
Ulva lactuca Extract and Fractions as Seed Priming Agents Mitigate Salinity Stress in Tomato Seedlings

 , ,
, ,
Abstract
:1. Introduction
2. Results
2.1. Biochemical Characterization of Ulva lactuca Extract Fractions and Yield
2.2. Effect of Ulva lactuca Extract on Tomato Growth Traits
2.3. Effect of Ulva lactuca Extract on Tomato Antioxidant Activity
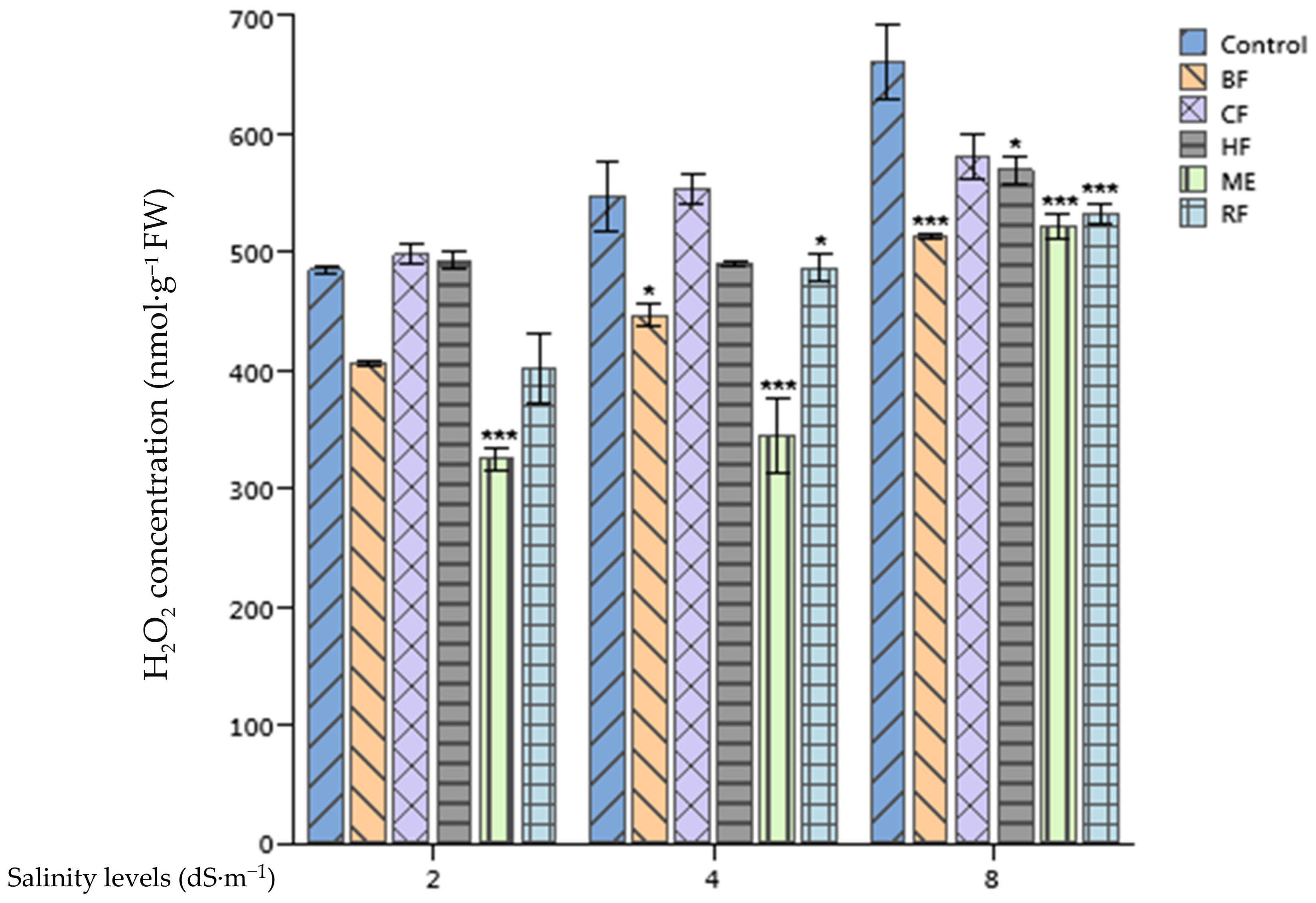
2.4. Effect of Ulva lactuca Extract Fractions on Tomato Hydrogen Peroxide Concentration
