
RNAs on the Go: Extracellular Transfer in Insects with Promising Prospects for Pest Management


Research Group of Molecular Developmental Physiology and Signal Transduction, Division of Animal Physiology and Neurobiology, Department of Biology, KU Leuven, Naamsestraat 59, 3000 Leuven, Belgium
*
Author to whom correspondence should be addressed.
Plants 2021, 10(3), 484; https://0-doi-org.brum.beds.ac.uk/10.3390/plants10030484
Submission received: 31 January 2021
/
Revised: 28 February 2021
/
Accepted: 1 March 2021
/
Published: 4 March 2021
(This article belongs to the Special Issue External RNA Application for Gene Regulation and Plant Resistance)
Abstract
:RNA-mediated pathways form an important regulatory layer of myriad biological processes. In the last decade, the potential of RNA molecules to contribute to the control of agricultural pests has not been disregarded, specifically via the RNA interference (RNAi) mechanism. In fact, several proofs-of-concept have been made in this scope. Furthermore, a novel research field regarding extracellular RNAs and RNA-based intercellular/interorganismal communication is booming. In this article, we review key discoveries concerning extracellular RNAs in insects, insect RNA-based cell-to-cell communication, and plant–insect transfer of RNA. In addition, we overview the molecular mechanisms implicated in this form of communication and discuss future biotechnological prospects, namely from the insect pest-control perspective.
1. Introduction: Regulatory RNAs and Insect-Plant Interactions
RNAs exist in a wide variety of structures and sizes, well suited to regulate a multitude of processes. Regulatory RNAs, also referred to as non-coding RNAs, do not contribute directly to protein synthesis but function at various control levels to modulate gene expression. These molecules act both at the transcriptional and post-transcriptional levels, by mediating chromatin modulation, regulating alternative splicing, inducing suppression of translation, or directing the degradation of target transcripts [1].
Eukaryotic regulatory RNAs are broadly classified into long (≥200 nt) and small (≤200 nt). While numerous of the so-called long non-coding RNAs are described to regulate gene expression at various levels, it has recently been shown that some might, in fact, have coding functions [1,2]. Nonetheless, long non-coding RNAs and the mechanisms by which they exert their functions are still poorly characterized and deserve further research efforts. On the other hand, small RNA (sRNA)-based regulatory mechanisms are well established. In particular, the discovery of the RNA interference (RNAi) mechanism in animals resulted in a Nobel Prize and motivated a boom of comprehensive studies unveiling the functional role of these molecules in post-transcriptional silencing [3,4,5,6]. In short, during RNAi, sRNAs of approximately 18–30 nt are incorporated into an RNA-induced silencing complex (RISC), which is then directed to a target transcript via Watson–Crick base pairing. Subsequently, an Argonaute (Ago) protein within RISC acts to inhibit or degrade the target transcript, resulting in suppressed gene expression [7,8].
Classification of sRNAs relies on their biogenesis mechanisms, size, complementarity to the target, associated proteins, and main regulatory processes in which they are involved. Based on these, several sRNAs are recognized among eukaryotes, of which two are common to plants and animals: microRNAs (miRNAs) and small interfering RNAs (siRNAs). In broad terms, miRNAs originate from the processing of endogenous stem-loop RNA precursors and act to regulate the expression of endogenous genes. In turn, siRNAs originate from long double-stranded RNA (dsRNA) structures and mainly function in the protection against viruses and transposons [9,10,11]. While many other sRNA types are distinguished, within and beyond the formerly described classes, these are not discussed in the context of the current review.
Although the mechanisms by which they act are not as extensively investigated as in eukaryotes, regulatory RNAs are also present in Archaea and Bacteria. In this regard, the RNA chaperone Hfq is well described to play a central role in several RNA-based regulatory systems in prokaryotes [12,13,14,15,16,17]. Furthermore, prokaryotic Ago proteins have been shown to contribute to some forms of RNA-guided gene regulation [18,19,20]. In addition, the CRISPR-Cas (clustered regularly inter-spaced short palindromic repeats and associated genes) system has attracted a lot of attention due to its exceptional potential for RNA-guided genome editing. In fact, this mechanism led to the 2020 Nobel Prize in Chemistry [21,22]. While prokaryotic regulatory RNAs differ from their eukaryotic counterparts in several aspects, such as size range and associated pathways, their contribution to regulatory mechanisms in multiple processes is also evident [23,24,25].
While regulatory RNAs and their pathways are mostly investigated at the intracellular level, discoveries regarding a role of extracellular RNAs in cell-to-cell communication are now making the scene. Numerous are the examples of interspecies and even interkingdom RNA-based communication, especially those concerning host–parasite/symbiont interactions. The study of RNA-based transfer of information promises not only to bring fundamental insights into a novel layer of regulatory complexity, but also to open doors to a handful of biotechnological applications [26,27,28].
Insects are the largest and most diverse group of animals on Earth. They form an important component of the ecosystems and affect many aspects of human life. A major part of insects’ impact occurs via their interaction with plants; for instance, some species are important pollinators, while others are agricultural pests and vectors of plant diseases [8,29,30]. In this scope, insects and their host plants have gone through a long coevolution and thereby developed several known forms of communication via specific effector proteins, volatiles, and other chemicals [31,32]. Recently, growing evidence suggests the existence of RNA-based cell-to-cell communication in insects and in plant–insect interactions. Understanding the mechanisms of intercellular RNA transfer in insects, as well as of the communication between Animalia and Plantae, promises to contribute to the development of novel technologies. These are crucial to cope with insect-related challenges, and valuable examples are RNAi-directed pest control strategies. In this manuscript, we review the current state of the art concerning RNA-based communication in and between insects, as well as the RNA transfer between plants and insects. In addition, we discuss the possible underlying transfer mechanisms and the related biotechnological prospects.
2. Extracellular RNA-Based Communication
The presence of RNA molecules in extracellular environments, mainly of sRNAs, has been systematically reported. Throughout the past decade, it became evident that RNA molecules mediate an important layer of communication, even between diverse and phylogenetically distant organisms [28,33,34]. In plants, the role of mobile RNAs in intercellular communication has been well-established [35,36]. In animals, extracellular RNAs are extensively described in mammalian biofluids. These molecules are known to be secreted followed by functional uptake into recipient cells [37,38,39,40,41,42,43,44,45,46,47,48,49,50,51,52,53,54,55,56]. In invertebrates, the presence of extracellular RNAs has been demonstrated in arthropods and nematodes, with a limited number of studies demonstrating functional RNA transfer [57,58,59,60,61,62,63,64,65,66,67,68,69,70,71,72,73,74,75,76,77,78,79,80,81,82]. Below, we review the current knowledge on extracellular RNAs in insects, establishing the distinct levels of this communication: intercellular/intraindividual, interindividual, interspecies, and interkingdom (insect–plant).
2.1. Intercellular/Intraindividual
In insects, little more than a handful of studies report extracellular RNAs and their (potential) role in intercellular/intraindividual communication, namely in beetles and flies: (i) extracellular sRNA populations have been observed in the hemolymph of the korean rhinoceros beetle, Allomyrina dichotoma [67]; (ii) miRNAs were detected in cell-free hemolymph of Drosophila melanogaster flies, and some were demonstrated to differentially accumulate with age [62]; (iii) several sRNA populations have been found in the extracellular medium of the D. melanogaster cell lines S2R+ and D17-c3 [63]; (iv) extracellular miRNAs have been identified in medium of cultured D. melanogaster cells, namely of S2 and Cl8 cell lines, with indications for differential secretion [65]; (v) in Drosophila flies, extracellular siRNAs have been shown to contribute to systemic antiviral immunity [64]; (vi) extracellular siRNAs were demonstrated to spread the RNAi signal in red flour beetle, Tribolium castaneum cultured cells [66]; and (vii) in the Colorado potato beetle, Leptinotarsa decemlineata, dsRNA has been identified in the extracellular medium of cultured cells previously treated with dsRNA [68]. These findings are summarized in Table 1.
2.2. Interindividual
Besides serving cell-to-cell communication within single organisms, RNA molecules have the potential to mediate interactions between individuals. This is particularly studied in insects, where interindividual RNA-based communication has been shown in social species. The first finding regards the honey bee, Apis mellifera, where caste determination has been shown to be influenced by miRNAs in nurse bee secretions [70]. More recently, a transmissible RNA pathway across generations, via the hemolymph and followed by secretion in the jellies, has been demonstrated [71,72]. In addition, miRNAs have been identified in the trophallactic fluid of the Florida carpenter ant, Camponotus floridanus, contributing to the concept of interindividual RNA transfer in social insects [73]. Table 1 resumes these reports.
2.3. Interspecies
Functional RNA transfer between different species is also currently described in animals. This is broadly termed interspecies RNA-based communication, and the so-far described examples mainly refer to host–parasite interactions [74,75,77]. A fascinating example is reported in insects and concerns the parasitic wasp Cotesia vestalis and its host Plutella xylostella, the diamondback moth. Specifically, the developing wasp larva has the capacity to release teratocyte cells into the host body. These cells then secrete extracellular vesicles (EVs) containing the miRNA Cve-miR-281. On another front, the wasp larva also releases the associated C. vestalis bracovirus, which expresses the Cve-miR-novel22 miRNA in the host. Remarkably, both miRNAs influence the host’s ecdysone cascade to delay its development, hereby granting the parasite additional time to develop [74]. In addition, sRNAs have been identified in saliva of mosquitos and other blood-feeding arthropods, allowing speculation about functions of these molecules in modulating host response [59,60,61,82]. In line with this, parasitic nematodes were shown to secrete sRNA molecules, which, in some cases, have been proven to regulate host biology [77,78,79,80]. The reports on (potential) interspecies RNA-based communication in insects are briefed in Table 1. Noteworthy, interspecies RNA communication is also reported within Plantae. The studied examples mainly regard parasitic plants from the genus Cuscuta and their hosts, where bidirectional RNA transfer has been demonstrated [83,84,85,86,87,88,89,90]. Furthermore, Cuscuta miRNAs have been reported to target Arabidopsis genes [91]. In addition, unidirectional movement of small RNAs from shoots to roots in a soybean–common bean (Glycine max-Phaseolus vulgaris) grafting system has been recently demonstrated [92].
2.4. Interkingdom
There are known cases of RNA transfer crossing the three domains of life and several examples of interkingdom RNA-based communication between plants or insects and their microorganisms. Within Eukarya, functional RNA transfer has been reported between plants (Plantae) or insects (Animalia) and their fungal (Fungi) or protist (Protista) pathogens [28,93,94,95,96,97]. In this scope, the currently available knowledge points towards the existence of functional RNA transfer systems between insects and plants, as addressed in the following sections.
2.4.1. Insect to Plant RNA-Based Communication
Until now, very little is known regarding RNA transfer from insects to plants, with only three reported studies. The first indication for a possible insect–plant functional RNA transfer was obtained in the green peach aphid, Myzus persicae. In this study, reduced aphid fecundity was observed when the insect fed on Arabidopsis mutants for the miRNA pathway. This led to speculation that processing of aphid miRNAs by the plant might mediate aphid infestation [98]. Later, sRNAs from the phloem-feeding whitefly Bemisia tabaci have been identified both in tissue and phloem of the tomato plant [99]. Interestingly, more recently, M. persicae aphids were demonstrated to transfer the Ya transcript to their host plants, where it moves to distal leaves. This transcript was identified as a long non-coding RNA virulence factor, with functional studies demonstrating that it promotes aphid fecundity [100].
2.4.2. Plant to Insect RNA-Based Communication
The presence and functionality of dietary RNAs in animals have been a matter of debate over the last decade [101,102,103,104,105,106]. Nevertheless, in insects, increasing functional evidence points towards the natural existence of some insect–plant RNA-based communication levels. Several studies identify plant-derived RNAs in herbivorous insects, and some even report functionality. The presence of Cucumis melo miRNAs has been shown in the cotton–melon aphid, Aphis gossypii, demonstrating that plant miRNAs are stable in the insect body [107]. In the silkworm, Bombyx mori, mulberry miRNAs were identified in both hemolymph and tissues [108]. Later, sorghum miRNAs were identified in the greenbug, Schizaphis graminum, and yellow sugarcane miRNAs in the yellow sugarcane aphid, Sipha flava. In silico target prediction indicated the involvement of these miRNAs in processes important for aphid fecundity, suggesting a role in decreasing aphid infestation [109]. Another study identified miRNAs of Brassica oleracea in the gut of M. persicae, the green peach aphid. The predicted miRNA targets in the insect were mainly carbohydrate transport and metabolism, RNA processing and modification, and nuclear structure genes [110]. Plant-derived miRNAs were also shown to be present in the hemolymph of P. xylostella, the diamondback moth. Importantly, two of the most abundant identified plant miRNAs were validated to target P. xylostella hemocyanin domain containing genes, that are known to play important roles in the hemolymph of arthropods. Moreover, other highly abundant plant miRNA was demonstrated to influence the pupal development and egg-hatching rate [111,112]. Remarkably, plant miRNAs found in honeybee larval food contribute to caste determination. Specifically, plant miRNAs are enriched in beebread in comparison with royal jelly, some of which were demonstrated to prevent larval differentiation into queens and to induce development into worker bees. Feeding miR162a to larvae resulted in worker bee phenotypes, with similar results in D. melanogaster larvae [113]. This miRNA targets A. mellifera TOR, a key player in cast development [113,114]. Table 2 summarizes the studies reviewed in this section.
2.4.3. Engineered Plant-to-Insect RNA Transfer
Adding to the previously described observations, it is relevant that plant–insect functional transfer of RNA can be engineered to occur. In short, host plants can be genetically modified to express specific RNA molecules targeting key insect genes. When the insect feeds on the plant, these RNA molecules enter the insect body, are taken up by recipient cells, and induce gene silencing by RNAi. In the context of insect pest control, there are several examples of transgenic plants designed to induce RNAi in insects, a process commonly designated as host-induced gene silencing. The first successful studies on achieving resistance to a pest–insect via transgenic plants expressing dsRNA date to 2007 [115,116]. Since then, multiple proofs of this concept have been reported [117,118,119,120,121,122,123,124,125,126,127,128,129,130,131]. Often, long dsRNA molecules are expressed. However, in planta expression of insect miRNAs has been suggested to be a good option as well [132,133,134,135]. In addition, a plant-expressed insect miRNA precursor was shown to be more effective than expressing the miRNA itself [136]. The authors suggested this was due to the fact that insect miRNA precursors are not processed by the plant, since they lack Drosha, an important nuclear processing enzyme of insect miRNA biogenesis [136]. In addition, expression of RNAi-inducing molecules in the chloroplast instead of in the cellular cytoplasm, via modification of the chloroplast genome, revealed to be quite promising [126,137,138,139]. Since no RNAi-processing machinery is present in this organelle, higher accumulation of the RNA molecules in the plant is obtained, resulting in a more effective RNAi response in the insect [126,137,138,139]. Nevertheless, this approach is not always effective, suggesting that, in some cases, RNA molecules from the chloroplast are not transferred to the insects [140]. Since recently, the first RNAi-based pest-resistant plant has been available on the market. It is named SmartStax Pro® and was developed by Monsanto and Dow Agrosciences [141]. This illustrates the economical relevance of this field and emphasizes the need for understanding the natural mechanisms of plant–insect RNA transfer, possibly contributing to the development of efficient pesticides based on exogenous RNA.
3. Mechanisms of RNA Transfer
Transfer of information via extracellular RNAs has been seen to occur in insects. In addition, bidirectional plant–insect RNA transfer has been reported and, in some cases, the RNA signal was shown to be functional. This was demonstrated both in the context of specific (possibly coevolved) plant–insect interactions, as well as of genetically engineered plants, to exert RNA-based insect pest control (see Section 2). Nevertheless, little is known regarding the mechanisms by which mobile RNAs spread in insects, or between plant and insect cells. To date, three main routes for cell-to-cell RNA based communication have been proposed, namely via transfer of naked RNA, RNA bound to RNA-binding proteins (RBPs), and RNA-containing extracellular vesicles (EVs).
