 50.5 |
| Minor or trace elements | Ga * | 0.06 a | 0.02 b | 64.9 | |||
| Fe | 72.82 a | 49.92 b | 31.4 | Sn | 0.05 a | 0.04 b | 20.1 |
| Zn * | 27.56 a | 8.53 b | 69.1 | Ge * | 0.05 a | 0.02 b | 58.1 |
| Mn | 15.30 a | 16.19 a | 5.9 | La * | 0.04 a | 0.03 b | 40.5 |
| Cu | 6.70 a | 6.34 a | 5.3 | Li | 0.04 b | 0.05 a | 19.1 |
| Co | 0.06 a | 0.06 a | 2.8 | Nb * | 0.04 a | 0.03 b | 34.2 |
| Ultratrace elements | Y * | 0.04 a | 0.02 b | 39.0 | |||
| Si | 9968.95 a | 7837.06 b | 21.4 | Nd * | 0.04 a | 0.03 b | 322.0 |
| Al | 36.85 a | 26.72 b | 27.5 | Cr | 0.29 a | 0.16 b | 45.3 |
| Ti * | 8.61 a | 4.18 b | 51.5 | Pr * | 0.01 a | 0.01 b | 33.5 |
| Sr | 3.99 b | 7.71 a | 92.9 | Th * | 0.01 a | 0.01 b | 26.6 |
| Ba * | 2.07 a | 0.53 b | 74.1 | Sm * | 0.01 a | 0.01 b | 35.8 |
| Rb * | 0.96 b | 1.42 a | 47.2 | ||||
| Mineral | Pericarp | Embryo Cotyledon Mesophyll | Endosperm | Perisperm | ||||||
|---|---|---|---|---|---|---|---|---|---|---|
| Non-Saline | Saline | Non-Saline | Saline | Non-Saline | Saline | Non-Saline | Saline | |||
| C | 51.89 ± 3.2 a | 50.5 ± 3.7 a | 61.32 ± 5.70 a | 57.37 ± 2.40 a | 61.64 ± 1.40 a | 66.17 ± 5.3 a | 68.60 ± 2.7 a | 74.23 ± 7.10 a | ||
| O | 39.96 ± 1.5 a | 41.4 ± 1.00 a | 26.51 ± 5.00 a | 30.70 ± 1.40 a | 25.76 ± 1.40 a | 22.41 ± 3.6 a | 26.30 ± 2.1 a | 23.89 ± 4.90 a | ||
| N | n.d. | n.d. | 9.75 ± 0.30 a | 10.53 ± 0.10 a | 11.46 ± 0.05 a | 7.31 ± 1.3 b | 4.97 ± 0.01 a | 2.50 ± 0.70 b | ||
| Na | 0.08 ± 0.04 b | 0.5 ± 0.03 a | n.d. | n.d. | n.d. | N.d. | n.d. | n.d. | ||
| Mg | 0.23 ± 0.10 a | 0.34 ± 0.10 a | 0.38 ± 0.07 a | 0.39 ± 0.04 a | 0.16 ± 0.03 b | 0.38 ± 0.04 a | 0.01 ± 0.01 a | 0.02 ± 0.01 a | ||
| Si | 0.26 ± 0.10 a | 0.21 ± 0.10 b | n.d. | n.d. | n.d. | n.d. | n.d. | n.d. | ||
| P | 0.06 ± 0.01 b | 0.08 ± 0.04 a | 1.01 ± 0.07 a | 0.65 ± 0.10 b | 0.17 ± 0.01 b | 1.23 ± 0.05 a | n.d. | n.d. | ||
| S | 0.19 ± 0.07 a | 0.22 ± 0.2 a | 0.45 ± 0.07 a | 0.15 ± 0.01 b | 0.11 ± 0.01 b | 0.69 ± 0.1 a | 0.02 ± 0.01 a | 0.06 ± 0.01 a | ||
| Cl | 0.60 ± 0.10 b | 1.24 ± 0.40 a | n.d. | 0.02 ± 0.01 a | 0.07 ± 0.02 a | n.d. b | 0.08 ± 0.03 b | 0.30 ± 0.20 a | ||
| K | 6.18 ± 10 a | 5.46 ± 3.20 a | 0.59 ± 0.05 a | 0.22 ± 0.01 b | 0.52 ± 0.07 b | 1.31 ± 0.10 a | 0.12 ± 0.10 b | 0.26 ± 0.05 a | ||
| Ca | 0.31 ± 0.20 a | 0.16 ± 0.10 b | n.d. | n.d. | 0.12 ± 0.01 a | 0.04 ± 0.01 b | n.d. | n.d. | ||
| Fe | 0.08 ± 0.05 a | 0.06 ± 0.04 b | n.d. | 0.01 ± 0.00 a | n.d. | n.d. | n.d. | n.d. | ||
| Br | 0.09 ± 0.09 b | 0.13 ± 0.10 a | n.d. | n.d. | n.d. | n.d. | n.d. | n.d. | ||
| Al | 0.05 ± 0.04 a | 0.03 ± 0.03 b | n.d. | n.d. | n.d. | n.d. | n.d. | n.d. | ||
| Cu | 0.05 ± 0.04 a | N.d. b | n.d. | n.d. | n.d. | n.d. | n.d. | n.d. | ||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
González, J.A.; Hinojosa, L.; Mercado, M.I.; Fernández-Turiel, J.-L.; Bazile, D.; Ponessa, G.I.; Eisa, S.; González, D.A.; Rejas, M.; Hussin, S.; et al. A Long Journey of CICA-17 Quinoa Variety to Salinity Conditions in Egypt: Mineral Concentration in the Seeds. Plants 2021, 10, 407. https://0-doi-org.brum.beds.ac.uk/10.3390/plants10020407
González JA, Hinojosa L, Mercado MI, Fernández-Turiel J-L, Bazile D, Ponessa GI, Eisa S, González DA, Rejas M, Hussin S, et al. A Long Journey of CICA-17 Quinoa Variety to Salinity Conditions in Egypt: Mineral Concentration in the Seeds. Plants. 2021; 10(2):407. https://0-doi-org.brum.beds.ac.uk/10.3390/plants10020407
Chicago/Turabian StyleGonzález, Juan A., Leonardo Hinojosa, María I. Mercado, José-Luis Fernández-Turiel, Didier Bazile, Graciela I. Ponessa, Sayed Eisa, Daniela A. González, Marta Rejas, Sayed Hussin, and et al. 2021. "A Long Journey of CICA-17 Quinoa Variety to Salinity Conditions in Egypt: Mineral Concentration in the Seeds" Plants 10, no. 2: 407. https://0-doi-org.brum.beds.ac.uk/10.3390/plants10020407