
Combining Ability and Heterotic Patterns of Tropical Early-Maturing Maize Inbred Lines under Individual and Combined Heat and Drought Environments

 ,
,  , , ,
, , ,  and
and 
Abstract
:1. Introduction
- Assess the importance of general combing ability (GCA) and specific combining ability (SCA) for grain yield and other key agronomic traits in a set of newly developed inbred lines under independent and combined drought and heat stress conditions;
- Identify inbred and hybrid testers across test environments;
- Assign the inbred lines into heterotic groups based on the GCA of multiple traits (HGCAMT) method and DArTseq markers, and assessed the performance and stability of the hybrids under the test environments.
2. Materials and Methods
2.1. Genetic Materials and Crossing Scheme
2.2. Field Evaluation and Agronomic Management
2.3. Traits Measured
2.4. DNA Extraction and DArTseq Genotyping
2.5. Statistical Analysis
3. Results
3.1. Genotypic Variation and General and Specific Combining Ability Analyses
3.2. Effects of General Combining Ability for Agronomic Traits
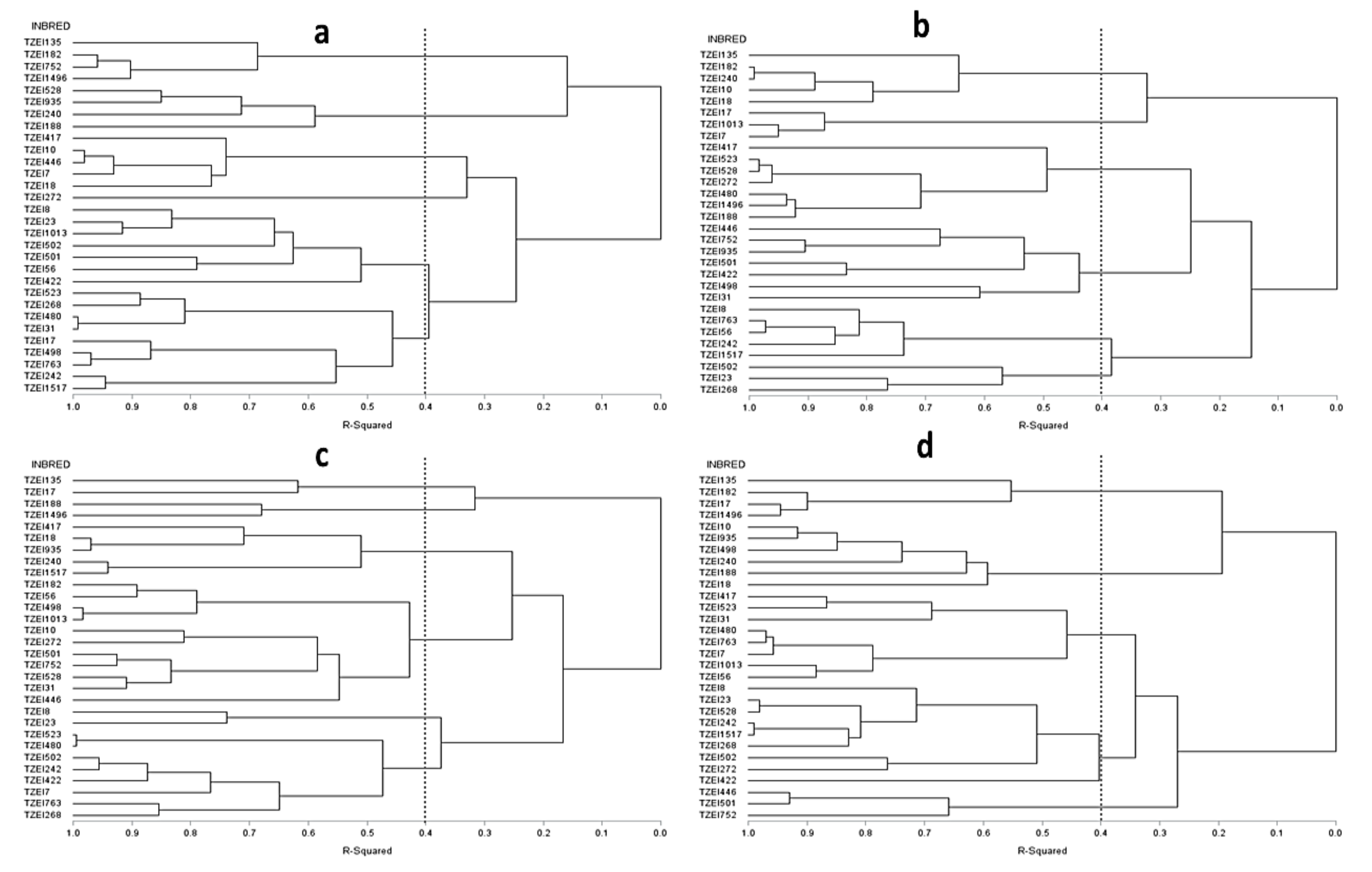
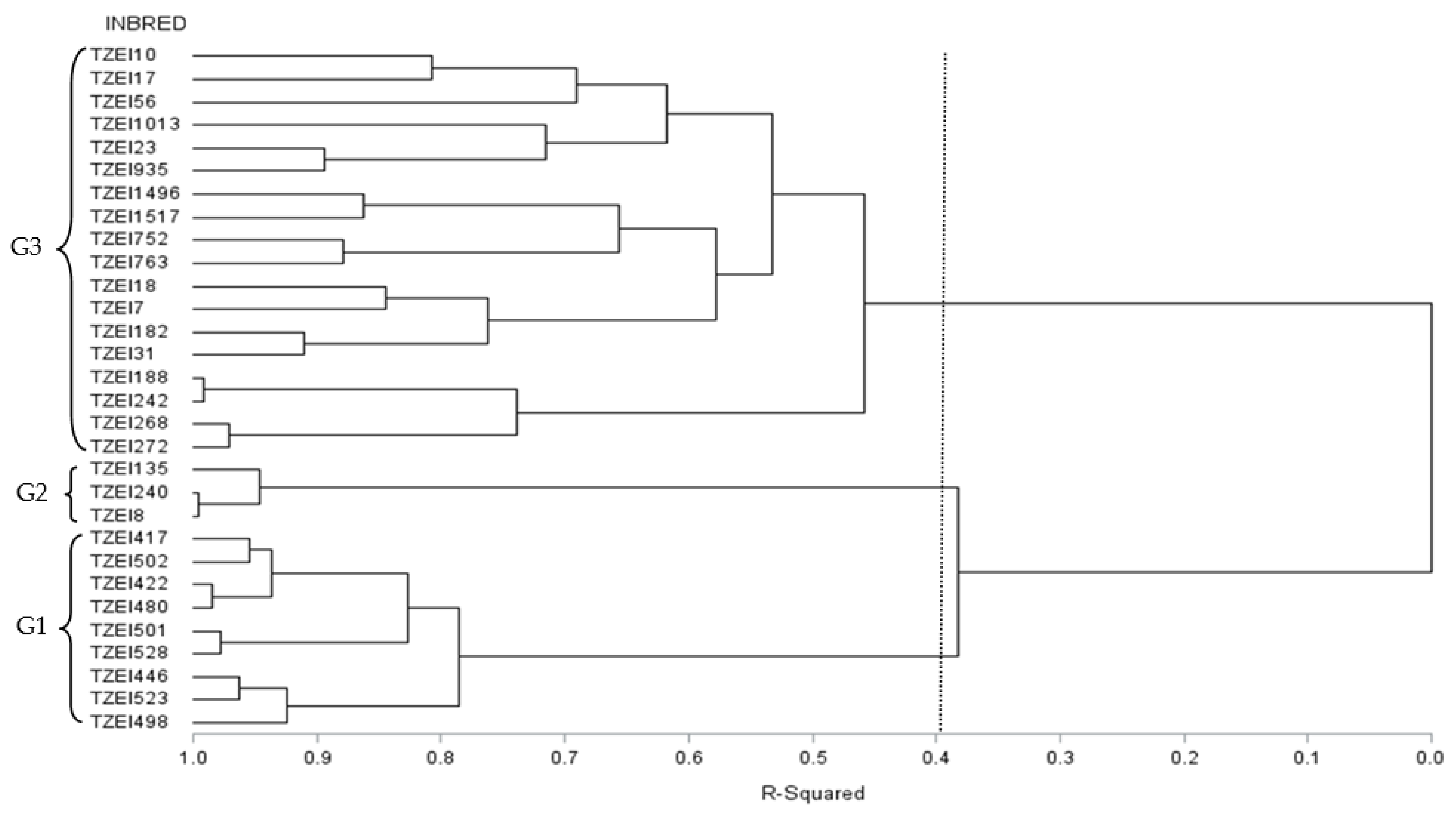
3.3. Heterotic Grouping and Relationship among Inbred Lines
3.3.1. Grouping Based on GCA of Multiple Traits
3.3.2. Grouping Based on DArTseq Marker Genetic Distances
3.4. Efficiency of Grouping Methods and Identification of Testers
3.5. Performance of Hybrids in Test Environments
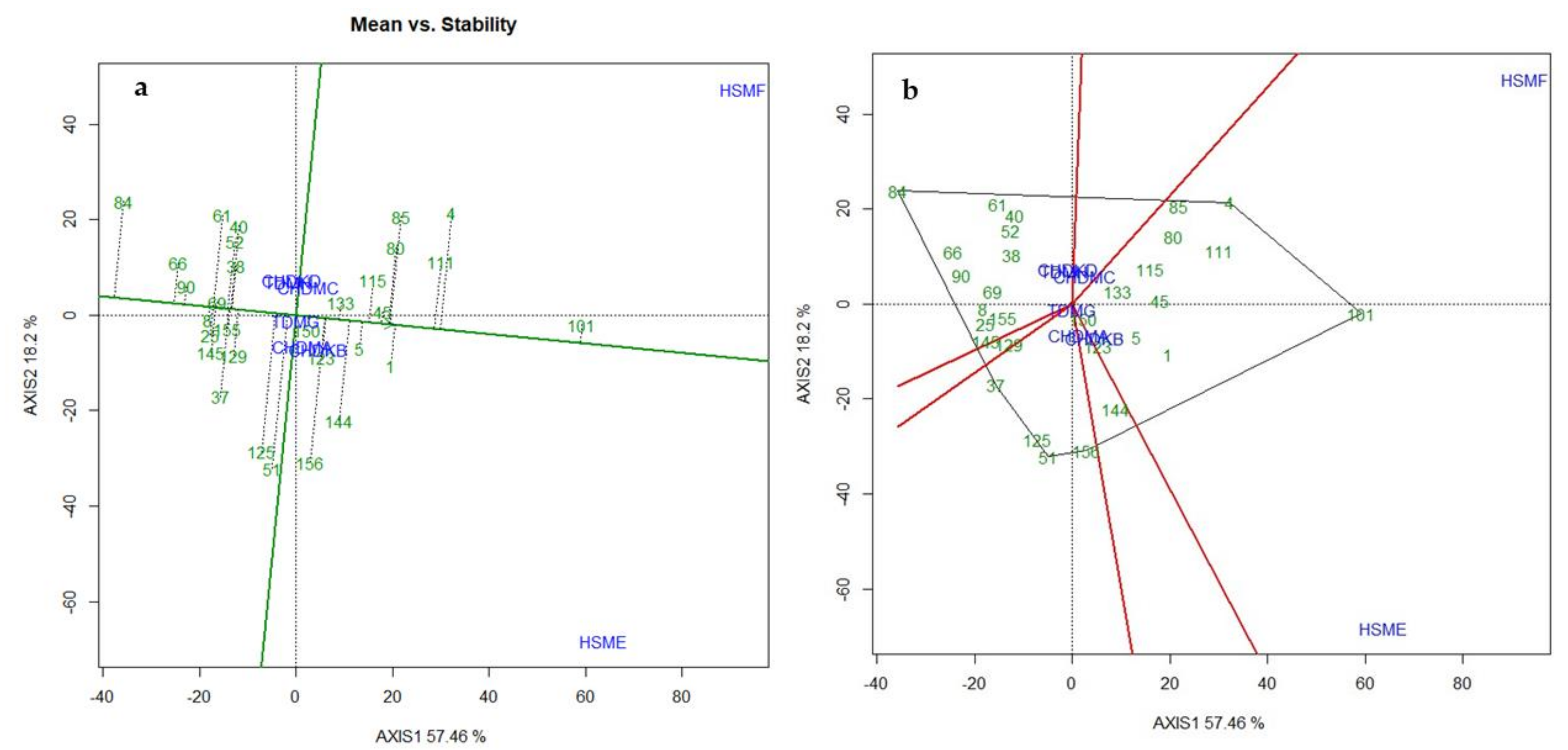
3.6. Stability of Hybrid Performance across Research Conditions
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Byerlee, D.; Edmeades, G. Fifty Years of Maize Research in the CGIAR: Diversity, Change and Ultimate Success; CIMMYT: Mexico City, Mexico, 2021. [Google Scholar]
- Badu-Apraku, B.; Fakorede, M.A.B. Improvement of early and extra-early maize for combined tolerance to drought and heat stress in sub-Saharan Africa. In Advances in Genetic Enhancement of Early and Extra-Early Maize for Sub-Saharan Africa; Springer: Cham, Switzerland, 2017; pp. 311–358. [Google Scholar]
- Daryanto, S.; Wang, L.; Jacinthe, P.A. Global synthesis of drought effects on maize and wheat production. PLoS ONE 2016, 11, e0156362. [Google Scholar] [CrossRef] [PubMed]
- NeSmith, D.S.; Ritchie, J.T. Effects of soil water-deficits during tassel emergence on development and yield components of maize (Zea mays L.). Field Crop. Res. 1992, 28, 251–256. [Google Scholar] [CrossRef]
- Fahad, S.; Hussain, S.; Saud, S.; Khan, F.; Hassan, S.; Nasim, W.; Huang, J. Exogenously applied plant growth regulators affect heat-stressed rice pollens. J. Agron. Crop Sci. 2016, 202, 139–150. [Google Scholar] [CrossRef]
