
Biotic and Abiotic Elicitors of Stilbenes Production in Vitis vinifera L. Cell Culture

 , , ,
, , ,
Abstract
:1. Introduction
2. Results
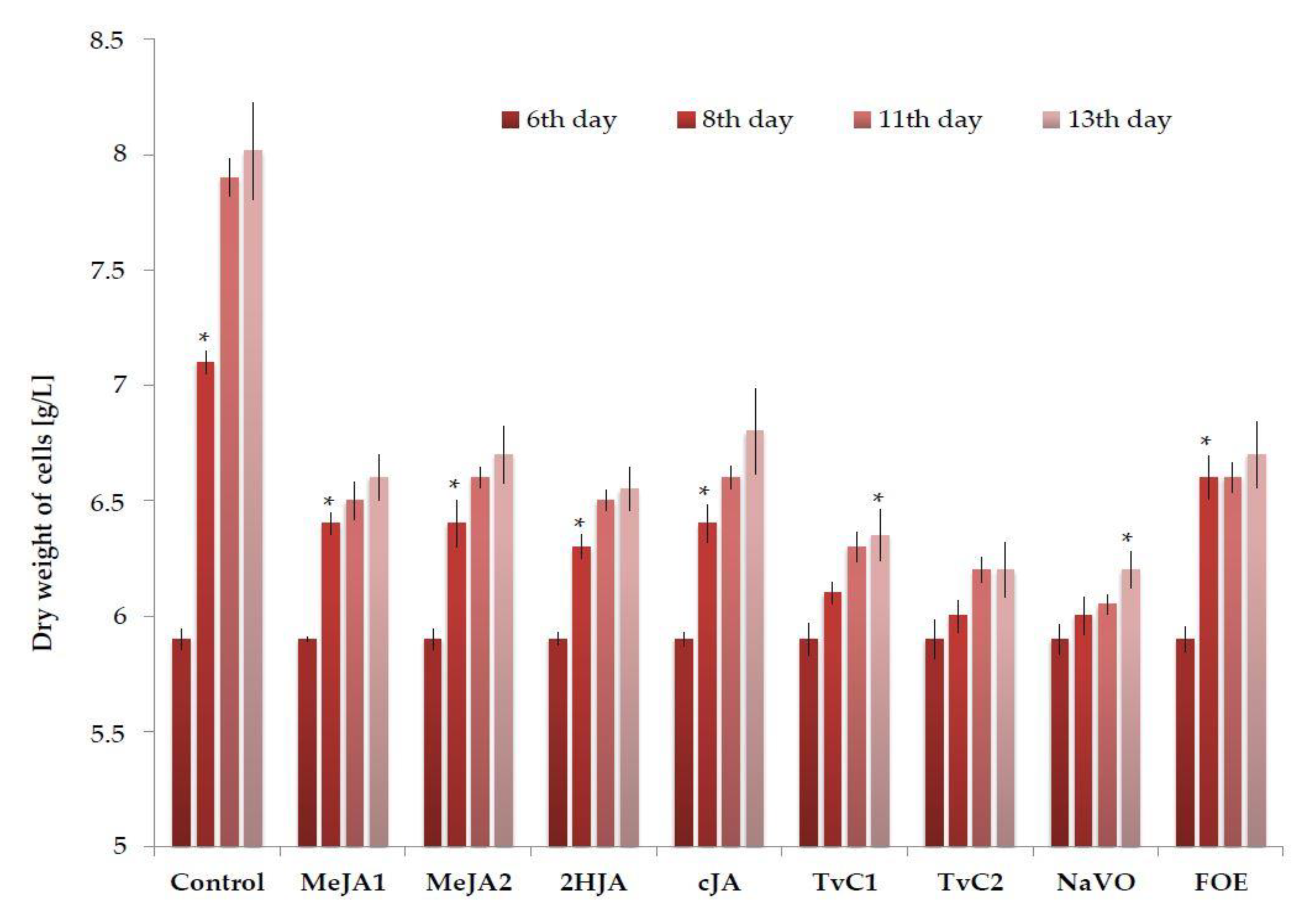
2.1. Cell Biomass Growth
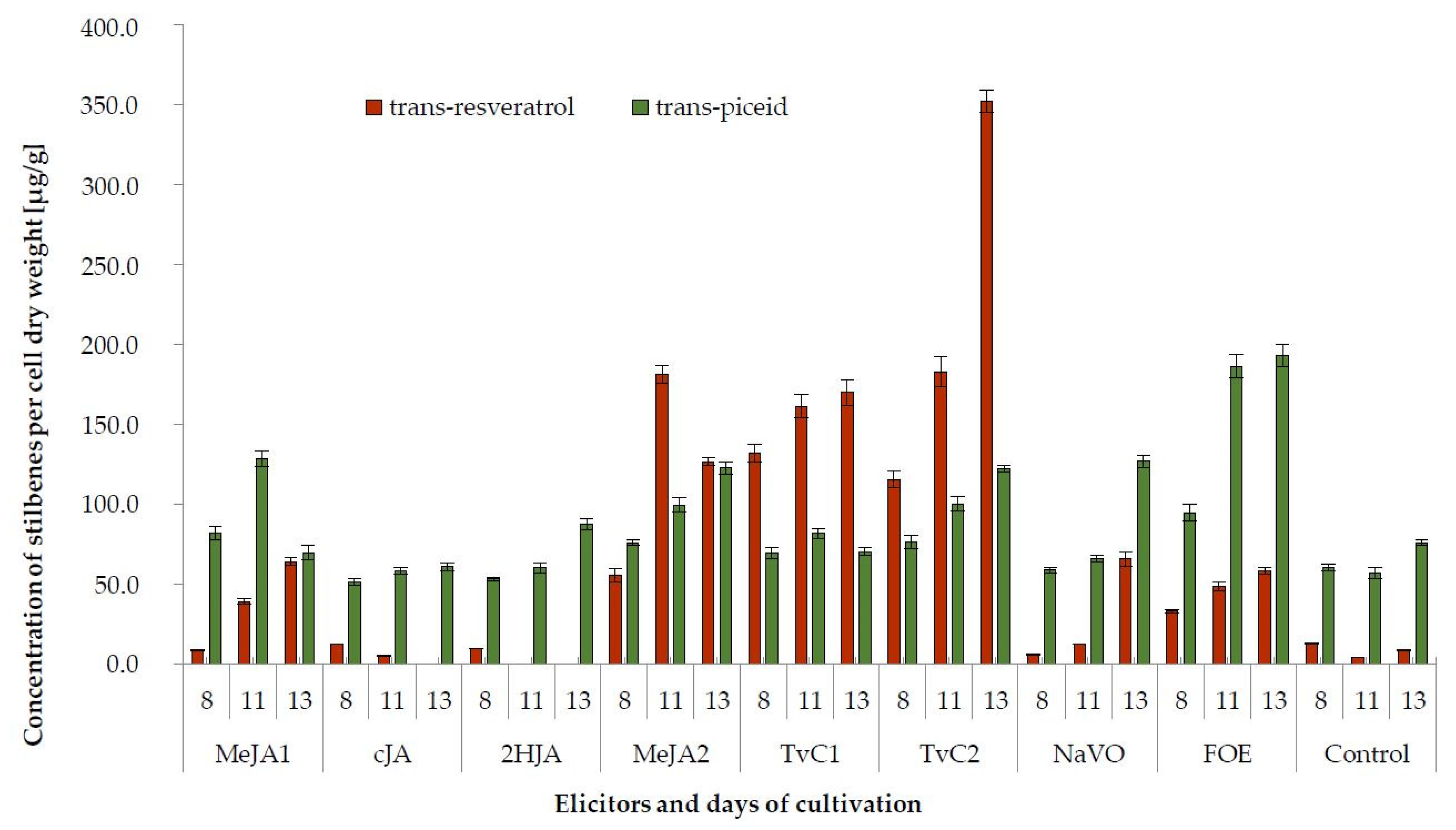
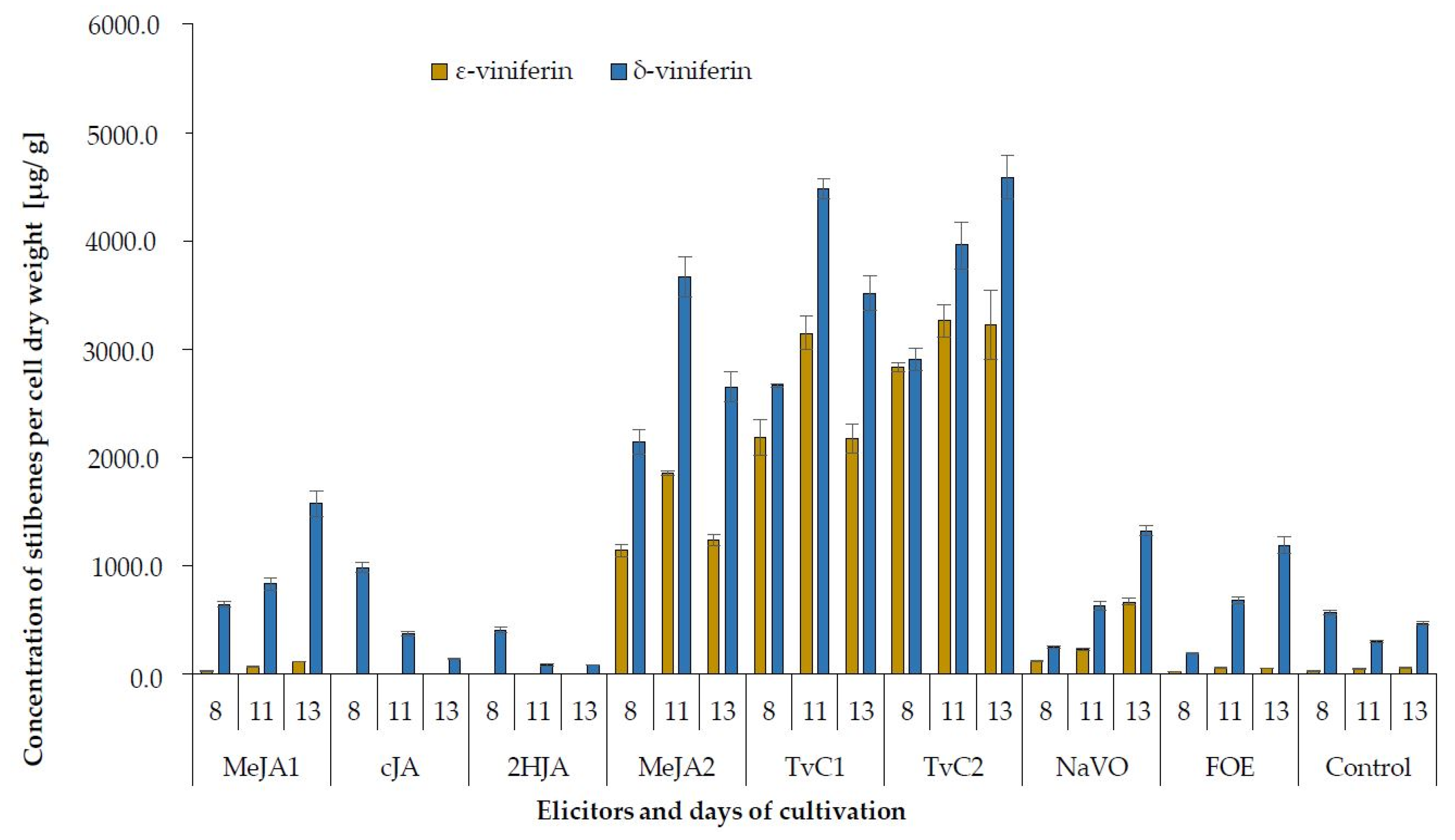
2.2. Production of Stilbenes
3. Discussion
4. Materials and Methods
4.1. Reagents and Standards
4.2. Cell Suspension Cultures
4.3. Elicitor Preparation and Elicitor Treatment
4.4. Extraction of Stilbenes
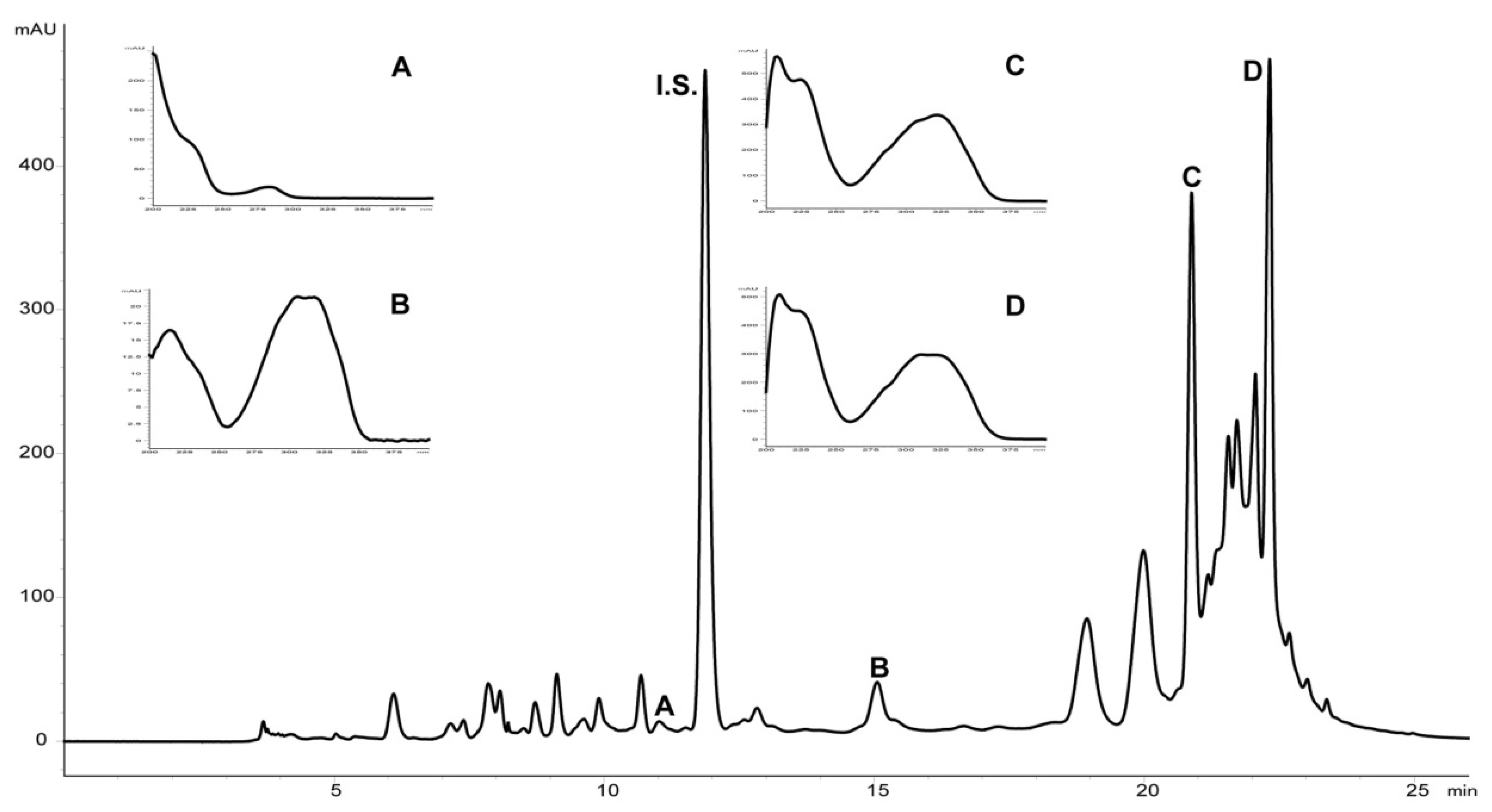
4.5. Chromatographic Analysis
4.6. Statistical Evaluation
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Hammerschmidt, R. PHYTOALEXINS: What Have We Learned After 60 Years? Annu. Rev. Phytopathol. 1999, 37, 285–306. [Google Scholar] [CrossRef] [Green Version]
- Langcake, P.; Pryce, R.J. A new class of phytoalexins from grapevines. Cell. Mol. Life Sci. 1977, 33, 151–152. [Google Scholar] [CrossRef] [PubMed]
- Jeandet, P.; Douillet-Breuil, A.-C.; Bessis, R.; Debord, S.; Sbaghi, M.; Adrian, M. Phytoalexins from the Vitaceae: Biosynthesis, Phytoalexin Gene Expression in Transgenic Plants, Antifungal Activity, and Metabolism. J. Agric. Food Chem. 2002, 50, 2731–2741. [Google Scholar] [CrossRef] [PubMed]
- Ahuja, I.; Kissen, R.; Bones, A.M. Phytoalexins in defense against pathogens. Trends Plant Sci. 2012, 17, 73–90. [Google Scholar] [CrossRef]
- Pastor, R.F.; Restani, P.; Di Lorenzo, C.; Orgiu, F.; Teissedre, P.-L.; Stockley, C.; Ruf, J.C.; Quini, C.I.; Tejedor, N.G.; Gargantini, R.; et al. Resveratrol, human health and winemaking perspectives. Crit. Rev. Food Sci. Nutr. 2019, 59, 1237–1255. [Google Scholar] [CrossRef]
- Baur, J.A.; Sinclair, D.A. Therapeutic potential of resveratrol: The in vivo evidence. Nat. Rev. Drug Discov. 2006, 5, 493–506. [Google Scholar] [CrossRef]
- Guerrero, R.F.; García-Parrilla, M.C.; Puertas, B.; Cantos-Villar, E. Wine, Resveratrol and Health: A Review. Nat. Prod. Commun. 2009, 4, 635–658. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Richard, T.; Pawlus, A.D.; Iglésias, M.-L.; Pedrot, E.; Waffo-Teguo, P.; Mérillon, J.-M.; Monti, J.-P. Neuroprotective properties of resveratrol and derivatives. Ann. N. Y. Acad. Sci. 2011, 1215, 103–108. [Google Scholar] [CrossRef] [PubMed]
- Giovannelli, L.; Innocenti, M.; Santamaria, A.R.; Bigagli, E.; Pasqua, G.; Mulinacci, N. Antitumoral activity of viniferin-enriched extracts from Vitis vinifera L. cell cultures. Nat. Prod. Res. 2014, 28, 2006–2016. [Google Scholar] [CrossRef] [PubMed]
- Gambini, J.; Inglés, M.; Olaso, G.; Lopez-Grueso, R.; Bonet-Costa, V.; Gimeno-Mallench, L.; Mas-Bargues, C.; Abdelaziz, K.M.; Gomez-Cabrera, M.C.; Vina, J.; et al. Properties of resveratrol: In vitro and in vivo studies about metabolism, bioavailability, and biological effects in animal models and humans. Oxid. Med. Cell. Longev. 2015, 215, 1–13. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Biais, B.; Krisa, S.; Cluzet, S.; Da Costa, G.; Waffo-Teguo, P.; Mérillon, J.-M.; Richard, T. Antioxidant and Cytoprotective Activities of Grapevine Stilbenes. J. Agric. Food Chem. 2017, 65, 4952–4960. [Google Scholar] [CrossRef]