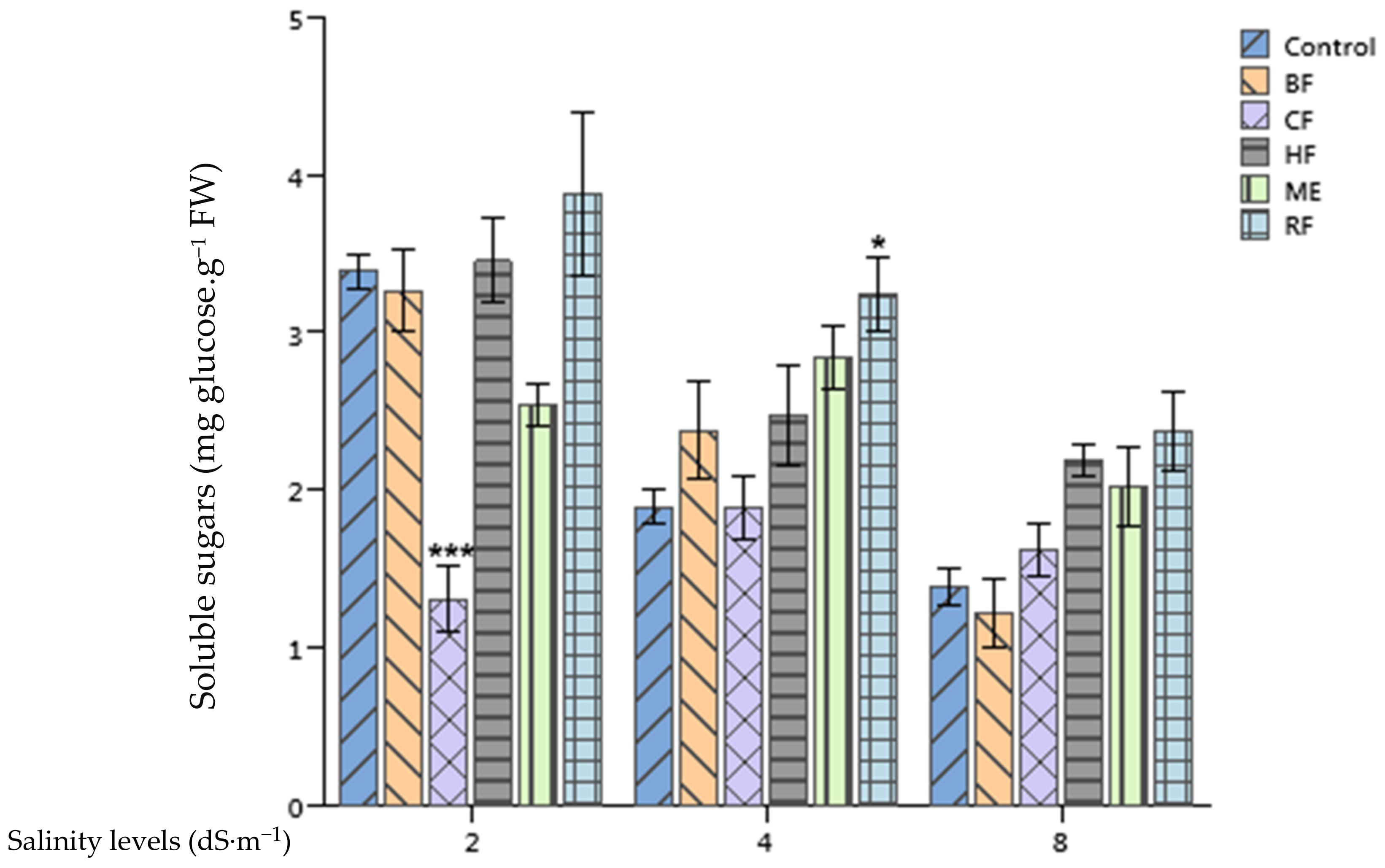
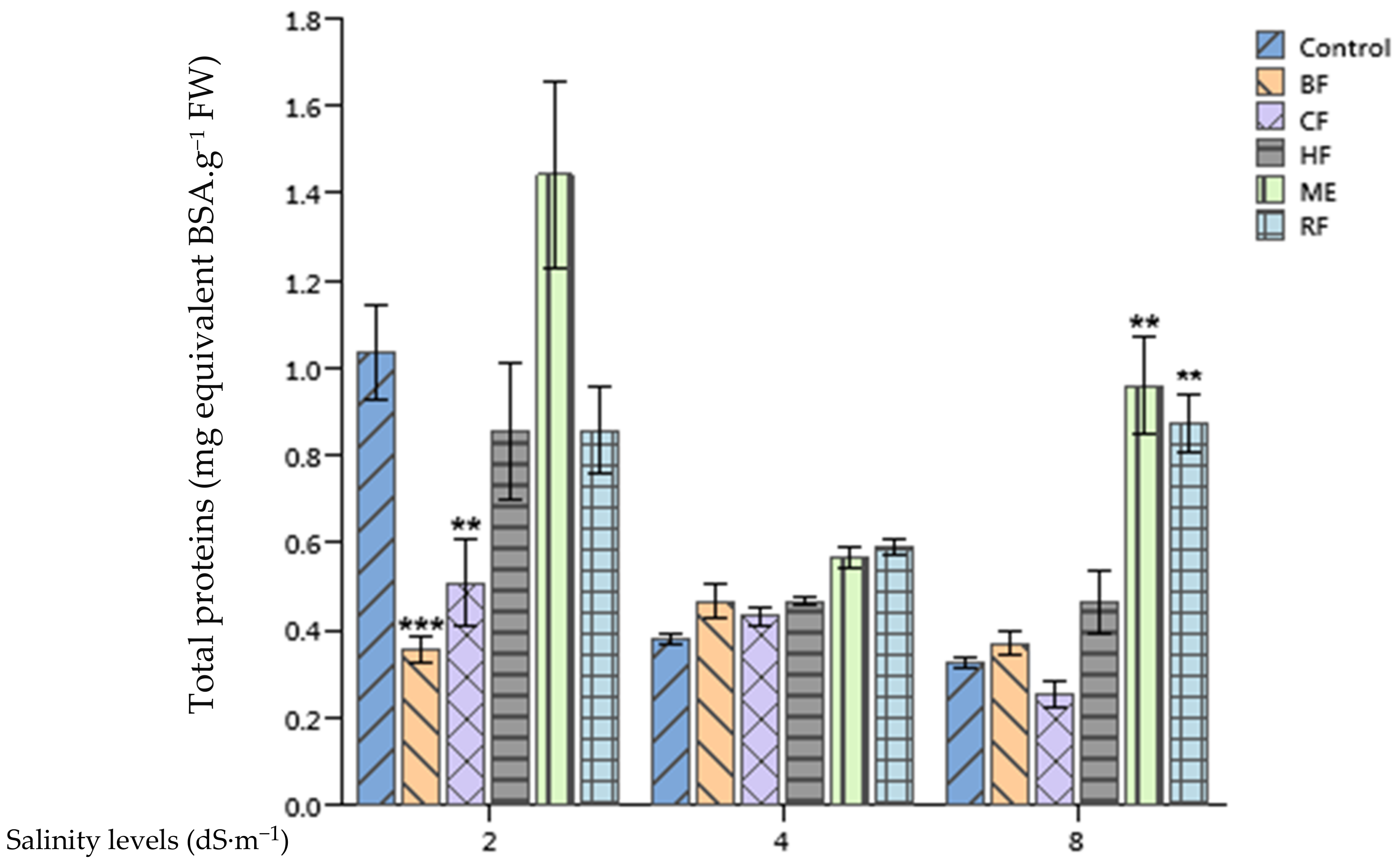
2.5. Effect of Ulva lactuca Extract Fractions on Tomato Soluble Sugars and Total Proteins Content
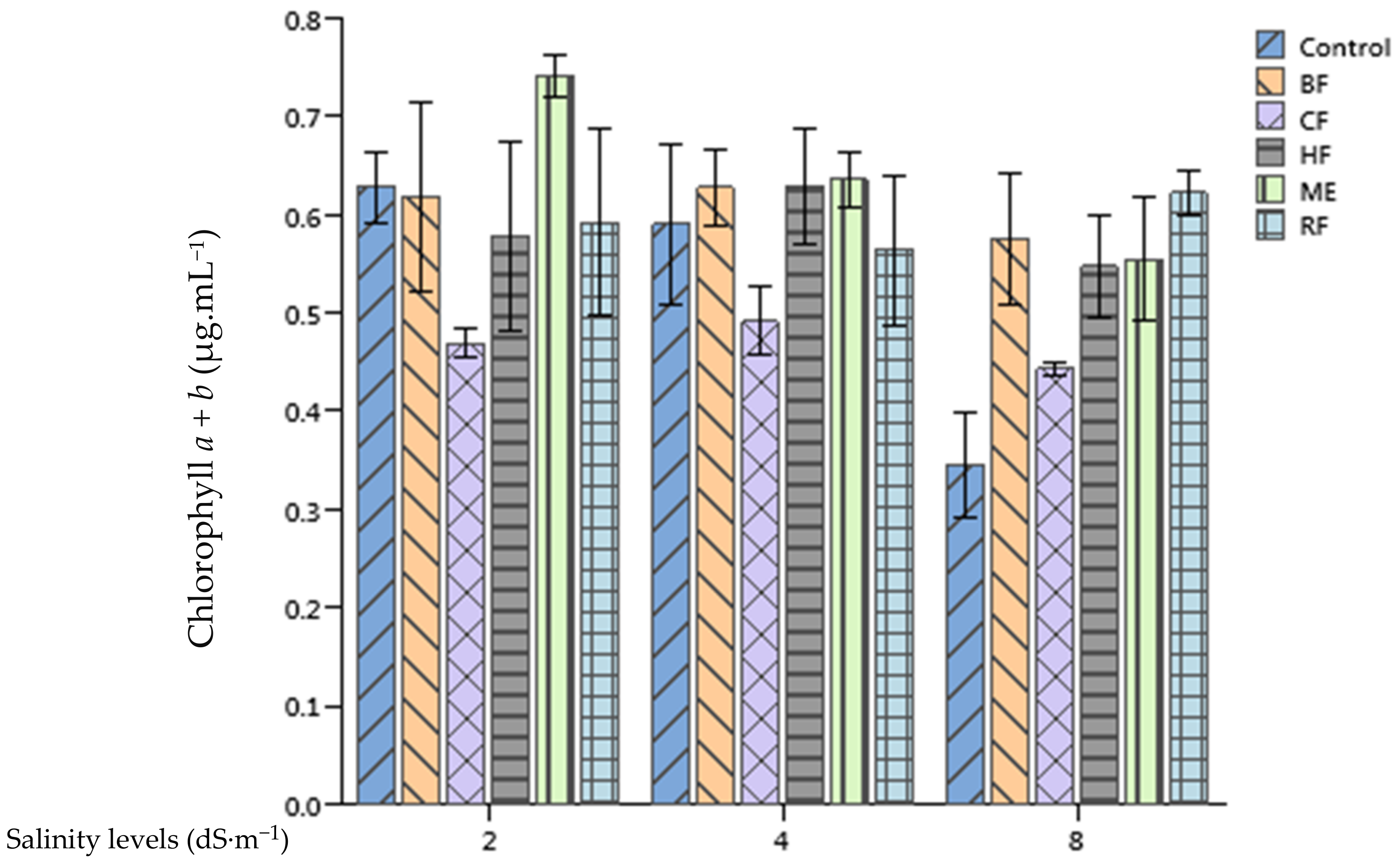
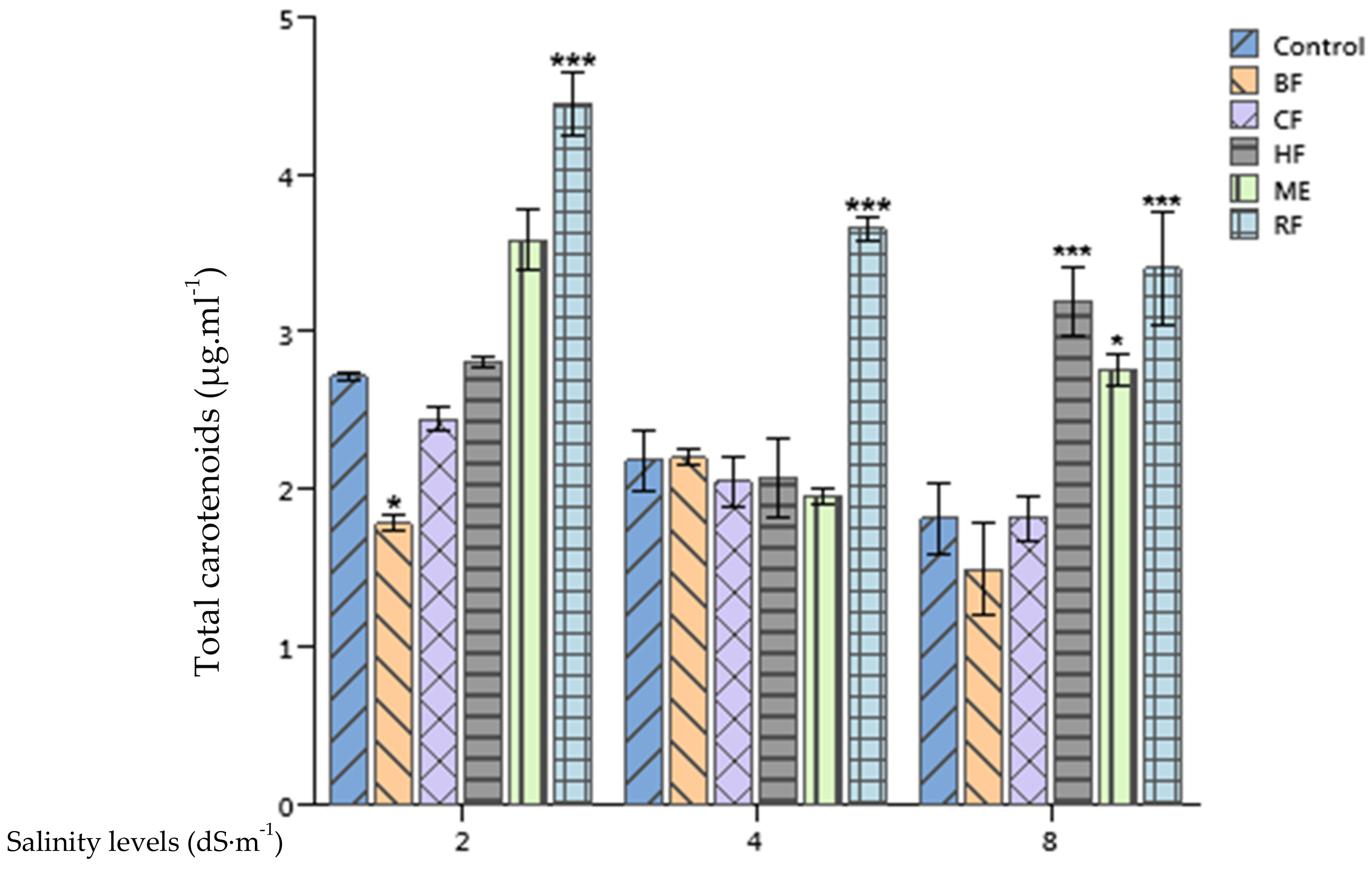
2.6. Effect of Ulva lactuca Extract Fractions on Tomato Leaves Chlorophyll and Carotenoides
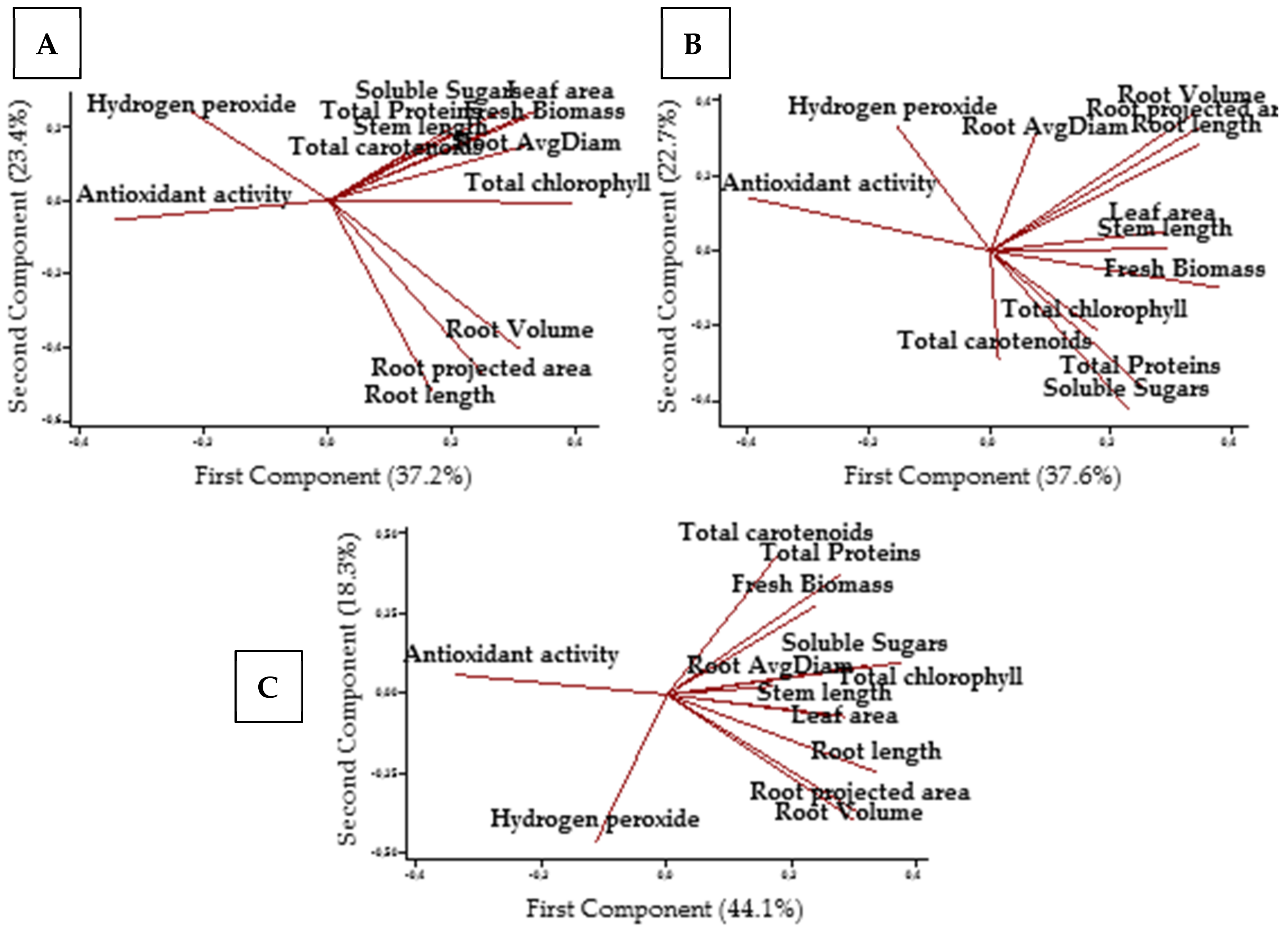
2.7. Principal Component Analysis (PCA)
3. Discussion
4. Material and Methods
4.1. Ulva lactuca Sampling and Characterization
4.2. Solvent Solvent Extraction and Fractionation
4.3. Seaweed Extract Fractions Characterization
4.3.1. Determination of Total Phenolic Content and Total Flavonoids
4.3.2. Determination of Glycine Betaine Concentration
4.3.3. Quantification of Soluble Sugars
4.4. Mode of Application of Seaweed Extracts as Seed Priming Agents
4.5. Plant Growth Conditions
4.5.1. Determination of Plant Shoot and Root Biomass
4.5.2. Salinity Stress Application
4.5.3. Determination of Antioxidant Activity
4.5.4. Hydrogen Peroxide (H2O2) Concentration
4.5.5. Soluble Sugars and Total Proteins Content
4.5.6. Chlorophyll a, b and Total Carotenoids
4.6. Statistical Analyses
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- The State of Food Security and Nutrition in the World 2020. Available online: https://data.unicef.org/resources/sofi-2020/ (accessed on 31 January 2021).