- Cairns, J.E.; Crossa, J.; Zaidi, P.H.; Grudloyma, P.; Sanchez, C.; Araus, J.L.; Menkir, A. Identification of drought, heat, and combined drought and heat tolerant donors in maize. Crop Sci. 2013, 53, 1335–1346. [Google Scholar] [CrossRef] [Green Version]
- Nelimor, C.; Badu-Apraku, B.; Tetteh, A.Y.; N’guetta, A.S.P. Assessment of Genetic Diversity for Drought, Heat and Combined Drought and Heat Stress Tolerance in Early Maturing Maize Landraces. Plants 2019, 8, 518. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nelimor, C.; Badu-Apraku, B.; Tetteh, A.Y.; Garcia-Oliveira, A.L.; N’guetta, A.S.P. Assessing the Potential of Extra-Early Maturing Landraces for Improving Tolerance to Drought, Heat, and Both Combined Stresses in Maize. Agronomy 2020, 10, 318. [Google Scholar] [CrossRef] [Green Version]
- Prasad, P.V.V.; Djanaguiraman, M.; Perumal, R.; Ciampitti, I.A. Impact of high temperature stress on floret fertility and individual grain weight of grain sorghum: Sensitive stages and thresholds for temperature and duration. Front. Plant Sci. 2020, 6, 820. [Google Scholar] [CrossRef] [PubMed]
- Zaidi, P.H.; Zaman-Allah, M.; Trachsel, S.; Seetharam, K.; Cairns, J.E.; Vinayan, M.T. Phenotyping for Abiotic Stress Tolerance in Maize Heat Stress: A Field Manual; CIMMYT: Mexico City, Mexico, 2016. [Google Scholar]
- Begna, T. Combining ability and heterosis in plant improvement. Open J. Plant Sci. 2020, 6, 108–117. [Google Scholar]
- Acquaah, G. Principles of Plant Genetics and Breeding, 2nd ed.; Wiley-Blackwell: Oxford, UK, 2012. [Google Scholar]
- Brieger, F.G. The genetic basis of heterosis in maize. Genetics 1950, 35, 420–445. [Google Scholar] [CrossRef]
- Lee, M. DNA markers and plant breeding programmes. Adv. Agron. 1995, 55, 265–344. [Google Scholar]
- Akinwale, R.O. Heterosis and heterotic grouping among tropical maize germplasm. Cereal Grains 2021, 2, 59. [Google Scholar]
- Badu-Apraku, B.; Akinwale, R.O. Biplot analysis of line × tester data of maize (Zea mays L.) inbred lines under stress and nonstress environments. Cereal Res. Commun. 2019, 47, 518–530. [Google Scholar] [CrossRef]
- Badu-Apraku, B.; Fakorede, M.A.B.; Talabi, A.O.; Oyekunle, M.; Akaogu, I.C.; Akinwale, R.O.; Annor, B.; Melaku, G.; Fasanmade, Y.; Aderounmu, M. Gene action and heterotic groups of early white quality protein maize inbreds under multiple stress environments. Crop Sci. 2016, 56, 183–199. [Google Scholar] [CrossRef] [Green Version]
- Annor, B.; Badu-Apraku, B. Gene action controlling grain yield and other agronomic traits of extra-early quality protein maize under stress and non-stress conditions. Euphytica 2016, 212, 213–228. [Google Scholar] [CrossRef]