3.1. Naked RNA
In plants, naked RNAs, i.e., not protected by RNA binding proteins or encapsulated in EVs, can be transferred over short and long distances [36,142,143,144]. Short range transfer of naked sRNAs in plants occurs via the plasmodesmata, by the space between the plasma membrane and the desmotubule, and by the desmotubule itself, which connects the endoplasmic reticulum of the two adjacent cells [144,145,146,147]. Similarly, in fungi, naked sRNA can move from cell-to-cell within short distances through the septal pore [144]. In mammals, transfer between adjacent cells can be mediated by gap junctions. In this respect, siRNA- and miRNA-mediated transfer via gap junctions has been described [148,149,150,151,152,153,154,155,156,157]. To our knowledge, the transfer of RNA molecules via gap junctions has not been investigated in other groups of animals. However, in the nematode Caenorhabditis elegans, transmembrane systemic RNA interference defective (SID) proteins act as channels for intercellular RNA movement [158,159,160,161]. In insects, short range transport of dsRNA and RISC components through nanotube-like structures was suggested between Drosophila cells [162]. However, whether these structures might effectively transfer (naked) RNAs between adjacent insect cells remains to be demonstrated. Taken together, these studies suggest that similar structures might function for short range RNA transfer in plants, fungi and animals.
In both plants and fungi, apoplastic movement of naked sRNA can also occur. In this case, naked sRNAs are secreted from the plasma membrane and move throughout the cell wall to extracellular spaces, where they can then enter both neighboring or distant cells [144,145,146,147].
Plants also transfer naked sRNAs via the phloem, using the vascular system to spread these molecules throughout the plant to distant cells [144,146,147]. In addition, it is noteworthy that several reports indicate the transfer of naked sRNA between plants and fungi [96,163,164,165], indicating bidirectional interkingdom RNAi between plants and fungi. Specialized infection structures of fungi and parasitic plants, termed haustoria, may act as a gateway for sRNA transfer between host and pathogen at the plant–plant and plant–fungi levels [91].
In human plasma, naked extracellular RNAs are rapidly degraded [166]. Similarly, naked RNA molecules are rapidly degraded in insect biofluids [8,167,168,169,170,171]. Nevertheless, it is by now clear that stable RNA molecules circulate in animal extracellular fluids (see Section 2). Together, these facts contribute to the idea that mobile RNAs in animal biofluids require protection form degradation in order to be functionally transferred.
3.2. RNA Associated with RNA Binding Proteins (RBPs)
In plants, RBPs are established to mediate short- and long-range RNA transport. The Cucurbita maxima Phloem Small RNA-Binding Protein 1 can bind sRNAs, transferring them between cells, both via the plasmodesmata and the phloem [172,173]. Furthermore, other RBPs have been identified in the phloem of different plants [174,175,176]. Interestingly, Ago proteins have also been suggested to be implicated in sRNA transfer in plants [177,178]. In addition, recently, a conserved family of sRNA-binding proteins–Small RNA-Binding Protein 1 family—that function in intercellular transfer of sRNAs has been identified in the phloem of several plants [179].
In 2008, Mitchell and colleagues demonstrated that extracellular sRNAs present in human plasma are protected from degradation due to their association with certain entities [166]. In line with this, most mammalian plasma miRNAs are associated with Ago proteins [180,181,182]. Interestingly, Neuropilin-1 has been reported to be a receptor for miRNA–Ago complexes [183]. Nevertheless, due to the remarkable extracellular stability reported for some Ago proteins, it is often suggested that extracellular RNA–Ago complexes are by-products of cell death [180,181,184]. In the nematode Heligmosomoides bakeri, secondary siRNAs are loaded into an extracellular Ago protein, and this complex is subsequently secreted in EVs, suggesting a role of this Ago protein in mediating the selective sorting of sRNAs in EVs in this species [79]. In the fruit fly, extracellular miRNAs have been shown to be stably present in the hemolymph, and an in vitro study with Drosophila-derived cell lines verified the presence of extracellular miRNAs associated with an Ago protein [62,65], suggesting that Ago proteins might also confer sRNA stability in insects (Figure 1).
Besides Ago proteins, the association of sRNAs to lipoproteins has been demonstrated as well. Lipoproteins have been shown to be associated with miRNAs, and high-density lipoproteins (HDLs) can functionally transfer miRNAs to recipient cells [185]. Furthermore, miRNA-delivery mediated by HDL was shown to be dependent on scavenger receptor class B type I [185]. Since then, other reports have emphasized the role of HDLs in intercellular RNA transfer, as well as the potential use of these lipoproteins as therapeutic-delivery vehicles [185,186,187,188,189,190,191]. Interestingly, insect and other arthropod lipoproteins were demonstrated to adhere to dsRNA, suggesting a potential role of these proteins in mediating RNA-based communication in this phylum [169,192,193] (Figure 1).
Additionally to lipoproteins and Ago proteins, other animal proteins have been suggested to bind RNA in the extracellular environments. Particularly, mammalian Nucleophosmin1 was demonstrated to bind miRNAs and protect them from nuclease degradation [194]. In insects, more specifically in honeybees, a secreted RBP named Major Royal Jelly Protein 3 (MRJP-3) binds to RNA in jelly, protecting it from degradation and enhancing its uptake [72] (Figure 1).
3.3. RNA-Containing Extracellular Vecicles (EVs)
EVs form a heterogeneous group consisting of exosomes, microvesicles and apoptotic bodies. Although long viewed as part of cellular waste disposal pathways, it is by now clear that EVs can functionally transfer their content (RNA, DNA, lipid, and protein) to recipient cells [195].
Despite previous debate regarding plant cell wall preventing formation and function of EVs, recent evidence shows that EVs are also produced by these organisms [97,165,196,197,198]. In addition, plant EVs have been shown to contain RNA [197,199,200,201], and selective sRNA loading in EVs has been observed [202]. Moreover, the transfer of sRNAs within EVs from plantae to fungi has been recently demonstrated [97]. Interestingly, specific RBPs, including Ago proteins, have been suggested to facilitate the packaging of RNAs into EVs in plants [178,203].
In 2007, a first study demonstrating that EVs mediate intercellular communication in mammalian cell lines, by transferring functional RNA from donor to recipient cells, was reported [37,38]. Since then, a myriad of reports indicate EV-mediated intercellular communication in mammals [39,40,41,42,43,44,45,46,47,48,49,50,51,52,53,54,55,56,204,205,206,207,208,209]. Currently, increasing evidence points towards the ubiquitous presence of RNA-containing EVs in animals, as suggested by studies in the nematodes C. elegans [57,58,69,76], Heligmosomoides polygyrus, Litomosoides sigmodontis [77], Brugia malayi [78], H. bakeri, and Trichuris muris [80]; in the ticks Ixodes Ricinus and Haemaphysalis longicornis [59,82]; as well as in the red swamp crayfish, Procambarus clarkia [81]. Also in insects, several reports from recent years suggest the involvement of EVs in a common mechanism for functional RNA transfer between cells. RNA-containing EVs have been reported in the fruit fly, namely in the hemolymph [62,64] and in cultured cells [63,65]; as well as in beetles, specifically in the hemolymph of A. dichotoma [67] and in cell lines of T. castaneum [66] and L. decemlineata [68]. Moreover, EV-specific miRNA profiles have been shown in Drosophila [62,65]. Noteworthy, functional transfer of RNA within EVs was demonstrated in three studies. First, hemocyte-derived EVs containing secondary viral siRNAs confer systemic RNAi antiviral immunity in D. melanogaster [64]. Second, in T. castaneum cell culture, siRNA-containing EVs spread an RNAi-based silencing signal to recipient cells [66]. Third, EVs of the parasitic wasp C. vestalis deliver miRNAs to its host P. xylostella, which target its ecdysone cascade, resulting in a developmental delay [74]. Likewise, the parasitoid wasp Leptopilina boulardi injects vesicles in its host Drosophila. Although transfer of RNA has not been investigated in this case, these EVs—venosomes—were shown to interfere with the hosts’ defense [210]. Moreover, EVs present in bee pollen, honey, and royal jelly were shown to function in the antibacterial properties of these products [211]. Together, these studies suggest that EVs function in transport of molecules in and between insects, including their RNA cargo (Figure 1).
Two potential RNA selective sorting mechanisms have been proposed in mammals. First, miRNAs packaged into EVs were demonstrated to contain specific nucleotide motifs with which specific ribonucleoproteins interact to allow selective sorting [212,213,214,215,216,217,218,219]. Moreover, several membrane proteins involved in EV biogenesis are also involved in selective miRNA sorting [50,220,221,222]. Second, certain miRNA post-transcriptional modifications appear to contribute to their selective sorting, as 3′ end uridylated miRNAs were found overrepresented in EVs while 3′ end adenylated miRNAs were relatively enriched intracellularly [223].
EVs are produced by all domains of life and are considered part of an ancient mechanism for RNA export [224,225]. In fact, several reports describe EV-mediated RNA transfer, within and among animals, plants, fungi and microbes [11,28,33,34,144,197,198,225,226,227]. Although further detailed research is needed to investigate potential mechanisms of RNA transfer between insects and plants, the current knowledge indicates EVs as promising candidates. Figure 1 summarizes the findings regarding RNA transfer mechanisms in insects.
4. Biotechnological Prospects
To sustain the currently growing food demand, agricultural pests are one of the many challenges that are faced. Pest insects destroy 18–20% of the annual global crop production, which is estimated at $470 billion [228]. The classical approach of combatting agricultural insect pests is via non-selective conventional insecticides. However, their limited target species selectivity has serious disadvantages, including a detrimental impact on environment and human health [229,230,231,232]. In fact, these concerns are increasing and have led to a ban in some commonly used insecticides [233,234,235]. In addition, some pest insect populations are already resistant against many of the conventionally used insecticides [236,237,238,239,240,241].
RNA-mediated insect gene silencing is a promising tool to contribute to highly specific pest control strategies. Nevertheless, the effect of RNA is not equally efficient in every insect, with high levels of variability between different species and even between different populations of the same species. RNA degradation by gut nucleases and extreme pH conditions upon ingestion is appointed as a reason for inefficient RNA-based silencing in several insect species. In addition, inefficient or absent functional RNA uptake systems in the gut, as well as the lack of an efficient systemic RNAi spreading mechanism from the gut to the rest of the insect body, are also considered potentially important limiting factors [8,168,242,243]. RNA might be available for ingestion by the insect via different approaches, namely via expression by genetically modified plants (see Section 2.4.3) or by exogenous in planta application. As such, several RNA insect delivery systems have been proposed, meant to exert RNA protection and mediate intracellular delivery. Well-known examples are based on nanoparticles, liposomes, RBPs, bacteria and viruses, among others [8]. In this scope, natural systems of RNA transfer within insects, as well as from plants to insects, represent key fundamental concepts that might lead the development of more efficient RNA-based insect gene silencing strategies.
In mammals, both RBPs and EVs have been explored for efficient cellular delivery of nucleic acids. These approaches are mainly explored in humans, in the context of targeted drug delivery therapeutics. Interesting examples are mammalian lipoproteins, which are often proposed for use in human siRNAs delivery [244,245,246,247,248,249]. In addition, a high percentage of mammalian extracellular miRNAs are bound to Ago proteins, and pre-assembled siRNA-Ago complexes, delivered via different carriers, can increase the gene silencing effect in mice [180,181,182,250]. Interestingly, insect lipoproteins (i.e., lipophorins) are known to bind exogenous dsRNA in the hemolymph [169,192], and Ago proteins have been identified in the extracellular medium of cultured insect cells [65]. Hence, these proteins may be promising candidates for design of exogenous RNA insect delivery systems, highlighting the importance of investigating natural RNA transfer mechanisms.
Besides RBPs, many studies appoint EVs as promising human drug-delivery vehicles [206,251,252]. In fact, engineering of EVs to deliver nucleic acid based therapeutics is already being explored in the market [253,254]. Considering the role of EVs in RNA-based intercellular, interspecies, and interkingdom communication, such structures might hold great potential for RNA-based pest management [226]. In addition, since increasing evidence indicates the role of EVs in transferring RNA molecules in insects (see Section 3.3), it is exciting to consider the development of EV-based RNA-delivery systems to control pests via exogenous RNA. Although RBP- and EV-based crop protection systems are still to be explored, these may lead to promising strategies for the future.
To ensure environment-friendly and biosafe insecticides, specificity is a watchword. The idea of RNA-based pest control strategies is highly popular since high species- and gene-specificity can be obtained at the level of the nucleic-acid sequence [131,255]. Currently, it is tempting to speculate that specificity of RNA-based insecticides could be achieved at additional levels.
First, several factors have been shown to influence the loading of sRNAs into Ago proteins, such as their sequence and structure [256,257,258,259,260,261,262,263,264]. In insects, generation of siRNAs with species-dependent length have been observed [265], and certain sRNA chemical modifications seem to vary among species. Specifically, D. melanogaster siRNAs are 2′-O-methylated, while this is not the case in the lepidopteran species P. xylostella, B. mori, and Trichoplusia ni [266,267]. In addition, RNAi genes have been shown to be fast evolving, resulting in lower levels of similarity between species (e.g., dicer2 or argonaute2) [268,269,270]. It is therefore interesting to conjecture that a second level of specificity could be achieved, determined by a species-specific ability to intracellularly recognize and process the exogenous RNA molecules. Although research efforts are needed to explore these possibilities, one can hypothesize that this could be based on the length of sRNA molecules, its chemical modifications, and/or its complexation with specific (modified) proteins of the RNAi machinery.
Second, exhaustively unravelling mechanisms of RNA transfer within and between a wide range of organisms can stimulate the design of highly specific RNA delivery vehicles. It is important to elucidate what EV populations and protein-complexes interact with RNAs in different species, as well as to determine their composition and structure. It is also relevant to determine their specificity and, in this regard, an example has recently been reported in bees. Specifically, the domain of MRJP-3 that provides its RNA binding activity seems to have emerged in the Apis genus and to only be associated with jelly-secreting bees [72]. Moreover, efforts should be made to understand uptake mechanisms of these structures, including the potential involvement of specific ligand–receptor interactions. In this scope, specialized host–pathogen/symbiont interactions might be particularly useful, as they would further contribute to refine specificity and efficiency of RNA delivery. Noteworthily, RNA delivery strategies based on host–pathogen/symbiont systems have already been proposed, emphasizing the potential of these interactions [271,272,273,274]. It is inspiring to consider that, based on specialized uptake of certain optimized RNA-containing vehicles, the target species could be solely affected.
In summary, three levels of specificity and efficiency could be aimed at, as determined by (1) the cellular delivery vehicle, (2) the recognition by the cellular machinery, and (3) the nucleotide sequence. Importantly, exploring these three levels in distinct contexts of integrated pest management has potential to delay, or avoid, the development of resistance. In addition, it is also relevant that the use of highly efficient and specific RNA delivery methods can contribute to the release of low quantities of the RNA-based insecticide in the environment, reducing potential toxicity concerns.
5. Conclusions
RNA molecules and their related pathways are classically described at the intracellular level. Recently, the existence of a novel RNA-based communication layer, via the spread of extracellular RNAs in biological fluids, became well established and gave rise to a novel research area that is currently booming in biomedical sciences. Although this research field remains underexplored in insects, growing evidence indicates that this route of communication is also functional in these animals. Specifically, extracellular RNAs are present in insects, are contained within EVs and/or interact with RBPs, and are transferred to recipient cells to exert their function. Nevertheless, only a limited number of studies are currently available, which do not cover the dimension and diversity of the class Insecta, or of the wide range of insect–plant interactions. A wider and deeper fundamental understanding of the intercellular RNA transfer mechanisms in these organisms is required. In this scope, there is an urgent need for well described, robust, and reproducible protocols for isolation of extracellular components, such as EVs and RBPs, from complex biofluids. In addition, although the advent of deep sequencing technologies has led to an increase of descriptive studies, these only form a starting point. In fact, functional studies regarding natural RNA-transfer mechanisms, as well as their potential for bioactive delivery of exogenous RNA molecules, are required. Noteworthily, it is critical to not only unravel conserved processes, but also species-specific mechanisms. Thus, besides model organisms, efforts must also be invested into several pest and beneficial systems. At last, on the long run, special attention should be given to the development of production/purification techniques of selected extracellular components bound to RNAs of interest; as well as to biosafety assays, namely concerning the specificity of the developed approaches.
