Metabolomics Response for Drought Stress Tolerance in Chinese Wheat Genotypes (Triticum aestivum)

Abstract
:1. Introduction
2. Results
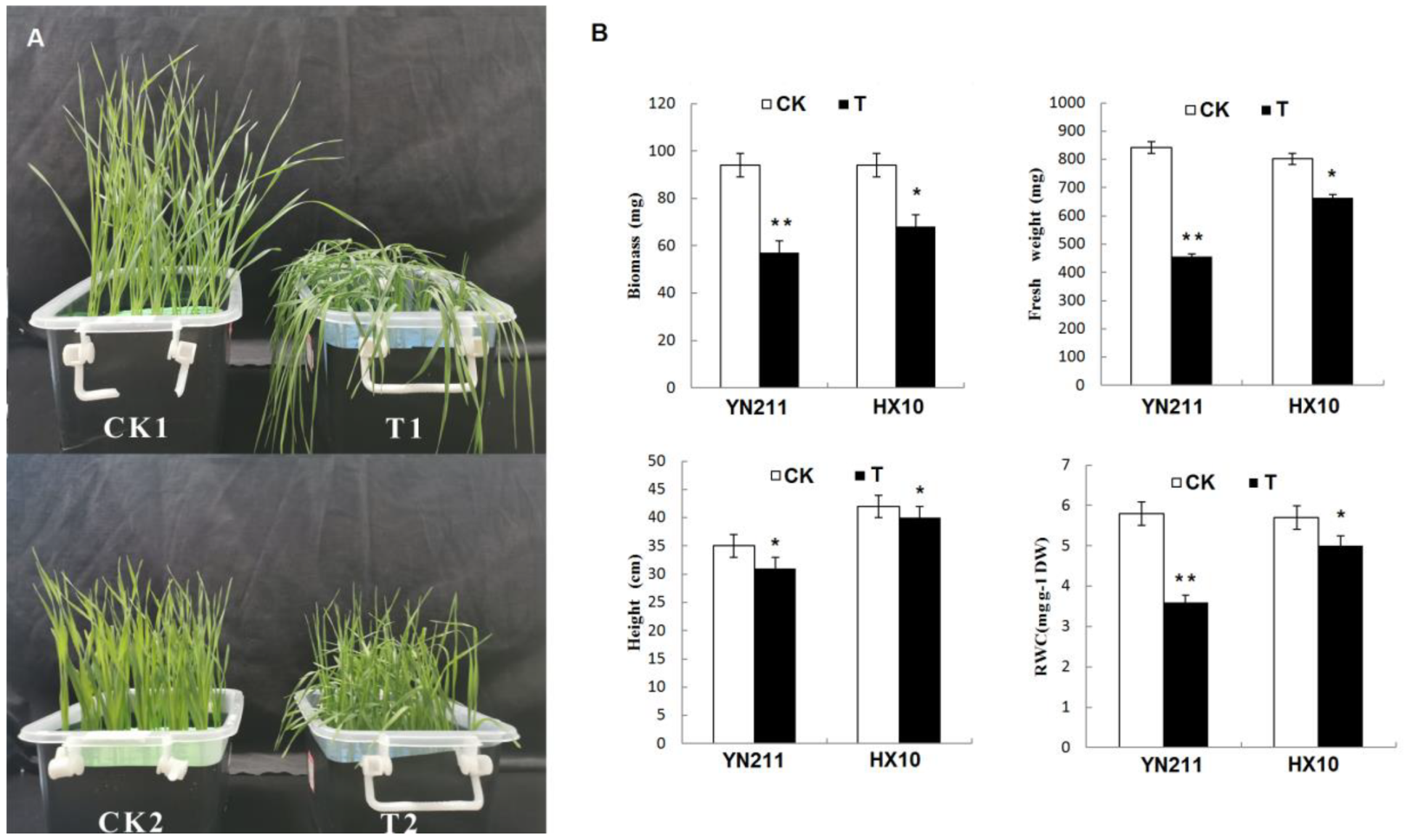
2.1. Physiological Responses to Drought Stress
2.2. Metabolite Profile under Drought Stress
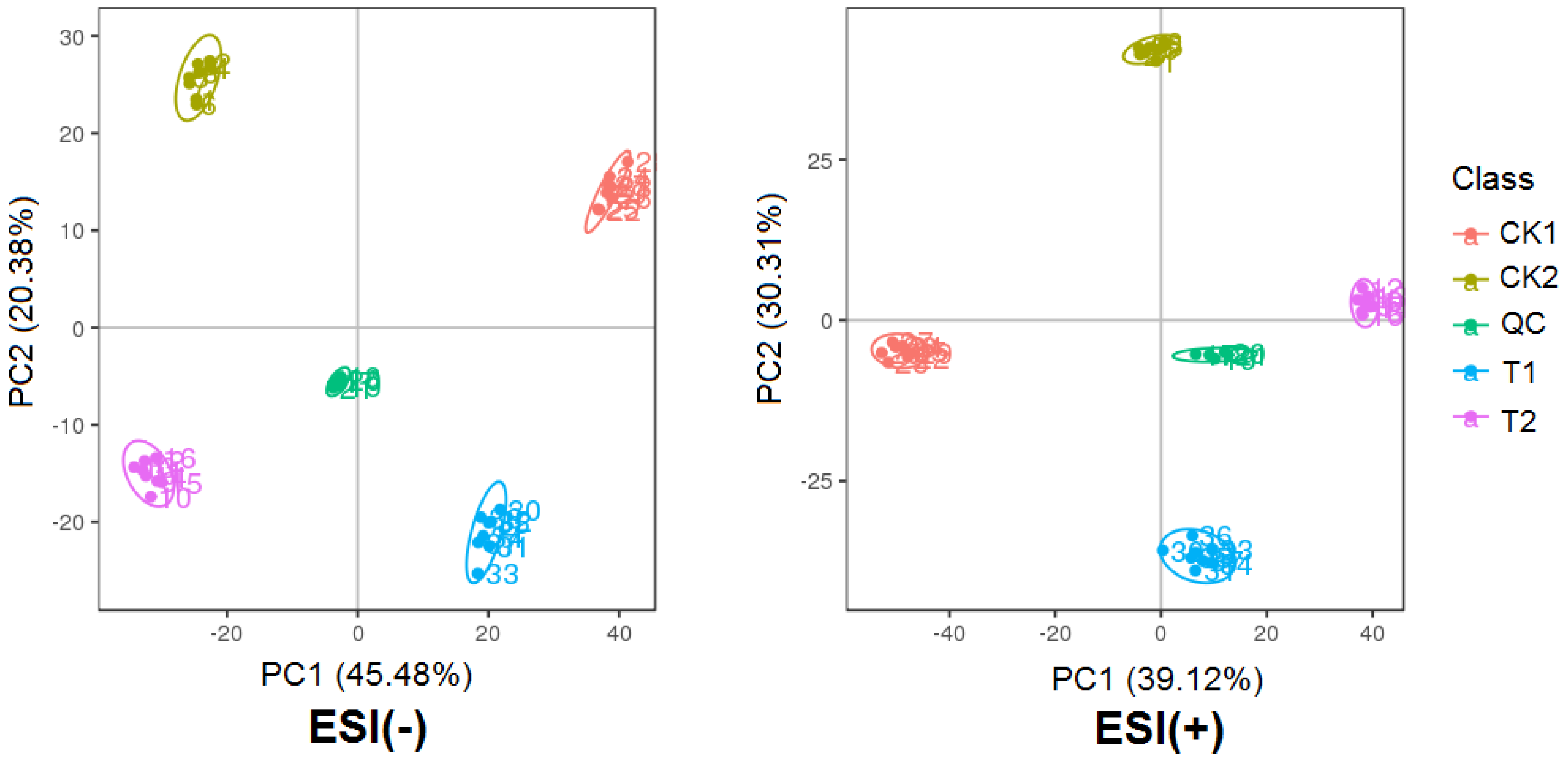
2.2.1. Principal Component Analysis of Metabolites Profiles
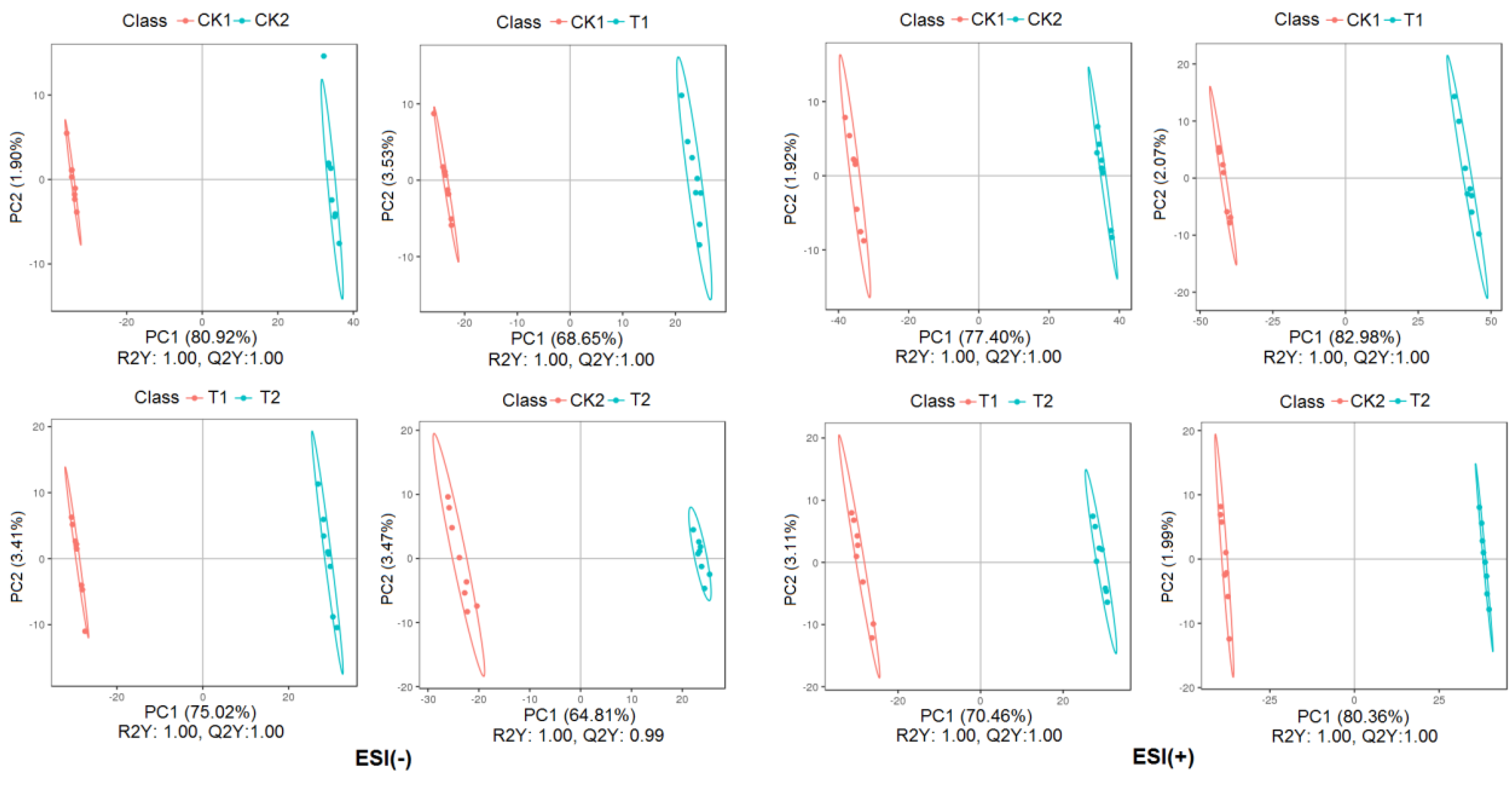
2.2.2. Partial Least Square Discriminant Analysis of Metabolites Profiles
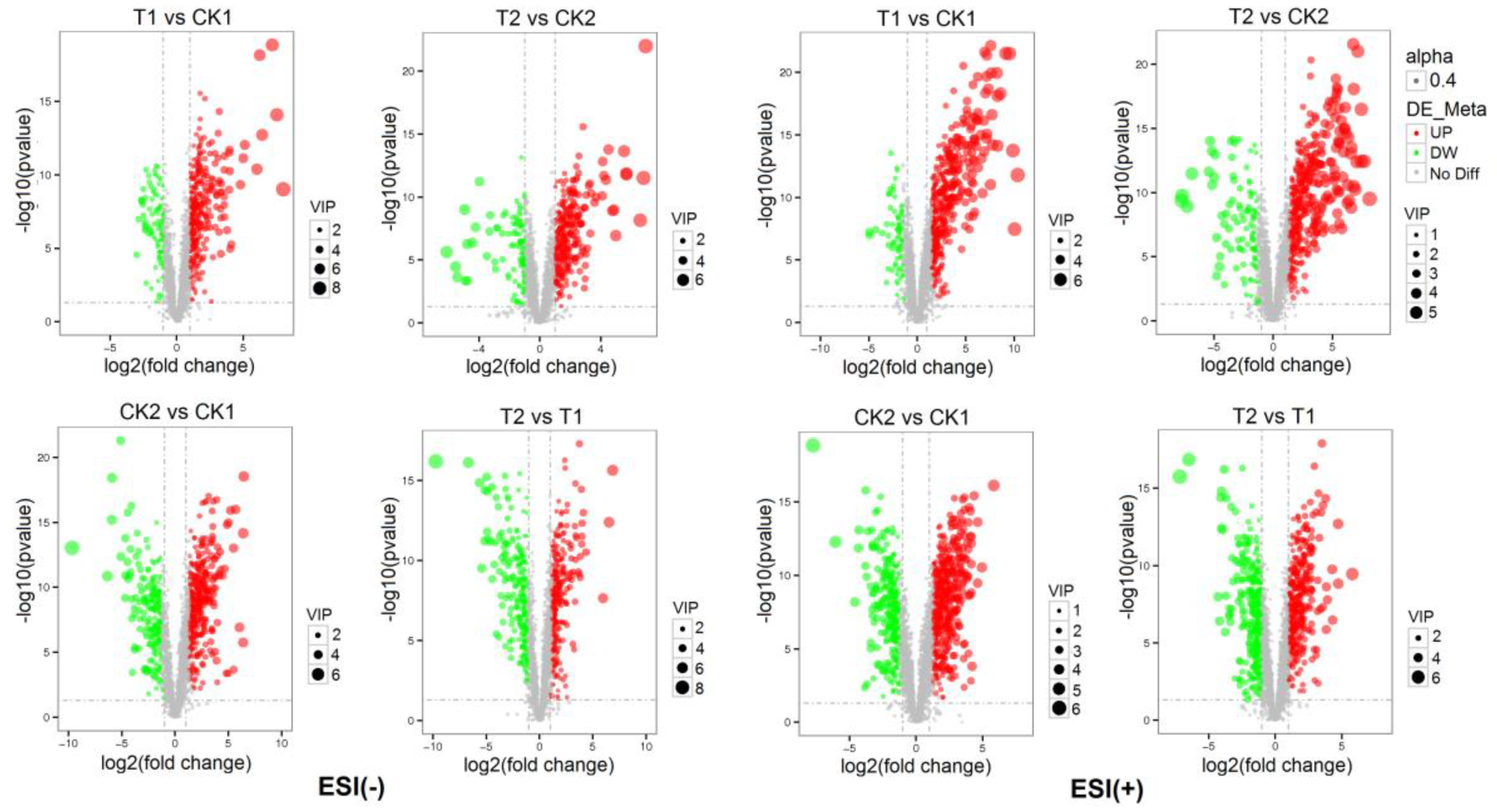
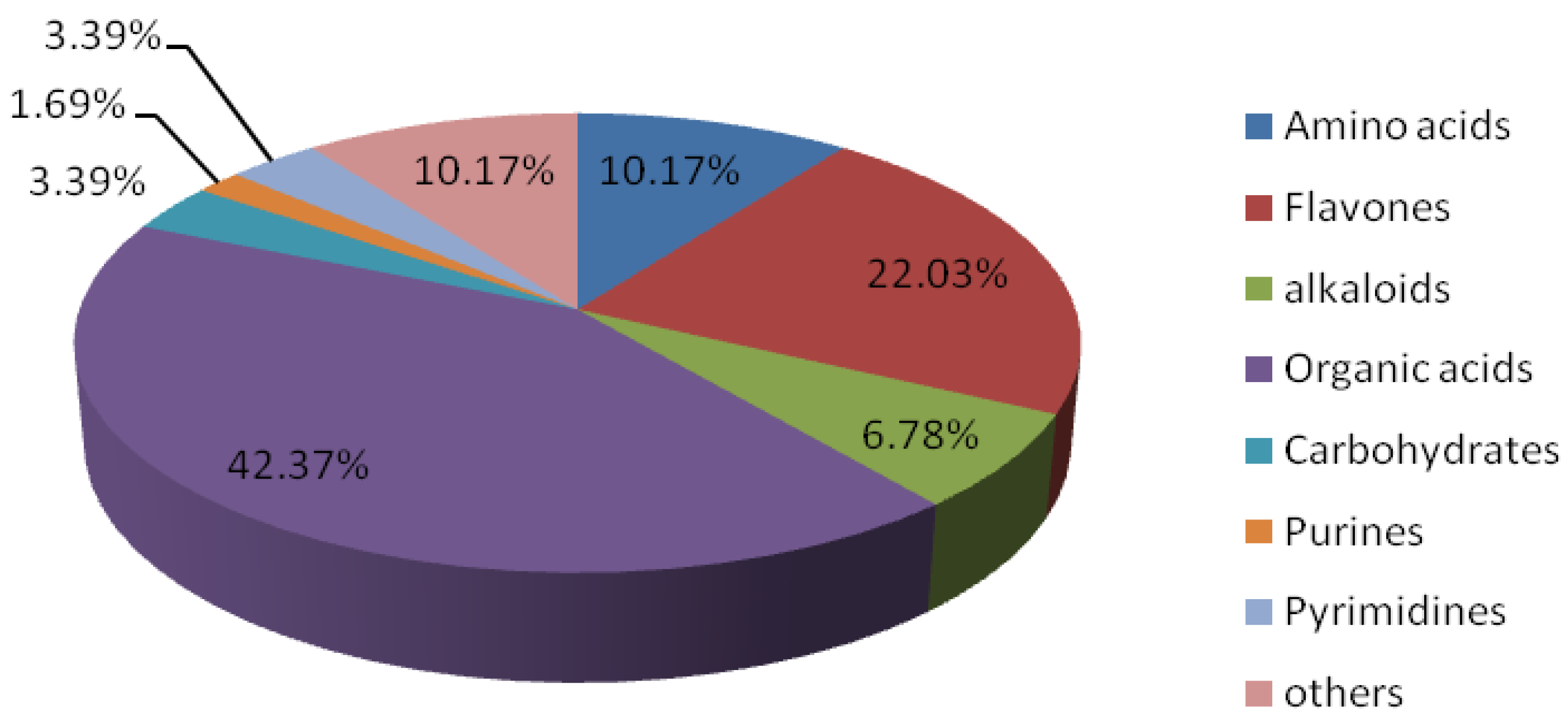
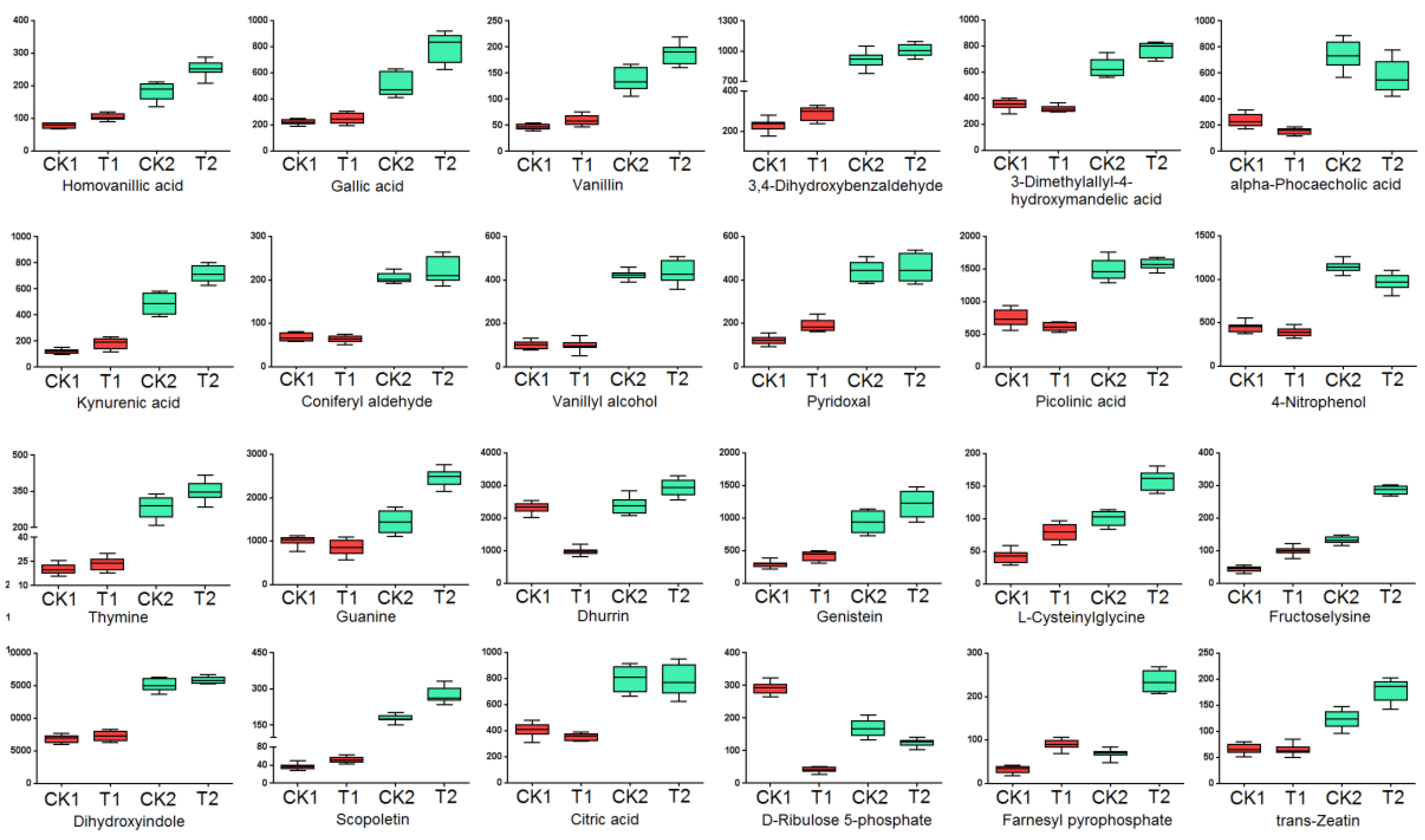
2.2.3. Dynamic Metabolic Changes of Drought-Tolerant Genotype and Drought-Sensitive Wheat Genotype under Normal Conditions or Drought Stress
3. Discussion
4. Materials and Methods
4.1. Materials and Growth Conditions
4.2. Treatments
4.3. Growth Parameters
4.4. Metabolite Extraction
4.5. Chromatographic Separation
4.6. Mass Spectrometry
4.7. Data Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Selim, D.A.-F.H.; Nassar, R.M.A.; Boghdady, M.S.; Bonfill, M. Physiological and anatomical studies of two wheat cultivars irrigated with magnetic water under drought stress conditions. Plant Physiol. Biochem. 2019, 135, 480–488. [Google Scholar] [CrossRef] [PubMed]
- Yu, X.; Wang, T.; Zhu, M.; Zhang, L.; Zhang, F.; Jing, E.; Ren, Y.; Wang, Z.; Xin, Z.; Lin, T. Transcriptome and physiological analyses for revealing genes involved in wheat response to endoplasmic reticulum stress. BMC Plant Biol. 2019, 19, 193. [Google Scholar] [CrossRef] [PubMed]
- McKay, J.K.; Richards, J.H.; Mitchell-Olds, T. Genetics of drought adaptation in Arabidopsis thaliana: I. Pleiotropy contributes to genetic correlations among ecological traits. Mol. Ecol. 2003, 12, 1137–1151. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bal, W.; Kozlowski, H.; Robbins, R.; Pettit, L.D. Improving Drought Tolerance by Exogenous Application of Glycinebetaine and Salicylic Acid in Sunflower. J. Agron. Crop Sci. 2010, 194, 193–199. [Google Scholar]
- Taiz, L.; Zeiger, E. Plant Physiology, 4th ed.; Sinauer Associates Inc Publishers: Sunderland, MA, USA, 2006. [Google Scholar]
- Steudle, E.; Peterson, C.A. How does water get through roots? J. Exp. Bot. 1998, 49, 775–788. [Google Scholar] [CrossRef]
