
Inhibition of Solar UV-Induced Matrix Metalloproteinase (MMP)-1 Expression by Non-Enzymatic Softening Cherry Blossom (Prunus yedoensis) Extract

Abstract
:1. Introduction
2. Results
2.1. Antioxidant Effects and Contents of Total Phenolics in NES-CBE
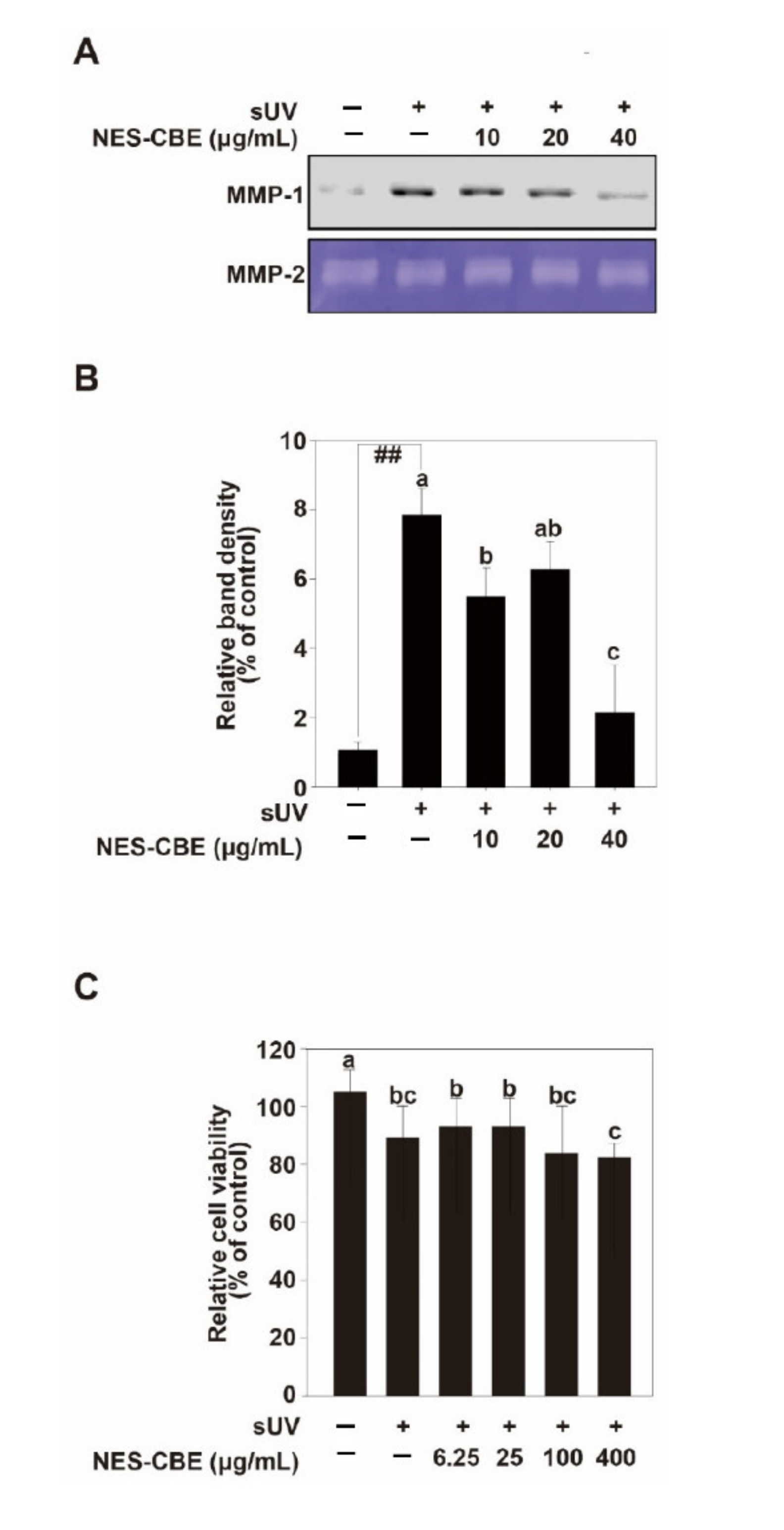
2.2. Effects of NES-CBE on sUV-Induced MMP-1 Expression
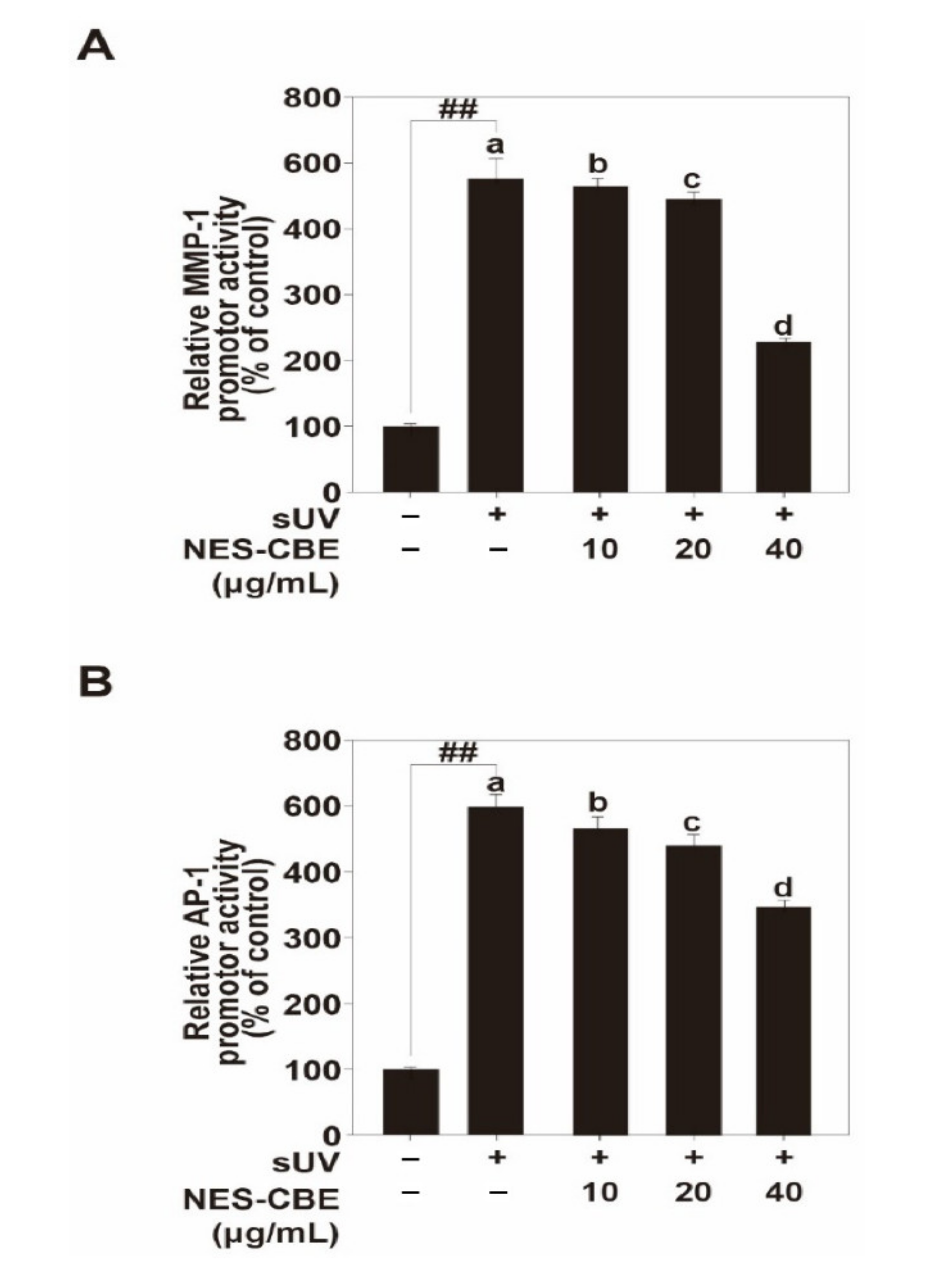
2.3. Effects of NES-CBE on sUV-Induced MMP-1 Promoter Activity by Reducing AP-1 Transactivation
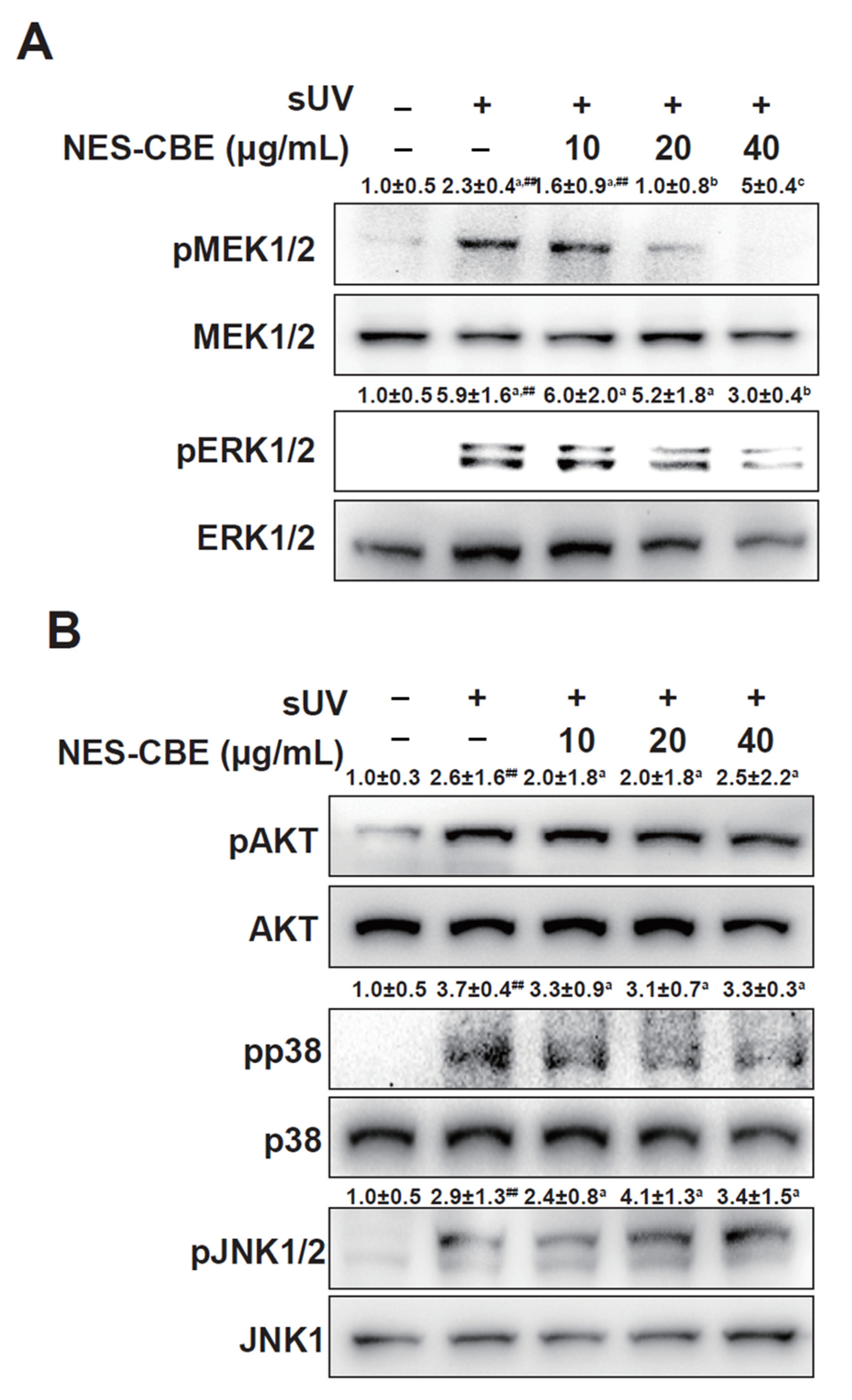
2.4. Effects of NES-CBE on sUV-Induced ERK1/2 Pathways
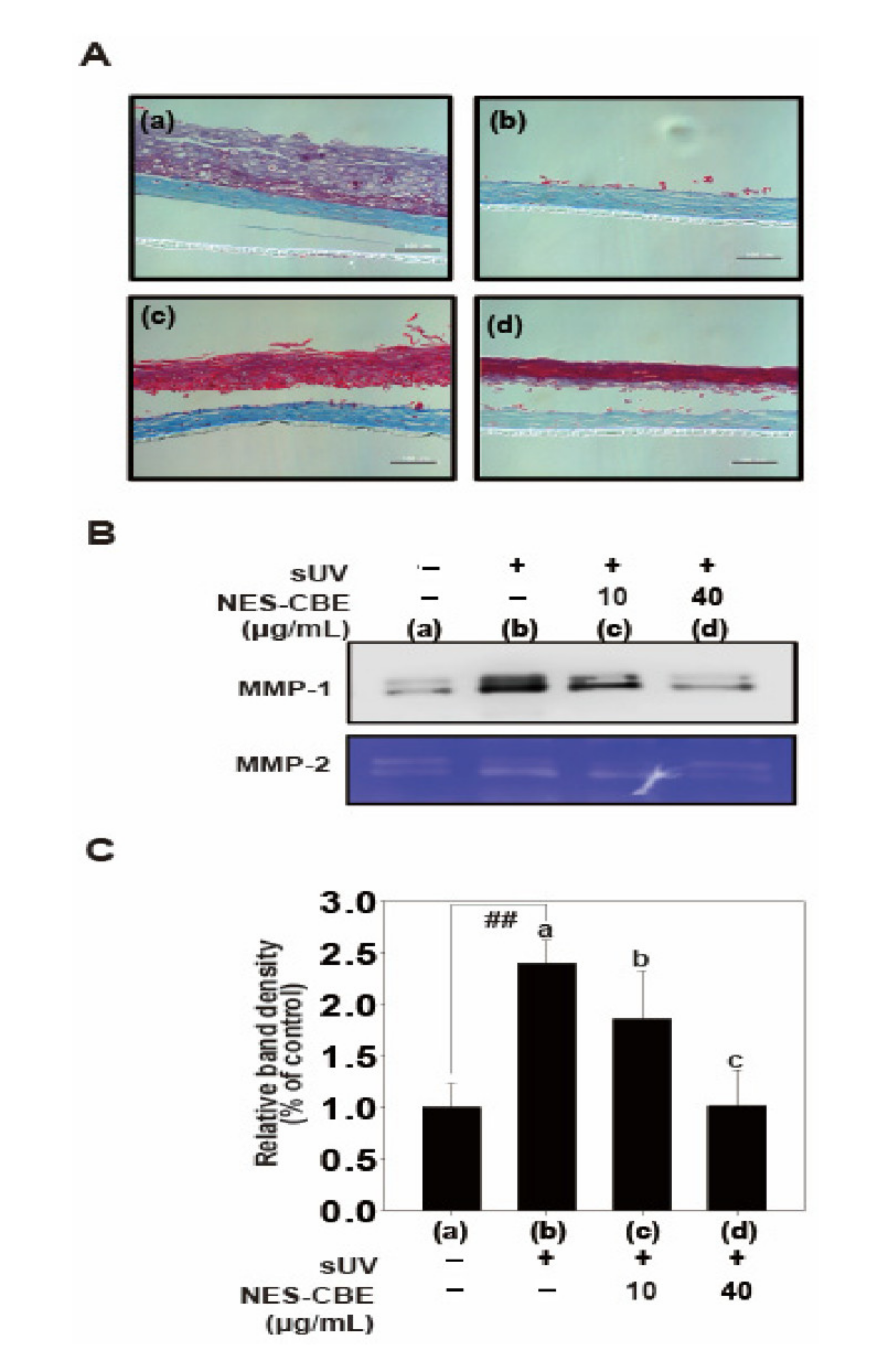
2.5. Effects of NES-CBE on sUV-Induced Skin Damage and MMP-1 Protein Expression in Human Skin Equivalent
3. Discussion
4. Materials and Methods
4.1. Chemicals
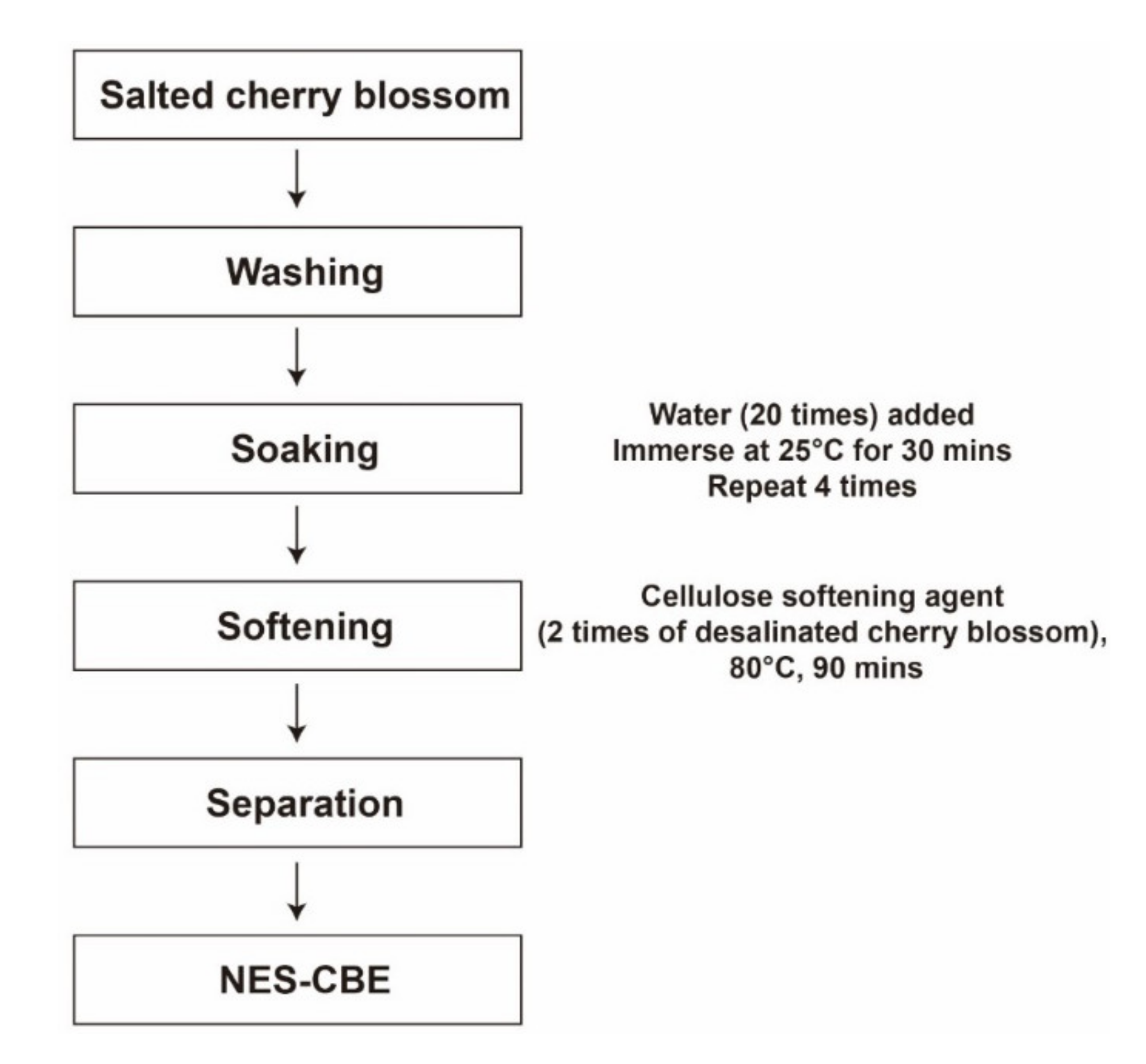
4.2. Preparing NES-CBE
4.3. ABTS Radical Scavenging Activity
4.4. DPPH Radical Scavenging Activity
4.5. Total Phenolic Content
4.6. Cell Culture
4.7. sUV Irradiation
4.8. Cell Viability
4.9. Western Blot
4.10. Gelatin Zymography
4.11. Luciferase Reporter Gene Assay