- Obeng-Bio, E. Genetic Analysis of Grain Yield and Other Traits of Early Maturing Provitamin A-Quality Protein Maize Inbred Lines under Drought and Low Soil Nitrogen Conditions. Ph.D. Thesis, University of Ghana, Accra, Ghana, 2018. [Google Scholar]
- Badu-Apraku, B.; Oyekunle, M.; Fakorede, M.A.B.; Vroh, I.; Akinwale, R.O.; Aderounmu, M. Combining ability, heterotic patterns and genetic diversity of extra-early yellow inbreds under contrasting environments. Euphytica 2013, 192, 413–433. [Google Scholar] [CrossRef]
- Badu-Apraku, B.; Oyekunle, M.; Akinwale, R.O.; Aderounmu., M. Combining ability and genetic diversity of extra-early white maize inbreds under stress and nonstress environments. Crop Sci. 2013, 53, 9–26. [Google Scholar] [CrossRef]
- Annor, B.; Badu-Apraku, B.; Nyadanu, D.; Akromah, R.; Fakorede, M.A.B. Testcross performance and combining ability of early maturing maize inbreds under multiple-stress environments. Sci. Rep. 2019, 9, 13809. [Google Scholar] [CrossRef] [Green Version]
- Nasser, L.M.; Badu-Apraku, B.; Gracen, V.E.; Mafouasson, H.N.A. Combining ability of early-maturing yellow maize inbreds under combined drought and heat stress and well-watered environments. Agronomy 2020, 10, 1585. [Google Scholar] [CrossRef]
- Osuman, A.S.; Badu-Apraku, B.; Ifie, B.E.; Tongoona, P.; Obeng-Bio, E.; Garcia-Oliveira, A.L. Genetic diversity, population structure and inter-trait relationships of combined heat and drought tolerant early-maturing maize inbred lines from west and central Africa. Agronomy 2020, 10, 1324. [Google Scholar] [CrossRef]
- Elshire, R.J.; Jeffrey, C.; Glaubitz, Q.S.; Jesse, A.; Poland, K.K.; Edward, S.B.; Sharon , E.M. A robust, simple genotyping-by-sequencing (GBS) approach for high diversity species. PloS one 2011, 6, e19379. [Google Scholar] [CrossRef] [Green Version]
- Kilian, A.; Sanewski, G.; Ko, L. The application of DArTseq technology to pineapple. Acta Hortic. 2016, 1111, 181–188. [Google Scholar] [CrossRef]
- Bradbury, P.J.; Zhang, Z.; Kroon, D.E.; Casstevens, T.M.; Ramdoss, Y.; Buckler, E.S. TASSEL: Software for association mapping of complex traits in diverse samples. Bioinformatics 2007, 23, 2633–2635. [Google Scholar] [CrossRef] [PubMed]
- SAS Institute Inc. SAS User’s Guide: Statistics; Version 9.4; SAS Institute Inc.: Cary, NC, USA, 2017. [Google Scholar]
- Cochran, W.G.; Cox, G.M. Experimental Designs; John Wiley and Sons: New York, NY, USA, 1960. [Google Scholar]
- Hallauer, A.R.; Miranda, J.B. Quantitative Genetics in Maize Breeding, 2nd ed.; Iowa State University Press: Ames, IA, USA, 1988. [Google Scholar]