- Cattivelli, L.; Rizza, F.; Badeck, F.-W.; Mazzucotelli, E.; Mastrangelo, A.M.; Francia, E.; Marè, C.; Tondelli, A.; Stanca, A.M. Drought tolerance improvement in crop plants: An integrated view from breeding to genomics. Field Crops Res. 2008, 105, 1–14. [Google Scholar] [CrossRef]
- Prasad, P.V.V.; Staggenborg, S.; Ristic, Z. Impacts of drought and/or heat stress on physiological, developmental, growth, and yield processes of crop plants. Adv. Agric. Syst. Model. Ser. 2008, 1, 301–355. [Google Scholar]
- Foulkes, M.J.; Sylvester-Bradley, R.; Weightman, R.; Snape, J.W. Identifying physiological traits associated with improved drought resistance in winter wheat. Field Crops Res. 2007, 103, 11–24. [Google Scholar] [CrossRef]
- Godfray, H.C.J.; Beddington, J.R.; Crute, I.R.; Lawrence, H.; David, L.; Muir, J.F.; Jules, P.; Sherman, R.; Thomas, S.M.; Camilla, T. Food security: The challenge of feeding 9 billion people. Science 2010, 327, 812. [Google Scholar] [CrossRef] [Green Version]
- Li, X.; Waddington, S.R.; Dixon, J.; Joshi, A.K.; Vicente, M.C.D. The relative importance of drought and other water-related constraints for major food crops in South Asian farming systems. Food Secur. 2011, 3, 19–33. [Google Scholar] [CrossRef]
- Dixon, R.A.; Paiva, N.L. Stress-Induced Phenylpropanoid Metabolism. Plant cell 1995, 7, 1085–1097. [Google Scholar] [CrossRef] [PubMed]
- Adams, H.D.; Guardiolaclaramonte, M.; Barrongafford, G.A.; Villegas, J.C.; Breshears, D.D.; Zou, C.B.; Troch, P.A.; Huxman, T.E. Reply to Leuzinger et al: Drought-induced tree mortality temperature sensitivity requires pressing forward with best available science. Proc. Natl Acad. Sci. USA 2009, 106, E107. [Google Scholar] [CrossRef] [Green Version]
- Spickett, C.M. Metabolic Response of MaizeRootstoHyperosmotic Shock’ AninVivo 31P Nuclear Magnetic Resonance Study. Plant Physiol. 1992, 99, 856–863. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hare, P.D.; Cress, W.A.; Van Staden, J. Dissecting the roles of osmolyte accumulation during stress. Plant Cell Environ. 1998, 21, 535–553. [Google Scholar]
