
Floral Scent Evolution in the Genus Jaborosa (Solanaceae): Influence of Ecological and Environmental Factors

 ,
,  ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
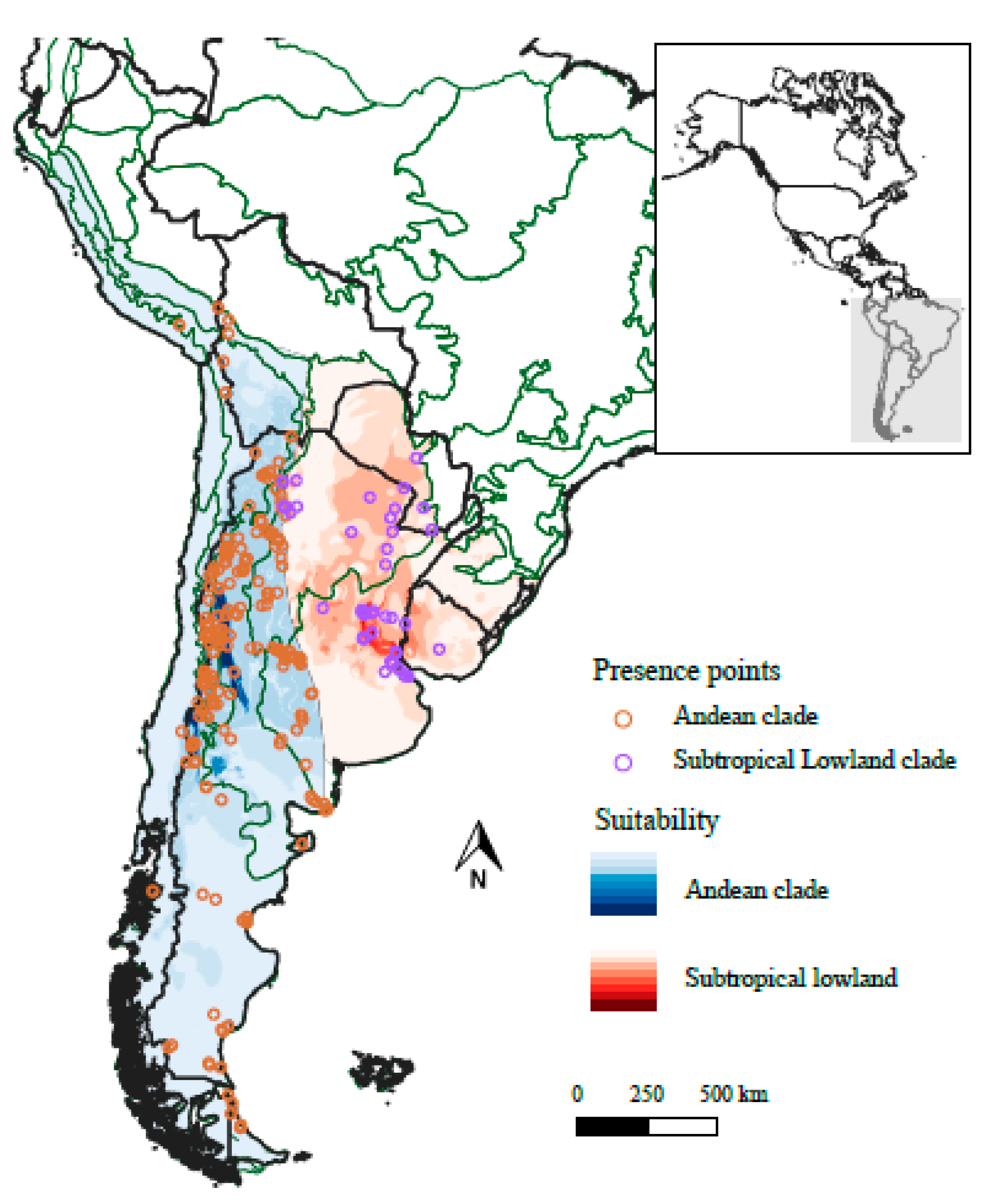
2.1. Ecological Niche Modelling of JABOROSA in Geographical Space
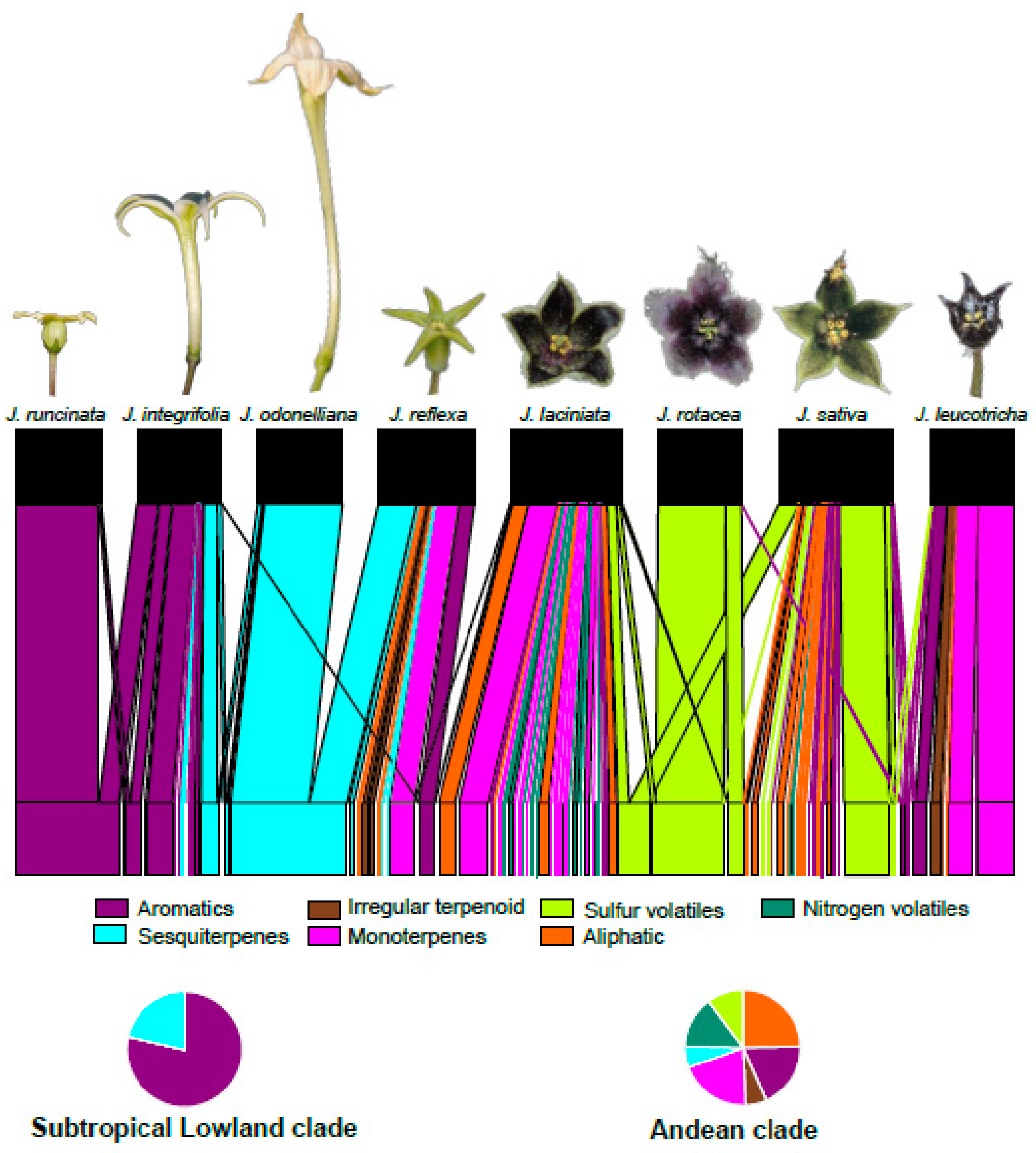
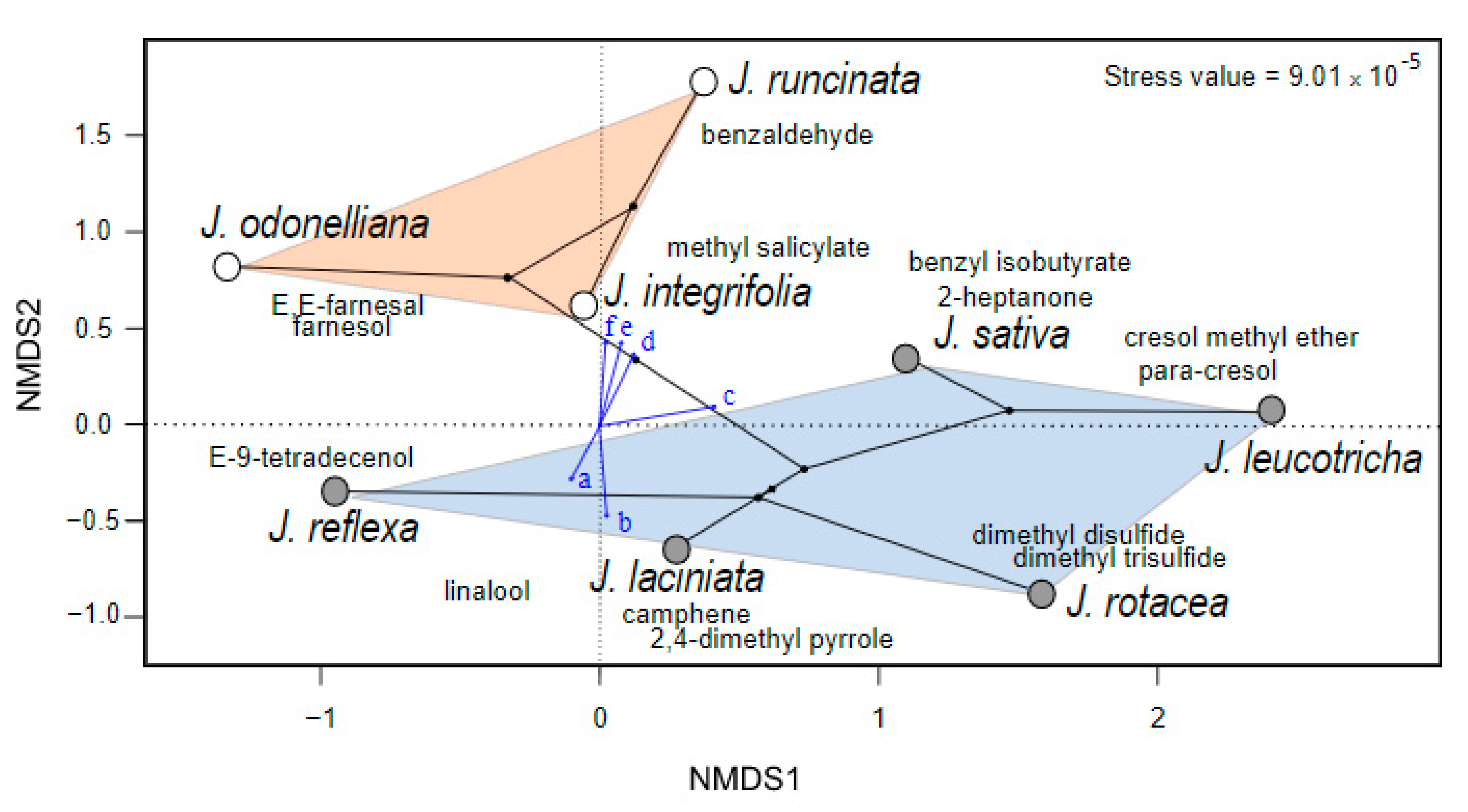
2.2. Composition and Variation of Floral Scent among Jaborosa Species
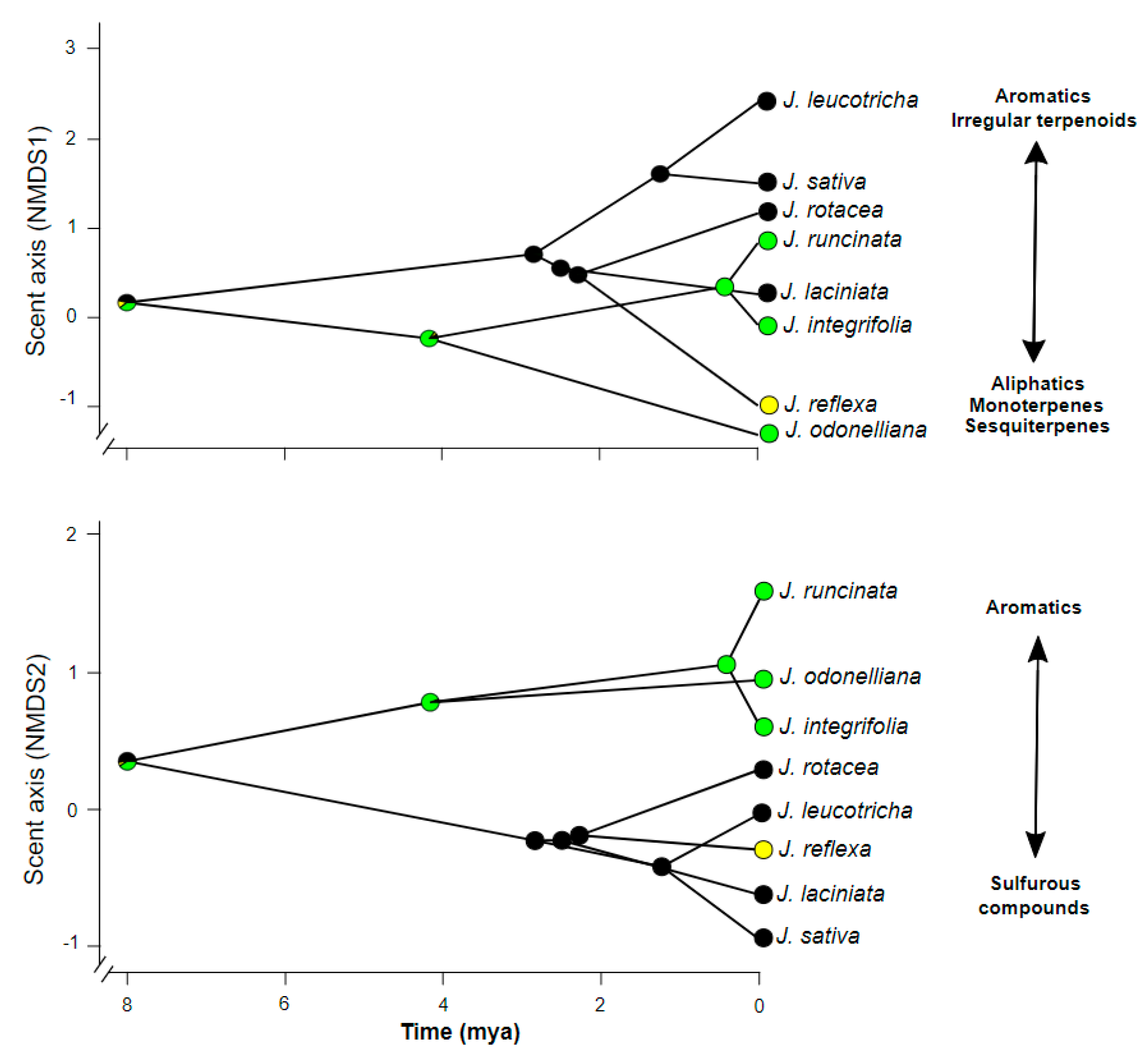
2.3. Evolution of Floral Scent in JABOROSA and Its Association with Pollination Mode and Climate
3. Discussion
3.1. Methodological Challenges
3.2. Clade-Specific Volatile Patterns
3.3. Niche Dimensions within Sapromyophily: A Closer Look at Scent in Fly-Pollinated Jaborosa Species
3.4. Jaborosa reflexa: Transition to Generalized Pollination in the Andes
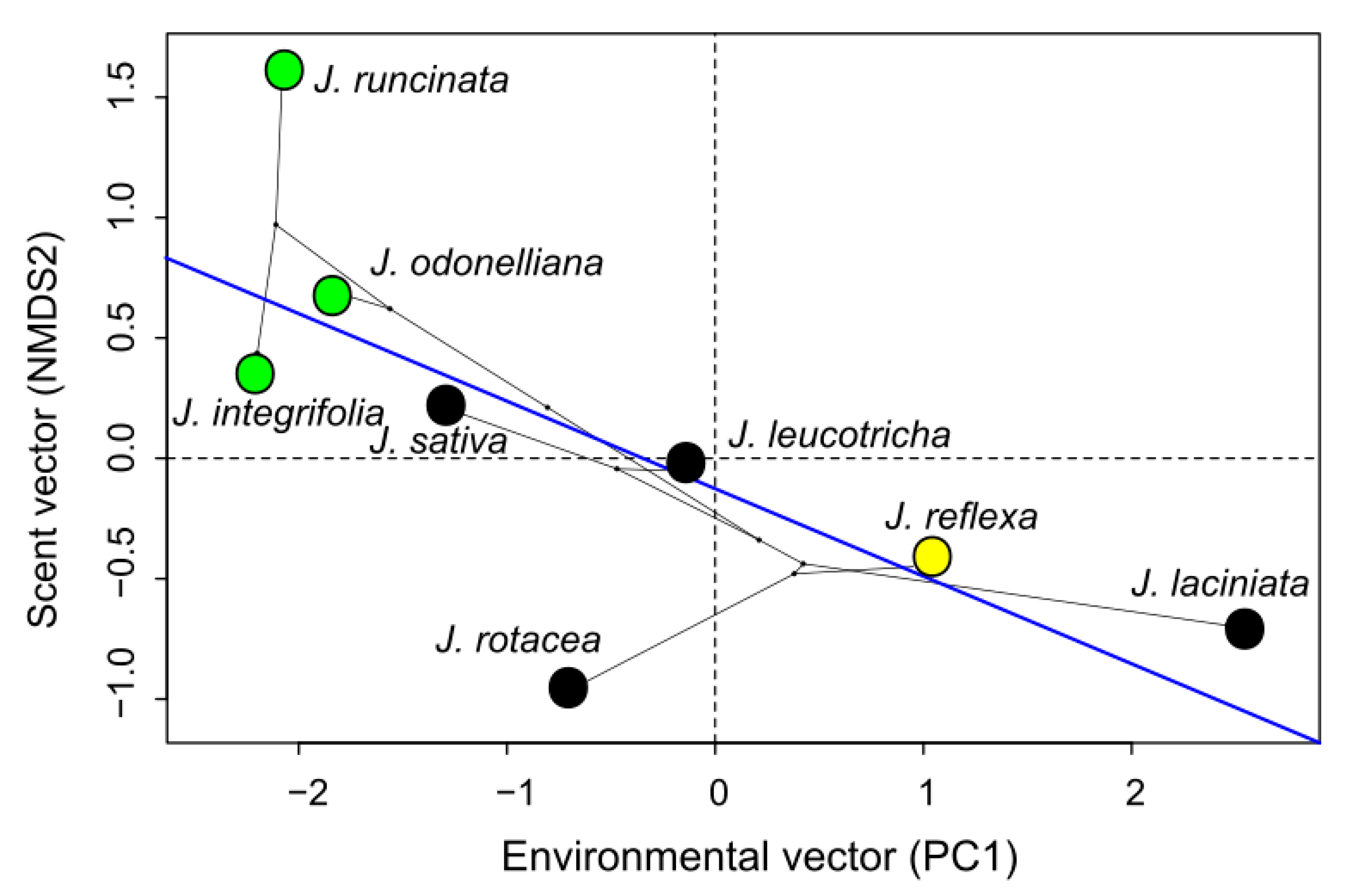
3.5. Environmental Influence on Floral VOCs and Pollinator Climate
4. Materials and Methods
4.1. Study System
4.2. Ecological Niche Modelling in Geographical Space
4.3. Floral Scent Collection and VOCs Identification
4.4. Floral Scent Variation among Jaborosa Species Associated with Pollination Mode and Climate
4.5. Evolution of Floral Scent across Environments
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Acknowledgments
Conflicts of Interest
References
- Knudsen, J.T.; Eriksson, R.; Gershenzon, J.; Ståhl, B. Diversity and distribution of floral scent. Bot. Rev. 2006, 72, 1. [Google Scholar] [CrossRef]
- Raguso, R.A. Wake up and smell the roses: The ecology and evolution of floral scent. Annu. Rev. Ecol. Evol. Syst. 2008, 39, 549–569. [Google Scholar] [CrossRef]
- Schiestl, F.P.; Johnson, S.D. Pollinator-mediated evolution of floral signals. Trends Ecol. Evol. 2013, 28, 307–315. [Google Scholar] [CrossRef]
- Leonard, A.S.; Dornhaus, A.; Papaj, D.R. Forget-me-not: Complex floral displays, inter-signal interactions, and pollinator cognition. Curr. Zool. 2011, 57, 215–224. [Google Scholar] [CrossRef] [Green Version]
- Junker, R.R.; Parachnowitsch, A.L. Working towards a holistic view on flower traits—How floral scents mediate plant–animal interactions in concert with other floral characters. J. Indian Inst. Sci. 2015, 95, 43–68. [Google Scholar]