- Baker, R.J. Issues in diallel analysis. Crop Sci. 1978, 18, 533–536. [Google Scholar] [CrossRef]
- Hung, H.Y.; Holland, J.B. Diallel analysis of resistance to fusarium ear rot and fumonisin contamination in maize. Crop Sci. 2012, 52, 2173–2181. [Google Scholar] [CrossRef] [Green Version]
- Badu-Apraku, B.; Oyekunle, M.; Akinwale, R.O.; Lum, A.F. Combining ability of early-maturing white maize inbreds under stress and non-stress environments. Agron. J. 2011, 103, 544–557. [Google Scholar] [CrossRef]
- Fan, X.M.; Zhang, Y.M.; Yao, W.H.; Chen, H.M.; Tan, J.; Xu, C.X.; Han, X.L.; Luo, L.M.; Kang, M.S. Classifying maize inbred lines into heterotic groups using a factorial mating design. Agron. J. 2009, 101, 106–112. [Google Scholar] [CrossRef]
- Badu-Apraku, B.; Fakorede, M.A.B.; Gedil, M.; Annor, B.; Talabi, A.O.; Akaogu, I.C.; Fasanmade, T.Y. Heterotic patterns of IITA and CIMMYT early-maturing yellow maize inbreds under contrasting environments. Agron. J. 2016, 108, 1321–1336. [Google Scholar] [CrossRef] [Green Version]
- Pswarayi, A.; Vivek, B.S. Combining ability amongst CIMMYT’s early maturing maize (Zea mays L.) germplasm under stress and non-stress conditions and identification of testers. Euphytica 2008, 162, 353–362. [Google Scholar] [CrossRef]
- Yan, W. GGE biplot: A windows application for graphical analysis of multi-environment trial data and other types of two-way data. Agron. J. 2001, 93, 1111–1118. [Google Scholar] [CrossRef] [Green Version]
- Edmeades, G.O.; Bolaños, J.; Chapman, S.C.; Lafiite, H.R.; Bänziger, M. Selection improves drought tolerance in tropical maize populations. I. Gains in biomass, grain yield and harvest index. Crop Sci. 1999, 39, 1306–1315. [Google Scholar] [CrossRef]
- Ziyomo, C.; Bernardo, R. Drought tolerance in maize: Indirect selection through secondary traits versus genome wide selection. Crop Sci. 2013, 53, 1269–1275. [Google Scholar] [CrossRef] [Green Version]
- Derera, J.; Tongoona, P.; Vivek, B.S.; Laing, M.D. Gene action controlling grain yield and secondary traits in southern African maize hybrids under drought and non-drought environments. Euphytica 2008, 162, 411–422. [Google Scholar] [CrossRef]
- Oyekunle, M.; Badu-Apraku, B. Genetic analysis of grain yield and other traits of early-maturing maize inbreds under drought and well-watered conditions. J. Agron. Crop Sci. 2014, 200, 92–107. [Google Scholar] [CrossRef]
- Adebayo, M.A.; Menkir, A.; Blay, E.; Gracen, V.; Danquah, E.; Hearne, S. Genetic analysis of drought tolerance in adapted× exotic crosses of maize inbred lines under managed stress conditions. Euphytica 2014, 196, 261–270. [Google Scholar] [CrossRef]