- Dobermann, A.; Nelson, R. Opportunities and Solutions for Sustainable Food Production; Sustainable Development Solutions Network: Paris, France, 2013. [Google Scholar]
- Mancosu, N.; Snyder, R.L.; Kyriakakis, G.; Spano, D. Water scarcity and future challenges for food production. Water 2015, 7, 975–992. [Google Scholar] [CrossRef]
- McKenzie, F.C.; Williams, J. Sustainable food production: Constraints, challenges and choices by 2050. Food Secur. 2015, 7, 221–233. [Google Scholar] [CrossRef]
- Shahid, S.A.; Zaman, M.; Heng, L. Soil salinity: Historical perspectives and a world overview of the problem. In Guideline for Salinity Assessment, Mitigation and Adaptation Using Nuclear and Related Techniques; Springer Science and Business Media LLC: Berlin/Heidelberg, Germany, 2018; pp. 43–53. [Google Scholar]
- He, Y.; Zhou, J.; Hu, Y.; Fang, C.; Yu, Y.; Yang, J.; Zhu, B.; Ruan, Y.-L.; Zhu, Z. Overexpression of sly-miR398b increased salt sensitivity likely via regulating antioxidant system and photosynthesis in tomato. Environ. Exp. Bot. 2021, 181, 104273. [Google Scholar] [CrossRef]
- Shariatinia, F.; Azari, A.; Rahimi, A.; Panahi, B.; Madahhosseini, S. Germination, growth, and yield of rocket populations show strong ecotypic variation under NaCl stress. Sci. Hortic. 2021, 278, 109841. [Google Scholar] [CrossRef]
- Cuevas, J.; Daliakopoulos, I.N.; Del Moral, F.; Hueso, J.J.; Tsanis, I.K. A review of soil-improving cropping systems for soil salinization. Agronomy 2019, 9, 295. [Google Scholar] [CrossRef] [Green Version]
- Du Jardin, P. Plant Biostimulants: Definition, concept, main categories and regulation. Sci. Hortic. 2015, 196, 3–14. [Google Scholar] [CrossRef] [Green Version]
- Yakhin, O.I.; Lubyanov, A.A.; Yakhin, I.A.; Brown, P.H. Biostimulants in plant science: A global perspective. Front. Plant Sci. 2017, 7, 2049. [Google Scholar] [CrossRef] [Green Version]
- Rouphael, Y.; Colla, G. Synergistic Biostimulatory action: Designing the next generation of plant Biostimulants for sustainable agriculture. Front. Plant Sci. 2018, 9, 1655. [Google Scholar] [CrossRef] [Green Version]
- Shukla, P.S.; Mantin, E.G.; Adil, M.; Bajpai, S.; Critchley, A.T.; Prithiviraj, B. Ascophyllum nodosum-based Biostimulants: Sustainable applications in agriculture for the stimulation of plant growth, stress tolerance, and disease management. Front. Plant Sci. 2019, 10, 655. [Google Scholar] [CrossRef] [Green Version]
- El Boukhari, M.E.; Barakate, M.; Bouhia, Y.; Lyamlouli, K. Trends in seaweed extract based Biostimulants: Manufacturing process and beneficial effect on soil-plant systems. Plants 2020, 9, 359. [Google Scholar] [CrossRef] [Green Version]
- Elansary, H.O.; Yessoufou, K.; Abdel-Hamid, A.; El-Esawi, M.A.; Ali, H.M.; Elshikh, M.S. Seaweed extracts enhance Salam turfgrass performance during prolonged irrigation intervals and saline shock. Front. Plant Sci. 2017, 8, 830. [Google Scholar] [CrossRef] [Green Version]
- Patel, K.; Agarwal, P.; Agarwal, P.K. Kappaphycus Alvarezii sap mitigates abiotic-induced stress in Triticum durum by modulating metabolic coordination and improves growth and yield. J. Appl. Phycol. 2018, 30, 2659–2673. [Google Scholar] [CrossRef]
- Trivedi, K.; Anand, K.V.; Vaghela, P.; Ghosh, A. Differential growth, yield and biochemical responses of maize to the exogenous application of Kappaphycus Alvarezii seaweed extract, at grain-filling stage under normal and drought conditions. Algal Res. 2018, 35, 236–244. [Google Scholar] [CrossRef]
- Liu, H.; Chen, X.; Song, L.; Li, K.; Zhang, X.; Liu, S.; Qin, Y.; Li, P. Polysaccharides from Grateloupia Filicina enhance tolerance of rice seeds (Oryza sativa L.) under salt stress. Int. J. Biol. Macromol. 2019, 124, 1197–1204. [Google Scholar] [CrossRef]
- Zou, P.; Lu, X.; Zhao, H.; Yuan, Y.; Meng, L.; Zhang, C.; Li, Y. Polysaccharides derived from the brown algae Lessonia Nigrescens enhance salt stress tolerance to wheat seedlings by enhancing the antioxidant system and modulating intracellular ion concentration. Front. Plant Sci. 2019, 10, 48. [Google Scholar] [CrossRef] [Green Version]