4.12. Human Skin Equivalent Preparation
4.13. Statistical Analysis
Author Contributions
Funding
Conflicts of Interest
References
- Lopez-Ojeda, W.; Pandey, A.; Alhajj, M.; Oakley, A.M. Anatomy, Skin (Integument). In StatPearls; StatPearls Publishing: Treasure Island, FL, USA, 2021. [Google Scholar]
- Gruber, F.; Marchetti-Deschmann, M.; Kremslehner, C.; Schosserer, M. The Skin Epilipidome in Stress, Aging, and Inflammation. Front. Endocrinol. 2020, 11, 607076. [Google Scholar] [CrossRef]
- Wang, A.S.; Dreesen, O. Biomarkers of Cellular Senescence and Skin Aging. Front. Genet. 2018, 9, 247. [Google Scholar] [CrossRef]
- Park, G.; Baek, S.; Kim, J.E.; Lim, T.G.; Lee, C.C.; Yang, H.; Kang, Y.G.; Park, J.S.; Augustin, M.; Mrosek, M.; et al. Flt3 is a target of coumestrol in protecting against UVB-induced skin photoaging. Biochem. Pharmacol. 2015, 98, 473–483. [Google Scholar] [CrossRef] [PubMed]
- Masamitsu Ichihashi, H.A.; Yoshida, M.; Niki, Y.; Matsui, M. Photoaging of the skin. Anti-Aging Med. 2009, 6, 46–59. [Google Scholar] [CrossRef] [Green Version]