Considering the potential of RNA-based insect pest control strategies, understanding the mechanisms by which RNAs are transferred within insects, as well as between plants and insects, promises to give leads to the development of effective strategies for exogenous RNA delivery. This has potential to lead to novel ways to protect crops, to combat insect pests, and to generate high economic, environmental and social value.
Author Contributions
Conceptualization: D.S.; investigation: D.S., S.R. and S.V.d.B.; writing—original draft preparation: D.S., S.R. and S.V.d.B.; writing—review and editing: D.S., S.R., S.V.d.B. and J.V.B.; supervision: D.S. and J.V.B.; funding acquisition: D.S. and J.V.B. All authors have read and agreed to the published version of the manuscript.
Funding
This work was funded by the Research Foundation of Flanders (FWO), grant number G093119N, and the Special Research Fund of KU Leuven, grant number C14/19/069. SR and SVdB are recipients of doctoral fellowships from the Research Foundation of Flanders (FWO), fellowship numbers 1SC7421N and 1169220N.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
This article reviews published studies.
Acknowledgments
The authors gratefully acknowledge the Research Foundation Flanders (FWO) and the Special Research Fund of KU Leuven for funding.
Conflicts of Interest
The authors declare no conflict of interest.
References
- A focus on regulatory RNAs. Nat. Cell Biol. 2019, 21, 535. [CrossRef]
- Wang, Q.; Boshoff, H.I.M.; Harrison, J.R.; Ray, P.C.; Green, S.R.; Wyatt, P.G.; Barry, C.E. PE/PPE proteins mediate nutrient transport across the outer membrane of Mycobacterium tuberculosis. Science 2020, 367, 1147–1151. [Google Scholar] [CrossRef]
- Fire, A.; Xu, S.; Montgomery, M.K.; Kostas, S.A.; Driver, S.E.; Mello, C.C. Potent and specific genetic interference by double-stranded RNA in Caenorhabditis elegans. Nature 1998, 391, 806–811. [Google Scholar] [CrossRef] [PubMed]
- Mello, C. Return to the RNAi world: Rethinking gene expression and evolution. Cell Death Differ. 2007, 14, 2013–2020. [Google Scholar] [CrossRef] [PubMed]
- Fire, A.Z. Nobel Lecture. Available online: https://www.nobelprize.org/prizes/medicine/2006/fire/lecture/ (accessed on 23 February 2021).
- Mello, C.C. Nobel Lecture. Available online: https://www.nobelprize.org/prizes/medicine/2006/mello/lecture/ (accessed on 23 February 2021).
- Olina, A.V.; Kulbachinskiy, A.V.; Aravin, A.A.; Esyunina, D.M. Argonaute Proteins and Mechanisms of RNA Interference in Eukaryotes and Prokaryotes. Biochemistry 2018, 83, 483–497. [Google Scholar] [CrossRef] [PubMed]
- Vogel, E.; Santos, D.; Mingels, L.; Verdonckt, T.-W.; Vanden Broeck, J. RNA Interference in Insects: Protecting Beneficials and Controlling Pests. Front. Physiol. 2019, 9, 1–21. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, V.N.; Han, J.; Siomi, M.C. Biogenesis of small RNAs in animals. Nat. Rev. Mol. Cell Biol. 2009, 10, 126–139. [Google Scholar] [CrossRef] [PubMed]
- Carthew, R.W.; Sontheimer, E.J. Origins and Mechanisms of miRNAs and siRNAs. Cell 2009, 136, 642–655. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huang, C.-Y.; Wang, H.; Hu, P.; Hamby, R.; Jin, H. Small RNAs—Big Players in Plant-Microbe Interactions. Cell Host Microbe 2019, 26, 173–182. [Google Scholar] [CrossRef] [Green Version]
- Bloch, S.; Węgrzyn, A.; Węgrzyn, G.; Nejman-Faleńczyk, B. Small and Smaller—sRNAs and MicroRNAs in the Regulation of Toxin Gene Expression in Prokaryotic Cells: A Mini-Review. Toxins 2017, 9, 181. [Google Scholar] [CrossRef] [PubMed]
- Fröhlich, K.S.; Velasco Gomariz, M. RNA-controlled regulation in Caulobacter crescentus. Curr. Opin. Microbiol. 2021, 60, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Evguenieva-Hackenberg, E.; Klug, G. New aspects of RNA processing in prokaryotes. Curr. Opin. Microbiol. 2011, 14, 587–592. [Google Scholar] [CrossRef]
- Hess, W.R.; Marchfelder, A. Regulatory RNAs in Prokaryotes; Springer Vienna: Vienna, Austria, 2012; ISBN 978-3-7091-0217-6. [Google Scholar]
- Saberi, F.; Kamali, M.; Najafi, A.; Yazdanparast, A.; Moghaddam, M.M. Natural antisense RNAs as mRNA regulatory elements in bacteria: A review on function and applications. Cell. Mol. Biol. Lett. 2016, 21, 6. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Watkins, D.; Arya, D.P. Regulatory roles of small RNAs in prokaryotes: Parallels and contrast with eukaryotic miRNA. Non-coding RNA Investig. 2019, 3, 28. [Google Scholar] [CrossRef]
- Li, Y.; Liu, X.; Huang, L.; Guo, H.; Wang, X.-J. Potential coexistence of both bacterial and eukaryotic small RNA biogenesis and functional related protein homologs in Archaea. J. Genet. Genomics 2010, 37, 493–503. [Google Scholar] [CrossRef]
- Makarova, K.S.; Wolf, Y.I.; van der Oost, J.; Koonin, E.V. Prokaryotic homologs of Argonaute proteins are predicted to function as key components of a novel system of defense against mobile genetic elements. Biol. Direct. 2009, 4, 29. [Google Scholar] [CrossRef] [Green Version]
- Hegge, J.W.; Swarts, D.C.; Van Der Oost, J. Prokaryotic argonaute proteins: Novel genome-editing tools? Nat. Rev. Microbiol. 2018. [Google Scholar] [CrossRef]
- Charpentier, E. Nobel Lecture. Available online: https://www.nobelprize.org/prizes/chemistry/2020/charpentier/lecture/ (accessed on 23 February 2021).
- Doudna, J.A. Nobel Lecture. Available online: https://www.nobelprize.org/prizes/chemistry/2020/doudna/lecture/ (accessed on 23 February 2021).
- Waters, L.S.; Storz, G. Regulatory RNAs in Bacteria. Cell 2009, 136, 615–628. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Babski, J.; Maier, L.-K.; Heyer, R.; Jaschinski, K.; Prasse, D.; Jäger, D.; Randau, L.; Schmitz, R.A.; Marchfelder, A.; Soppa, J. Small regulatory RNAs in Archaea. RNA Biol. 2014, 11, 484–493. [Google Scholar] [CrossRef] [Green Version]
- Dutta, T.; Srivastava, S. Small RNA-mediated regulation in bacteria: A growing palette of diverse mechanisms. Gene 2018, 656, 60–72. [Google Scholar] [CrossRef] [PubMed]
- Asgari, S. RNA as a means of inter-species communication and manipulation: Progresses and shortfalls. RNA Biol. 2017, 14, 389–390. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Das, S.; Ansel, K.M.; Bitzer, M.; Breakefield, X.O.; Charest, A.; Galas, D.J.; Gerstein, M.B.; Gupta, M.; Milosavljevic, A.; McManus, M.T.; et al. The Extracellular RNA Communication Consortium: Establishing Foundational Knowledge and Technologies for Extracellular RNA Research. Cell 2019, 177, 231–242. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hudzik, C.; Hou, Y.; Ma, W.; Axtell, M.J. Exchange of Small Regulatory RNAs between Plants and Their Pests. Plant Physiol. 2020, 182, 51–62. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gill, R.J.; Baldock, K.C.R.; Brown, M.J.F.; Cresswell, J.E.; Dicks, L.V.; Fountain, M.T.; Garratt, M.P.D.; Gough, L.A.; Heard, M.S.; Holland, J.M.; et al. Protecting an Ecosystem Service. In Advances in Ecological Research; Academic Press: Cambridge, MA, USA, 2016; pp. 135–206. ISBN 9780081009789. [Google Scholar]
- Rader, R.; Bartomeus, I.; Garibaldi, L.A.; Garratt, M.P.D.; Howlett, B.G.; Winfree, R.; Cunningham, S.A.; Mayfield, M.M.; Arthur, A.D.; Andersson, G.K.S.; et al. Non-bee insects are important contributors to global crop pollination. Proc. Natl. Acad. Sci. USA 2016, 113, 146–151. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bruce, T.J.A. Interplay between insects and plants: Dynamic and complex interactions that have coevolved over millions of years but act in milliseconds. J. Exp. Bot. 2015, 66, 455–465. [Google Scholar] [CrossRef] [PubMed]
- Bouwmeester, H.; Schuurink, R.C.; Bleeker, P.M.; Schiestl, F. The role of volatiles in plant communication. Plant J. 2019, 100, 892–907. [Google Scholar] [CrossRef] [Green Version]
- Lefebvre, F.A.; Lécuyer, E. Small Luggage for a Long Journey: Transfer of Vesicle-Enclosed Small RNA in Interspecies Communication. Front. Microbiol. 2017, 8, 1–15. [Google Scholar] [CrossRef] [Green Version]
- Zeng, J.; Gupta, V.K.; Jiang, Y.; Yang, B.; Gong, L.; Zhu, H. Cross-Kingdom Small RNAs Among Animals, Plants and Microbes. Cells 2019, 8, 371. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ham, B.-K.; Lucas, W.J. Phloem-Mobile RNAs as Systemic Signaling Agents. Annu. Rev. Plant Biol. 2017, 68, 173–195. [Google Scholar] [CrossRef]
- Liu, L.; Chen, X. Intercellular and systemic trafficking of RNAs in plants. Nat. Plants 2018, 4, 869–878. [Google Scholar] [CrossRef]
- Lotvall, J.; Valadi, H. Cell to Cell Signalling via Exosomes Through esRNA. Cell Adh. Migr. 2007, 1, 156–158. [Google Scholar] [CrossRef] [PubMed]
- Valadi, H.; Ekström, K.; Bossios, A.; Sjöstrand, M.; Lee, J.J.; Lötvall, J.O. Exosome-mediated transfer of mRNAs and microRNAs is a novel mechanism of genetic exchange between cells. Nat. Cell Biol. 2007, 9, 654–659. [Google Scholar] [CrossRef] [Green Version]
- Ying, W.; Riopel, M.; Bandyopadhyay, G.; Dong, Y.; Birmingham, A.; Seo, J.B.; Ofrecio, J.M.; Wollam, J.; Hernandez-Carretero, A.; Fu, W.; et al. Adipose Tissue Macrophage-Derived Exosomal miRNAs Can Modulate In Vivo and In Vitro Insulin Sensitivity. Cell 2017, 171, 372–384.e12. [Google Scholar] [CrossRef] [Green Version]
- Thomou, T.; Mori, M.A.; Dreyfuss, J.M.; Konishi, M.; Sakaguchi, M.; Wolfrum, C.; Rao, T.N.; Winnay, J.N.; Garcia-Martin, R.; Grinspoon, S.K.; et al. Adipose-derived circulating miRNAs regulate gene expression in other tissues. Nature 2017, 542, 450–455. [Google Scholar] [CrossRef] [PubMed]
- Chen, S.; Tang, Y.; Liu, Y.; Zhang, P.; Lv, L.; Zhang, X.; Jia, L.; Zhou, Y. Exosomes derived from miR-375-overexpressing human adipose mesenchymal stem cells promote bone regeneration. Cell Prolif. 2019, 52, 1–14. [Google Scholar] [CrossRef] [Green Version]
- Abels, E.R.; Maas, S.L.N.; Nieland, L.; Wei, Z.; Cheah, P.S.; Tai, E.; Kolsteeg, C.-J.; Dusoswa, S.A.; Ting, D.T.; Hickman, S.; et al. Glioblastoma-Associated Microglia Reprogramming Is Mediated by Functional Transfer of Extracellular miR-21. Cell Rep. 2019, 28, 3105–3119.e7. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, Q.; Peng, F.; Chen, J. The Role of Exosomal MicroRNAs in the Tumor Microenvironment of Breast Cancer. Int. J. Mol. Sci. 2019, 20, 3884. [Google Scholar] [CrossRef] [Green Version]
- Shen, M.; Dong, C.; Ruan, X.; Yan, W.; Cao, M.; Pizzo, D.; Wu, X.; Yang, L.; Liu, L.; Ren, X.; et al. Chemotherapy-Induced Extracellular Vesicle miRNAs Promote Breast Cancer Stemness by Targeting ONECUT2. Cancer Res. 2019, 79, 3608–3621. [Google Scholar] [CrossRef] [PubMed]
- Ghamloush, F.; Ghayad, S.E.; Rammal, G.; Fahs, A.; Ayoub, A.J.; Merabi, Z.; Harajly, M.; Zalzali, H.; Saab, R. The PAX3-FOXO1 oncogene alters exosome miRNA content and leads to paracrine effects mediated by exosomal miR-486. Sci. Rep. 2019, 9, 14242. [Google Scholar] [CrossRef]
- Wang, L.; Jia, Q.; Xinnong, C.; Xie, Y.; Yang, Y.; Zhang, A.; Liu, R.; Zhuo, Y.; Zhang, J. Role of cardiac progenitor cell-derived exosome-mediated microRNA-210 in cardiovascular disease. J. Cell. Mol. Med. 2019, 23, 7124–7131. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ma, Y.; Li, C.; Huang, Y.; Wang, Y.; Xia, X.; Zheng, J.C. Exosomes released from neural progenitor cells and induced neural progenitor cells regulate neurogenesis through miR-21a. Cell Commun. Signal. 2019, 17, 96. [Google Scholar] [CrossRef] [Green Version]
- Lucero, R.; Zappulli, V.; Sammarco, A.; Murillo, O.D.; Cheah, P.S.; Srinivasan, S.; Tai, E.; Ting, D.T.; Wei, Z.; Roth, M.E.; et al. Glioma-Derived miRNA-Containing Extracellular Vesicles Induce Angiogenesis by Reprogramming Brain Endothelial Cells. Cell Rep. 2020, 30, 2065–2074.e4. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, Y.; Liu, D.; Chen, X.; Li, J.; Li, L.; Bian, Z.; Sun, F.; Lu, J.; Yin, Y.; Cai, X.; et al. Secreted Monocytic miR-150 Enhances Targeted Endothelial Cell Migration. Mol. Cell 2010, 39, 133–144. [Google Scholar] [CrossRef] [Green Version]
- Kosaka, N.; Iguchi, H.; Yoshioka, Y.; Takeshita, F.; Matsuki, Y.; Ochiya, T. Secretory mechanisms and intercellular transfer of microRNAs in living cells. J. Biol. Chem. 2010, 285, 17442–17452. [Google Scholar] [CrossRef] [Green Version]
- Kogure, T.; Lin, W.-L.; Yan, I.K.; Braconi, C.; Patel, T. Intercellular nanovesicle-mediated microRNA transfer: A mechanism of environmental modulation of hepatocellular cancer cell growth. Hepatology 2011, 54, 1237–1248. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mittelbrunn, M.; Gutiérrez-Vázquez, C.; Villarroya-Beltri, C.; González, S.; Sánchez-Cabo, F.; González, M.Á.; Bernad, A.; Sánchez-Madrid, F. Unidirectional transfer of microRNA-loaded exosomes from T cells to antigen-presenting cells. Nat. Commun. 2011, 2, 282. [Google Scholar] [CrossRef] [Green Version]
- Hergenreider, E.; Heydt, S.; Tréguer, K.; Boettger, T.; Horrevoets, A.J.G.G.; Zeiher, A.M.; Scheffer, M.P.; Frangakis, A.S.; Yin, X.; Mayr, M.; et al. Atheroprotective communication between endothelial cells and smooth muscle cells through miRNAs. Nat. Cell Biol. 2012, 14, 249–256. [Google Scholar] [CrossRef]
- Ismail, N.; Wang, Y.; Dakhlallah, D.; Moldovan, L.; Agarwal, K.; Batte, K.; Shah, P.; Wisler, J.; Eubank, T.D.; Tridandapani, S.; et al. Macrophage microvesicles induce macrophage differentiation and miR-223 transfer. Blood 2013, 121, 984–995. [Google Scholar] [CrossRef] [Green Version]