- Raguso, R.A. Behavioral responses to floral scent: Experimental manipulations and multimodal plant-pollinator communication. In Biology of Plant Volatiles; Pichersky, E., Dudareva, N., Eds.; CRC Press: Boca Raton, FL, USA, 2020; pp. 297–325. [Google Scholar]
- Jürgens, A. The hidden language of flowering plants: Floral odours as a key for understanding angiosperm evolution? New Phytol. 2009, 183, 240–243. [Google Scholar] [CrossRef]
- Galen, C.; Kaczorowski, R.; Todd, S.L.; Geib, J.; Raguso, R.A. Dosage-dependent impacts of a floral volatile compound on pollinators, larcenists, and the potential for floral evolution in the alpine skypilot Polemonium viscosum. Am. Nat. 2011, 177, 258–272. [Google Scholar] [CrossRef]
- Kessler, D.; Diezel, C.; Clark, D.G.; Colquhoun, T.A.; Baldwin, I.T. Petunia flowers solve the defence/apparency dilemma of pollinator attraction by deploying complex floral blends. Ecol. Lett. 2013, 16, 299–306. [Google Scholar] [CrossRef] [PubMed]
- Wester, P.; Lunau, K. Plant–pollinator communication. Adv. Bot. Res. 2017, 82, 225–257. [Google Scholar]
- Wang, T.N.; Clifford, M.R.; Martínez-Gómez, J.; Johnson, J.C.; Riffell, J.A.; Di Stilio, V.S. Scent matters: Differential contribution of scent to insect response in flowers with insect vs. wind pollination traits. Ann. Bot. 2019, 123, 289–301. [Google Scholar] [CrossRef]
- Lahondère, C.; Vinauger, C.; Okubo, R.P.; Wolff, G.H.; Chan, J.K.; Akbari, O.S.; Riffell, J.A. The olfactory basis of orchid pollination by mosquitoes. Proc. Natl. Acad. Sci. USA 2020, 117, 708–716. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Raguso, R.A.; Levin, R.A.; Foose, S.E.; Holmberg, M.W.; McDade, L.A. Fragrance chemistry, nocturnal rhythms and pollination “syndromes” in Nicotiana. Phytochemistry 2003, 63, 265–284. [Google Scholar] [CrossRef]
- Schiestl, F.P.; Dötterl, S. The evolution of floral scent and olfactory preferences in pollinators: Coevolution or pre-existing bias? Evolution 2012, 66, 2042–2055. [Google Scholar] [CrossRef]
- Levin, R.A.; McDade, L.A.; Raguso, R.A. The systematic utility of floral and vegetative fragrance in two genera of Nyctaginaceae. Syst. Biol. 2003, 52, 334–351. [Google Scholar] [CrossRef] [PubMed]