- Kim, J.E.; Lee, K.W. Molecular Targets of Phytochemicals for Skin Inflammation. Curr. Pharm. Des. 2018, 24, 1533–1550. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.E.; Heo, Y.S.; Lee, K.W. Osajin Inhibits Solar UV-Induced Cyclooxygenase-2 Expression Through Direct Inhibition of RSK2. J. Cell Biochem. 2017, 118, 4080–4087. [Google Scholar] [CrossRef]
- Gu, Y.; Han, J.; Jiang, C.; Zhang, Y. Biomarkers, oxidative stress and autophagy in skin aging. Ageing Res. Rev. 2020, 59, 101036. [Google Scholar] [CrossRef]
- Lee, B.B.; Cha, M.R.; Kim, S.Y.; Park, E.; Park, H.R.; Lee, S.C. Antioxidative and anticancer activity of extracts of cherry (Prunus serrulata var. spontanea) blossoms. Plant. Foods Hum. Nutr. 2007, 62, 79–84. [Google Scholar] [CrossRef]
- Matsuura, R.; Moriyama, H.; Takeda, N.; Yamamoto, K.; Morita, Y.; Shimamura, T.; Ukeda, H. Determination of antioxidant activity and characterization of antioxidant phenolics in the plum vinegar extract of cherry blossom (Prunus lannesiana). J. Agric. Food Chem. 2008, 56, 544–549. [Google Scholar] [CrossRef]