- Fan, E.; Zhang, K.; Zhu, M.; Wang, Q. Obtaining Resveratrol: From Chemical Synthesis to Biotechnological Production. Mini-Rev. Org. Chem. 2010, 7, 272–281. [Google Scholar] [CrossRef]
- Donnez, D.; Jeandet, P.; Clément, C.; Courot, E. Bioproduction of resveratrol and stilbene derivatives by plant cells and microorganisms. Trends Biotechnol. 2009, 27, 706–713. [Google Scholar] [CrossRef] [PubMed]
- Vuong, T.V.; Franco, C.M.M.; Zhang, W. Treatment strategies for high resveratrol induction in Vitis vinifera L. cell suspension culture. Biotechnol. Rep. 2014, 1–2, 15–21. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sae-Lee, N.; Kerdchoechuen, O.; Laohakunjit, N. Enhancement of Phenolics, Resveratrol and Antioxidant Activity by Nitrogen Enrichment in Cell Suspension Culture of Vitis vinifera. Molecules 2014, 19, 7901–7912. [Google Scholar] [CrossRef] [Green Version]
- Almagro, L.; Belchí-Navarro, S.; Martínez-Márquez, A.; Bru, R.; Pedreño, M.A. Enhanced extracellular production of trans-resveratrol in Vitis vinifera suspension cultured cells by using cyclodextrins and coronatine. Plant Physiol. Biochem. 2015, 97, 361–367. [Google Scholar] [CrossRef] [Green Version]
- Ismail, A.; Seo, M.; Takebayashi, Y.; Kamiya, Y.; Nick, P. A balanced JA/ABA status may correlate with adaptation to osmot-ic stress in Vitis cells. J. Plant Physiol. 2015, 185, 57–64. [Google Scholar] [CrossRef] [PubMed]
- Decendit, A.; Ramawat, K.G.; Waffo, P.; Badoc, A. Anthocyanins, catechins, condensed tannins and piceid production in Vitis vinifera cell bioreactor cultures. Biotechnol. Lett. 1996, 18, 659–662. [Google Scholar] [CrossRef]
- Ferri, M.; Dipalo, S.C.; Bagni, N.; Tassoni, A. Chitosan elicits mono-glucosylated stilbene production and release in fed-batch bioreactor cultures of grape cells. Food Chem. 2011, 124, 1473–1479. [Google Scholar] [CrossRef]
- Donnez, D.; Kim, K.-H.; Antoine, S.; Conreux, A.; De Luca, V.; Jeandet, P.; Clément, C.; Courot, E. Bioproduction of resveratrol and viniferins by an elicited grapevine cell culture in a 2 L stirred bioreactor. Process. Biochem. 2011, 46, 1056–1062. [Google Scholar] [CrossRef]
- Jeandet, P.; Clément, C.; Courot, E. Resveratrol production at large scale using plant cell suspensions. Eng. Life Sci. 2014, 14, 622–632. [Google Scholar] [CrossRef]
- Flamini, R.; Zanzotto, A.; de Rosso, M.; Lucchetta, G.; Vedova, A.D.; Bavaresco, L. Stilbene oligomer phytoalexins in grape as a response to Aspergillus carbonarius infection. Physiol. Mol. Plant Pathol. 2016, 93, 112–118. [Google Scholar] [CrossRef]
- Kiselev, K.; Dubrovina, A.; Veselova, M.; Bulgakov, V.; Fedoreyev, S.; Zhuravlev, Y. The rolB gene-induced overproduction of resveratrol in Vitis amurensis transformed cells. J. Biotechnol. 2007, 128, 681–692. [Google Scholar] [CrossRef]