- Ridder, K.; Keller, S.; Dams, M.; Rupp, A.-K.; Schlaudraff, J.; Del Turco, D.; Starmann, J.; Macas, J.; Karpova, D.; Devraj, K.; et al. Extracellular Vesicle-Mediated Transfer of Genetic Information between the Hematopoietic System and the Brain in Response to Inflammation. PLoS Biol. 2014, 12, e1001874. [Google Scholar] [CrossRef]
- Lai, C.P.; Kim, E.Y.; Badr, C.E.; Weissleder, R.; Mempel, T.R.; Tannous, B.A.; Breakefield, X.O. Visualization and tracking of tumour extracellular vesicle delivery and RNA translation using multiplexed reporters. Nat. Commun. 2015, 6, 7029. [Google Scholar] [CrossRef]
- Wang, J.; Silva, M.; Haas, L.A.; Morsci, N.S.; Nguyen, K.C.Q.; Hall, D.H.; Barr, M.M. C. elegans ciliated sensory neurons release extracellular vesicles that function in animal communication. Curr. Biol. 2014, 24, 519–525. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhou, Y.; Wang, X.; Song, M.; He, Z.; Cui, G.; Peng, G.; Dieterich, C.; Antebi, A.; Jing, N.; Shen, Y. A secreted microRNA disrupts autophagy in distinct tissues of Caenorhabditis elegans upon ageing. Nat. Commun. 2019, 10, 4827. [Google Scholar] [CrossRef] [Green Version]
- Nawaz, M.; Malik, M.I.; Zhang, H.; Gebremedhin, M.B.; Cao, J.; Zhou, Y.; Zhou, J. miRNA profile of extracellular vesicles isolated from saliva of Haemaphysalis longicornis tick. Acta Trop. 2020, 212, 105718. [Google Scholar] [CrossRef]
- Maharaj, P.D.; Widen, S.G.; Huang, J.; Wood, T.G.; Thangamani, S. Discovery of Mosquito Saliva MicroRNAs during CHIKV Infection. PLoS Negl. Trop. Dis. 2015, 9, e0003386. [Google Scholar] [CrossRef] [Green Version]
- Arcà, B.; Colantoni, A.; Fiorillo, C.; Severini, F.; Benes, V.; Di Luca, M.; Calogero, R.A.; Lombardo, F. MicroRNAs from saliva of anopheline mosquitoes mimic human endogenous miRNAs and may contribute to vector-host-pathogen interactions. Sci. Rep. 2019, 9, 2955. [Google Scholar] [CrossRef]
- Dhahbi, J.M.; Atamna, H.; Li, R.; Yamakawa, A.; Guerrero, N.; Lam, H.T.; Mote, P.; Spindler, S.R. MicroRNAs Circulate in the Hemolymph of Drosophila and Accumulate Relative to Tissue microRNAs in an Age-Dependent Manner. Genomics Insights 2016, 9, GEI.S38147. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lefebvre, F.A.; Benoit Bouvrette, L.P.; Perras, L.; Blanchet-Cohen, A.; Garnier, D.; Rak, J.; Lécuyer, É. Comparative transcriptomic analysis of human and Drosophila extracellular vesicles. Sci. Rep. 2016, 6, 27680. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tassetto, M.; Kunitomi, M.; Andino, R. Circulating Immune Cells Mediate a Systemic RNAi-Based Adaptive Antiviral Response in Drosophila. Cell 2017, 169, 314–325.e13. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Van den Brande, S.; Gijbels, M.; Wynant, N.; Santos, D.; Mingels, L.; Gansemans, Y.; Van Nieuwerburgh, F.; Vanden Broeck, J. The presence of extracellular microRNAs in the media of cultured Drosophila cells. Sci. Rep. 2018, 8, 17312. [Google Scholar] [CrossRef] [PubMed]
- Mingels, L.; Wynant, N.; Santos, D.; Peeters, P.; Gansemans, Y.; Billen, J.; Van Nieuwerburgh, F.; Vanden Broeck, J. Extracellular vesicles spread the RNA interference signal of Tribolium castaneum TcA cells. Insect Biochem. Mol. Biol. 2020, 122, 103377. [Google Scholar] [CrossRef]
- Kim, H.G.; Kwon, K.; Suh, H.-W.; Lee, S.; Park, K.-H.; Kwon, O.-Y.; Choi, J.-Y. Exosome isolation from hemolymph of Korean rhinoceros beetle, A llomyrina dichotoma (Coleoptera: Scarabaeidae). Entomol. Res. 2015, 45, 339–344. [Google Scholar] [CrossRef]
- Yoon, J.-S.; Kim, K.; Palli, S.R. Double-stranded RNA in exosomes: Potential systemic RNA interference pathway in the Colorado potato beetle, Leptinotarsa decemlineata. J. Asia. Pac. Entomol. 2020, 23, 1160–1164. [Google Scholar] [CrossRef]
- Russell, J.C.; Kim, T.K.; Noori, A.; Merrihew, G.E.; Robbins, J.E.; Golubeva, A.; Wang, K.; MacCoss, M.J.; Kaeberlein, M. Composition of Caenorhabditis elegans extracellular vesicles suggests roles in metabolism, immunity, and aging. GeroScience 2020, 42, 1133–1145. [Google Scholar] [CrossRef] [PubMed]
- Guo, X.; Su, S.; Skogerboe, G.; Dai, S.; Li, W.; Li, Z.; Liu, F.; Ni, R.; Guo, Y.; Chen, S.; et al. Recipe for a Busy Bee: MicroRNAs in Honey Bee Caste Determination. PLoS ONE 2013, 8, e81661. [Google Scholar] [CrossRef] [PubMed]
- Maori, E.; Garbian, Y.; Kunik, V.; Mozes-Koch, R.; Malka, O.; Kalev, H.; Sabath, N.; Sela, I.; Shafir, S. A Transmissible RNA Pathway in Honey Bees. Cell Rep. 2019, 27, 1949–1959.e6. [Google Scholar] [CrossRef] [Green Version]
- Maori, E.; Navarro, I.C.; Boncristiani, H.; Seilly, D.J.; Rudolph, K.L.M.; Sapetschnig, A.; Lin, C.-C.C.; Ladbury, J.E.; Evans, J.D.; Heeney, J.L.; et al. A Secreted RNA Binding Protein Forms RNA-Stabilizing Granules in the Honeybee Royal Jelly. Mol. Cell 2019, 74, 598–608.e6. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- LeBoeuf, A.C.; Waridel, P.; Brent, C.S.; Gonçalves, A.N.; Menin, L.; Ortiz, D.; Riba-Grognuz, O.; Koto, A.; Soares, Z.G.; Privman, E.; et al. Oral transfer of chemical cues, growth proteins and hormones in social insects. Elife 2016, 5, 1–28. [Google Scholar] [CrossRef]
- Wang, Z.; Ye, X.; Shi, M.; Li, F.; Wang, Z.; Zhou, Y.; Gu, Q.; Wu, X.; Yin, C.; Guo, D.; et al. Parasitic insect-derived miRNAs modulate host development. Nat. Commun. 2018, 9, 2205. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Quintana, J.F.; Kumar, S.; Ivens, A.; Chow, F.W.N.; Hoy, A.M.; Fulton, A.; Dickinson, P.; Martin, C.; Taylor, M.; Babayan, S.A.; et al. Comparative analysis of small RNAs released by the filarial nematode Litomosoides sigmodontis in vitro and in vivo. PLoS Negl. Trop. Dis. 2019, 13, e0007811. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Duguet, T.B.; Soichot, J.; Kuzyakiv, R.; Malmström, L.; Tritten, L. Extracellular Vesicle-Contained microRNA of C. elegans as a Tool to Decipher the Molecular Basis of Nematode Parasitism. Front. Cell. Infect. Microbiol. 2020, 10, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Buck, A.H.; Coakley, G.; Simbari, F.; McSorley, H.J.; Quintana, J.F.; Le Bihan, T.; Kumar, S.; Abreu-Goodger, C.; Lear, M.; Harcus, Y.; et al. Exosomes secreted by nematode parasites transfer small RNAs to mammalian cells and modulate innate immunity. Nat. Commun. 2014, 5, 5488. [Google Scholar] [CrossRef]
- Zamanian, M.; Fraser, L.M.; Agbedanu, P.N.; Harischandra, H.; Moorhead, A.R.; Day, T.A.; Bartholomay, L.C.; Kimber, M.J. Release of Small RNA-containing Exosome-like Vesicles from the Human Filarial Parasite Brugia malayi. PLoS Negl. Trop. Dis. 2015, 9, e0004069. [Google Scholar] [CrossRef] [Green Version]
- Chow, F.W.-N.; Koutsovoulos, G.; Ovando-Vázquez, C.; Neophytou, K.; Bermúdez-Barrientos, J.R.; Laetsch, D.R.; Robertson, E.; Kumar, S.; Claycomb, J.M.; Blaxter, M.; et al. Secretion of an Argonaute protein by a parasitic nematode and the evolution of its siRNA guides. Nucleic Acids Res. 2019, 47, 3594–3606. [Google Scholar] [CrossRef] [PubMed]
- White, R.; Kumar, S.; Chow, F.W.-N.; Robertson, E.; Hayes, K.S.; Grencis, R.K.; Duque-Correa, M.A.; Buck, A.H. Extracellular vesicles from Heligmosomoides bakeri and Trichuris muris contain distinct microRNA families and small RNAs that could underpin different functions in the host. Int. J. Parasitol. 2020, 50, 719–729. [Google Scholar] [CrossRef] [PubMed]
- Yang, H.; Li, X.; Ji, J.; Yuan, C.; Gao, X.; Zhang, Y.; Lu, C.; Li, F.; Zhang, X. Changes of microRNAs expression profiles from red swamp crayfish (Procambarus clarkia) hemolymph exosomes in response to WSSV infection. Fish Shellfish Immunol. 2019, 84, 169–177. [Google Scholar] [CrossRef]
- Hackenberg, M.; Langenberger, D.; Schwarz, A.; Erhart, J.; Kotsyfakis, M. In silico target network analysis of de novo-discovered, tick saliva-specific microRNAs reveals important combinatorial effects in their interference with vertebrate host physiology. RNA 2017, 23, 1259–1269. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Roney, J.K.; Khatibi, P.A.; Westwood, J.H. Cross-Species Translocation of mRNA from Host Plants into the Parasitic Plant Dodder. Plant Physiol. 2007, 143, 1037–1043. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- David-Schwartz, R.; Runo, S.; Townsley, B.; MacHuka, J.; Sinha, N. Long-distance transport of mRNA via parenchyma cells and phloem across the host-parasite junction in Cuscuta. New Phytol. 2008, 1133–1141. [Google Scholar] [CrossRef] [PubMed]
- Tomilov, A.A.; Tomilova, N.B.; Wroblewski, T.; Michelmore, R.; Yoder, J.I. Trans-specific gene silencing between host and parasitic plants. Plant J. 2008, 56, 389–397. [Google Scholar] [CrossRef] [PubMed]
- Aly, R.; Cholakh, H.; Joel, D.M.; Leibman, D.; Steinitz, B.; Zelcer, A.; Naglis, A.; Yarden, O.; Gal-On, A. Gene silencing of mannose 6-phosphate reductase in the parasitic weed Orobanche aegyptiaca through the production of homologous dsRNA sequences in the host plant. Plant Biotechnol. J. 2009, 7, 487–498. [Google Scholar] [CrossRef]
- Alakonya, A.; Kumar, R.; Koenig, D.; Kimura, S.; Townsley, B.; Runo, S.; Garces, H.M.; Kang, J.; Yanez, A.; David-Schwartz, R.; et al. Interspecific RNA Interference of SHOOT MERISTEMLESS-Like Disrupts Cuscuta pentagona Plant Parasitism. Plant Cell 2012, 24, 3153–3166. [Google Scholar] [CrossRef] [Green Version]
- Kim, G.; LeBlanc, M.L.; Wafula, E.K.; DePamphilis, C.W.; Westwood, J.H. Genomic-scale exchange of mRNA between a parasitic plant and its hosts. Science 2014, 345, 808–811. [Google Scholar] [CrossRef] [PubMed]
- Kim, G.; Westwood, J.H. Macromolecule exchange in Cuscuta–host plant interactions. Curr. Opin. Plant Biol. 2015, 26, 20–25. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Westwood, J.H.; Kim, G. RNA mobility in parasitic plant—Host interactions. RNA Biol. 2017, 14, 450–455. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shahid, S.; Kim, G.; Johnson, N.R.; Wafula, E.; Wang, F.; Coruh, C.; Bernal-Galeano, V.; Phifer, T.; DePamphilis, C.W.; Westwood, J.H.; et al. MicroRNAs from the parasitic plant Cuscuta campestris target host messenger RNAs. Nature 2018, 553, 82–85. [Google Scholar] [CrossRef]
- Li, S.; Wang, X.; Xu, W.; Liu, T.; Cai, C.; Chen, L.; Clark, C.B.; Ma, J. Unidirectional movement of small RNAs from shoots to roots in interspecific heterografts. Nat. Plants 2021, 7, 50–59. [Google Scholar] [CrossRef] [PubMed]
- Hou, Y.; Zhai, Y.; Feng, L.; Karimi, H.Z.; Rutter, B.D.; Zeng, L.; Choi, D.S.; Zhang, B.; Gu, W.; Chen, X.; et al. A Phytophthora Effector Suppresses Trans-Kingdom RNAi to Promote Disease Susceptibility. Cell Host Microbe 2019, 25, 153–165.e5. [Google Scholar] [CrossRef] [Green Version]
- Cui, C.; Wang, Y.; Liu, J.; Zhao, J.; Sun, P.; Wang, S. A fungal pathogen deploys a small silencing RNA that attenuates mosquito immunity and facilitates infection. Nat. Commun. 2019, 10, 4298. [Google Scholar] [CrossRef] [PubMed]
- Dunker, F.; Trutzenberg, A.; Rothenpieler, J.S.; Kuhn, S.; Pröls, R.; Schreiber, T.; Tissier, A.; Kemen, A.; Kemen, E.; Hückelhoven, R.; et al. Oomycete small RNAs bind to the plant RNA-induced silencing complex for virulence. Elife 2020, 9, 1–23. [Google Scholar] [CrossRef] [PubMed]
- Weiberg, A.; Wang, M.; Lin, F.-M.; Zhao, H.; Zhang, Z.; Kaloshian, I.; Huang, H.-D.; Jin, H. Fungal Small RNAs Suppress Plant Immunity by Hijacking Host RNA Interference Pathways. Science 2013, 342, 118–123. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cai, Q.; Qiao, L.; Wang, M.; He, B.; Lin, F.-M.; Palmquist, J.; Huang, S.-D.; Jin, H. Plants send small RNAs in extracellular vesicles to fungal pathogen to silence virulence genes. Science 2018, 360, 1126–1129. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kettles, G.J.; Drurey, C.; Schoonbeek, H.; Maule, A.J.; Hogenhout, S.A. Resistance of Arabidopsis thaliana to the green peach aphid, Myzus persicae, involves camalexin and is regulated by microRNAs. New Phytol. 2013, 198, 1178–1190. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- van Kleeff, P.J.M.; Galland, M.; Schuurink, R.C.; Bleeker, P.M. Small RNAs from Bemisia tabaci Are Transferred to Solanum lycopersicum Phloem during Feeding. Front. Plant Sci. 2016, 7, 1–13. [Google Scholar] [CrossRef] [Green Version]
- Chen, Y.; Singh, A.; Kaithakottil, G.G.; Mathers, T.C.; Gravino, M.; Mugford, S.T.; van Oosterhout, C.; Swarbreck, D.; Hogenhout, S.A. An aphid RNA transcript migrates systemically within plants and is a virulence factor. Proc. Natl. Acad. Sci. USA 2020, 117, 12763–12771. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Wiggins, B.; Lawrence, C.; Petrick, J.; Ivashuta, S.; Heck, G. Analysis of plant-derived miRNAs in animal small RNA datasets. BMC Genomics 2012, 13, 381. [Google Scholar] [CrossRef] [Green Version]
- Witwer, K.W.; Hirschi, K.D. Transfer and functional consequences of dietary microRNAs in vertebrates: Concepts in search of corroboration. BioEssays 2014, 36, 394–406. [Google Scholar] [CrossRef] [PubMed]
- Chan, S.Y.; Snow, J.W. Uptake and impact of natural diet-derived small RNA in invertebrates: Implications for ecology and agriculture. RNA Biol. 2017, 14, 402–414. [Google Scholar] [CrossRef] [PubMed]
- Li, Z.; Xu, R.; Li, N. MicroRNAs from plants to animals, do they define a new messenger for communication? Nutr. Metab. 2018, 15, 68. [Google Scholar] [CrossRef]
- Snow, J.W.; Hale, A.E.; Isaacs, S.K.; Baggish, A.L.; Chan, S.Y. Ineffective delivery of diet-derived microRNAs to recipient animal organisms. RNA Biol. 2013, 10, 1107–1116. [Google Scholar] [CrossRef] [Green Version]