- Lanza, L.L.B.; de Souza, C.L., Jr.; Ottoboni, L.M.M.; Vieira, M.L.C.; de Souza, A.P. Genetic distance of inbred lines and prediction of maize single-cross performance using RAPD markers. Theor. Appl. Genet. 1997, 94, 1023–1030. [Google Scholar] [CrossRef]
- Badu-Apraku, B.; Fakorede, M.A.B.; Oyekunle, M.; Akinwale, R.O. Genetic gains in grain yield under nitrogen stress following three decades of breeding for drought tolerance and Striga resistance in early maturing maize. J. Agric. Sci. 2015, 154, 647–661. [Google Scholar] [CrossRef]
- Akinwale, R.O.; Badu-Apraku, B.; Fakorede, M.A.B.; Vroh-Bi, I. Heterotic grouping of tropical early-maturing maize inbred lines based on combining ability in Striga-infested and Striga-free environments and the use of SSR markers for genotyping. Field Crops Res. 2014, 156, 48–62. [Google Scholar] [CrossRef]
- Smith, O.S.; Smith, J.S.C.; Bowen, S.L.; Tenborg, R.A.; Wall, S.J. Similarities among a group of elite maize inbreds as measured by pedigree, F 1 grain yield, grain yield, heterosis, and RFLPs. Theor. Appl. Genet. 1990, 80, 833–840. [Google Scholar] [CrossRef]
- Senior, M.L.; Murphy, J.P.; Goodman, M.M.; Stuber, C.W. Utility of SSRs for determining genetic similarities an relationships in maize using an agarose gel system. Crop Sci. 1998, 38, 1088–1098. [Google Scholar] [CrossRef]
- Betran, F.J.; Beck, D.; Bänziger, M.; Edmeades, G.O. Genetic analysis of inbred and hybrid grain yield under stress and non-stress environments in tropical maize. Crop Sci. 2003, 43, 807–817. [Google Scholar] [CrossRef]
- Badu-Apraku, B.; Akinwale, R. Identification of early-maturing maize inbred lines based on multiple traits under drought and low N environments for hybrid development and population improvement. Can. J. Plant Sci. 2011, 91, 931–942. [Google Scholar] [CrossRef] [Green Version]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Source | a DF | b YIELD (kg/ha) | d SD (Days) | e ASI (Days) | j PLASP (Scale:1–9) | k EASP (Scale:1–9) | n EPP | p LF (%) | q TB |
|---|---|---|---|---|---|---|---|---|---|
| Combined Heat and Drought Stress | |||||||||
| 1 Env | 3 | 12,331,663.3 *** | 3859.85 *** | 1535.63 *** | 261.49 *** | 336.88 *** | 0.09 ** | 0.21 *** | 0.41 *** |
| Set | 5 | 3,434,567.8 *** | 59.27 *** | 11.34 | 2.75 *** | 7.85 *** | 0.15 *** | 0.15 *** | 0.39 *** |
| Env * set | 15 | 297,357.4 *** | 43.38 *** | 12.72 * | 3.29 *** | 4.85 *** | 0.17 *** | 0.02 * | 0.02 |
| Genotype | 155 | 465,564.8 *** | 18.73 *** | 7 | 1.03 *** | 1.21 *** | 0.07 *** | 0.02 *** | 0.04 *** |
| Male (set) | 24 | 486,510.5 *** | 28.05 *** | 6.19 | 1.54 *** | 1.16 *** | 0.09 *** | 0.03 *** | 0.05 *** |
| Female (set) | 24 | 729,472.5 *** | 24.23 *** | 7.92 | 1.71 *** | 2.11 *** | 0.09 *** | 0.02 ** | 0.04 ** |