- Ramirez-Estrada, K.; Vidal-Limon, H.; Hidalgo, D.; Moyano, E.; Golenioswki, M.; Cusidó, R.M.; Palazon, J. Elicitation, an Effective Strategy for the Biotechnological Production of Bioactive High-Added Value Compounds in Plant Cell Factories. Molecules 2016, 21, 182. [Google Scholar] [CrossRef]
- Lijavetzky, D.; Almagro, L.; Belchi-Navarro, S.; Martínez-Zapater, J.M.; Bru, R.; Pedreño, M. Synergistic effect of methyljasmonate and cyclodextrin on stilbene biosynthesis pathway gene expression and resveratrol production in Monastrell grapevine cell cultures. BMC Res. Notes 2008, 1, 132. [Google Scholar] [CrossRef] [Green Version]
- Radman, R.; Saez, T.; Bucke, C.; Keshavarz, T. Elicitation of plants and microbial cell systems. Biotechnol. Appl. Biochem. 2003, 37, 91–102. [Google Scholar] [CrossRef]
- Namdeo, A.G. Plant cell elicitation for production of secondary metabolites: A review. Pharmacogn. Rev. 2007, 1, 69–79. [Google Scholar]
- Vasconsuelo, A.; Boland, R. Molecular aspects of the early stages of elicitation of secondary metabolites in plants. Plant Sci. 2007, 172, 861–875. [Google Scholar] [CrossRef]
- Kombrink, E. Chemical and genetic exploration of jasmonate biosynthesis and signaling paths. Planta 2012, 236, 1351–1366. [Google Scholar] [CrossRef]
- Zhang, W.; Curtin, C.; Kikuchi, M.; Franco, C. Integration of jasmonic acidand light irradiation for enhancement of antho-cyanin biosynthesis in Vitis vinifera suspension cultures. Plant Sci. 2002, 162, 459–468. [Google Scholar] [CrossRef]
- Belchí-Navarro, S.; Almagro, L.; Sabater-Jara, A.B.; Fernández-Pérez, F.; Bru, R.; Pedreño, M.A. Induction of trans-resveratrol and extracellular pathogenesis-related proteins in elicited suspension cultured cells of Vitis vinifera cv Monastrell. J. Plant Physiol. 2013, 170, 258–264. [Google Scholar] [CrossRef]
- Creelman, R.A.; Mullet, J.E. Biosynthesis and action of jasmonates in plants. Annu. Rev. Plant Biol. 1997, 48, 355–381. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Martinez-Esteso, M.J.; Sellés-Marchart, S.; Vera-Urbina, J.C.; Pedreño, M.A.; Bru-Martinez, R. DIGE analysis of proteome changes accompanying large resveratrol production by grapevine (Vitis vinifera cv Gamay) cell cultures in response to me-thyl-β-cyclodextrin and methyl jasmonate elicitors. J. Proteom. 2011, 74, 1421–1436. [Google Scholar] [CrossRef] [PubMed]
- Nopo-Olazabal, C.; Condori, J.; Nopo-Olazabal, L.; Medina-Bolivar, F. Differential induction of antioxidant stilbenoids in hairy roots of Vitis rotundifolia treated with methyl jasmonate and hydrogen peroxide. Plant Physiol. Biochem. 2014, 74, 50–69. [Google Scholar] [CrossRef]
- Santamaria, A.R.; Mulinacci, N.; Valletta, A.; Innocenti, M.; Pasqua, G. Effects of Elicitors on the Production of Resveratrol and Viniferins in Cell Cultures ofVitis viniferaL. cv Italia. J. Agric. Food Chem. 2011, 59, 9094–9101. [Google Scholar] [CrossRef] [PubMed]
- Dar, T.A.; Uddin, M.; Khan, M.M.A.; Hakeem, K.; Jaleel, H. Jasmonates counter plant stress: A Review. Environ. Exp. Bot. 2015, 115, 49–57. [Google Scholar] [CrossRef]