- Lee, K.W.; Kim, Y.J.; Lee, H.J.; Lee, C.Y. Cocoa has more phenolic phytochemicals and a higher antioxidant capacity than teas and red wine. J. Agric. Food Chem. 2003, 51, 7292–7295. [Google Scholar] [CrossRef] [PubMed]
- Roh, E.; Kim, J.E.; Kwon, J.Y.; Park, J.S.; Bode, A.M.; Dong, Z.; Lee, K.W. Molecular mechanisms of green tea polyphenols with protective effects against skin photoaging. Crit. Rev. Food Sci. Nutr. 2017, 57, 1631–1637. [Google Scholar] [CrossRef]
- Ozanne, B.W.; Spence, H.J.; McGarry, L.C.; Hennigan, R.F. Transcription factors control invasion: AP-1 the first among equals. Oncogene 2007, 26, 1–10. [Google Scholar] [CrossRef] [Green Version]
- Xu, Y.; Fisher, G.J. Ultraviolet (UV) light irradiation induced signal transduction in skin photoaging. J. Dermatol. Sci. Suppl. 2005, 1, S1–S8. [Google Scholar] [CrossRef]
- Todorova, K.; Mandinova, A. Novel approaches for managing aged skin and nonmelanoma skin cancer. Adv. Drug Deliv. Rev. 2020, 153, 18–27. [Google Scholar] [CrossRef] [PubMed]