- Dennison, K.L.; Robertson, W.R.; Lewis, B.D.; Hirsch, R.E.; Sussman, M.R.; Spalding, E.P. Functions of AKT1 and AKT2 potassium channels determined by studies of single and double mutants of Arabidopsis. Plant Physiol. 2001, 127, 1012–1019. [Google Scholar] [CrossRef] [PubMed]
- Chen, T.H.H.; Murata, N. Enhancement of tolerance of abiotic stress by metabolic engineering of betaines and other compatible solutes. Curr. Opin. Plant Biol. 2002, 5, 250–257. [Google Scholar] [CrossRef]
- Yasar, F.; Uzal, O.; Tufenkci, S.; Yildiz, K.; Pasa, O. Ion accumulation in different organs of green bean genotypes grown under salt stress. Plant Soil Environ. 2006, 52, 476. [Google Scholar] [CrossRef] [Green Version]
- Wang, H. Effects of salt stress on ion balance and nitrogen metabolism of old and young leaves in rice (Oryza sativa L.). BMC Plant Biol. 2012, 12, 194. [Google Scholar] [CrossRef] [Green Version]
- Oliver, F. Metabolomics—The link between genotypes and phenotypes. Plant Mol. Biol. 2002, 48, 155–171. [Google Scholar]
- Hillenmeyer, M.E.; Ericson, E.; Davis, R.W.; Nislow, C.; Koller, D.; Giaever, G. Systematic analysis of genome-wide fitness data in yeast reveals novel gene function and drug action. Genome Biol. 2010, 11, R30. [Google Scholar] [CrossRef] [Green Version]
- Marco, O.; Stephan, L.; Simon, P.; Kirsten, S.; Nadine, S.; Christiane, W. Metabolomics: Current state and evolving methodologies and tools. Appl. Microbiol. Biotechnol. 2007, 76, 495–511. [Google Scholar]
- Meng, J.; Zhang, X.; Wu, H.; Bu, J.; Shi, C.; Deng, C.; Mao, Y. Morphine-induced conditioned place preference in mice: Metabolomic profiling of brain tissue to find “molecular switch” of drug abuse by gas chromatography/mass spectrometry. Anal. Chim. Acta 2012, 710, 125–130. [Google Scholar] [CrossRef] [PubMed]
- Ruan, C.J.; Ja, T.D.S. Metabolomics: Creating new potentials for unraveling the mechanisms in response to salt and drought stress and for the biotechnological improvement of xero-halophytes. Crit. Rev. Biotechnol. 2011, 31, 153–169. [Google Scholar] [CrossRef] [PubMed]
- Barding, G.A.; Szabolcs, B.; Takeshi, F.; Julia, B.S.; Larive, C.K. Comparison of GC-MS and NMR for metabolite profiling of rice subjected to submergence stress. J. Proteome Res. 2013, 12, 898–909. [Google Scholar] [CrossRef]