- Chrysargyris, A.; Xylia, P.; Anastasiou, M.; Pantelides, I.; Tzortzakis, N. Effects of Ascophyllum Nodosum seaweed extracts on lettuce growth, physiology and fresh-cut salad storage under potassium deficiency. J. Sci. Food Agric. 2018, 98, 5861–5872. [Google Scholar] [CrossRef]
- De Carvalho, R.P.; Pasqual, M.; Silveira, H.R.D.O.; De Melo, P.C.; Bispo, D.F.A.; Laredo, R.R.; Lima, L.D.A.S. Niágara Rosada table grape cultivated with seaweed extracts: Physiological, nutritional, and yielding behavior. J. Appl. Phycol. 2019, 31, 2053–2064. [Google Scholar] [CrossRef]
- Rosa, V.D.R.; Dos Santos, A.L.F.; Da Silva, A.A.; Sab, M.P.V.; Germino, G.H.; Cardoso, F.B.; Silva, M.D.A. Increased soybean tolerance to water deficiency through Biostimulant based on fulvic acids and Ascophyllum nodosum (L.) seaweed extract. Plant Physiol. Biochem. 2021, 158, 228–243. [Google Scholar] [CrossRef]
- Sharma, H.S.S.; Fleming, C.C.; Selby, C.; Rao, J.R.; Martin, T. Plant Biostimulants: A review on the processing of macroalgae and use of extracts for crop management to reduce abiotic and biotic stresses. J. Appl. Phycol. 2013, 26, 465–490. [Google Scholar] [CrossRef]
- Masondo, N.A.; Kulkarni, M.G.; Finnie, J.F.; Van Staden, J. Influence of Biostimulants-seed-priming on Ceratotheca triloba germination and seedling growth under low temperatures, low osmotic potential and salinity stress. Ecotoxicol. Environ. Saf. 2018, 147, 43–48. [Google Scholar] [CrossRef]
- Hussein, M.H.; Eltanahy, E.; Al Bakry, A.F.; Elsafty, N.; Elshamy, M.M. Seaweed extracts as prospective plant growth bio-stimulant and salinity stress alleviator for Vigna sinensis and Zea mays. J. Appl. Phycol. 2021, 33, 1273–1291. [Google Scholar] [CrossRef]
- Houghton, P.J.; Raman, A. Laboratory Handbook for the Fractionation of Natural Extracts; Metzler, J.B., Ed.; Springer: Berlin/Heidelberg, Germany, 1998. [Google Scholar]
- Bannour, M.; Khadhri, A.; Venskutonis, P.R.; Baranauskiene, R.; Lachenmeier, D.W. Conventional and innovative extraction methods applied on Calligonum Azel Maire leaves and roots: A comparative study. Eur. Food Res. Technol. 2021, 247, 637–649. [Google Scholar] [CrossRef]
- Chanthini, K.M.-P.; Stanley-Raja, V.; Thanigaivel, A.; Karthi, S.; Palanikani, R.; Sundar, N.S.; Sivanesh, H.; Soranam, R.; Senthil-Nathan, S. Sustainable agronomic strategies for enhancing the yield and nutritional quality of wild tomato, Solanum Lycopersicum (L.) var Cerasiforme mill. Agronomy 2019, 9, 311. [Google Scholar] [CrossRef] [Green Version]
- Cisse, E.H.M.; Miao, L.-F.; Yang, F.; Huang, J.-F.; Li, D.-D.; Zhang, J. Gly Betaine surpasses melatonin to improve salt tolerance in Dalbergia Odorifera. Front. Plant Sci. 2021, 12. [Google Scholar] [CrossRef]
- Acosta-Motos, J.R.; Penella, C.; Hernández, J.A.; Díaz-Vivancos, P.; Sánchez-Blanco, M.J.; Navarro, J.M.; Gómez-Bellot, M.J.; Barba-Espín, G. Towards a sustainable agriculture: Strategies involving Phytoprotectants against salt stress. Agronomy 2020, 10, 194. [Google Scholar] [CrossRef] [Green Version]
- Giri, J. Glycinebetaine and abiotic stress tolerance in plants. Plant Signal. Behav. 2011, 6, 1746–1751. [Google Scholar] [CrossRef]
- Di Stasio, E.; Cirillo, V.; Raimondi, G.; Giordano, M.; Esposito, M.; Maggio, A. Osmo-priming with seaweed extracts enhances yield of salt-stressed tomato plants. Agronomy 2020, 10, 1559. [Google Scholar] [CrossRef]
- Debnath, M.; Pandey, M.; Bisen, P. An omics approach to understand the plant abiotic stress. OMICS J. Integr. Biol. 2011, 15, 739–762. [Google Scholar] [CrossRef]
- Mittler, R. Oxidative stress, antioxidants and stress tolerance. Trends Plant Sci. 2002, 7, 405–410. [Google Scholar] [CrossRef]
- Das, K.; Roychoudhury, A. Reactive oxygen species (ROS) and response of antioxidants as ROS-scavengers during environmental stress in plants. Front. Environ. Sci. 2014, 2, 1–13. [Google Scholar] [CrossRef] [Green Version]
- Gill, S.S.; Tuteja, N. Reactive oxygen species and antioxidant machinery in abiotic stress tolerance in crop plants. Plant Physiol. Biochem. 2010, 48, 909–930. [Google Scholar] [CrossRef] [PubMed]