- Peakall, R.; Ebert, D.; Poldy, J.; Barrow, R.A.; Francke, W.; Bower, C.C.; Schiestl, F.P. Pollinator specificity, floral odour chemistry and the phylogeny of Australian sexually deceptive Chiloglottis orchids: Implications for pollinator-driven speciation. New Phytol. 2010, 188, 437–450. [Google Scholar] [CrossRef] [PubMed]
- Dobson, H.E.; Arroyo, J.; Bergström, G.; Groth, I. Interspecific variation in floral fragrances within the genus Narcissus (Amaryllidaceae). Biochem. Syst. Ecol. 1997, 25, 685–706. [Google Scholar] [CrossRef]
- Williams, N.H.; Whitten, W.M. Molecular phylogeny and floral fragrances of male euglossine bee-pollinated orchids: A study of Stanhopea (Orchidaceae). Plant Species Biol. 1999, 14, 129–136. [Google Scholar] [CrossRef]
- Friberg, M.; Schwind, C.; Guimarães, P.R.; Raguso, R.A.; Thompson, J.N. Extreme diversification of floral volatiles within and among species of Lithophragma (Saxifragaceae). Proc. Natl. Acad. Sci. USA 2019, 116, 4406–4415. [Google Scholar] [CrossRef] [Green Version]
- Delle-Vedove, R.; Schatz, B.; Dufay, M. Understanding intraspecific variation of floral scent in light of evolutionary ecology. Ann. Bot. 2017, 120, 1–20. [Google Scholar] [CrossRef] [Green Version]
- Gervasi, D.D.; Schiestl, F.P. Real-time divergent evolution in plants driven by pollinators. Nat. Commun. 2017, 8, 14691. [Google Scholar] [CrossRef] [Green Version]
- Raguso, R.A.; Kelber, A.; Pfaff, M.; Levin, R.A.; McDade, L.A. Floral biology of North American Oenothera sect. Lavauxia (Onagraceae): Advertisements, rewards, and extreme variation in floral depth. Ann. Mo. Bot. Gard. 2007, 94, 236–257. [Google Scholar]
- Doubleday, L.A.; Raguso, R.A.; Eckert, C.G. Dramatic vestigialization of floral fragrance across a transition from outcrossing to selfing in Abronia umbellata (Nyctaginaceae). Am. J. Bot. 2013, 100, 2280–2292. [Google Scholar] [CrossRef]
- Theis, N.; Adler, L.S. Advertising to the enemy: Enhanced floral fragrance increases beetle attraction and reduces plant reproduction. Ecology 2012, 93, 430–435. [Google Scholar] [CrossRef] [PubMed]