- Gopalan, S.; Bauer, D.W.; Alfano, J.R.; Loniello, A.O.; He, S.Y.; Collmer, A. Expression of the Pseudomonas syringae avirulence protein AvrB in plant cells alleviates its dependence on the hypersensitive response and pathogenicity (Hrp) secretion sys-tem in eliciting genotype-specific hypersensitive cell death. Plant Cell 1996, 8, 1095–1105. [Google Scholar]
- Tassoni, A.; Fornalè, S.; Franceschetti, M.; Musiani, F.; Michael, A.J.; Perry, B.; Bagni, N. Jasmonates and Na-orthovanadate promote resveratrol production in Vitis vinifera cv. Barbera cell cultures. New Phytol. 2005, 166, 895–905. [Google Scholar] [CrossRef]
- Calderón, A.A.; Zapata, J.M.; Muñoz, R.; Pedreño, M.A.; Barceló, A.R. Resveratrol production as a part of the hypersensitive-like response of grapevine cells to an elicitor from Trichoderma viride. New Phytol. 1993, 124, 455–463. [Google Scholar] [CrossRef]
- Calderón, A.; Zapata, J.; Barceló, A.R. Peroxidase-mediated formation of resveratrol oxidation products during the hypersensitive-like reaction of grapevine cells to an elicitor from Trichoderma viride. Physiol. Mol. Plant Pathol. 1994, 44, 289–299. [Google Scholar] [CrossRef]
- Barceló, A.R.; Zapata, J.M.; Calderón, A.A. A Basic Peroxidase Isoenzyme, Marker of Resistance Against Plasmopara viticola in Grapevines, is Induced by an Elicitor from Trichoderma viride in Susceptible Grapevines. J. Phytopathol. 1996, 144, 309–313. [Google Scholar] [CrossRef]
- Mutawila, C.; Stander, C.; Halleen, F.; Vivier, M.A.; Mostert, L. Response of Vitis vinifera cell cultures to Eutypa lata and Trichoderma atroviride culture filtrates: Expression of defence-related genes and phenotypes. Protoplasma 2016, 254, 863–879. [Google Scholar] [CrossRef]
- Namdeo, A.; Patil, S.; Fulzele, D.P. Influence of Fungal Elicitors on Production of Ajmalicine by Cell Cultures of Catharanthus roseus. Biotechnol. Prog. 2002, 18, 159–162. [Google Scholar] [CrossRef]
- Sák, M.; Dokupilová, I.; Mihálik, D.; Lakatošová, J.; Gubišová, M.; Kraic, J. Elicitation Phenolic Compounds in Cell Culture of Vitis vinifera L. by Phaeomoniella chlamydospora. Nova Biotechnol. Chim. 2014, 13, 162–171. [Google Scholar] [CrossRef]
- Highet, A.; Nair, N. Fusarium oxysporum associated with grapevine decline in the Hunter Valley, NSW, Australia. Aust. J. Grape Wine Res. 1995, 1, 48–50. [Google Scholar] [CrossRef]
- Omer, A.D.; Granett, J.; Wakeman, R.J. Pathogenicity of Fusarium oxysporum on different Vitis rootstocks. J. Phytopathol. 1999, 147, 433–436. [Google Scholar] [CrossRef]
- Reveglia, P.; Cinelli, T.; Cimmino, A.; Masi, M.; Evidente, A. The main phytotoxic metabolite produced by a strain of Fusarium oxysporim inducing grapevine plant declining in Italy. Nat. Prod. Res. 2018, 32, 2397–2407. [Google Scholar] [CrossRef]
- Zhai, X.; Jia, M.; Chen, L.; Zheng, C.-J.; Rahman, K.; Han, T.; Qin, L.-P. The regulatory mechanism of fungal elicitor-induced secondary metabolite biosynthesis in medical plants. Crit. Rev. Microbiol. 2017, 43, 238–261. [Google Scholar] [CrossRef]