- Masood, M.; Everett, C.P.; Chan, S.Y.; Snow, J.W. Negligible uptake and transfer of diet-derived pollen microRNAs in adult honey bees. RNA Biol. 2016, 13, 109–118. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sattar, S.; Addo-Quaye, C.; Song, Y.; Anstead, J.A.; Sunkar, R.; Thompson, G.A. Expression of Small RNA in Aphis gossypii and Its Potential Role in the Resistance Interaction with Melon. PLoS ONE 2012, 7, e48579. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jia, L.; Zhang, D.; Xiang, Z.; He, N. Nonfunctional ingestion of plant miRNAs in silkworm revealed by digital droplet PCR and transcriptome analysis. Sci. Rep. 2015, 5, 12290. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, H.; Zhang, C.; Dou, Y.; Yu, B.; Liu, Y.; Heng-Moss, T.M.; Lu, G.; Wachholtz, M.; Bradshaw, J.D.; Twigg, P.; et al. Insect and plant-derived miRNAs in greenbug (Schizaphis graminum) and yellow sugarcane aphid (Sipha flava) revealed by deep sequencing. Gene 2017, 599, 68–77. [Google Scholar] [CrossRef] [Green Version]
- Thompson, M.C.; Feng, H.; Wuchty, S.; Wilson, A.C.C. The green peach aphid gut contains host plant microRNAs identified by comprehensive annotation of Brassica oleracea small RNA data. Sci. Rep. 2019, 9, 18904. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, L.; Jing, X.; Chen, W.; Wang, Y.; Lin, J.; Zheng, L.; Dong, Y.; Zhou, L.; Li, F.; Yang, F.; et al. Host Plant-Derived miRNAs Potentially Modulate the Development of a Cosmopolitan Insect Pest, Plutella xylostella. Biomolecules 2019, 9, 602. [Google Scholar] [CrossRef] [Green Version]
- Burmester, T. Origin and evolution of arthropod hemocyanins and related proteins. J. Comp. Physiol. B Biochem. Syst. Environ. Physiol. 2002, 172, 95–107. [Google Scholar]
- Zhu, K.; Liu, M.; Fu, Z.; Zhou, Z.; Kong, Y.; Liang, H.; Lin, Z.; Luo, J.; Zheng, H.; Wan, P.; et al. Plant microRNAs in larval food regulate honeybee caste development. PLOS Genet. 2017, 13, e1006946. [Google Scholar] [CrossRef] [PubMed]
- Patel, A.; Fondrk, M.K.; Kaftanoglu, O.; Emore, C.; Hunt, G.; Frederick, K.; Amdam, G.V. The Making of a Queen: TOR Pathway Is a Key Player in Diphenic Caste Development. PLoS ONE 2007, 2, e509. [Google Scholar] [CrossRef] [Green Version]
- Baum, J.A.; Bogaert, T.; Clinton, W.; Heck, G.R.; Feldmann, P.; Ilagan, O.; Johnson, S.; Plaetinck, G.; Munyikwa, T.; Pleau, M.; et al. Control of coleopteran insect pests through RNA interference. Nat. Biotechnol. 2007, 25, 1322–1326. [Google Scholar] [CrossRef]
- Mao, Y.-B.; Cai, W.-J.; Wang, J.-W.; Hong, G.-J.; Tao, X.-Y.; Wang, L.-J.; Huang, Y.-P.; Chen, X.-Y. Silencing a cotton bollworm P450 monooxygenase gene by plant-mediated RNAi impairs larval tolerance of gossypol. Nat. Biotechnol. 2007, 25, 1307–1313. [Google Scholar] [CrossRef]
- Mao, Y.-B.; Tao, X.-Y.; Xue, X.-Y.; Wang, L.-J.; Chen, X.-Y. Cotton plants expressing CYP6AE14 double-stranded RNA show enhanced resistance to bollworms. Transgenic Res. 2011, 20, 665–673. [Google Scholar] [CrossRef] [Green Version]
- Zha, W.; Peng, X.; Chen, R.; Du, B.; Zhu, L.; He, G. Knockdown of Midgut Genes by dsRNA-Transgenic Plant-Mediated RNA Interference in the Hemipteran Insect Nilaparvata lugens. PLoS ONE 2011, 6, e20504. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bhatia, V.; Bhattacharya, R.; Uniyal, P.L.; Singh, R.; Niranjan, R.S. Host Generated siRNAs Attenuate Expression of Serine Protease Gene in Myzus persicae. PLoS ONE 2012, 7, e46343. [Google Scholar] [CrossRef] [Green Version]
- Xiong, Y.; Zeng, H.; Zhang, Y.; Xu, D.; Qiu, D. Silencing the HaHR3 gene by transgenic plant-mediated RNAi to disrupt Helicoverpa armigera development. Int. J. Biol. Sci. 2013, 9, 370–381. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mao, Y.-B.; Xue, X.-Y.; Tao, X.-Y.; Yang, C.-Q.; Wang, L.-J.; Chen, X.-Y. Cysteine protease enhances plant-mediated bollworm RNA interference. Plant Mol. Biol. 2013, 83, 119–129. [Google Scholar] [CrossRef] [Green Version]
- Yu, R.; Xu, X.; Liang, Y.; Tian, H.; Pan, Z.; Jin, S.; Wang, N.; Zhang, W. The Insect Ecdysone Receptor is a Good Potential Target for RNAi-based Pest Control. Int. J. Biol. Sci. 2014, 10, 1171–1180. [Google Scholar] [CrossRef] [Green Version]
- Mao, J.; Zeng, F. Plant-mediated RNAi of a gap gene-enhanced tobacco tolerance against the Myzus persicae. Transgenic Res. 2014, 23, 145–152. [Google Scholar] [CrossRef]
- Thakur, N.; Upadhyay, S.K.; Verma, P.C.; Chandrashekar, K.; Tuli, R.; Singh, P.K. Enhanced Whitefly Resistance in Transgenic Tobacco Plants Expressing Double Stranded RNA of v-ATPase A Gene. PLoS ONE 2014, 9, e87235. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, F.; Wang, X.; Zhao, Y.; Li, Y.; Liu, Y.; Sun, J. Silencing the HaAK Gene by Transgenic Plant-Mediated RNAi Impairs Larval Growth of Helicoverpa armigera. Int. J. Biol. Sci. 2015, 11, 67–74. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jin, S.; Singh, N.D.; Li, L.; Zhang, X.; Daniell, H. Engineered chloroplast dsRNA silences cytochrome p450 monooxygenase, V—ATPase and chitin synthase genes in the insect gut and disrupts Helicoverpa armigera larval development and pupation. Plant Biotechnol. J. 2015, 13, 435–446. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Malik, H.J.; Raza, A.; Amin, I.; Scheffler, J.A.; Scheffler, B.E.; Brown, J.K.; Mansoor, S. RNAi-mediated mortality of the whitefly through transgenic expression of double-stranded RNA homologous to acetylcholinesterase and ecdysone receptor in tobacco plants. Sci. Rep. 2016, 6, 38469. [Google Scholar] [CrossRef] [Green Version]
- Mamta; Reddy, K.R.K.; Rajam, M.V. Targeting chitinase gene of Helicoverpa armigera by host-induced RNA interference confers insect resistance in tobacco and tomato. Plant Mol. Biol. 2016, 90, 281–292. [Google Scholar] [CrossRef] [PubMed]
- Pitino, M.; Coleman, A.D.; Maffei, M.E.; Ridout, C.J.; Hogenhout, S.A. Silencing of Aphid Genes by dsRNA Feeding from Plants. PLoS ONE 2011, 6, e25709. [Google Scholar] [CrossRef] [PubMed]
- Zhu, J.-Q.; Liu, S.; Ma, Y.; Zhang, J.-Q.; Qi, H.-S.; Wei, Z.-J.; Yao, Q.; Zhang, W.-Q.; Li, S. Improvement of Pest Resistance in Transgenic Tobacco Plants Expressing dsRNA of an Insect-Associated Gene EcR. PLoS ONE 2012, 7, e38572. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kumar, P.; Pandit, S.S.; Baldwin, I.T. Tobacco Rattle Virus Vector: A Rapid and Transient Means of Silencing Manduca sexta Genes by Plant Mediated RNA Interference. PLoS ONE 2012, 7, e31347. [Google Scholar] [CrossRef] [Green Version]
- Guo, H.; Song, X.; Wang, G.; Yang, K.; Wang, Y.; Niu, L.; Chen, X.; Fang, R. Plant-Generated Artificial Small RNAs Mediated Aphid Resistance. PLoS ONE 2014, 9, e97410. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Agrawal, A.; Rajamani, V.; Reddy, V.S.; Mukherjee, S.K.; Bhatnagar, R.K. Transgenic plants over-expressing insect-specific microRNA acquire insecticidal activity against Helicoverpa armigera: An alternative to Bt-toxin technology. Transgenic Res. 2015, 24, 791–801. [Google Scholar] [CrossRef]
- Li, T.; Chen, J.; Fan, X.; Chen, W.; Zhang, W. MicroRNA and dsRNA targeting chitin synthase A reveal a great potential for pest management of the hemipteran insect Nilaparvata lugens. Pest Manag. Sci. 2017, 73, 1529–1537. [Google Scholar] [CrossRef]
- Saini, R.P.; Raman, V.; Dhandapani, G.; Malhotra, E.V.; Sreevathsa, R.; Kumar, P.A.; Sharma, T.R.; Pattanayak, D. Silencing of HaAce1 gene by host-delivered artificial microRNA disrupts growth and development of Helicoverpa armigera. PLoS ONE 2018, 13, e0194150. [Google Scholar] [CrossRef] [PubMed]
- Bally, J.; Fishilevich, E.; Doran, R.L.; Lee, K.; Campos, S.B.; German, M.A.; Narva, K.E.; Waterhouse, P.M. Plin-amiR, a pre-microRNA-based technology for controlling herbivorous insect pests. Plant Biotechnol. J. 2020, 18, 1925–1932. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Khan, S.A.; Hasse, C.; Ruf, S.; Heckel, D.G.; Bock, R. Full crop protection from an insect pest by expression of long double-stranded RNAs in plastids. Science 2015, 347, 991–994. [Google Scholar] [CrossRef] [PubMed]
- Bally, J.; McIntyre, G.J.; Doran, R.L.; Lee, K.; Perez, A.; Jung, H.; Naim, F.; Larrinua, I.M.; Narva, K.E.; Waterhouse, P.M. In-Plant Protection against Helicoverpa armigera by Production of Long hpRNA in Chloroplasts. Front. Plant Sci. 2016, 7, 1453. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- He, W.; Xu, W.; Xu, L.; Fu, K.; Guo, W.; Bock, R.; Zhang, J. Length-dependent accumulation of double-stranded RNAs in plastids affects RNA interference efficiency in the Colorado potato beetle. J. Exp. Bot. 2020, 71, 2670–2677. [Google Scholar] [CrossRef]
- Dong, Y.; Yang, Y.; Wang, Z.; Wu, M.; Fu, J.; Guo, J.; Chang, L.; Zhang, J. Inaccessibility to double-stranded RNAs in plastids restricts RNA interference in Bemisia tabaci (whitefly). Pest Manag. Sci. 2020, 76, 3168–3176. [Google Scholar] [CrossRef] [PubMed]
- Head, G.P.; Carroll, M.W.; Evans, S.P.; Rule, D.M.; Willse, A.R.; Clark, T.L.; Storer, N.P.; Flannagan, R.D.; Samuel, L.W.; Meinke, L.J. Evaluation of SmartStax and SmartStax PRO maize against western corn rootworm and northern corn rootworm: Efficacy and resistance management. Pest Manag. Sci. 2017, 73, 1883–1899. [Google Scholar] [CrossRef] [Green Version]
- Nazim Uddin, M.; Kim, J.Y. Intercellular and systemic spread of RNA and RNAi in plants. Wiley Interdiscip. Rev. RNA 2013, 4, 279–293. [Google Scholar] [CrossRef] [PubMed]
- Mermigka, G.; Verret, F.; Kalantidis, K. RNA silencing movement in plants. J. Integr. Plant Biol. 2016, 58, 328–342. [Google Scholar] [CrossRef] [PubMed]
- Wang, M.; Dean, R.A. Movement of small RNAs in and between plants and fungi. Mol. Plant Pathol. 2020, 21, 589–601. [Google Scholar] [CrossRef] [PubMed]
- Hyun, T.K.; Uddin, M.N.; Rim, Y.; Kim, J.-Y. Cell-to-cell trafficking of RNA and RNA silencing through plasmodesmata. Protoplasma 2011, 248, 101–116. [Google Scholar] [CrossRef]
- Melnyk, C.W.; Molnar, A.; Baulcombe, D.C. Intercellular and systemic movement of RNA silencing signals. EMBO J. 2011, 30, 3553–3563. [Google Scholar] [CrossRef]
- Zhang, S.; Hong, Z. Mobile RNAs—the magical elf traveling between plant and the associated organisms. ExRNA 2019, 1, 8. [Google Scholar] [CrossRef] [Green Version]
- Valiunas, V.; Polosina, Y.Y.; Miller, H.; Potapova, I.A.; Valiuniene, L.; Doronin, S.; Mathias, R.T.; Robinson, R.B.; Rosen, M.R.; Cohen, I.S.; et al. Connexin-specific cell-to-cell transfer of short interfering RNA by gap junctions. J. Physiol. 2005, 568, 459–468. [Google Scholar] [CrossRef] [Green Version]
- Katakowski, M.; Buller, B.; Wang, X.; Rogers, T.; Chopp, M. Functional MicroRNA Is Transferred between Glioma Cells. Cancer Res. 2010, 70, 8259–8263. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lim, P.K.; Bliss, S.A.; Patel, S.A.; Taborga, M.; Dave, M.A.; Gregory, L.A.; Greco, S.J.; Bryan, M.; Patel, P.S.; Rameshwar, P. Gap Junction–Mediated Import of MicroRNA from Bone Marrow Stromal Cells Can Elicit Cell Cycle Quiescence in Breast Cancer Cells. Cancer Res. 2011, 71, 1550–1560. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aucher, A.; Rudnicka, D.; Davis, D.M. MicroRNAs Transfer from Human Macrophages to Hepato-Carcinoma Cells and Inhibit Proliferation. J. Immunol. 2013, 191, 6250–6260. [Google Scholar] [CrossRef] [PubMed]
- Hong, X.; Sin, W.C.; Harris, A.L.; Naus, C.C. Gap junctions modulate glioma invasion by direct transfer of microRNA. Oncotarget 2015, 6, 15566–15577. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhu, Y.; Zong, L.; Mei, L.; Zhao, H.-B. Connexin26 gap junction mediates miRNA intercellular genetic communication in the cochlea and is required for inner ear development. Sci. Rep. 2015, 5, 15647. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zong, L.; Zhu, Y.; Liang, R.; Zhao, H.-B. Gap junction mediated miRNA intercellular transfer and gene regulation: A novel mechanism for intercellular genetic communication. Sci. Rep. 2016, 6, 19884. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Thuringer, D.; Boucher, J.; Jego, G.; Pernet, N.; Cronier, L.; Hammann, A.; Solary, E.; Garrido, C. Transfer of functional microRNAs between glioblastoma and microvascular endothelial cells through gap junctions. Oncotarget 2016, 7, 73925–73934. [Google Scholar] [CrossRef] [Green Version]
- Thuringer, D.; Jego, G.; Berthenet, K.; Hammann, A.; Solary, E.; Garrido, C. Gap junction-mediated transfer of miR-145-5p from microvascular endothelial cells to colon cancer cells inhibits angiogenesis. Oncotarget 2016, 7, 28160–28168. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fan, X.; Teng, Y.; Ye, Z.; Zhou, Y.; Tan, W.-S. The effect of gap junction-mediated transfer of miR-200b on osteogenesis and angiogenesis in a co-culture of MSCs and HUVECs. J. Cell Sci. 2018, 131, jcs216135. [Google Scholar] [CrossRef] [Green Version]
- Winston, W.M.; Molodowitch, C.; Hunter, C.P. Systemic RNAi in C. elegans requires the putative transmembrane protein SID-1. Science 2002, 295, 2456–2459. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Feinberg, E.H.; Hunter, C.P. Transport of dsRNA into cells by the transmembrane protein SID-1. Science 2003, 301, 1545–1547. [Google Scholar] [CrossRef] [Green Version]
- Winston, W.M.; Sutherlin, M.; Wright, A.J.; Feinberg, E.H.; Hunter, C.P. Caenorhabditis elegans SID-2 is required for environmental RNA interference. Proc. Natl. Acad. Sci. USA 2007, 104, 10565–10570. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hinas, A.; Wright, A.J.; Hunter, C.P. SID-5 Is an Endosome-Associated Protein Required for Efficient Systemic RNAi in C. elegans. Curr. Biol. 2012, 22, 1938–1943. [Google Scholar] [CrossRef] [Green Version]