- Li, L.; Hwang, E.; Ngo, H.T.T.; Lin, P.; Gao, W.; Liu, Y.; Yi, T.H. Antiphotoaging Effect of Prunus yeonesis Blossom Extract via Inhibition of MAPK/AP-1 and Regulation of the TGF-βI/Smad and Nrf2/ARE Signaling Pathways. Photochem. Photobiol. 2018, 94, 725–732. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.Q.; Guan, L.; Zhong, Z.Y.; Chang, M.; Zhang, D.K.; Li, H.; Lai, W. The anti-inflammatory effect of cherry blossom extract (Prunus yedoensis) used in soothing skincare product. Int. J. Cosmet. Sci. 2014, 36, 527–530. [Google Scholar] [CrossRef]
- Song, Y.R.; Lim, W.C.; Han, A.; Lee, M.H.; Shin, E.J.; Lee, K.M.; Nam, T.G.; Lim, T.G. Rose Petal Extract (Rosa gallica) Exerts Skin Whitening and Anti-Skin Wrinkle Effects. J. Med. Food 2020, 23, 870–878. [Google Scholar] [CrossRef] [PubMed]
- Shin, E.J.; Han, A.R.; Lee, M.H.; Song, Y.R.; Lee, K.M.; Nam, T.G.; Lee, P.; Lee, S.Y.; Lim, T.G. Extraction conditions for Rosa gallica petal extracts with anti-skin aging activities. Food Sci. Biotechnol. 2019, 28, 1439–1446. [Google Scholar] [CrossRef]
- Shin, J.; Kim, J.E.; Pak, K.J.; Kang, J.I.; Kim, T.S.; Lee, S.Y.; Yeo, I.H.; Park, J.H.; Kim, J.H.; Kang, N.J.; et al. A Combination of Soybean and Haematococcus Extract Alleviates Ultraviolet B-Induced Photoaging. Int. J. Mol. Sci. 2017, 18. [Google Scholar] [CrossRef] [Green Version]