- Cai, Z.; Kastell, A.; Mewis, I.; Knorr, D.; Smetanska, I. Polysaccharide elicitors enhance anthocyanin and phenolic acid accumulation in cell suspension cultures of Vitis vinifera. Plant Cell Tissue Organ Cult. 2011, 108, 401–409. [Google Scholar] [CrossRef]
- Rühmann, S.; Pfeiffer, J.; Brunner, P.; Szankowski, I.; Fischer, T.C.; Forkmann, G.; Treutter, D. Induction of stilbene phytoa-lexins in grapevine (Vitis vinifera) and transgenic stilbene synthase-apple plants (Malus domestica) by a culture filtrate of Au-reobasidium pullulans. Plant Physiol. Biochem. 2013, 72, 62–71. [Google Scholar] [CrossRef]
- Cai, Z.; Riedel, H.; Saw, T.N.; Mewis, I.; Reineke, K.; Knorr, D.; Smetanska, I. Effects of elicitors and high hydrostatic pres-sure on secondary metabolism of Vitis vinifera suspension culture. Process Biochem. 2011, 46, 1411–1416. [Google Scholar] [CrossRef]
- Sák, M.; Dokupilová, I.; Mihalik, D.; Sajbidor, J. Response of Vitis vinifera cell cultures on stress caused by methyl jasmonate. Curr. Opin. Biotechnol. 2013, 24, S127. [Google Scholar] [CrossRef]
- Steponkus, P.L.; Lanphear, F.O. Refinement of the triphenyltetrazolium chloride method of determining cold injury. Plant Physiol. 1967, 42, 1423–1426. [Google Scholar] [CrossRef] [Green Version]
- Pezet, R.; Perret, C.; Jean-Denis, J.B.; Tabacchi, R.; Gindro, A.K.; Viret, O. δ-Viniferin, a Resveratrol Dehydrodimer: One of the Major Stilbenes Synthesized by Stressed Grapevine Leaves. J. Agric. Food Chem. 2003, 51, 5488–5492. [Google Scholar] [CrossRef] [PubMed]
- Creelman, R.A.; Mullet, J. Oligosaccharins, brassinolides, and jasmonates: Nontraditional regulators of plant growth, development, and gene expression. Plant Cell 1997, 9, 1211–1223. [Google Scholar] [CrossRef] [Green Version]
- Angelova, Z.; Georgiev, S.; Roos, W. Elicitation of Plants. Biotechnol. Biotechnol. Equip. 2006, 20, 72–83. [Google Scholar] [CrossRef] [Green Version]
- Almagro, L.; Carbonell-Bejerano, P.; Belchí-Navarro, S.; Bru, R.; Martínez-Zapater, J.M.; Lijavetzky, D.; Pedreño, M.A. Dissecting the Transcriptional Response to Elicitors in Vitis vinifera Cells. PLoS ONE 2014, 9, e109777. [Google Scholar] [CrossRef] [Green Version]
- Rajesh, R.W.; Rahul, M.S.; Ambalal, N.S.; Waghund, R.R.; Shelake, R.M.; Sabalpara, A.N. Trichoderma: A significant fungus for agriculture and environment. Afr. J. Agric. Res. 2016, 11, 1952–1965. [Google Scholar] [CrossRef] [Green Version]
- Wang, H.; Liu, L.; Guo, Y.-X.; Dong, Y.-S.; Zhang, D.-J.; Xiu, Z.-L. Biotransformation of piceid in Polygonum cuspidatum to resveratrol by Aspergillus oryzae. Appl. Microbiol. Biotechnol. 2007, 75, 763–768. [Google Scholar] [CrossRef] [PubMed]
- Chong, J.; Poutaraud, A.; Hugueney, P. Metabolism and roles of stilbenes in plants. Plant Sci. 2009, 177, 143–155. [Google Scholar] [CrossRef]
- Hipskind, J.D.; Paiva, N.L. Constitutive Accumulation of a Resveratrol-Glucoside in Transgenic Alfalfa Increases Resistance to Phoma medicaginis. Mol. Plant-Microbe Interact. 2000, 13, 551–562. [Google Scholar] [CrossRef] [Green Version]