- Karlikow, M.; Goic, B.; Mongelli, V.; Salles, A.; Schmitt, C.; Bonne, I.; Zurzolo, C.; Saleh, M.C. Drosophila cells use nanotube-like structures to transfer dsRNA and RNAi machinery between cells. Sci. Rep. 2016, 6, 1–9. [Google Scholar] [CrossRef] [Green Version]
- Ellendorff, U.; Fradin, E.F.; de Jonge, R.; Thomma, B.P.H.J. RNA silencing is required for Arabidopsis defence against Verticillium wilt disease. J. Exp. Bot. 2009, 60, 591–602. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, M.; Weiberg, A.; Lin, F.M.; Thomma, B.P.H.J.; Huang, H.D.; Jin, H. Bidirectional cross-kingdom RNAi and fungal uptake of external RNAs confer plant protection. Nat. Plants 2016, 2, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Chung, K.P.; Lin, W.; Jiang, L. Protein secretion in plants: Conventional and unconventional pathways and new techniques. J. Exp. Bot. 2018, 69, 21–37. [Google Scholar] [CrossRef] [Green Version]
- Mitchell, P.S.; Parkin, R.K.; Kroh, E.M.; Fritz, B.R.; Wyman, S.K.; Pogosova-Agadjanyan, E.L.; Peterson, A.; Noteboom, J.; O’Briant, K.C.; Allen, A.; et al. Circulating microRNAs as stable blood-based markers for cancer detection. Proc. Natl. Acad. Sci. USA 2008, 105, 10513–10518. [Google Scholar] [CrossRef] [Green Version]
- Garbutt, J.S.; Bellés, X.; Richards, E.H.; Reynolds, S.E. Persistence of double-stranded RNA in insect hemolymph as a potential determiner of RNA interference success: Evidence from Manduca sexta and Blattella germanica. J. Insect Physiol. 2013, 59, 171–178. [Google Scholar] [CrossRef]
- Wynant, N.; Santos, D.; Verdonck, R.; Spit, J.; Van Wielendaele, P.; Vanden Broeck, J. Identification, functional characterization and phylogenetic analysis of double stranded RNA degrading enzymes present in the gut of the desert locust, Schistocerca gregaria. Insect Biochem. Mol. Biol. 2014, 46, 1–8. [Google Scholar] [CrossRef]
- Wynant, N.; Duressa, T.F.; Santos, D.; Van Duppen, J.; Proost, P.; Huybrechts, R.; Vanden Broeck, J. Lipophorins can adhere to dsRNA, bacteria and fungi present in the hemolymph of the desert locust: A role as general scavenger for pathogens in the open body cavity. J. Insect Physiol. 2014, 64, 7–13. [Google Scholar] [CrossRef]
- Wynant, N.; Santos, D.; Vanden Broeck, J. Biological Mechanisms Determining the Success of RNA Interference in Insects. Int. Rev. Cell Mol. Biol. 2014, 312, 139–167. [Google Scholar]
- Peng, Y.; Wang, K.; Fu, W.; Sheng, C.; Han, Z. Biochemical Comparison of dsRNA Degrading Nucleases in Four Different Insects. Front. Physiol. 2018, 9, 1–14. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yoo, B.C.; Kragler, F.; Varkonyi-Gasic, E.; Haywood, V.; Archer-Evans, S.; Lee, Y.M.; Lough, T.J.; Lucas, W.J. A systematic small RNA signaling system in plants. Plant Cell 2004, 16, 1979–2000. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ham, B.K.; Li, G.; Jia, W.; Leary, J.A.; Lucas, W.J. Systemic delivery of siRNA in pumpkin by a plant PHLOEM SMALL RNA-BINDING PROTEIN 1-ribonucleoprotein complex. Plant J. 2014, 80, 683–694. [Google Scholar] [CrossRef]
- Barnes, A.; Bale, J.; Constantinidou, C.; Ashton, P.; Jones, A.; Pritchard, J. Determining protein identity from sieve element sap in Ricinus communis L. by quadrupole time of flight (Q-TOF) mass spectrometry. J. Exp. Bot. 2004, 55, 1473–1481. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gómez, G.; Pallás, V. A Long-Distance Translocatable Phloem Protein from Cucumber Forms a Ribonucleoprotein Complex In Vivo with Hop Stunt Viroid RNA. J. Virol. 2004, 78, 10104–10110. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Giavalisco, P.; Kapitza, K.; Kolasa, A.; Buhtz, A.; Kehr, J. Towards the proteome ofBrassica napus phloem sap. Proteomics 2006, 6, 896–909. [Google Scholar] [CrossRef]
- Marín-González, E.; Suárez-López, P. “And yet it moves”: Cell-to-cell and long-distance signaling by plant microRNAs. Plant Sci. 2012, 196, 18–30. [Google Scholar] [CrossRef] [PubMed]
- He, B.; Cai, Q.; Qiao, L.; Huang, C.-Y.; Wang, S.; Miao, W.; Ha, T.; Wang, Y.; Jin, H. RNA-binding proteins contribute to small RNA loading in plant extracellular vesicles. Nat. Plants 2021, 1–22. [Google Scholar]
- Yan, Y.; Ham, B.-K.; Chong, Y.H.; Yeh, S.-D.; Lucas, W.J. A Plant SMALL RNA-BINDING PROTEIN 1 Family Mediates Cell-to-Cell Trafficking of RNAi Signals. Mol. Plant 2020, 13, 321–335. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Turchinovich, A.; Weiz, L.; Langheinz, A.; Burwinkel, B. Characterization of extracellular circulating microRNA. Nucleic Acids Res. 2011, 39, 7223–7233. [Google Scholar] [CrossRef] [PubMed]
- Arroyo, J.D.; Chevillet, J.R.; Kroh, E.M.; Ruf, I.K.; Pritchard, C.C.; Gibson, D.F.; Mitchell, P.S.; Bennett, C.F.; Pogosova-Agadjanyan, E.L.; Stirewalt, D.L.; et al. Argonaute2 complexes carry a population of circulating microRNAs independent of vesicles in human plasma. Proc. Natl. Acad. Sci. USA 2011, 108, 5003–5008. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Turchinovich, A.; Burwinkel, B. Distinct AGO1 and AGO2 associated miRNA profiles in human cells and blood plasma. RNA Biol. 2012, 9, 1066–1075. [Google Scholar] [CrossRef] [Green Version]
- Prud’homme, G.J.; Glinka, Y.; Lichner, Z.; Yousef, G.M. Neuropilin-1 is a receptor for extracellular miRNA and AGO2/miRNA complexes and mediates the internalization of miRNAs that modulate cell function. Oncotarget 2016, 7, 68057–68071. [Google Scholar] [CrossRef] [Green Version]
- Turchinovich, A.; Tonevitsky, A.G.; Cho, W.C.; Burwinkel, B. Check and mate to exosomal extracellular miRNA: New lesson from a new approach. Front. Mol. Biosci. 2015, 2, 10–12. [Google Scholar] [CrossRef] [Green Version]
- Vickers, K.C.; Palmisano, B.T.; Shoucri, B.M.; Shamburek, R.D.; Remaley, A.T. MicroRNAs are transported in plasma and delivered to recipient cells by high-density lipoproteins. Nat. Cell Biol. 2011, 13, 423–433. [Google Scholar] [CrossRef] [Green Version]
- Vickers, K.C.; Remaley, A.T. Lipid-based carriers of microRNAs and intercellular communication. Curr. Opin. Lipidol. 2012, 23, 91–97. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wagner, J.; Riwanto, M.; Besler, C.; Knau, A.; Fichtlscherer, S.; Röxe, T.; Zeiher, A.M.; Landmesser, U.; Dimmeler, S. Characterization of levels and cellular transfer of circulating lipoprotein-bound microRNAs. Arterioscler. Thromb. Vasc. Biol. 2013, 33, 1392–1400. [Google Scholar] [CrossRef] [Green Version]
- Tabet, F.; Vickers, K.C.; Cuesta Torres, L.F.; Wiese, C.B.; Shoucri, B.M.; Lambert, G.; Catherinet, C.; Prado-Lourenco, L.; Levin, M.G.; Thacker, S.; et al. HDL-transferred microRNA-223 regulates ICAM-1 expression in endothelial cells. Nat. Commun. 2014, 5, 3292. [Google Scholar] [CrossRef] [Green Version]
- Michell, D.L.; Vickers, K.C. Lipoprotein carriers of microRNAs. Biochim. Biophys. Acta Mol. Cell Biol. Lipids 2016, 1861, 2069–2074. [Google Scholar] [CrossRef] [Green Version]
- Michell, D.L.; Vickers, K.C. HDL and microRNA therapeutics in cardiovascular disease. Pharmacol. Ther. 2016, 168, 43–52. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ben-Aicha, S.; Escate, R.; Casaní, L.; Padró, T.; Peña, E.; Arderiu, G.; Mendieta, G.; Badimón, L.; Vilahur, G. High-density lipoprotein remodelled in hypercholesterolaemic blood induce epigenetically driven down-regulation of endothelial HIF-1α expression in a preclinical animal model. Cardiovasc. Res. 2020, 116, 1288–1299. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sakashita, K.; Tatsuke, T.; Masaki, Y.; Lee, J.M.; Kawaguch, Y.; Kusakabe, T. DsRNA binding activity of silkworm larval hemolymph is mediated by lipophorin complex. J. Fac. Agric. Kyushu Univ. 2009, 54, 401–406. [Google Scholar] [CrossRef]
- Banks, T.M.; Wang, T.; Fitzgibbon, Q.P.; Smith, G.G.; Ventura, T. Double-Stranded RNA Binding Proteins in Serum Contribute to Systemic RNAi Across Phyla—Towards Finding the Missing Link in Achelata. Int. J. Mol. Sci. 2020, 21, 6967. [Google Scholar] [CrossRef] [PubMed]
- Wang, K.; Zhang, S.; Weber, J.; Baxter, D.; Galas, D.J. Export of microRNAs and microRNA-protective protein by mammalian cells. Nucleic Acids Res. 2010, 38, 7248–7259. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vidal, M. Exosomes: Revisiting their role as “garbage bags”. Traffic 2019, 20, 815–828. [Google Scholar] [CrossRef] [PubMed]
- Hansen, L.L.; Nielsen, M.E. Plant exosomes: Using an unconventional exit to prevent pathogen entry? J. Exp. Bot. 2018, 69, 59–68. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cui, Y.; Gao, J.; He, Y.; Jiang, L. Plant extracellular vesicles. Protoplasma 2020, 257, 3–12. [Google Scholar] [CrossRef] [PubMed]
- Rutter, B.D.; Innes, R.W. Extracellular Vesicles Isolated from the Leaf Apoplast Carry Stress-Response Proteins. Plant Physiol. 2017, 173, 728–741. [Google Scholar] [CrossRef] [Green Version]
- Rybak, K.; Robatzek, S. Functions of Extracellular Vesicles in Immunity and Virulence. Plant Physiol. 2019, 179, 1236–1247. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Limpens, E. Extracellular membranes in symbiosis. Nat. Plants 2019, 5, 131–132. [Google Scholar] [CrossRef] [PubMed]
- Rutter, B.D.; Innes, R.W. Growing pains: Addressing the pitfalls of plant extracellular vesicle research. New Phytol. 2020, 228, 1505–1510. [Google Scholar] [CrossRef]
- Baldrich, P.; Rutter, B.D.; Karimi, H.Z.; Podicheti, R.; Meyers, B.C.; Innes, R.W. Plant Extracellular Vesicles Contain Diverse Small RNA Species and Are Enriched in 10- to 17-Nucleotide “Tiny” RNAs. Plant Cell 2019, 31, 315–324. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lv, Z.; Wei, Y.; Wang, D.; Zhang, C.-Y.; Zen, K.; Li, L. Argonaute 2 in Cell-Secreted Microvesicles Guides the Function of Secreted miRNAs in Recipient Cells. PLoS ONE 2014, 9, e103599. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Varcianna, A.; Myszczynska, M.A.; Castelli, L.M.; O’Neill, B.; Kim, Y.; Talbot, J.; Nyberg, S.; Nyamali, I.; Heath, P.R.; Stopford, M.J.; et al. Micro-RNAs secreted through astrocyte-derived extracellular vesicles cause neuronal network degeneration in C9orf72 ALS. EBioMedicine 2019, 40, 626–635. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Parolini, I.; Federici, C.; Raggi, C.; Lugini, L.; Palleschi, S.; De Milito, A.; Coscia, C.; Iessi, E.; Logozzi, M.; Molinari, A.; et al. Microenvironmental pH Is a Key Factor for Exosome Traffic in Tumor Cells. J. Biol. Chem. 2009, 284, 34211–34222. [Google Scholar] [CrossRef] [Green Version]
- O’Brien, K.; Breyne, K.; Ughetto, S.; Laurent, L.C.; Breakefield, X.O. RNA delivery by extracellular vesicles in mammalian cells and its applications. Nat. Rev. Mol. Cell Biol. 2020, 21, 585–606. [Google Scholar] [CrossRef]
- Danielson, K.M.; Shah, R.; Yeri, A.; Liu, X.; Camacho Garcia, F.; Silverman, M.; Tanriverdi, K.; Das, A.; Xiao, C.; Jerosch-Herold, M.; et al. Plasma Circulating Extracellular RNAs in Left Ventricular Remodeling Post-Myocardial Infarction. EBioMedicine 2018, 32, 172–181. [Google Scholar] [CrossRef]
- Hoshino, A.; Costa-Silva, B.; Shen, T.-L.; Rodrigues, G.; Hashimoto, A.; Tesic Mark, M.; Molina, H.; Kohsaka, S.; Di Giannatale, A.; Ceder, S.; et al. Tumour exosome integrins determine organotropic metastasis. Nature 2015, 527, 329–335. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yu, H.; Wang, Z. Cardiomyocyte-Derived Exosomes: Biological Functions and Potential Therapeutic Implications. Front. Physiol. 2019, 10, 1049. [Google Scholar] [CrossRef] [PubMed]
- Wan, B.; Goguet, E.; Ravallec, M.; Pierre, O.; Lemauf, S.; Volkoff, A.-N.; Gatti, J.-L.; Poirié, M. Venom Atypical Extracellular Vesicles as Interspecies Vehicles of Virulence Factors Involved in Host Specificity: The Case of a Drosophila Parasitoid Wasp. Front. Immunol. 2019, 10, 1–14. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schuh, C.M.A.P.; Aguayo, S.; Zavala, G.; Khoury, M. Exosome-like vesicles in Apis mellifera bee pollen, honey and royal jelly contribute to their antibacterial and pro-regenerative activity. J. Exp. Biol. 2019, 222, jeb208702. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guduric-Fuchs, J.; O’Connor, A.; Camp, B.; O’Neill, C.L.; Medina, R.J.; Simpson, D.A. Selective extracellular vesicle-mediated export of an overlapping set of microRNAs from multiple cell types. BMC Genom. 2012, 13, 357. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Villarroya-Beltri, C.; Gutiérrez-Vázquez, C.; Sánchez-Cabo, F.; Pérez-Hernández, D.; Vázquez, J.; Martin-Cofreces, N.; Martinez-Herrera, D.J.; Pascual-Montano, A.; Mittelbrunn, M.; Sánchez-Madrid, F. Sumoylated hnRNPA2B1 controls the sorting of miRNAs into exosomes through binding to specific motifs. Nat. Commun. 2013, 4, 2980. [Google Scholar] [CrossRef] [Green Version]
- Shurtleff, M.J.; Temoche-Diaz, M.M.; Karfilis, K.V.; Ri, S.; Schekman, R. Y-box protein 1 is required to sort microRNAs into exosomes in cells and in a cell-free reaction. Elife 2016, 5, 1–23. [Google Scholar] [CrossRef]