| Female * male (set) | 96 | 222,641.04 *** | 11.51 | 6.74 | 0.67 *** | 0.64 *** | 0.04 *** | 0.01 * | 0.02 |
| Genotype * env | 465 | 189,430.7 *** | 13.86 *** | 7.11 | 0.72 *** | 1.04 *** | 0.06 *** | 0.01 | 0.02 |
| Env * male (set) | 72 | 167,949.31 *** | 14.15 * | 6.71 | 0.7 *** | 0.71 *** | 0.05 *** | 0.01 | 0.02 |
| Env * female (set) | 72 | 247,017.7 *** | 18.09 *** | 6.39 | 0.89 *** | 1.21 *** | 0.07 *** | 0.01 *** | 0.03 * |
| Env * female * male (set) | 288 | 162,849.2 *** | 10.5 | 7.03 | 0.54 ** | 0.65 *** | 0.05 *** | 0.01 | 0.02 |
| Error | 522 | 7707.2 | 10.43 | 6.76 | 0.41 | 0.42 | 0.02 | 0.01 | 0.02 |
| Heat Stress | |||||||||
| Env | 1 | 872,905,949.1 *** | 11574.18 *** | 1196.44 | 22.19 *** | 61.97 *** | 0.76 *** | 0.05 | 0.38 |
| Set | 5 | 19,992,488.1 *** | 55.95 *** | 423.37 | 2.94 *** | 4.21 *** | 0.16 *** | 0.03 * | 0.09 |
| Env * set | 5 | 3,670,697.2 *** | 6.17 | 424.03 | 1.98 *** | 2.11 ** | 0.58 *** | 0.02 | 0.21 |
| Genotype | 155 | 4,807,572.3 *** | 15.94 *** | 439.79 | 0.91 *** | 1.39 *** | 0.07 *** | 0.18 *** | 0.25 *** |
| Male (set) | 24 | 7,941,325.8 *** | 23.48 *** | 391.07 | 0.83 *** | 2.49 *** | 0.06 ** | 0.01 | 0.15 |
| Female (set) | 24 | 7,003,187.1 *** | 27.12 *** | 387.34 | 1.49 *** | 1.49 *** | 0.07 *** | 0.03 ** | 0.17 |
| Female * male (set) | 96 | 2,683,972.1 *** | 10.07 | 484.01 | 0.68 *** | 0.99 *** | 0.05 *** | 0.02 | 0.11 |
| Genotype * env | 155 | 2,878,702.1 *** | 12.18 *** | 436.93 | 0.700 *** | 0.83 ** | 0.09 *** | 0.19 *** | 0.28 *** |
| Env * male (set) | 24 | 2,070,038.2 *** | 16.69 ** | 366.51 | 0.61 * | 0.86 | 0.06 ** | 0.02 | 0.17 * |
| Env * female (set) | 24 | 3,425,366.8 *** | 8.49 | 371.51 | 0.69 ** | 0.94 * | 0.07 *** | 0.01 | 0.11 |
| Env * female * male (set) | 96 | 2,657,296.4 *** | 11.03 * | 488.09 | 0.64 *** | 0.81 * | 0.06 *** | 0.02 | 0.12 |
| Error | 262 | 191,640 | 7.68166 | 437.585 | 0.38 | 0.59 | 0.03 | 0.05 | 0.13 |
| Source | a DF | b YIELD (kg/ha) | d SD (Days) | e ASI (Days) | j PLASP (Scale:1–9) | k EASP (Scale:1–9) | n EPP | p LF (%) | q TB (%) |
|---|---|---|---|---|---|---|---|---|---|
| Terminal Drought Stress | |||||||||
| 1 Env | 1 | 8,547,258.20 *** | 141.27 *** | 0.0005 | 1552.78 *** | 10.32 *** | 1.24 ** | 788.57 *** | |
| Set | 5 | 2,768,389.49 *** | 24.87 *** | 9.06 *** | 3.97 *** | 1.54 *** | 0.27 | 0.88 ** | |
| Env * set | 5 | 546,048.98 * | 1.18 | 2.05 * | 0.41 | 0.14 | 0.14 | 0.78 ** | - |
| Genotype | 155 | 855,779.0 *** | 5.58 *** | 1.61 *** | 0.63 *** | 0.42 *** | 0.21 * | 0.42 *** | - |
| Male (set) | 24 | 702,898.04 *** | 10.13 *** | 1.86 ** | 0.63 *** | 0.36 ** | 0.15 | 0.51 ** | - |