- Bowne, J.B.; Erwin, T.A.; Juttner, J.; Schnurbusch, T.; Langridge, P.; Bacic, A.; Roessner, U. Drought Responses of Leaf Tissues from Wheat Cultivars of Differing Drought Tolerance at the Metabolite Level. Mol. Plant 2012, 5, 418–429. [Google Scholar] [CrossRef] [Green Version]
- Alvarez, S.; Marsh, E.L.; Schroeder, S.G.; Schachtman, D.P. Metabolomic and proteomic changes in the xylem sap of maize under drought. Plant Cell Environ. 2008, 31, 325–340. [Google Scholar] [CrossRef]
- Charlton, A.J.; Donarski, J.A.; Harrison, M.; Jones, S.A.; Godward, J.; Oehlschlager, S.; Arques, J.L.; Ambrose, M.; Chinoy, C.; Mullineaux, P.M.; et al. Responses of the pea (Pisumsativum L.) leaf metabolome to drought stress assessed by nuclear magnetic resonance spectroscopy. Metabolomics 2008, 4, 312. [Google Scholar] [CrossRef]
- Rice-Evans, C.; Miller, N.; Paganga, G. Antioxidant properties of phenolic compounds. Trends Plant Sci. 1997, 2, 152–159. [Google Scholar] [CrossRef]
- Varela, M.C.; Arslan, I.; Reginato, M.A.; Cenzano, A.M.; Luna, M.V. Phenolic compounds as indicators of drought resistance in shrubs from Patagonian shrublands (Argentina). Plant Physiol. Biochem. 2016, 104, 81–91. [Google Scholar] [CrossRef]
- Sarker, U.; Oba, S. Drought stress enhances nutritional and bioactive compounds, phenolic acids and antioxidant capacity of Amaranthus leafy vegetable. BMC Plant Biol. 2018, 18, 258. [Google Scholar] [CrossRef] [Green Version]
- Quan, N.T.; Anh, L.H.; Khang, D.T.; Tuyen, P.T.; Toan, N.P.; Minh, T.N.; Minh, L.T.; Bach, D.T.; Ha, P.T.T.; Elzaawely, A.A.; et al. Involvement of Secondary Metabolites in Response to Drought Stress of Rice (Oryza sativa L.). Agriculture 2016, 6, 23. [Google Scholar] [CrossRef] [Green Version]
- Baier, M.; Kandlbinder, A.; Golldack, D.; Dietz, K.-J. Oxidative stress and ozone: Perception, signalling and response. Plant Cell Environ. 2005, 28, 1012–1020. [Google Scholar] [CrossRef]
- Tarchoune, I.; Sgherri, C.; Izzo, R.; Lachaal, M.; Ouerghi, Z.; Navari-Izzo, F. Antioxidative responses of Ocimum basilicum to sodium chloride or sodium sulphate salinization. Plant Physiol. Biochem. 2010, 48, 772–777. [Google Scholar] [CrossRef] [PubMed]