- Ramos, S.E.; Schiestl, F.P. Rapid plant evolution driven by the interaction of pollination and herbivory. Science 2019, 364, 193–196. [Google Scholar] [CrossRef]
- Shuttleworth, A.; Johnson, S.D. The missing stink: Sulphur compounds can mediate a shift between fly and wasp pollination systems. Proc. R. Soc. B Biol. Sci. 2010, 277, 2811–2819. [Google Scholar] [CrossRef] [Green Version]
- Peng, F.; Byers, K.J.; Bradshaw, H.D., Jr. Less is more: Independent loss-of-function ocimene synthase alleles parallel pollination syndrome diversification in monkeyflowers (Mimulus). Am. J. Bot. 2017, 104, 1055–1059. [Google Scholar] [CrossRef] [Green Version]
- Johnson, S.D. The pollination niche and its role in the diversification and maintenance of the southern African flora. Philos. Trans. R. Soc. B Biol. Sci. 2010, 365, 499–516. [Google Scholar] [CrossRef] [Green Version]
- Kay, K.M.; Zepeda, A.M.; Raguso, R.A. Experimental sympatry reveals geographic variation in floral isolation by hawkmoths. Ann. Bot. 2019, 123, 405–413. [Google Scholar] [CrossRef]
- Grant, V. Origin of floral isolation between ornithophilous and sphingophilous plant species. Proc. Natl. Acad. Sci. USA 1993, 90, 7729–7733. [Google Scholar] [CrossRef] [Green Version]
- Streisfeld, M.A.; Kohn, J.R. Environment and pollinator-mediated selection on parapatric floral races of Mimulus aurantiacus. J. Evol. Biol. 2007, 20, 122–132. [Google Scholar] [CrossRef]
- Streisfeld, M.A.; Kohn, J.R. Contrasting patterns of floral and molecular variation across a cline in Mimulus aurantiacus. Evolution 2005, 59, 2548–2559. [Google Scholar] [CrossRef]
- Gross, K.; Sun, M.; Schiestl, F.P. Why do floral perfumes become different? Region-specific selection on floral scent in a terrestrial orchid. PLoS ONE 2016, 11, e0147975. [Google Scholar] [CrossRef] [PubMed]
- Gfrerer, E.; Laina, D.; Gibernau, M.; Fuchs, R.; Happ, M.; Tolasch, T.; Trutschnig, W.; Hörger, A.; Comes, H.P.; Dötterl, S. Floral Scents of a Deceptive Plant Are Hyperdiverse and under Population Specific Phenotypic Selection. Available online: https://www.biorxiv.org/content/10.1101/2021.04.28.441155v1 (accessed on 5 June 2021).