| Female (set) | 24 | 1,442,564.23 *** | 7.47 *** | 1.69 ** | 0.71 *** | 0.35 ** | 0.21 | 0.62 *** | - |
| Female * male (set) | 96 | 663,950.50 *** | 2.75 ** | 1.13 | 0.44 *** | 0.29 ** | 0.22 * | 0.31 | - |
| Genotype * env | 155 | 418,299.9 *** | 2.03 | 1.27 ** | 0.37 ** | 0.26 * | 0.17 | 0.37 ** | - |
| Env * male (set) | 24 | 348,748.92 * | 1.86 | 1.50 * | 0.36 | 0.26 | 0.15 | 0.48 ** | - |
| Env * female (set) | 24 | 581,963.92 *** | 1.81 | 1.33 | 0.41 * | 0.33 * | 0.13 | 0.59 *** | - |
| Env * female * male (set) | 96 | 345,575.29 ** | 2.01 | 1.18 | 0.32 | 0.21 | 0.19 | 0.24 | - |
| Error | 262 | 209,950.4 | 2 | 0.95 | 0.26 | 0.21 | 0.16 | 0.25 | - |
| Across Test Environments | |||||||||
| 1 Env | 7 | 459,751,108 *** | 11343.41 *** | 1333.61 *** | 387.24 *** | 483.81 *** | 1.83 *** | 0.26 *** | 0.39 *** |
| Set | 5 | 13,383,275 *** | 12.22 | 176.86 | 6.21 *** | 7.36 *** | 0.15 * | 0.17 *** | 0.44 *** |
| Env *set | 35 | 2,502,667 *** | 36.99 *** | 104.84 | 2.28 *** | 3.32 *** | 0.24 *** | 0.02 * | 0.06 |
| 2 Rep(env * set) | 40 | 104,024 | 7.09 | 50.14 | 0.42 | 0.68 ** | 0.08 * | 0.01 | 0.07 |
| Block(env * rep) | 194 | 116,728 | 17.87 *** | 109.61 | 0.71 *** | 0.82 *** | 0.07 | 0.01 ** | 0.07 * |
| Genotype | 155 | 378,998,432 *** | 16.59 *** | 111.53 | 1.33 *** | 1.47 *** | 0.11 *** | 0.08 *** | 0.12 *** |
| Male (set) | 24 | 3,849,768 *** | 34.35 *** | 97.13 | 1.79 *** | 2.11 *** | 0.09 * | 0.03 *** | 0.07 |
| Female (set) | 24 | 4,043,843 *** | 24.33 *** | 95.69 | 2.56 *** | 2.73 *** | 0.16 *** | 0.03 *** | 0.08 * |
| Female * male (set) | 96 | 1,135,578 *** | 10.26 * | 121.45 | 0.72 *** | 0.68 *** | 0.09 *** | 0.02 *** | 0.06 |
| Genotype * env | 1085 | 1,1805,42,807 *** | 11.31 *** | 113.59 | 0.64 *** | 0.83 *** | 0.11 *** | 0.07 *** | 0.11 *** |
| Env * male (set) | 168 | 1,220,148 *** | 12.44 *** | 99.54 | 0.61 *** | 0.76 *** | 0.08 *** | 0.01 | 0.07 ** |
| Env * female (set) | 168 | 1,442,533 *** | 14.24 *** | 100.63 | 0.72 *** | 0.88 *** | 0.09 *** | 0.01 ** | 0.06 |
| Env * female * male (set) | 672 | 847,896 *** | 8.36 | 125.31 | 0.52 ** | 0.61 *** | 0.09 *** | 0.01 | 0.05 |
| Error | 1046 | 104,436 | 7.63 | 113.21 | 0.36 | 0.41 | 0.06 | 0.02 | 0.06 |
| GY (kg/ha) | SD (Days) | ASI (Days) | PLASP (Scale:1–9) | EASP (Scale:1–9) | EPP | LF (%) | TB (%) | |
|---|---|---|---|---|---|---|---|---|
| Combined Heat and Drought Stress | ||||||||
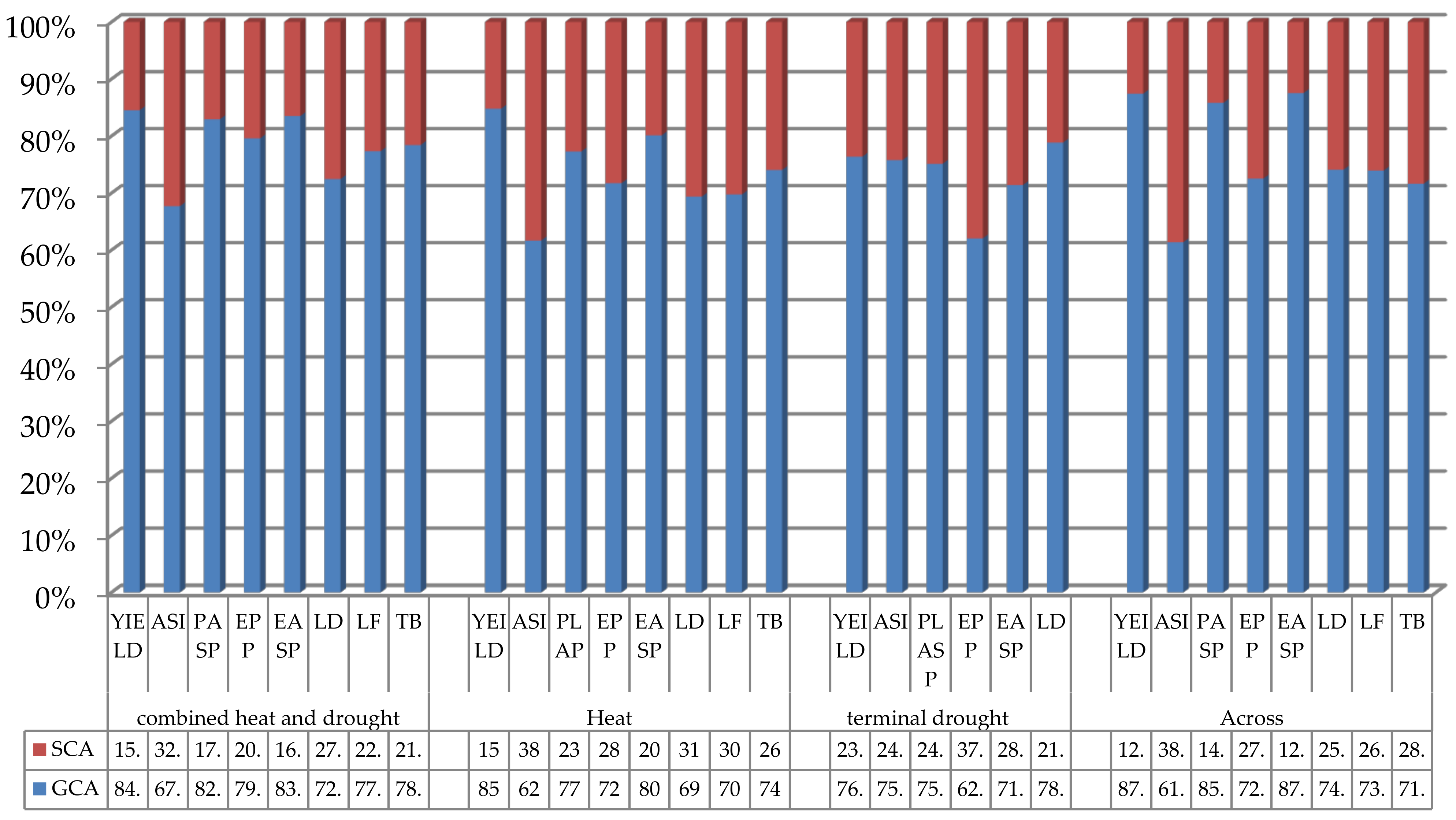
| GCA | 1,215,983 | 52.28 | 14.11 | 3.25 | 3.27 | 0.18 | 0.05 | 0.09 |
| SCA | 222,641 | 11.51 | 6.74 | 0.67 | 0.64 | 0.04 | 0.01 | 0.02 |
| 0.845 | 0.82 | 0.677 | 0.829 | 0.836 | 0.818 | 0.833 | 0.818 | |