- Suzuki, N.; Koussevitzky, S.; Mittler, R.; Miller, G. ROS and redox signalling in the response of plants to abiotic stress. Plant Cell Environ. 2012, 35, 259–270. [Google Scholar] [CrossRef]
- Rice-Evans, C.A.; Miller, N.J.; Paganga, G. Structure-antioxidant activity relationships of flavonoids and phenolic acids. Free Rad. Biol. Med. 1996, 20, 933–956. [Google Scholar] [CrossRef]
- Rice-evans, C.A.; Miller, N.J.; Bolwell, P.G.; Bramley, P.M.; Pridham, J.B. The Relative Antioxidant Activities of Plant-Derived Polyphenolic Flavonoids. Free Radic. Res. 1995, 22, 375–383. [Google Scholar] [CrossRef]
- Ma, D.; Sun, D.; Wang, C.; Li, Y.; Guo, T. Expression of flavonoid biosynthesis genes and accumulation of flavonoid in wheat leaves in response to drought stress. Plant Physiol. Biochem. 2014, 80, 60–66. [Google Scholar] [CrossRef]
- Yang, C.; Zhang, C.; Zhang, J.; Gong, P.; Zhang, X.; Zhang, Y.; Khurram, Z.; Ye, Z.; Li, H.; Wang, T.; et al. Transcriptional profiles of drought-responsive genes in modulating transcription signal transduction, and biochemical pathways in tomato. J. Exp. Bot. 2010, 61, 3563–3575. [Google Scholar] [CrossRef]
- Stasolla, C.; Katahira, R.; Thorpe, T.A.; Ashihara, H. Purine and pyrimidine nucleotide metabolism in higher plants. J. Plant Physiol. 2003, 160, 1271–1295. [Google Scholar] [CrossRef]
- Watanabe, S.; Matsumoto, M.; Hakomori, Y.; Takagi, H.; Shimada, H.; Sakamoto, A. The purine metabolite allantoin enhances abiotic stress tolerance through synergistic activation of abscisic acid metabolism. Plant Cell Environ. 2014, 37, 1022–1036. [Google Scholar] [CrossRef] [Green Version]
- Watanabe, S.; Kounosu, Y.; Shimada, H.; Sakamoto, A. Arabidopsis Xanthine dehydrogenase mutants defective in purine degradation show a compromised protective response to drought and oxidative stress. Plant Biotechnol. 2014, 31, 173–178. [Google Scholar] [CrossRef] [Green Version]
- Das, A.; Rushton, P.J.; Rohila, J.S. Metabolomic Profiling of Soybeans (Glycine max L.) Reveals the Importance of Sugar and Nitrogen Metabolism under Drought and Heat Stress. Plants 2017, 6, 21. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, S.-C.; Jin, J.-Q.; Ma, J.-Q.; Yao, M.-Z.; Ma, C.-L.; Li, C.-F.; Ding, Z.-T.; Chen, L. Transcriptomic Analysis of Tea Plant Responding to Drought Stress and Recovery. PLoS ONE 2016, 11, e0147306. [Google Scholar] [CrossRef] [PubMed]