- Kováčik, J.; Grúz, J.; Klejdus, B.; Štork, F.; Marchiosi, R.; Ferrarese-Filho, O. Lignification and related parameters in cop-per-exposed Matricaria chamomilla roots: Role of H2O2 and NO in this process. Plant Sci. 2010, 179, 383–389. [Google Scholar] [CrossRef]
- Kouakou-Siransy, G.; Sahpaz, S.; Irié-Nguessan, G.; Datte, Y.; Kablan, J.; Gressier, B.; Bailleul, F. Oxygen species scavenger activities and phenolic contents of four West African plants. Food Chem. 2010, 118, 430–435. [Google Scholar] [CrossRef]
- De Maria, S.; Scognamiglio, I.; Lombardi, A.; Amodio, N.; Caraglia, M.; Cartenì, M.; Ravagnan, G.; Stiuso, P. Polydatin, a natural precursor of resveratrol, induces cell cycle arrest and differentiation of human colorectal Caco-2 cell. J. Transl. Med. 2013, 11, 264. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, H.-L.; Gao, J.-P.; Han, Y.-L.; Xu, X.; Wu, R.; Gao, Y.; Cui, X.-H. Comparative studies of polydatin and resveratrol on mutual transformation and antioxidative effect in vivo. Phytomedicine 2015, 22, 553–559. [Google Scholar] [CrossRef]
- Barceló, A.R.; Pomar, F.; López-Serrano, M.; Pedreño, M. Ángeles Peroxidase: A multifunctional enzyme in grapevines. Funct. Plant Biol. 2003, 30, 577–591. [Google Scholar] [CrossRef] [PubMed]
- Donati, L.; Ferretti, L.; Frallicciardi, J.; Rosciani, R.; Valletta, A.; Pasqua, G. Stilbene biosynthesis and gene expression in response to methyl jasmonate and continuous light treatment in Vitis vinifera cv. Malvasia del Lazio and Vitis rupestris Du Lot cell cultures. Plant Physiol. 2019, 166, 646–662. [Google Scholar] [CrossRef]
- Santamaria, A.R.; Antonacci, D.; Caruso, G.; Cavaliere, C.; Gubbiott, R.; Laganà, A.; Valletta, A.; Pasqua, G. Stilbene production in cell cultures ofVitis viniferaL. cvs Red Globe and Michele Palieri elicited by methyl jasmonate. Nat. Prod. Res. 2009, 24, 1488–1498. [Google Scholar] [CrossRef]
- Duan, D.; Halter, D.; Baltenweck, R.; Tisch, C.; Tröster, V.; Kortekamp, A.; Hugueney, P.; Nick, P. Genetic diversity of stilbene metabolism in Vitis sylvestris. J. Exp. Bot. 2015, 66, 3243–3257. [Google Scholar] [CrossRef]
- Suwalsky, M.; Fierro, P.; Villena, F.; Aguilar, L.F.; Sotomayor, C.P.; Jemiola-Rzeminska, M.; Szutowicz, A. Human erythro-cytes and neuroblastoma cells are in vitro affected by sodium orthovanadate. Biochim. Biophys Acta 2012, 1818, 2260–2270. [Google Scholar] [CrossRef] [Green Version]
- Belhadj, A.; Telef, N.; Saigne, C.; Cluzet, S.; Barrieu, F.; Hamdi, S.; Mérillon, J.-M. Effect of methyl jasmonate in combination with carbohydrates on gene expression of PR proteins, stilbene and anthocyanin accumulation in grapevine cell cultures. Plant Physiol. Biochem. 2008, 46, 493–499. [Google Scholar] [CrossRef]
- Belchí-Navarro, S.; Almagro, L.; Lijavetzky, D.; Bru, R.; Pedreño, M.A. Enhanced extracellular production of trans-resveratrol in Vitis vinifera suspension cultured cells by using cyclodextrins and methyljasmonate. Plant Cell Rep. 2011, 31, 81–89. [Google Scholar] [CrossRef] [PubMed]