- Santangelo, L.; Giurato, G.; Cicchini, C.; Montaldo, C.; Mancone, C.; Tarallo, R.; Battistelli, C.; Alonzi, T.; Weisz, A.; Tripodi, M. The RNA-Binding Protein SYNCRIP Is a Component of the Hepatocyte Exosomal Machinery Controlling MicroRNA Sorting. Cell Rep. 2016, 17, 799–808. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Statello, L.; Maugeri, M.; Garre, E.; Nawaz, M.; Wahlgren, J.; Papadimitriou, A.; Lundqvist, C.; Lindfors, L.; Collén, A.; Sunnerhagen, P.; et al. Identification of RNA-binding proteins in exosomes capable of interacting with different types of RNA: RBP-facilitated transport of RNAs into exosomes. PLoS ONE 2018, 13, e0195969. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Temoche-Diaz, M.M.; Shurtleff, M.J.; Nottingham, R.M.; Yao, J.; Fadadu, R.P.; Lambowitz, A.M.; Schekman, R. Distinct mechanisms of microRNA sorting into cancer cell-derived extracellular vesicle subtypes. Elife 2019, 8, 1–34. [Google Scholar] [CrossRef] [PubMed]
- Zietzer, A.; Hosen, M.R.; Wang, H.; Goody, P.R.; Sylvester, M.; Latz, E.; Nickenig, G.; Werner, N.; Jansen, F. The RNA-binding protein hnRNPU regulates the sorting of microRNA-30c-5p into large extracellular vesicles. J. Extracell. Vesicles 2020, 9, 1786967. [Google Scholar] [CrossRef] [PubMed]
- McKenzie, A.J.; Hoshino, D.; Hong, N.H.; Cha, D.J.; Franklin, J.L.; Coffey, R.J.; Patton, J.G.; Weaver, A.M. KRAS-MEK Signaling Controls Ago2 Sorting into Exosomes. Cell Rep. 2016, 15, 978–987. [Google Scholar] [CrossRef] [Green Version]
- Wei, J.; Lv, L.; Wan, Y.; Cao, Y.; Li, G.; Lin, H.; Zhou, R.; Shang, C.; Cao, J.; He, H.; et al. Vps4A functions as a tumor suppressor by regulating the secretion and uptake of exosomal microRNAs in human hepatoma cells. Hepatology 2015, 61, 1284–1294. [Google Scholar] [CrossRef] [PubMed]
- Jackson, C.E.; Scruggs, B.S.; Schaffer, J.E.; Hanson, P.I. Effects of Inhibiting VPS4 Support a General Role for ESCRTs in Extracellular Vesicle Biogenesis. Biophys. J. 2017, 113, 1342–1352. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, H.; Li, C.; Zhang, Y.; Zhang, D.; Otterbein, L.E.; Jin, Y. Caveolin-1 selectively regulates microRNA sorting into microvesicles after noxious stimuli. J. Exp. Med. 2019, 216, 2202–2220. [Google Scholar] [CrossRef]
- Koppers-Lalic, D.; Hackenberg, M.; Bijnsdorp, I.V.; van Eijndhoven, M.A.J.; Sadek, P.; Sie, D.; Zini, N.; Middeldorp, J.M.; Ylstra, B.; de Menezes, R.X.; et al. Nontemplated Nucleotide Additions Distinguish the Small RNA Composition in Cells from Exosomes. Cell Rep. 2014, 8, 1649–1658. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gill, S.; Catchpole, R.; Forterre, P. Extracellular membrane vesicles in the three domains of life and beyond. FEMS Microbiol. Rev. 2019, 273–303. [Google Scholar] [CrossRef]
- Tsatsaronis, J.A.; Franch-Arroyo, S.; Resch, U.; Charpentier, E. Extracellular Vesicle RNA: A Universal Mediator of Microbial Communication? Trends Microbiol. 2018, 26, 401–410. [Google Scholar] [CrossRef]
- Cai, Q.; He, B.; Weiberg, A.; Buck, A.H.; Jin, H. Small RNAs and extracellular vesicles: New mechanisms of cross-species communication and innovative tools for disease control. PLOS Pathog. 2019, 15, e1008090. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bielska, E.; May, R.C. Extracellular vesicles of human Pathogenic fungi. Curr. Opin. Microbiol. 2019, 52, 90–99. [Google Scholar] [CrossRef]
- Sharma, S.; Kooner, R.; Arora, R. Insect pests and crop losses. In Breeding Insect Resistant Crops for Sustainable Agriculture; Springer: Singapore, 2017; pp. 45–66. ISBN 9789811060564. [Google Scholar]
- Ansari, M.S.; Moraiet, M.A.; Ahmad, S. Insecticides: Impact on the Environment and Human Health. In Environmental Deterioration and Human Health; Springer Netherlands: Dordrecht, The Netherlands, 2014; Volume 9789400778, pp. 99–123. ISBN 9789400778900. [Google Scholar]
- Ndakidemi, B.; Mtei, K.; Ndakidemi, P.A. Impacts of Synthetic and Botanical Pesticides on Beneficial Insects. Agric. Sci. 2016, 07, 364–372. [Google Scholar] [CrossRef] [Green Version]
- Caldas, E.D. Toxicological Aspects of Pesticides. In Sustainable Agrochemistry; Springer International Publishing: Cham, Switzerland, 2019; pp. 275–305. ISBN 9783030178918. [Google Scholar]
- Siviter, H.; Muth, F. Do novel insecticides pose a threat to beneficial insects? Proc. R. Soc. B Biol. Sci. 2020, 287, 20201265. [Google Scholar] [CrossRef]
- Dewar, A.M. The adverse impact of the neonicotinoid seed treatment ban on crop protection in oilseed rape in the United Kingdom. Pest Manag. Sci. 2017, 73, 1305–1309. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mc Namara, L.; Gauthier, K.; Walsh, L.; Thébaud, G.; Gaffney, M.; Jacquot, E. Management of yellow dwarf disease in Europe in a post-neonicotinoid agriculture. Pest Manag. Sci. 2020, 76, 2276–2285. [Google Scholar] [CrossRef] [PubMed]
- Jactel, H.; Verheggen, F.; Thiéry, D.; Escobar-Gutiérrez, A.J.; Gachet, E.; Desneux, N. Alternatives to neonicotinoids. Environ. Int. 2019, 129, 423–429. [Google Scholar] [CrossRef] [PubMed]
- Alyokhin, A.; Baker, M.; Mota-Sanchez, D.; Dively, G.; Grafius, E. Colorado potato beetle resistance to insecticides. Am. J. Potato Res. 2008, 85, 395–413. [Google Scholar] [CrossRef]
- Zhang, H.; Yin, W.; Zhao, J.; Jin, L.; Yang, Y.; Wu, S.; Tabashnik, B.E.; Wu, Y. Early Warning of Cotton Bollworm Resistance Associated with Intensive Planting of Bt Cotton in China. PLoS ONE 2011, 6, e22874. [Google Scholar] [CrossRef] [PubMed]
- Tabashnik, B.E.; Brévault, T.; Carrière, Y. Insect resistance to Bt crops: Lessons from the first billion acres. Nat. Biotechnol. 2013, 31, 510–521. [Google Scholar] [CrossRef]
- Alvi, A.H.K.; Sayyed, A.H.; Naeem, M.; Ali, M. Field Evolved Resistance in Helicoverpa armigera (Lepidoptera: Noctuidae) to Bacillus thuringiensis Toxin Cry1Ac in Pakistan. PLoS ONE 2012, 7, e47309. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ahmad, S.; Cheema, H.M.N.; Khan, A.A.; Khan, R.S.A.; Ahmad, J.N. Resistance status of Helicoverpa armigera against Bt cotton in Pakistan. Transgenic Res. 2019, 28, 199–212. [Google Scholar] [CrossRef] [PubMed]
- Souza, D.; Vieira, B.C.; Fritz, B.K.; Hoffmann, W.C.; Peterson, J.A.; Kruger, G.R.; Meinke, L.J. Western corn rootworm pyrethroid resistance confirmed by aerial application simulations of commercial insecticides. Sci. Rep. 2019, 9, 1–10. [Google Scholar] [CrossRef] [Green Version]
- Santos, D.; Vanden Broeck, J.; Wynant, N. Systemic RNA interference in locusts: Reverse genetics and possibilities for locust pest control. Curr. Opin. Insect Sci. 2014, 6, 9–14. [Google Scholar] [CrossRef] [Green Version]
- Spit, J.; Philips, A.; Wynant, N.; Santos, D.; Plaetinck, G.; Vanden Broeck, J. Knockdown of nuclease activity in the gut enhances RNAi efficiency in the Colorado potato beetle, Leptinotarsa decemlineata, but not in the desert locust, Schistocerca gregaria. Insect Biochem. Mol. Biol. 2017, 81, 103–116. [Google Scholar] [CrossRef] [PubMed]
- Ng, K.K.; Lovell, J.F.; Zheng, G. Lipoprotein-Inspired Nanoparticles for Cancer Theranostics. Acc. Chem. Res. 2011, 44, 1105–1113. [Google Scholar] [CrossRef] [PubMed]
- Lacko, A.G.; Sabnis, N.A.; Nagarajan, B.; McConathy, W.J. HDL as a drug and nucleic acid delivery vehicle. Front. Pharmacol. 2015, 6, 1–6. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Thaxton, C.S.; Rink, J.S.; Naha, P.C.; Cormode, D.P. Lipoproteins and lipoprotein mimetics for imaging and drug delivery. Adv. Drug Deliv. Rev. 2016, 106, 116–131. [Google Scholar] [CrossRef] [Green Version]
- Mahmoudian, M.; Salatin, S.; Khosroushahi, A.Y. Natural low- and high-density lipoproteins as mighty bio-nanocarriers for anticancer drug delivery. Cancer Chemother. Pharmacol. 2018, 82, 371–382. [Google Scholar] [CrossRef]
- Raut, S.; Dasseux, J.-L.; Sabnis, N.A.; Mooberry, L.; Lacko, A. Lipoproteins for therapeutic delivery: Recent advances and future opportunities. Ther. Deliv. 2018, 9, 257–268. [Google Scholar] [CrossRef] [PubMed]
- Shotwell, C.R.; Cleary, J.D.; Berglund, J.A. The potential of engineered eukaryotic RNA-binding proteins as molecular tools and therapeutics. WIREs RNA 2020, 11, 1–21. [Google Scholar] [CrossRef]
- Li, J.; Wu, C.; Wang, W.; He, Y.; Elkayam, E.; Joshua-Tor, L.; Hammond, P.T. Structurally modulated codelivery of siRNA and Argonaute 2 for enhanced RNA interference. Proc. Natl. Acad. Sci. USA 2018, 115, E2696–E2705. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jiang, L.; Vader, P.; Schiffelers, R.M. Extracellular vesicles for nucleic acid delivery: Progress and prospects for safe RNA-based gene therapy. Gene Ther. 2017, 24, 157–166. [Google Scholar] [CrossRef]
- Massaro, C.; Sgueglia, G.; Frattolillo, V.; Baglio, S.R.; Altucci, L.; Dell’Aversana, C. Extracellular Vesicle-Based Nucleic Acid Delivery: Current Advances and Future Perspectives in Cancer Therapeutic Strategies. Pharmaceutics 2020, 12, 980. [Google Scholar] [CrossRef] [PubMed]
- Zickler, A.M.; EL Andaloussi, S. Functional extracellular vesicles aplenty. Nat. Biomed. Eng. 2020, 4, 9–11. [Google Scholar] [CrossRef]
- Witwer, K.W.; Wolfram, J. Extracellular vesicles versus synthetic nanoparticles for drug delivery. Nat. Rev. Mater. 2021, 1–4. [Google Scholar]
- Whyard, S.; Singh, A.D.; Wong, S. Ingested double-stranded RNAs can act as species-specific insecticides. Insect Biochem. Mol. Biol. 2009, 39, 824–832. [Google Scholar] [CrossRef]
- Khvorova, A.; Reynolds, A.; Jayasena, S.D. Functional siRNAs and miRNAs Exhibit Strand Bias. Cell 2003, 115, 209–216. [Google Scholar] [CrossRef] [Green Version]
- Schwarz, D.S.; Hutvágner, G.; Du, T.; Xu, Z.; Aronin, N.; Zamore, P.D. Asymmetry in the Assembly of the RNAi Enzyme Complex. Cell 2003, 115, 199–208. [Google Scholar] [CrossRef] [Green Version]
- Frank, F.; Sonenberg, N.; Nagar, B. Structural basis for 5′-nucleotide base-specific recognition of guide RNA by human AGO2. Nature 2010, 465, 818–822. [Google Scholar] [CrossRef] [PubMed]
- Ghildiyal, M.; Xu, J.; Seitz, H.; Weng, Z.; Zamore, P.D. Sorting of Drosophila small silencing RNAs partitions microRNA* strands into the RNA interference pathway. RNA 2010, 16, 43–56. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Okamura, K.; Ladewig, E.; Zhou, L.; Lai, E.C. Functional small RNAs are generated from select miRNA hairpin loops in flies and mammals. Genes Dev. 2013, 27, 778–792. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chak, L.-L.; Okamura, K. Argonaute-dependent small RNAs derived from single-stranded, non-structured precursors. Front. Genet. 2014, 5, 1–15. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nakanishi, K. Anatomy of RISC: How do small RNAs and chaperones activate Argonaute proteins? Wiley Interdiscip. Rev. RNA 2016, 7, 637–660. [Google Scholar] [CrossRef] [PubMed]
- Goh, E.; Okamura, K. Gateway to Understanding Argonaute Loading of Single-Stranded RNAs: Preparation of Deep Sequencing Libraries with In Vitro Loading Samples. In Methods in Molecular Biology; Humana Press: New York, NY, USA, 2018; pp. 41–63. [Google Scholar]
- Goh, E.; Okamura, K. Hidden sequence specificity in loading of single-stranded RNAs onto Drosophila Argonautes. Nucleic Acids Res. 2019, 47, 3101–3116. [Google Scholar] [CrossRef] [Green Version]
- Santos, D.; Mingels, L.; Vogel, E.; Wang, L.; Christiaens, O.; Cappelle, K.; Wynant, N.; Gansemans, Y.; Van Nieuwerburgh, F.; Smagghe, G.; et al. Generation of Virus- and dsRNA-Derived siRNAs with Species-Dependent Length in Insects. Viruses 2019, 11, 738. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pélisson, A.; Sarot, E.; Payen-Groschéne, G.; Bucheton, A. A Novel Repeat-Associated Small Interfering RNA-Mediated Silencing Pathway Downregulates Complementary Sense gypsy Transcripts in Somatic Cells of the Drosophila Ovary. J. Virol. 2007, 81, 1951–1960. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fu, Y.; Yang, Y.; Zhang, H.; Farley, G.; Wang, J.; Quarles, K.A.; Weng, Z.; Zamore, P.D. The genome of the Hi5 germ cell line from Trichoplusia ni, an agricultural pest and novel model for small RNA biology. Elife 2018, 7, 1–41. [Google Scholar] [CrossRef]
- Obbard, D.J.; Jiggins, F.M.; Halligan, D.L.; Little, T.J. Natural Selection Drives Extremely Rapid Evolution in Antiviral RNAi Genes. Curr. Biol. 2006, 16, 580–585. [Google Scholar] [CrossRef] [Green Version]
- Mukherjee, K.; Campos, H.; Kolaczkowski, B. Evolution of Animal and Plant Dicers: Early Parallel Duplications and Recurrent Adaptation of Antiviral RNA Binding in Plants. Mol. Biol. Evol. 2013, 30, 627–641. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wynant, N.; Santos, D.; Vanden Broeck, J. The evolution of animal Argonautes: Evidence for the absence of antiviral AGO Argonautes in vertebrates. Sci. Rep. 2017, 7, 9230. [Google Scholar] [CrossRef] [Green Version]
- Kumar, A.; Wang, S.; Ou, R.; Samrakandi, M.; Beerntsen, B.T.T.; Sayre, R.T.T. Development of an RNAi based microalgal larvicide to control mosquitoes. Malar. World J. 2013, 4, 1–7. [Google Scholar]
- Whitten, M.M.A.; Facey, P.D.; Del Sol, R.; Fernández-Martínez, L.T.; Evans, M.C.; Mitchell, J.J.; Bodger, O.G.; Dyson, P.J. Symbiont-mediated RNA interference in insects. Proc. R. Soc. B Biol. Sci. 2016, 283, 20160042. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Murphy, K.A.; Tabuloc, C.A.; Cervantes, K.R.; Chiu, J.C. Ingestion of genetically modified yeast symbiont reduces fitness of an insect pest via RNA interference. Sci. Rep. 2016, 6, 22587. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kolliopoulou, A.; Taning, C.N.T.; Smagghe, G.; Swevers, L. Viral Delivery of dsRNA for Control of Insect Agricultural Pests and Vectors of Human Disease: Prospects and Challenges. Front. Physiol. 2017, 8, 399. [Google Scholar] [CrossRef] [Green Version]
Figure 1.