- Mibei, E.K.; Owino, W.O.; Ambuko, J.; Giovannoni, J.J.; Onyango, A.N. Metabolomic analyses to evaluate the effect of drought stress on selected African Eggplant accessions. J. Sci. Food Agric. 2018, 98, 205–216. [Google Scholar] [CrossRef] [PubMed]
- Lima, M.R.M.; Machado, A.F.; Gubler, W.D. Metabolomic Study of Chardonnay Grapevines Double Stressed with Esca-Associated Fungi and Drought. Phytopathology 2017, 107, 669–680. [Google Scholar] [CrossRef] [PubMed]
- Liang, X.; Zhang, L.; Natarajan, S.K.; Becker, D.F. Proline mechanisms of stress survival. Antioxid. Redox Signal. 2013, 19, 998–1011. [Google Scholar] [CrossRef] [Green Version]
- Hayat, S.; Hayat, Q.; Alyemeni, M.N.; Wani, A.S.; Pichtel, J.; Ahmad, A. Role of proline under changing environments: A review. Plant Signal. Behav. 2012, 7, 1456–1466. [Google Scholar] [CrossRef] [Green Version]
- Mwadzingeni, L.; Shimelis, H.; Tesfay, S.; Tsilo, T.J. Screening of bread wheat genotypes for drought tolerance using phenotypic and proline analyses. Front. Plant Sci. 2016, 7, 1276. [Google Scholar] [CrossRef] [Green Version]
- Anjum, S.A.; Farooq, M.; Xie, X.-y.; Liu, X.-j.; Ijaz, M.F. Antioxidant defense system and proline accumulation enables hot pepper to perform better under drought. Sci. Horticult. 2012, 140, 66–73. [Google Scholar] [CrossRef]
- Gill, S.S.; Anjum, N.A.; Hasanuzzaman, M.; Gill, R.; Trivedi, D.K.; Ahmad, I.; Pereira, E.; Tuteja, N. Glutathione and glutathione reductase: A boon in disguise for plant abiotic stress defense operations. Plant. Physiol. Biochem. 2013, 70, 204–212. [Google Scholar] [CrossRef]
- Jozefczak, M.; Remans, T.; Vangronsveld, J.; Cuypers, A. Glutathione is a key player in metal-induced oxidative stress defenses. Int. J. Mol. Sci. 2012, 13, 3145–3175. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bechtold, U.; Rabbani, N.; Mullineaux, P.M.; Thornalley, P.J. Quantitative measurement of specific biomarkers for protein oxidation, nitration and glycation in Arabidopsis leaves. Plant J. 2009, 59, 661–671. [Google Scholar] [CrossRef]
- Jing, H.; Kitts, D.D. Comparison of the antioxidative and cytotoxic properties of glucose-lysine and fructose-lysine Maillard reaction products. Food Res. Int. 2000, 33, 509–516. [Google Scholar] [CrossRef]