- Kowalczewski, P.Ł.; Radzikowska, D.; Ivanišová, E.; Szwengiel, A.; Kačániová, M.; Sawinska, Z. Influence of abiotic stress factors on the antioxidant properties and polyphenols profile composition of green barley (Hordeum vulgare L.). Int. J. Mol. Sci. 2020, 21, 397. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dawange, P.; Jaiswar, S. Effects of Ascophyllum marine plant extract powder (AMPEP) on tissue growth, proximate, phenolic contents, and free radical scavenging activities in endemic red seaweed Gracilaria Corticata var. cylindrica from India. J. Appl. Phycol. 2020, 32, 4127–4135. [Google Scholar] [CrossRef]
- Al-Ghamdi, A.A.; Elansary, H.O. Synergetic effects of 5-aminolevulinic acid and Ascophyllum nodosum seaweed extracts on Asparagus phenolics and stress related genes under saline irrigation. Plant Physiol. Biochem. 2018, 129, 273–284. [Google Scholar] [CrossRef]
- Di Mola, I.; Ottaiano, L.; Cozzolino, E.; Senatore, M.; Giordano, M.; El-Nakhel, C.; Sacco, A.; Rouphael, Y.; Colla, G.; Mori, M. Plant-based Biostimulants influence the agronomical, physiological, and qualitative responses of baby rocket leaves under diverse nitrogen conditions. Plants 2019, 8, 522. [Google Scholar] [CrossRef] [Green Version]
- Jang, S.J.; Park, H.H.; Kuk, Y.I. Growth promotion, nutrition levels, and antioxidant activity in Peucedanum japonicum under various plant extracts. Agronomy 2020, 10, 1494. [Google Scholar] [CrossRef]
- González-González, M.F.; Ocampo-Alvarez, H.; Santacruz-Ruvalcaba, F.; Sánchez-Hernández, C.V.; Casarrubias-Castillo, K.; Becerril-Espinosa, A.; Castañeda-Nava, J.J.; Hernández-Herrera, R.M. Physiological, ecological, and biochemical implications in tomato plants of two plant Biostimulants: Arbuscular mycorrhizal fungi and seaweed extract. Front. Plant Sci. 2020, 11, 999. [Google Scholar] [CrossRef]
- Carmody, N.; Goñi, O.; Łangowski, Ł.; O’Connell, S. Ascophyllum nodosum extract Biostimulant processing and its impact on enhancing heat stress tolerance during tomato fruit set. Front. Plant Sci. 2020, 11, 807. [Google Scholar] [CrossRef]
- Castellanos-Barriga, L.G.; Santacruz-Ruvalcaba, F.; Hernández-Carmona, G.; Ramírez-Briones, E.; Hernández-Herrera, R.M. Effect of seaweed liquid extracts from Ulva lactuca on seedling growth of mung bean (Vigna radiata). J. Appl. Phycol. 2017, 29, 2479–2488. [Google Scholar] [CrossRef]
- Michalak, I.; Górka, B.; Wieczorek, P.P.; Rój, E.; Lipok, J.; Łęska, B.; Messyasz, B.; Wilk, R.; Schroeder, G.; Dobrzyńska-Inger, A.; et al. Supercritical fluid extraction of algae enhances levels of biologically active compounds promoting plant growth. Eur. J. Phycol. 2016, 51, 243–252. [Google Scholar] [CrossRef]
- Whapham, C.A.; Blunden, G.; Jenkins, T.; Hankins, S.D. Significance of betaines in the increased chlorophyll content of plants treated with seaweed extract. J. Appl. Phycol. 1993, 5, 231–234. [Google Scholar] [CrossRef]
- Hamani, A.K.M.; Wang, G.; Soothar, M.K.; Shen, X.; Gao, Y.; Qiu, R.; Mehmood, F. Responses of leaf gas exchange attributes, photosynthetic pigments and antioxidant enzymes in NaCl-stressed cotton (Gossypium Hirsutum L.) seedlings to exogenous glycine betaine and salicylic acid. BMC Plant Biol. 2020, 20, 434. [Google Scholar] [CrossRef]
- Wadas, W.; Dziugieł, T. Changes in Assimilation Area and Chlorophyll Content of Very Early Potato (Solanum tuberosum L.) Cultivars as Influenced by Biostimulants. Agronomy 2020, 10, 387. [Google Scholar] [CrossRef] [Green Version]
- Ainsworth, E.A.; Gillespie, K.M. Estimation of total phenolic content and other oxidation substrates in plant tissues using Folin–Ciocalteu reagent. Nat. Protoc. 2007, 2, 875–877. [Google Scholar] [CrossRef]
- Herald, T.J.; Gadgil, P.; Tilley, M. High-throughput micro plate assays for screening flavonoid content and DPPH-scavenging activity in sorghum bran and flour. J. Sci. Food Agric. 2012, 92, 2326–2331. [Google Scholar] [CrossRef]
- Grieve, C.M.; Grattan, S.R. Rapid assay for determination of water soluble quaternary ammonium compounds. Plant Soil 1983, 70, 303–307. [Google Scholar] [CrossRef]
- Dubois, M.; Gilles, K.A.; Hamilton, J.K.; Rebers, P.A.; Smith, F. Colorimetric Method for Determination of Sugars and Related Substances. Anal. Chem. 1956, 28, 350–356. [Google Scholar] [CrossRef]
- Hoagland, D.R.; Arnon, D.I. The Water-Culture Method for Growing Plants without Soil. Circular. Calif. Agric. Exp. Stn. 1950, 347, 32. [Google Scholar]