- Majetic, C.J.; Raguso, R.A.; Ashman, T.L. The sweet smell of success: Floral scent affects pollinator attraction and seed fitness in Hesperis matronalis. Funct. Ecol. 2009, 23, 480–487. [Google Scholar] [CrossRef]
- Farré-Armengol, G.; Peñuelas, J.; Li, T.; Yli-Pirilä, P.; Filella, I.; Llusia, J.; Blande, J.D. Ozone degrades floral scent and reduces pollinator attraction to flowers. New Phytol. 2016, 209, 152–160. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Campbell, D.R.; Sosenski, P.; Raguso, R.A. Phenotypic plasticity of floral volatiles in response to increasing drought stress. Ann. Bot. 2019, 123, 601–610. [Google Scholar] [CrossRef] [PubMed]
- Glenny, W.R.; Runyon, J.B.; Burkle, L.A. Drought and increased CO2 alter floral visual and olfactory traits with context-dependent effects on pollinator visitation. New Phytol. 2018, 220, 785–798. [Google Scholar] [CrossRef] [Green Version]
- Fenske, M.P.; Hazelton, K.D.H.; Hempton, A.K.; Shim, J.S.; Yamamoto, B.M.; Riffell, J.A.; Imaizumi, T. Circadian clock gene late elongated hypocotyl directly regulates the timing of floral scent emission in Petunia. Proc. Natl. Acad. Sci. USA 2015, 112, 9775–9780. [Google Scholar] [CrossRef] [Green Version]
- Cna’ani, A.; Dener, E.; Ben-Zeev, E.; Günther, J.; Köllner, T.G.; Tzin, V.; Seifan, M. Phylogeny and abiotic conditions shape the diel floral emission patterns of desert Brassicaceae species. Plant Cell Environ. 2021, 44, 2656–2671. [Google Scholar] [CrossRef]
- Fenske, M.P.; Nguyen, L.P.; Horn, E.K.; Riffell, J.A.; Imaizumi, T. Circadian clocks of both plants and pollinators influence flower seeking behavior of the pollinator hawkmoth Manduca sexta. Sci. Rep. 2018, 8, 2842. [Google Scholar] [CrossRef]
- Grant, K.A.; Grant, V. Flower Pollination in the Phlox Family; Columbia University Press: New York, NY, USA, 1965. [Google Scholar]
- Cruden, R.W.; Kinsman, S.; Stockhouse, R.E.; Linhart, Y.B. Pollination, fecundity, and the distribution of moth-flowered plants. Biotropica 1976, 8, 204–210. [Google Scholar] [CrossRef]
- Chapurlat, E.; Anderson, J.; Ågren, J.; Friberg, M.; Sletvold, N. Diel pattern of floral scent emission matches the relative importance of diurnal and nocturnal pollinators in populations of Gymnadenia conopsea. Ann. Bot. 2018, 121, 711–721. [Google Scholar] [CrossRef]
- Braunschmid, H.; Guilhot, R.; Dötterl, S. Floral scent and pollinators of Cypripedium calceolus L. at different latitudes. Diversity 2021, 13, 5. [Google Scholar] [CrossRef]
- Hazzi, N.A.; Moreno, J.S.; Ortiz-Movliav, C.; Palacio, R.D. Biogeographic regions and events of isolation and diversification of the endemic biota of the tropical Andes. Proc. Natl. Acad. Sci. USA 2018, 115, 7985–7990. [Google Scholar] [CrossRef] [Green Version]
- Nevado, B.; Contreras-Ortiz, N.; Hughes, C.; Filatov, D.A. Pleistocene glacial cycles drive isolation, gene flow and speciation in the high-elevation Andes. New Phytol. 2018, 219, 779–793. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pérez, F.; Arroyo, M.T.; Medel, R.; Hershkovitz, M.A. Ancestral reconstruction of flower morphology and pollination systems in Schizanthus (Solanaceae). Am. J. Bot. 2006, 93, 1029–1038. [Google Scholar] [CrossRef]
- Schlumpberger, B.O.; Raguso, R.A. Geographic variation in floral scent of Echinopsis ancistrophora (Cactaceae); evidence for constraints on hawkmoth attraction. Oikos 2008, 117, 801–814. [Google Scholar] [CrossRef]
- Strelin, M.M.; Arroyo, J.I.; Fliesswasser, S.; Ackermann, M. Diversification of Caiophora (Loasaceae subfam. Loasoideae) during the uplift of the Central Andes. Org. Divers. Evol. 2017, 17, 29–41. [Google Scholar] [CrossRef]
- Ibañez, A.C.; Moré, M.; Salazar, G.; Leiva, S.; Barboza, G.E.; Cocucci, A.A. Crescendo, diminuendo and subito of the trumpets: Winds of change in the concerted evolution between flowers and pollinators in Salpichroa (Solanaceae). Mol. Phylogenetics Evol. 2019, 132, 90–99. [Google Scholar] [CrossRef]
- Givnish, T.J. Adaptive radiation and molecular systematics: Issues and approaches. In Molecular Evolution and Adaptive Radiation; Givnish, T.J., Sytsma, T.J., Eds.; Cambridge University Press: Cambridge, UK, 1997; pp. 1–54. [Google Scholar]
- Schluter, D. The Ecology of Adaptive Radiation; OUP Oxford: Oxford, UK, 2000. [Google Scholar]
- Rundle, H.D.; Nosil, P. Ecological speciation. Ecol. Lett. 2005, 8, 336–352. [Google Scholar] [CrossRef]
- Moré, M.; Cocucci, A.A.; Sérsic, A.N.; Barboza, G.E. Phylogeny and floral trait evolution in Jaborosa (Solanaceae). Taxon 2015, 64, 523–534. [Google Scholar] [CrossRef]
- Moré, M.; Ibañez, A.C.; Drewniak, M.E.; Cocucci, A.A.; Raguso, R.A. Flower diversification across “pollinator climates”: Sensory aspects of corolla color evolution in the florally diverse South American genus Jaborosa (Solanaceae). Front. Plant Sci. 2020, 11, 601975. [Google Scholar] [CrossRef]
- Moré, M.; Cocucci, A.A.; Raguso, R.A. The importance of oligosulfides in the attraction of fly pollinators to the brood-site deceptive species Jaborosa rotacea (Solanaceae). Int. J. Plant Sci. 2013, 174, 863–876. [Google Scholar] [CrossRef] [Green Version]
- Moré, M.; Mulieri, P.; Battán-Horenstein, M.; Cocucci, A.A.; Raguso, R.A. The role of fetid olfactory signals in the shift to saprophilous fly pollination in Jaborosa (Solanaceae). Arthropod-Plant Interact. 2019, 13, 375–386. [Google Scholar] [CrossRef]
- Zito, P.; Dötterl, S.; Sajeva, M. Floral volatiles in a sapromyiophilous plant and their importance in attracting house fly pollinators. J. Chem. Ecol. 2015, 41, 340–349. [Google Scholar] [CrossRef] [PubMed]
- Chen, G.; Ma, X.K.; Jürgens, A.; Lu, J.; Liu, E.X.; Sun, W.B.; Cai, X.H. Mimicking livor mortis: A well-known but unsubstantiated color profile in sapromyiophily. J. Chem. Ecol. 2015, 41, 808–815. [Google Scholar] [CrossRef]
- Dobson, H.E.M. Relationship between floral fragrance composition and type of pollinator. In Biology of Floral Scent; Dudareva, N., Pichersky, E., Eds.; CRC Press: Boca Raton, FL, USA, 2006; pp. 147–198. [Google Scholar]