- Zhang, Y.; Ma, X.-M.; Wang, X.-C.; Liu, J.-H.; Huang, B.-Y.; Guo, X.-Y.; Xiong, S.-P.; La, G.-X. UPLC-QTOF analysis reveals metabolomic changes in the flag leaf of wheat (Triticum aestivum L.) under low-nitrogen stress. Plant Physiol. Biochem. 2017, 111, 30–38. [Google Scholar] [CrossRef] [PubMed]
- Ma, X.; Xin, Z.; Wang, Z.; Yang, Q.; Guo, S.; Guo, X.; Cao, L.; Lin, T. Identification and comparative analysis of differentially expressed miRNAs in leaves of two wheat (Triticum aestivum L.) genotypes during dehydration stress. BMC Plant Biol. 2015, 15, 21. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Barrs, H.; Weatherley, P. A re-examination of the relative turgidity technique for estimating water deficits in leaves. Aust. J. Biol. Sci. 1962, 15, 413–428. [Google Scholar] [CrossRef] [Green Version]
- Zhang, W.; Tan, B.; Ye, G.; Wang, J.; Dong, X.; Yang, Q.; Chi, S.; Liu, H.; Zhang, S.; Zhang, H. Identification of potential biomarkers for soybean meal-induced enteritis in juvenile pearl gentian grouper, Epinephelus lanceolatus♂ × Epinephelus fuscoguttatus♀. Aquaculture 2019, 512, 734337. [Google Scholar] [CrossRef]
- Luo, P.; Yin, P.; Hua, R.; Tan, Y.; Li, Z.; Qiu, G.; Yin, Z.; Xie, X.; Wang, X.; Chen, W.; et al. A Large-scale, multicenter serum metabolite biomarker identification study for the early detection of hepatocellular carcinoma. Hepatology 2018, 67, 662–675. [Google Scholar] [CrossRef]
- Liu, X.; Yang, Q.; Lu, Y.; Li, Y.; Li, T.; Zhou, B.; Qiao, L. Effect of purslane (Portulaca oleracea L.) extract on anti-browning of fresh-cut potato slices during storage. Food Chem. 2019, 283, 445–453. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Compared Samples | Num. of Total Ident | Num. of Total Sig | Num. of Sig. Up | Num. of Sig. Down |
|---|---|---|---|---|
| T1 vs. CK1_pos | 3028 | 527 | 449 | 78 |
| T2 vs. CK2_pos | 3028 | 499 | 397 | 102 |
| CK2 vs. CK1_pos | 3028 | 811 | 523 | 288 |
| T2 vs. T1_pos | 3028 | 797 | 425 | 372 |
| T1 vs. CK1_neg | 2544 | 464 | 366 | 98 |
| T2 vs. CK2_neg | 2544 | 429 | 318 | 111 |
| CK2 vs. CK1_neg | 2544 | 610 | 390 | 220 |
| T2 vs. T1_neg | 2544 | 584 | 326 | 258 |