| Heat Stress | ||||||||
| GCA | 14,944,513 | 50.6 | 778.41 | 2.32 | 3.98 | 0.13 | 0.04 | 0.32 |
| SCA | 2,683,972 | 10.07 | 484.01 | 0.68 | 0.99 | 0.05 | 0.02 | 0.11 |
| 0.848 | 0.834 | 0.617 | 0.773 | 0.801 | 0.722 | 0.667 | 0.744 | |
| Terminal Drought Stress | ||||||||
| GCA | 2,145,462 | 17.6 | 3.55 | 1.34 | 0.71 | 0.36 | 1.13 | - |
| SCA | 663,950.5 | 2.75 | 1.13 | 0.44 | 0.29 | 0.22 | 0.31 | - |
| 0.764 | 0.865 | 0.759 | 0.753 | 0.71 | 0.621 | 0.785 | - | |
| Across Environments | ||||||||
| GCA | 7,893,611 | 58.68 | 192.82 | 4.35 | 4.84 | 0.25 | 0.06 | 0.15 |
| SCA | 1,135,578 | 10.26 | 121.45 | 0.72 | 0.68 | 0.09 | 0.02 | 0.06 |
| 0.874 | 0.851 | 0.614 | 0.858 | 0.877 | 0.735 | 0.75 | 0.714 | |
| Yield Group | Cross Type | HGCAMT | DArTseq Markers |
|---|---|---|---|
| 1 | Inter | 45 | 30 |
| 1 | Intra | 5 | 20 |
| 2 | Inter | 39 | 30 |
| 2 | Intra | 11 | 20 |
| 3 | Inter | 38 | 26 |
| 3 | Intra | 12 | 24 |
| Breeding efficiency (%) | 39.8 | 36.2 | |
| 1 | Inter | 43 | 32 |
| 1 | Intra | 7 | 18 |
| 2 | Inter | 46 | 34 |
| 2 | Intra | 6 | 16 |
| 3 | Inter | 39 | 18 |
| 3 | Intra | 11 | 32 |
| Breeding efficiency (%) | 41.6 | 43.2 | |
| 1 | Inter | 37 | 31 |
| 1 | Intra | 13 | 19 |
| 2 | Inter | 41 | 29 |
| 2 | Intra | 9 | 21 |
| 3 | Inter | 40 | 24 |
| 3 | Intra | 10 | 26 |
| Breeding efficiency (%) | 31.3 | 38.14 | |
| 1 | Inter | 41 | 30 |
| 1 | Intra | 9 | 20 |
| 2 | Inter | 38 | 33 |
| 2 | Intra | 12 | 17 |
| 3 | Inter | 41 | 22 |
| 3 | Intra | 9 | 28 |
| Breeding efficiency (%) | 32 | 39.2 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Osuman, A.S.; Badu-Apraku, B.; Ifie, B.E.; Nelimor, C.; Tongoona, P.; Obeng-Bio, E.; Karikari, B.; Danquah, E.Y. Combining Ability and Heterotic Patterns of Tropical Early-Maturing Maize Inbred Lines under Individual and Combined Heat and Drought Environments. Plants 2022, 11, 1365. https://0-doi-org.brum.beds.ac.uk/10.3390/plants11101365
Osuman AS, Badu-Apraku B, Ifie BE, Nelimor C, Tongoona P, Obeng-Bio E, Karikari B, Danquah EY. Combining Ability and Heterotic Patterns of Tropical Early-Maturing Maize Inbred Lines under Individual and Combined Heat and Drought Environments. Plants. 2022; 11(10):1365. https://0-doi-org.brum.beds.ac.uk/10.3390/plants11101365
Chicago/Turabian StyleOsuman, Alimatu Sadia, Baffour Badu-Apraku, Beatrice Elohor Ifie, Charles Nelimor, Pangirayi Tongoona, Ebenezer Obeng-Bio, Benjamin Karikari, and Eric Yirenkyi Danquah. 2022. "Combining Ability and Heterotic Patterns of Tropical Early-Maturing Maize Inbred Lines under Individual and Combined Heat and Drought Environments" Plants 11, no. 10: 1365. https://0-doi-org.brum.beds.ac.uk/10.3390/plants11101365