- Jürgens, A.; Dötterl, S.; Meve, U. The chemical nature of fetid floral odours in stapeliads (Apocynaceae-Asclepiadoideae-Ceropegieae). New Phytol. 2006, 172, 452–468. [Google Scholar] [CrossRef] [PubMed]
- Jürgens, A.; Wee, S.L.; Shuttleworth, A.; Johnson, S.D. Chemical mimicry of insect oviposition sites: A global analysis of convergence in angiosperms. Ecol. Lett. 2013, 16, 1157–1167. [Google Scholar] [CrossRef] [PubMed]
- Urru, I.; Stensmyr, M.C.; Hansson, B.S. Pollination by brood-site deception. Phytochemistry 2011, 72, 1655–1666. [Google Scholar] [CrossRef] [PubMed]
- Kaiser, R. The Scent of Orchids: Olfactory and Chemical Investigations; Elsevier Science Publishers: Amsterdam, The Netherlands, 1993. [Google Scholar]
- Raguso, R.A.; Willis, M.A. Synergy between visual and olfactory cues in nectar feeding by wild hawkmoths, Manduca sexta. Anim. Behav. 2005, 69, 407–418. [Google Scholar] [CrossRef]
- Knudsen, J.T.; Tollsten, L. Trends in floral scent chemistry in pollination syndromes: Floral scent composition in moth-pollinated taxa. Bot. J. Linn. Soc. 1993, 113, 263–284. [Google Scholar] [CrossRef]
- Raguso, R.A.; Light, D.M.; Pickersky, E. Electroantennogram responses of Hyles lineata (Sphingidae: Lepidoptera) to volatile compounds from Clarkia breweri (Onagraceae) and other moth-pollinated flowers. J. Chem. Ecol. 1996, 22, 1735–1766. [Google Scholar] [CrossRef] [PubMed]
- Fraser, A.M.; Mechaber, W.L.; Hildebrand, J.G. Electroantennographic and behavioral responses of the sphinx moth Manduca sexta to host plant headspace volatiles. J. Chem. Ecol. 2003, 29, 1813–1833. [Google Scholar] [CrossRef] [PubMed]
- Balao, F.; Herrera, J.; Talavera, S.; Dötterl, S. Spatial and temporal patterns of floral scent emission in Dianthus inoxianus and electroantennographic responses of its hawkmoth pollinator. Phytochemistry 2011, 72, 601–609. [Google Scholar] [CrossRef]
- Hoballah, M.E.; Stuurman, J.; Turlings, T.C.; Guerin, P.M.; Connetable, S.; Kuhlemeier, C. The composition and timing of flower odour emission by wild Petunia axillaris coincide with the antennal perception and nocturnal activity of the pollinator Manduca sexta. Planta 2005, 222, 141–150. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Riffell, J.A.; Lei, H.; Hildebrand, J.G. Neural correlates of behavior in the moth Manduca sexta in response to complex odors. Proc. Natl. Acad. Sci. USA 2009, 106, 19219–19226. [Google Scholar] [CrossRef] [Green Version]
- Löwenberg-Neto, P. Neotropical region: A shapefile of Morrone’s (2014) biogeographical regionalization. Zootaxa 2014, 3802, 300. [Google Scholar] [CrossRef]
- Runfola, D.; Anderson, A.; Baier, H.; Crittenden, M.; Dowker, E.; Fuhrig, S. geoBoundaries: A global database of political administrative boundaries. PLoS ONE 2020, 15, e0231866. [Google Scholar] [CrossRef] [Green Version]
- Ollerton, J.; Raguso, R.A. The sweet stench of decay. New Phytol. 2006, 172, 382–385. [Google Scholar] [CrossRef]
- Stensmyr, M.C.; Urru, I.; Collu, I.; Celander, M.; Hansson, B.S.; Angioy, A.M. Rotting smell of dead-horse arum florets. Nature 2002, 420, 625–626. [Google Scholar] [CrossRef]
- Johnson, S.D.; Jürgens, A. Convergent evolution of carrion and faecal scent mimicry in fly-pollinated angiosperm flowers and a stinkhorn fungus. S. Afr. J. Bot. 2010, 76, 796–807. [Google Scholar] [CrossRef] [Green Version]
- Zito, P.; Sajeva, M.; Raspi, A.; Dötterl, S. Dimethyl disulfide and dimethyl trisulfide: So similar yet so different in evoking biological responses in saprophilous flies. Chemoecology 2014, 24, 261–267. [Google Scholar] [CrossRef]
- Goodrich, K.R.; Zjhra, M.L.; Ley, C.A.; Raguso, R.A. When flowers smell fermented: The chemistry and ontogeny of yeasty floral scent in pawpaw (Asimina triloba: Annonaceae). Int. J. Plant Sci. 2006, 167, 33–46. [Google Scholar] [CrossRef] [Green Version]
- Riffell, J.A.; Lei, H.; Abrell, L.; Hildebrand, J.G. Neural basis of a pollinator’s buffet: Olfactory specialization and learning in Manduca sexta. Science 2013, 339, 200–204. [Google Scholar] [CrossRef] [Green Version]
- Rubini Pisano, A.; Moré, M.; Cisternas, M.A.; Raguso, R.A.; Benitez-Vieyra, S. Breakdown of species boundaries in Mandevilla: Floral morphological intermediacy, novel fragrances and asymmetric pollen flow. Plant Biol. 2019, 21, 206–215. [Google Scholar] [CrossRef]
- Levin, R.A.; Raguso, R.A.; McDade, L.A. Fragrance chemistry and pollinator affinities in Nyctaginaceae. Phytochemistry 2001, 58, 429–440. [Google Scholar] [CrossRef] [Green Version]
- Sazatornil, F.D.; Moré, M.; Benitez-Vieyra, S.; Cocucci, A.A.; Kitching, I.J.; Schlumpberger, B.O.; Oliveira, P.E.; Sazima, M.; Amorim, F.W. Beyond neutral and forbidden links: Morphological matches and the assembly of mutualistic hawkmoth–plant networks. J. Anim. Ecol. 2016, 85, 1586–1594. [Google Scholar] [CrossRef]
- Raguso, R.A.; Willis, M.A. Synergy between visual and olfactory cues in nectar feeding by naıve hawkmoths, Manduca sexta. Anim. Behav. 2002, 64, 685–695. [Google Scholar] [CrossRef] [Green Version]
- Klahre, U.; Gurba, A.; Hermann, K.; Saxenhofer, M.; Bossolini, E.; Guerin, P.; Kuhlemeier, C. Pollinator choice in Petunia depends on two major genetic loci for floral scent production. Curr. Biol. 2011, 21, 730–739. [Google Scholar] [CrossRef] [Green Version]
- Haverkamp, A.; Bing, J.; Badeke, E.; Hansson, B.S.; Knaden, M. Innate olfactory preferences for flowers matching proboscis length ensure optimal energy gain in a hawkmoth. Nat. Commun. 2016, 7, 11644. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Daly, K.C.; Smith, B.H. Associative olfactory learning in the moth Manduca Sexta. J. Exp. Biol. 2000, 203, 2025–2038. [Google Scholar] [CrossRef]
- Riffell, J.A.; Alarcón, R.; Abrell, L.; Davidowitz, G.; Bronstein, J.L.; Hildebrand, J.G. Behavioral consequences of innate preferences and olfactory learning in hawkmoth–flower interactions. Proc. Natl. Acad. Sci. USA 2008, 105, 3404–3409. [Google Scholar] [CrossRef] [Green Version]
- Marino, P.; Raguso, R.; Goffinet, B. The ecology and evolution of fly dispersed dung mosses (Family Splachnaceae): Manipulating insect behaviour through odour and visual cues. Symbiosis 2009, 47, 61–76. [Google Scholar] [CrossRef]