- Blois, M.S. Antioxidant Determinations by the Use of a Stable Free Radical. Nat. Cell Biol. 1958, 181, 1199–1200. [Google Scholar] [CrossRef]
- Velikova, V.; Yordanov, I.; Edreva, A. Oxidative stress and some antioxidant systems in acid rain-treated bean plants. Plant Sci. 2000, 151, 59–66. [Google Scholar] [CrossRef]
- Bradford, M.M. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-Dye binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef]
- Lichtenthaler, H.K. Chlorophylls and carotenoids: Pigments of photosynthetic biomembranes. In Methods in Enzymology; Academic Press: Orlando, FL, USA, 1987; Volume 148, pp. 350–382. [Google Scholar]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Extract Fraction | Total Phenols Content (mg GAE·g−1 DW) | Total Flavonoids (mg Quercitine·g−1 DW) | Glycine Betaine (mg·g−1 DW) | Soluble Sugars (mg Glucose·g−1 DW) | Extraction Yield (%) |
|---|---|---|---|---|---|
| ME | 112.640 a | 4.870 a | 7.290 b | 0.296 a | 20.47 bc |
| HF | 84.047 b | 3.073 b | 1.97 e | 0.140 c | 24.46 b |
| CF | 54.057 c | 1.953 c | 3.79 c | 0.156 e | 5.99 d |
| BF | 61.904 c | 1.230 d | 2.293 d | 0.150 d | 18.65 c |
| RF | 79.971 b | 1.955 c | 8.033 a | 0.178 b | 49.39 a |
| Fractions | Shoot Weight (g) | Leaf Area (cm2) | Root Length (cm) | ||||||
| 2 dS·m−1 | 4 dS·m−1 | 8 dS·m−1 | 2 dS·m−1 | 4 dS·m−1 | 8 dS·m−1 | 2 dS·m−1 | 4 dS·m−1 | 8 dS·m−1 | |
| Control | 2.49 ± 0.31 | 1.68 ± 0.28 | 1.22 ± 0.15 | 94.86 ± 7.29 | 85.82 ± 5.27 | 73.18 ± 6.79 | 747.4 ± 99.8 | 756.3 ± 221.4 | 419.7 ± 22.7 |
| ME | 2.68 ± 0.44 | 2.35 ± 0.23 * | 1.44 ± 0.21 | 100.38 ± 6.55 | 94.8 ± 28.8 | 79.4 ± 18 | 1021 ± 233 | 872.4 ± 167.9 | 509.4 ± 113.4 |
| HF | 2.94 ± 0.33 | 2.09 ± 0.19 | 1.42 ± 0.36 | 104.61 ± 8.76 | 93 ± 20.2 | 73.51 ± 6.2 | 828 ± 152.8 | 877.2 ± 72.5 | 628.4 ± 15.9 |
| CF | 2.21 ± 0.46 | 1.52 ± 0.19 | 1.04 ± 0.22 | 81.56 ± 9.56 | 72.26 ± 4.36 | 55.84 ± 5.91 | 917 ± 188 | 747.4 ± 99.8 | 419.7 ± 22.3 |
| BF | 1.78 ± 0.34 * | 1.36 ± 0.22 | 1.18 ± 0.42 | 82.66 ± 8.58 | 68.44 ± 3.11 | 55.76 ± 8.71 | 939.4 ± 32.1 | 483.1 ± 96.8 | 450.5 ± 39.6 |
| RF | 3.10 ± 0.38 | 2.24 ± 0.37 | 2.08 ± 0.32 ** | 111.8 ± 21.1 | 83.47 ± 12.62 | 82.03 ± 10.35 | 1001.3 ± 38.6 | 756.3 ± 221.4 | 499.6 ± 15.6 |
| Fractions | Root Projected Area (cm2) | Root Volume (cm3) | Average Diameter (mm) | ||||||
| 2 dS·m−1 | 4 dS·m−1 | 8 dS·m−1 | 2 dS·m−1 | 4 dS·m−1 | 8 dS·m−1 | 2 dS·m−1 | 4 dS·m−1 | 8 dS·m−1 | |
| Control | 18.6 ± 2.08 | 18.03 ± 5.01 | 10.73 ± 0.54 | 0.36 ± 0.02 | 0.34 ± 0.05 | 0.22 ± 0.01 | 0.25 ± 0.01 | 0.24 ± 0.01 | 0.25 ± 0.01 |
| ME | 25.41 ± 6.25 | 20.38 ± 3.88 | 12.47 ± 2.83 | 0.50 ± 0.08 | 0.37 ± 0.04 | 0.24 ± 0.03 | 0.25 ± 0.01 | 0.23 ± 0.01 | 0.24 ± 0.01 |
| HF | 19.6 ± 2.87 | 20.49 ± 1.58 | 15.79 ± 1.94 | 0.36 ± 0.03 | 0.38 ± 0.02 | 0.31 ± 0.04 | 0.24 ± 0.01 | 0.23 ± 0.01 | 0.25 ± 0.02 |
| CF | 21.05 ± 4.43 | 18.6 ± 2.08 | 10.714 ± 0.49 | 0.38 ± 0.05 | 0.36 ± 0.02 | 0.21 ± 0.01 | 0.23 ± 0.01 | 0.25 ± 0.01 | 0.26 ± 0.01 |
| BF | 22.29 ± 0.34 | 10.75 ± 2.18 | 10.70 ± 1.28 | 0.42 ± 0.01 | 0.19 ± 0.02 | 0.20 ± 0.02 | 0.24 ± 0.01 | 0.22 ± 0.01 | 0.24 ± 0.01 |
| RF | 25.18 ± 0.28 | 18.03 ± 5.01 | 11.56 ± 0.77 | 0.50 ± 0.01 | 0.34 ± 0.05 | 0.24 ± 0.02 | 0.25 ± 0.01 | 0.24 ± 0.01 | 0.28 ± 0.01 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
El Boukhari, M.E.M.; Barakate, M.; Choumani, N.; Bouhia, Y.; Lyamlouli, K. Ulva lactuca Extract and Fractions as Seed Priming Agents Mitigate Salinity Stress in Tomato Seedlings. Plants 2021, 10, 1104. https://0-doi-org.brum.beds.ac.uk/10.3390/plants10061104
El Boukhari MEM, Barakate M, Choumani N, Bouhia Y, Lyamlouli K. Ulva lactuca Extract and Fractions as Seed Priming Agents Mitigate Salinity Stress in Tomato Seedlings. Plants. 2021; 10(6):1104. https://0-doi-org.brum.beds.ac.uk/10.3390/plants10061104
Chicago/Turabian StyleEl Boukhari, Mohammed El Mehdi, Mustapha Barakate, Nadia Choumani, Youness Bouhia, and Karim Lyamlouli. 2021. "Ulva lactuca Extract and Fractions as Seed Priming Agents Mitigate Salinity Stress in Tomato Seedlings" Plants 10, no. 6: 1104. https://0-doi-org.brum.beds.ac.uk/10.3390/plants10061104