Summary of the known mechanisms involved in the presence of extracellular RNAs and their functional transfer in insects. (A) D. melanogaster—miRNAs were identified in immunoprecipitates of extracellular Ago proteins and in EVs from the culture medium of D. melanogaster cells, namely the Cl8 and the S2 cell lines [65]. In addition, miRNAs and other sRNA populations were identified in EVs from the culture medium of the D. melanogaster S2R+ and D17-c3 cell lines [63]. (B) D. melanogaster—EVs from D. melanogaster hemocytes contain secondary viral siRNAs, synthesized from viral DNA. These EVs circulate in the hemolymph and functionally spread these viral siRNAs, thereby inducing systemic antiviral immunity [64]. (C), T. castaneum—dsRNA-derived siRNAs are found in EVs from the culture medium of T. castaneum TcA cells. These siRNA-containing EVs trigger RNAi in recipient cells. miRNAs and other sRNAs were also identified in these EVs [66]. (D) L. decemlineata—dsRNA was identified in EVs from the culture medium of L. decemlineata Lepd-SL1 cells, previously treated with dsRNA [68]. (E) S. gregaria—upon microinjection in the hemocoel, dsRNA binds to lipophorins in the hemolymph [169,192]. (F) A. mellifera—Major Royal Jelly Protein 3 (MRJP-3) binds to dsRNA in the jelly, protecting it from degradation and enhancing its uptake. MRJP-3 also binds single-stranded RNA and several populations of sRNAs in the jellies [71,72]. In parallel, ingested dsRNA was shown to spread in the hemolymph and to be secreted in worker and royal jellies, via which it passes to larvae, triggering target silencing [71]. (G) C. vestalis/P. xylostella—Larva of the parasitic wasp C. vestalis secretes teratocyte cells into its host, P. xylostella. These teratocytes secrete miRNA-containing EVs that enter host’ cells, where the miRNAs induce a delay in host development [74].
Figure 1.
Summary of the known mechanisms involved in the presence of extracellular RNAs and their functional transfer in insects. (A) D. melanogaster—miRNAs were identified in immunoprecipitates of extracellular Ago proteins and in EVs from the culture medium of D. melanogaster cells, namely the Cl8 and the S2 cell lines [65]. In addition, miRNAs and other sRNA populations were identified in EVs from the culture medium of the D. melanogaster S2R+ and D17-c3 cell lines [63]. (B) D. melanogaster—EVs from D. melanogaster hemocytes contain secondary viral siRNAs, synthesized from viral DNA. These EVs circulate in the hemolymph and functionally spread these viral siRNAs, thereby inducing systemic antiviral immunity [64]. (C), T. castaneum—dsRNA-derived siRNAs are found in EVs from the culture medium of T. castaneum TcA cells. These siRNA-containing EVs trigger RNAi in recipient cells. miRNAs and other sRNAs were also identified in these EVs [66]. (D) L. decemlineata—dsRNA was identified in EVs from the culture medium of L. decemlineata Lepd-SL1 cells, previously treated with dsRNA [68]. (E) S. gregaria—upon microinjection in the hemocoel, dsRNA binds to lipophorins in the hemolymph [169,192]. (F) A. mellifera—Major Royal Jelly Protein 3 (MRJP-3) binds to dsRNA in the jelly, protecting it from degradation and enhancing its uptake. MRJP-3 also binds single-stranded RNA and several populations of sRNAs in the jellies [71,72]. In parallel, ingested dsRNA was shown to spread in the hemolymph and to be secreted in worker and royal jellies, via which it passes to larvae, triggering target silencing [71]. (G) C. vestalis/P. xylostella—Larva of the parasitic wasp C. vestalis secretes teratocyte cells into its host, P. xylostella. These teratocytes secrete miRNA-containing EVs that enter host’ cells, where the miRNAs induce a delay in host development [74].


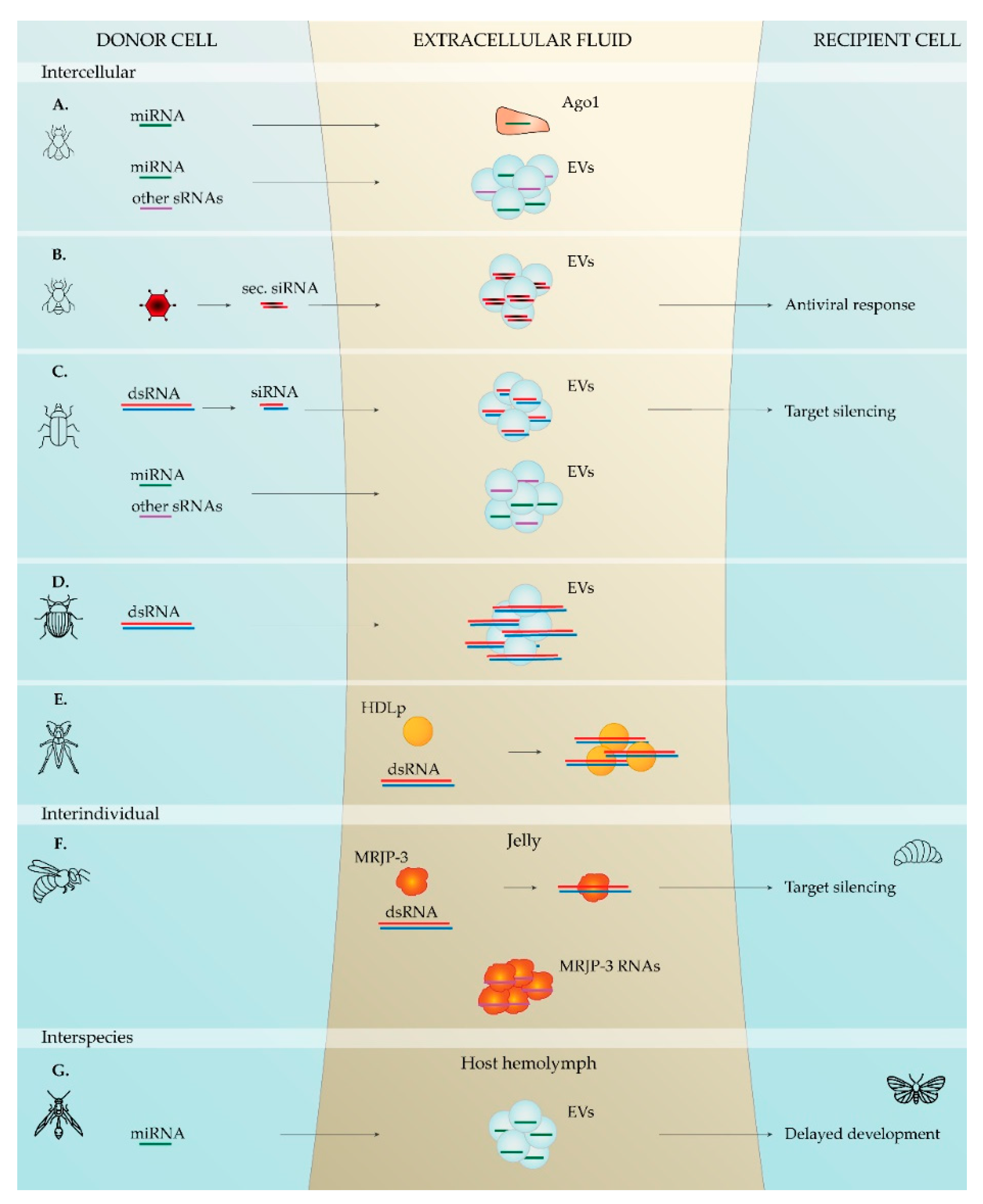{kind=link}
Table 1.
Summary of the reported studies demonstrating extracellular RNAs and their functional transfer in insects. Plasma refers to cell-free hemolymph. * Indicates that functionality was reported.
Table 1.
Summary of the reported studies demonstrating extracellular RNAs and their functional transfer in insects. Plasma refers to cell-free hemolymph. * Indicates that functionality was reported.
| Insect | Sample | RNA | Reference | |
|---|---|---|---|---|
| Intercellular / Intraindividual | Korean rhinoceros beetle, Allomyrina dichotoma | plasma | sRNAs | [67] |
| Fruit fly, D. melanogaster | plasma | miRNAs | [62] | |
| Fruit fly, D. melanogaster | medium of cultured cells (S2R+ and D17-c3) | sRNAs | [63] | |
| Fruit fly, D. melanogaster | medium of cultured cells (S2 and Cl8 cell lines) | miRNAs | [65] | |
| Fruit fly, D. melanogaster | plasma | siRNAs * | [64] | |
| Red flour beetle, T. castaneum | medium of cultured cells (TcA cell line) | siRNAs *, miRNAs and other sRNAs | [66] | |
| Colorado potato beetle, L. decemlineata | medium of cultured cells (Lepd-SL1 cell line) | dsRNA | [68] | |
| Interindividual | Honey bee, A. mellifera | nurse bee secretions | miRNAs * | [70] |
| Honey bee, A. mellifera | hemolymph, worker and royal jellies | dsRNA *, ssRNA and sRNAs | [71,72] | |
| Florida carpenter ant, C. floridanus | trophallactic fluid | miRNAs | [73] | |
| Interspecies | Mosquito, Aedes aegypti and Aedes albopictus | saliva | miRNAs * | [60] |
| Mosquito, Anopheles coluzzii | saliva | miRNAs and other sRNAs | [61] | |
| Parasitic wasp, C. vestalis | host body (Diamondback moth, P. xylostella) | miRNAs * | [74] |
Table 2.
Summary of the reported studies indicating natural RNA transfer from plant to insects. * Indicates that functionality was reported.
Table 2.
Summary of the reported studies indicating natural RNA transfer from plant to insects. * Indicates that functionality was reported.
| Plant | Insect | Sample | RNA | Reference |
|---|---|---|---|---|
| C. melo | Cotton-melon aphid, A. gossypii | whole insect | miRNAs | [107] |
| Morus notabilis | Silkworm, B. mori | hemolymph, fat body, and silk gland | miRNAs | [108] |
| Sorghum bicolor | Greenbug, S. graminum | whole insect | miRNAs | [109] |
| Hordeum vulgare | Yellow sugarcane aphid, S. flava | whole insect | miRNAs | [109] |
| B. oleracea | Green peach aphid, M. persicae | gut | miRNAs | [110] |
| Arabidopsis thaliana | Diamondback moth, P. xylostella | hemolymph | miRNAs * | [111] |
| Brassica campestris | Honey bee, A. mellifera | beebread and royal jelly | miRNAs * | [113] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Santos, D.; Remans, S.; Van den Brande, S.; Vanden Broeck, J. RNAs on the Go: Extracellular Transfer in Insects with Promising Prospects for Pest Management. Plants 2021, 10, 484. https://0-doi-org.brum.beds.ac.uk/10.3390/plants10030484
AMA Style
Santos D, Remans S, Van den Brande S, Vanden Broeck J. RNAs on the Go: Extracellular Transfer in Insects with Promising Prospects for Pest Management. Plants. 2021; 10(3):484. https://0-doi-org.brum.beds.ac.uk/10.3390/plants10030484
Chicago/Turabian StyleSantos, Dulce, Simon Remans, Stijn Van den Brande, and Jozef Vanden Broeck. 2021. "RNAs on the Go: Extracellular Transfer in Insects with Promising Prospects for Pest Management" Plants 10, no. 3: 484. https://0-doi-org.brum.beds.ac.uk/10.3390/plants10030484
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.