- van der Niet, T.; Hansen, D.M.; Johnson, S.D. Carrion mimicry in a South African orchid: Flowers attract a narrow subset of the fly assemblage on animal carcasses. Ann. Bot. 2011, 107, 981–992. [Google Scholar] [CrossRef] [Green Version]
- Shuttleworth, A.; Johnson, S.D.; Jürgens, A. Entering through the narrow gate: A morphological filter explains specialized pollination of a carrion-scented stapeliad. Flora 2017, 232, 92–103. [Google Scholar] [CrossRef]
- Johnson, S.D.; Sivechurran, J.; Doarsamy, S.; Shuttleworth, A. Dung mimicry: The function of volatile emissions and corolla patterning in fly-pollinated Wurmbea flowers. New Phytol. 2020, 228, 1662–1673. [Google Scholar] [CrossRef]
- Wee, S.L.; Tan, S.B.; Jürgens, A. Pollinator specialization in the enigmatic Rafflesia cantleyi: A true carrion flower with species-specific and sex-biased blow fly pollinators. Phytochemistry 2018, 153, 120–128. [Google Scholar] [CrossRef]
- Lamberet, G.; Auberger, B.; Bergere, J.L. Aptitude of cheese bacteria for volatile S-methyl thioester synthesis I. Effect of substrates and pH on their formation by Brevibacterium linens GC171. Appl. Microbiol. Biotechnol. 1997, 47, 279–283. [Google Scholar] [CrossRef]
- Kaiser, R.; Tollsten, L. An introduction to the scent of cacti. Flavour Fragr. J. 1995, 10, 153–164. [Google Scholar] [CrossRef]
- Zito, P.; Guarino, S.; Peri, E.; Sajeva, M.; Colazza, S. Electrophysiological and behavioural responses of the housefly to “sweet” volatiles of the flowers of Caralluma europaea (Guss.) NE Br. Arthropod-Plant Interact. 2013, 7, 485–489. [Google Scholar] [CrossRef]
- Marotz-Clausen, G.; Jürschik, S.; Fuchs, R.; Schäffler, I.; Sulzer, P.; Gibernau, M.; Dötterl, S. Incomplete synchrony of inflorescence scent and temperature patterns in Arum maculatum L.(Araceae). Phytochemistry 2018, 154, 77–84. [Google Scholar] [CrossRef] [PubMed]
- Lukas, K.; Harig, T.; Schulz, S.; Hadersdorfer, J.; Dötterl, S. Flowers of European pear release common and uncommon volatiles that can be detected by honey bee pollinators. Chemoecology 2019, 29, 211–223. [Google Scholar] [CrossRef] [Green Version]
- Kite, G.C.; Hetterscheid, W.L.; Lewis, M.J.; Boyce, P.C.; Ollerton, J.; Cocklin, E.; Díaz, A.; Simmonds, M.S.J. Inflorescence odours and pollinators of Arum and Amorphophallus (Araceae). In Reproductive Biology; Owens, S.J., Rudall, P.J., Eds.; Royal Botanic Gardens Kew: Richmond, UK, 1998; pp. 295–315. [Google Scholar]
- McCuaig, B.; Dufour, S.C.; Raguso, R.A.; Bhatt, A.P.; Marino, P. Structural changes in plastids of developing Splachnum ampullaceum sporophytes and relationship to odour production. Plant Biol. 2015, 17, 466–473. [Google Scholar] [CrossRef] [Green Version]
- Parreira, L.A.; Azevedo, A.F.; Menini, L.; Gusevskaya, E.V. Functionalization of the naturally occurring linalool and nerol by the palladium catalyzed oxidation of their trisubstituted olefinic bonds. J. Mol. Catal. A Chem. 2017, 426, 429–434. [Google Scholar] [CrossRef]
- Burkle, L.A.; Runyon, J.B. Drought and leaf herbivory influence floral volatiles and pollinator attraction. Glob. Chang. Biol. 2016, 22, 1644–1654. [Google Scholar] [CrossRef] [PubMed]
- Copolovici, L.; Niinements, Ü. Environmental impacts on plant volatile emission. In Deciphering Chemical Language of Plant Communication; Blande, J.D., Glinwood, R., Eds.; Springer: Cham, Switzerland, 2016. [Google Scholar]
- Junker, R.R.; Blüthgen, N. Floral scents repel facultative flower visitors, but attract obligate ones. Ann. Bot. 2010, 105, 777–782. [Google Scholar] [CrossRef] [Green Version]
- Farré-Armengol, G.; Filella, I.; Llusià, J.; Niinemets, Ü.; Peñuelas, J. Changes in floral bouquets from compound-specific responses to increasing temperatures. Glob. Chang. Biol. 2014, 20, 3660–3669. [Google Scholar] [CrossRef] [Green Version]
- Terry, L.I.; Roemer, R.B.; Booth, D.T.; Moore, C.J.; Walter, G.H. Thermogenic respiratory processes drive the exponential increase of volatile organic compound emissions in Macrozamia cycad cones. Plant Cell Environ. 2016, 39, 1588–1600. [Google Scholar] [CrossRef]
- Hodkinson, I.D. Terrestrial insects along elevation gradients: Species and community responses to altitude. Biol. Rev. 2005, 80, 489–513. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Seymour, R.S.; Gibernau, M.; Ito, K. Thermogenesis and respiration of inflorescences of the dead horse arum Helicodiceros muscivorus, a pseudo-thermoregulatory aroid associated with fly pollination. Funct. Ecol. 2003, 17, 886–894. [Google Scholar] [CrossRef]
- Moré, M.; Kitching, I.J.; Cocucci, A.A. Sphingidae. In Biodiversidad de Artrópodos Argentinos Volumen 4; Roig-Juñent, S., Claps, L.E., Morrone, J.J., Eds.; INSUE-UNT Ediciones: San Miguel de Tucumán, Argentina, 2014; pp. 281–295. [Google Scholar]
- Haber, W.A.; Frankie, G.W. A tropical hawkmoth community: Costa Rican dry forest Sphingidae. Biotropica 1989, 21, 155–172. [Google Scholar] [CrossRef]
- Arroyo, M.T.K.; Primack, R.; Armesto, J. Community studies in pollination ecology in the high temperate Andes of central Chile. I. Pollination mechanisms and altitudinal variation. Am. J. Bot. 1982, 69, 82–97. [Google Scholar] [CrossRef]
- Pérez, F.; Arroyo, M.T.K.; Armesto, J.J. Evolution of autonomous selfing accompanies increased specialization in the pollination system of Schizanthus (Solanaceae). Am. J. Bot. 2009, 96, 1168–1176. [Google Scholar] [CrossRef]
- Mulieri, P.R.; Mariluis, J.C. New species and new records of Microcerella Macquart (Diptera: Sarcophagidae) belonging to the M. spinigena species-group. Neotrop. Entomol. 2009, 38, 101–103. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Revell, L.J. phytools: An R package for phylogenetic comparative biology (and other things). Methods Ecol. Evol. 2012, 3, 217–223. [Google Scholar] [CrossRef]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2020. [Google Scholar]
- QGIS Development Team. QGIS Geographic Information System. 3.10.4-A Coruña. Open Source Geospatial Foundation Project. 2019. Available online: http://www.qgis.org/ (accessed on 20 October 2020).