- Faurie, B.; Cluzet, S.; Corio-Costet, M.F.; Mérillon, J.M. Methyl jasmonate/ethephon cotretment synergistically induces stilbene production in Vitis vinifera cell suspensions but fails to trigger resistance to Erysiphe necator. J. Int. Sci. Vigne. Vin. 2009, 43, 99–110. [Google Scholar]
- Santamaria, A.R.; Innocenti, M.; Mulinacci, N.; Melani, F.; Valletta, A.; Schiandra, I.; Pasqua, G. Enhancement of viniferin production bin Vitis vinifera L. cv. Alphonse Lavallé cell suspensions by low-energy ultrasound alone and in combination with methyl jasmonate. J. Agric. Food Chem. 2012, 60, 11135–11142. [Google Scholar] [CrossRef] [PubMed]
- Douillet-Breuil, A.-C.; Jeandet, P.; Adrian, M.; Bessis, R. Changes in the Phytoalexin Content of VariousVitisSpp. in Response to Ultraviolet C Elicitation. J. Agric. Food Chem. 1999, 47, 4456–4461. [Google Scholar] [CrossRef]
- Coutos-Thévenot, P.; Poinssot, B.; Bonomelli, A.; Yean, H.; Breda, C.; Buffard, D.; Esnault, R.; Hain, R.; Boulay, M. In vitro tolerance to Botrytis cinerea of grapevine 41B rootstock in transgenic plants expressing the stilbene synthase Vst1 gene under the control of a pathogen-inducible PR 10 promoter. J. Exp. Bot. 2001, 52, 901–910. [Google Scholar] [CrossRef] [Green Version]
- Chu, M.; Pedreño, M.; Alburquerque, N.; Faize, L.; Burgos, L.; Almagro, L. A new strategy to enhance the biosynthesis of trans -resveratrol by overexpressing stilbene synthase gene in elicited Vitis vinifera cell cultures. Plant Physiol. Biochem. 2017, 113, 141–148. [Google Scholar] [CrossRef]
- Jeandet, P.; Courot, E.; Clément, C.; Ricord, S.; Crouzet, J.; Aziz, A.; Cordelier, S. Molecular Engineering of Phytoalexins in Plants: Benefits and Limitations for Food and Agriculture. J. Agric. Food Chem. 2017, 65, 2643–2644. [Google Scholar] [CrossRef] [PubMed]
- Martínez-Márquez, A.; Morante-Carriel, J.A.; Ramírez-Estrada, K.; Cusidó, R.M.; Palazon, J.; Bru-Martínez, R. Production of highly bioactive resveratrol analogues pterostilbene and piceatannol in metabolically engineered grapevine cell cultures. Plant Biotechnol. J. 2016, 14, 1813–1825. [Google Scholar] [CrossRef] [Green Version]
- Murashige, T.; Skoog, F. A revised medium for rapid growth and bioassays with tobacco tissue cultures. Physiol. Plant 1962, 15, 473–497. [Google Scholar] [CrossRef]
- Wenjun, W.; Longjiang, Y.; Pengpeng, Z. Effects of different fungal elicitors on growth, total carotenoids and astaxanthin formation by Xanthophyllomyces dendrorhous. Bioresour. Technol. 2006, 97, 26–31. [Google Scholar]
- Li-Wen, Z.; Jian-Jiang, Z.; Ya-Jie, T. Significance of fungal elicitors on the production of ganoderic acid and Ganoderma polysaccharides by the submerged culture of medicinal mushroom Ganoderma lucidum. Process. Biochem. 2008, 43, 1359–1370. [Google Scholar]
- Albalasmeh, A.; Berhe, A.A.; Ghezzehei, T.A. A new method for rapid determination of carbohydrate and total carbon con-centrations using UV spectrophotometry. Carbohyd. Polym. 2013, 97, 253–261. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Elicitor | Days of Cultivation | trans-Resveratrol | trans-Piceid | ε-Viniferin | δ-Viniferin |