- Marionnet, C.; Tricaud, C.; Bernerd, F. Exposure to non-extreme solar UV daylight: Spectral characterization, effects on skin and photoprotection. Int. J. Mol. Sci. 2014, 16, 68–90. [Google Scholar] [CrossRef]
- Kim, J.E.; Song, D.; Kim, J.; Choi, J.; Kim, J.R.; Yoon, H.S.; Bae, J.S.; Han, M.; Lee, S.; Hong, J.S.; et al. Oral Supplementation with Cocoa Extract Reduces UVB-Induced Wrinkles in Hairless Mouse Skin. J. Investig. Dermatol. 2016, 136, 1012–1021. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Han, S.; Lim, T.G.; Kim, J.E.; Yang, H.; Oh, D.K.; Yoon Park, J.H.; Kim, H.J.; Rhee, Y.K.; Lee, K.W. The Ginsenoside Derivative 20(S)-Protopanaxadiol Inhibits Solar Ultraviolet Light-Induced Matrix Metalloproteinase-1 Expression. J. Cell Biochem. 2017, 118, 3756–3764. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Method | ABTS | DPPH | Total Phenolic Content |
|---|---|---|---|
| Comparison | Vitamin C equivalents | Vitamin C equivalents | tannic acid equivalents |
| Unit | mg VCEAC (1)/100 g | mg VCEAC/100 g | mg TAE (2)/g |
| Value | 1499.31 ± 7.64 | 564.64 ± 6.40 | 36.86 ± 0.61 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jung, Y.-A.; Lee, J.-Y.; Lee, P.; Shin, H.-S.; Kim, J.-E. Inhibition of Solar UV-Induced Matrix Metalloproteinase (MMP)-1 Expression by Non-Enzymatic Softening Cherry Blossom (Prunus yedoensis) Extract. Plants 2021, 10, 1016. https://0-doi-org.brum.beds.ac.uk/10.3390/plants10051016
Jung Y-A, Lee J-Y, Lee P, Shin H-S, Kim J-E. Inhibition of Solar UV-Induced Matrix Metalloproteinase (MMP)-1 Expression by Non-Enzymatic Softening Cherry Blossom (Prunus yedoensis) Extract. Plants. 2021; 10(5):1016. https://0-doi-org.brum.beds.ac.uk/10.3390/plants10051016
Chicago/Turabian StyleJung, Yeong-A, Ji-Yoon Lee, Pomjoo Lee, Han-Seung Shin, and Jong-Eun Kim. 2021. "Inhibition of Solar UV-Induced Matrix Metalloproteinase (MMP)-1 Expression by Non-Enzymatic Softening Cherry Blossom (Prunus yedoensis) Extract" Plants 10, no. 5: 1016. https://0-doi-org.brum.beds.ac.uk/10.3390/plants10051016