- Barve, N.; Barve, V.; Jiménez-Valverde, A.; Lira-Noriega, A.; Maher, S.P.; Peterson, A.T.; Soberón, J.; Villalobos, F. The crucial role of the accessible area in ecological niche modeling and species distribution modeling. Ecol. Model. 2011, 222, 1810–1819. [Google Scholar] [CrossRef]
- Peterson, A.T.; Soberón, J.; Sánchez-Cordero, V. Conservatism of ecological niches in evolutionary time. Science 1999, 285, 1265–1267. [Google Scholar] [CrossRef] [PubMed]
- Vega, G.C.; Pertierra, L.R.; Olalla-Tárraga, M.A. MERRAclim, a high-resolution global dataset of remotely sensed bioclimatic variables for ecological modelling. Sci. Data 2017, 4, 170078. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Busby, J.R. BIOCLIM. A bioclimate analysis and prediction system. In Nature Conservation: Cost Effective Biological Surveys and Data Analysis; Margules, C.R., Austin, M.P., Eds.; CSIRO: Canberra, Australia, 1991; pp. 64–68. [Google Scholar]
- Carpenter, G.; Gillison, A.N.; Winter, J. Domain: A flexible modelling procedure for mapping potential distributions of plants and animals. Biodivers. Conserv. 1993, 2, 667–680. [Google Scholar] [CrossRef]
- Phillips, S.J.; Anderson, R.P.; Schapire, R.E. Maximum entropy modeling of species geographic distributions. Ecol. Model. 2006, 190, 231–259. [Google Scholar] [CrossRef] [Green Version]
- Hijmans, R.J.; Phillips, S.; Leathwick, J.; Elith, J. Package ‘dismo’. Species Distribution Modeling. Version 1.1-4. 2017. Available online: https://cran.r-project.org/package=dismo (accessed on 20 October 2020).
- Guisan, A.; Edwards, T.C., Jr.; Hastie, T. Generalized linear and generalized additive models in studies of species distributions: Setting the scene. Ecol. Model. 2002, 157, 89–100. [Google Scholar] [CrossRef] [Green Version]
- Breiman, L. Random Forests. Mach. Learn. 2001, 45, 5–32. [Google Scholar] [CrossRef] [Green Version]
- Liaw, A.; Wiener, M. Classification and regression by random forest. R News 2002, 2, 18–22. [Google Scholar]
- Vapnik, V. Statistical Learning Theory; Wiley: New York, NY, USA, 1998. [Google Scholar]
- Karatzoglou, A.; Smola, A.; Hornik, K.; Zeileis, A. Kernlab—An S4 Package for Kernel Methods in R. J. Stat. Softw. 2004, 11, 1–20. [Google Scholar] [CrossRef] [Green Version]
- Muscarella, R.; Galante, P.J.; Soley-Guardia, M.; Boria, R.A.; Kass, J.M.; Uriarte, M.; Anderson, R.P. Package ENMeval. Automated Runs and Evaluations of Ecological Niche Models. Version 0.2.2. 2017. Available online: https://cran.r-project.org/package=ENMeval (accessed on 20 October 2020).
- Osorio-Olvera, L.; Lira-Noriega, A.; Soberón, J.; Townsend Peterson, A.; Falconi, M.; Contreras-Díaz, R.G.; Martínez-Meyer, E.; Barve, V.; Barve, N. ntbox: An R package with graphical user interface for modeling and evaluating multidimensional ecological niches. Methods Ecol. Evol. 2020, 11, 1199–1206. [Google Scholar] [CrossRef]
- Raguso, R.A.; Schlumpberger, B.O.; Kaczorowski, R.L.; Holtsford, T.P. Phylogenetic fragrance patterns in Nicotiana sections Alatae and Suaveolentes. Phytochemistry 2006, 67, 1931–1942. [Google Scholar] [CrossRef] [PubMed]
- Dötterl, S.; Wolfe, L.M.; Jürgens, A. Qualitative and quantitative analyses of flower scent in Silene latifolia. Phytochemistry 2005, 66, 203–213. [Google Scholar] [CrossRef]
- Dobson, H.E.M.; Raguso, R.A.; Knudsen, J.T.; Ayasse, M. Scent as an attractant. In Practical Pollination Biology; Dafni, A., Kevan, P.G., Husband, B.C., Eds.; Cambridge Enviroquest Ltd.: Cambridge, ON, Canada, 2005; pp. 197–230. [Google Scholar]
- Goodrich, K.R.; Raguso, R.A. The olfactory component of floral display in Asimina and Deeringothamnus (Annonaceae). New Phytol. 2009, 183, 457–469. [Google Scholar] [CrossRef] [PubMed]
- Kantsa, A.; Raguso, R.A.; Dyer, A.G.; Sgardelis, S.P.; Olesen, J.M.; Petanidou, T. Community-wide integration of floral colour and scent in a Mediterranean scrubland. Nat. Ecol. Evol. 2017, 1, 1502–1510. [Google Scholar] [CrossRef]
- Dormann, C.F. Using bipartite to describe and plot two-mode networks in R. R Package Version 2020, 4, 1–28. [Google Scholar]
- Oksanen, J.; Blanchet, F.G.; Kindt, R.; Legendre, P.; Minchin, P.R.; O’hara, R.B.; Oksanen, M.J. Package ‘vegan’. Community Ecol. Package Version 2013, 2, 1–295. [Google Scholar]
- Dötterl, S.; Jürgens, A. Spatial fragrance patterns in flowers of Silene latifolia: Lilac compounds as olfactory nectar guides? Plant Syst. Evol. 2005, 255, 99–109. [Google Scholar] [CrossRef]
- Felsenstein, J. Maximum-likelihood estimation of evolutionary trees from continuous characters. Am. J. Hum. Genet. 1973, 25, 471. [Google Scholar] [PubMed]
- Martins, E.P.; Hansen, T.F. Phylogenies and the comparative method: A general approach to incorporating phylogenetic information into the analysis of interspecific data. Am. Nat. 1997, 149, 646–667. [Google Scholar] [CrossRef] [Green Version]
- Pinheiro, J.; Bates, D.; DebRoy, S.; Sarkar, D.; Team, R.C. nlme: Linear and nonlinear mixed effects models. R Package Version 2006, 3, 109. [Google Scholar]





Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Moré, M.; Soteras, F.; Ibañez, A.C.; Dötterl, S.; Cocucci, A.A.; Raguso, R.A. Floral Scent Evolution in the Genus Jaborosa (Solanaceae): Influence of Ecological and Environmental Factors. Plants 2021, 10, 1512. https://0-doi-org.brum.beds.ac.uk/10.3390/plants10081512
Moré M, Soteras F, Ibañez AC, Dötterl S, Cocucci AA, Raguso RA. Floral Scent Evolution in the Genus Jaborosa (Solanaceae): Influence of Ecological and Environmental Factors. Plants. 2021; 10(8):1512. https://0-doi-org.brum.beds.ac.uk/10.3390/plants10081512
Chicago/Turabian StyleMoré, Marcela, Florencia Soteras, Ana C. Ibañez, Stefan Dötterl, Andrea A. Cocucci, and Robert A. Raguso. 2021. "Floral Scent Evolution in the Genus Jaborosa (Solanaceae): Influence of Ecological and Environmental Factors" Plants 10, no. 8: 1512. https://0-doi-org.brum.beds.ac.uk/10.3390/plants10081512