|---|---|---|---|---|---|
| MeJA1 | 8 | 8.5 ± 0.52 ab | 82.0 ± 4.10 e | 29.2 ± 1.46 a | 647.1 ± 22.42 c |
| cJA | 12.0 ± 0.07 b | 51.6 ± 1.81 a | n.d. | 985.5 ± 48.61 d | |
| 2HJA | 9.4 ± 0.14 ab | 53.1 ± 0.81 a | n.d. | 410.9 ± 22.63 b | |
| MeJA2 | 55.6 ± 4.17 d | 76.1 ± 1.58 d | 1141.4 ± 57.45 b | 2144.4 ± 113.47 e | |
| TvC1 | 132.2 ± 5.76 f | 69.2 ± 3.48 c | 2188.1 ± 167.12 c | 2662.9 ± 15.37 f | |
| TvC2 | 115.6 ± 5.21 e | 76.4 ± 4.34 d | 2836.5 ± 43.33 d | 2907.2 ± 102.10 g | |
| NaVO | 5.5 ± 0.22 a | 58.9 ± 1.80 bc | 121.7 ± 6.25 a | 248.3 ± 7.59 a | |
| FOE | 32.9 ± 0.95 c | 94.7 ± 5.22 f | 25.4 ± 0.76 a | 193.9 ± 5.13 a | |
| Control | 12.8 ± 0.56 b | 60.4 ± 2.12 ab | 28.6 ± 1.03 a | 571.3 ± 20.60 c | |
| MeJA1 | 11 | 39.0 ± 1.62 c | 128.5 ± 4.86 f | 71.5 ± 6.11 a | 834.2 ± 55.54 d |
| cJA | 5.2 ± 0.16 ab | 58.4 ± 2.21 a | n.d. | 376.3 ± 17.38 b | |
| 2HJA | n.d. | 60.3 ± 3.04 a | n.d. | 85.6 ± 4.71 a | |
| MeJA2 | 181.3 ± 5.44 f | 99.6 ± 4.60 e | 1859.6 ± 18.60 c | 3670.3 ± 184.73 e | |
| TvC1 | 161.5 ± 7.28 e | 81.7 ± 3.30 c | 3151.3 ± 157.57 d | 4486.2 ± 89.72 f | |
| TvC2 | 183.0 ± 9.15 f | 100.1 ± 4.59 d | 3267.6 ± 149.74 e | 3960.7 ± 218.14 e | |
| NaVO | 12.2 ± 0.25 b | 66.2 ± 2.32 b | 233.6 ± 11.68 b | 634.3 ± 44.40 c | |
| FOE | 48.6 ± 2.85 d | 186.2 ± 7.45 g | 61.8 ± 3.73 a | 684.7 ± 27.39 cd | |
| Control | 4.1 ± 0.17 a | 57.0 ± 3.42 a | 47.2 ± 3.35 a | 304.1 ± 13.71 b | |
| MeJA1 | 13 | 64.1 ± 2.59 b | 69.8 ± 4.58 b | 116.2 ± 2.92 a | 1579.7 ± 118.57 d |
| cJA | n.d. | 61.1 ± 2.44 a | n.d. | 143.2 ± 5.16 a | |
| 2HJA | n.d. | 87.6 ± 3.65 d | n.d. | 85.7 ± 0.49 a | |
| MeJA2 | 126.9 ± 2.64 c | 122.6 ± 3.75 f | 1236.9 ± 49.48 c | 2652.6 ± 136.12 e | |
| TvC1 | 169.9 ± 7.79 d | 70.3 ± 2.47 b | 2179.6 ± 132.58 d | 3518.2 ± 158.64 f | |
| TvC2 | 352.3 ± 7.05 e | 122.5 ± 2.12 e | 3226.6 ± 322.66 e | 4587.2 ± 199.95 g | |
| NaVO | 65.6 ± 4.47 b | 127.0 ± 3.88 f | 667.9 ± 34.27 b | 1327.2 ± 53.09 c | |
| FOE | 58.1 ± 2.01 b | 193.0 ± 6.78 g | 56.2 ± 2.58 a | 1189.1 ± 79.17 c | |
| Control | 8.8 ± 0.18 a | 76.0 ± 1.91 c | 60.8 ± 4.86 a | 473.4 ± 17.07 b |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sák, M.; Dokupilová, I.; Kaňuková, Š.; Mrkvová, M.; Mihálik, D.; Hauptvogel, P.; Kraic, J. Biotic and Abiotic Elicitors of Stilbenes Production in Vitis vinifera L. Cell Culture. Plants 2021, 10, 490. https://0-doi-org.brum.beds.ac.uk/10.3390/plants10030490
Sák M, Dokupilová I, Kaňuková Š, Mrkvová M, Mihálik D, Hauptvogel P, Kraic J. Biotic and Abiotic Elicitors of Stilbenes Production in Vitis vinifera L. Cell Culture. Plants. 2021; 10(3):490. https://0-doi-org.brum.beds.ac.uk/10.3390/plants10030490
Chicago/Turabian StyleSák, Martin, Ivana Dokupilová, Šarlota Kaňuková, Michaela Mrkvová, Daniel Mihálik, Pavol Hauptvogel, and Ján Kraic. 2021. "Biotic and Abiotic Elicitors of Stilbenes Production in Vitis vinifera L. Cell Culture" Plants 10, no. 3: 490. https://0-doi-org.brum.beds.ac.uk/10.3390/plants10030490