| Ionization Mode | Classification | Metabolite | RT (min) | m/z | Formula | VIP | FC | p Value | |
|---|---|---|---|---|---|---|---|---|---|
| ESI (+) | 1 | Amino acid | L-Isoleucine | 1.648 | 131.0946 | C6 H13 N O2 | 1.51 | 0.33 | 1.49 × 10−7 |
| ESI (+) | 2 | Amino acid | L-Tyrosine | 1.413 | 181.0738 | C9 H11 N O3 | 1.40 | 0.37 | 1.12 × 10−5 |
| ESI (+) | 3 | Amino acid | L-Phenylalanine | 3.199 | 165.07893 | C9 H11 N O2 | 2.37 | 0.22 | 0.014226 |
| ESI (+) | 4 | Flavonoids | Dhurrin | 6.244 | 311.09967 | C14 H17 N O7 | 1.49 | 2.97 | 1.55 × 10−12 |
| ESI (+) | 5 | Flavonoids | Amygdalin | 8.234 | 457.15781 | C20 H27 N O11 | 2.36 | 0.18 | 8.67 × 10−10 |
| ESI (+) | 6 | Flavonoids | Luteolin | 8.624 | 286.04691 | C15 H10 O6 | 1.37 | 0.36 | 1.83 × 10−10 |
| ESI (+) | 7 | Flavonoids | Salidroside | 7.593 | 300.11993 | C14 H20 O7 | 3.30 | 0.09 | 1.41 × 10−7 |
| ESI (+) | 8 | Flavonoids | Quercetin 3-O-sophoroside | 7.941 | 626.14635 | C27 H30 O17 | 1.38 | 0.43 | 0.00944 |
| ESI (+) | 9 | Flavonoids | Arbutin | 6.052 | 272.08903 | C12 H16 O7 | 1.05 | 0.46 | 2.43 × 10−6 |
| ESI (+) | 10 | Flavonoids | Keracyanin | 10.441 | 595.1657 | C27 H31 O15 | 1.53 | 0.32 | 1.40 × 10−9 |
| ESI (+) | 11 | Alkaloids | N-Methyltryptamine | 5.986 | 174.1156 | C11 H14 N2 | 3.42 | 0.07 | 1.98 × 10−6 |
| ESI (+) | 12 | Alkaloids | Harmine | 9.559 | 212.09404 | C13 H12 N2 O | 2.21 | 0.20 | 4.11 × 10−12 |
| ESI (+) | 13 | Alkaloids | Tryptamine | 5.638 | 160.09996 | C10 H12 N2 | 3.98 | 0.05 | 1.02 × 10−8 |
| ESI (+) | 14 | Organic acids | Dihydroxyindole | 7.901 | 149.04764 | C8 H7 N O2 | 1.06 | 2.16 | 2.82 × 10−8 |
| ESI (+) | 15 | Alkaloids | Indole | 5.788 | 117.05778 | C8 H7 N | 1.01 | 0.48 | 1.32 × 10−9 |
| ESI (+) | 16 | Amino acid | L-Glutamic acid | 1.088 | 147.05307 | C5 H9 N O4 | 1.15 | 0.43 | 1.65 × 10−6 |
| ESI (+) | 17 | Phenolics | Vanillin | 2.477 | 152.04724 | C8 H8 O3 | 1.65 | 3.36 | 2.90 × 10−10 |
| ESI (+) | 18 | Phenolics | 3,4-Dihydroxybenzaldehyde | 6.419 | 138.03162 | C7 H6 O3 | 1.74 | 3.56 | 8.94 × 10−12 |
| ESI (+) | 19 | Alkaloids | Tyramine | 3.173 | 137.08408 | C8 H11 N O | 2.63 | 0.23 | 0.045753 |
| ESI (+) | 20 | Phenolics | 3-Dimethylallyl-4-hydroxymandelic acid | 8.564 | 236.1046 | C13 H16 O4 | 1.21 | 2.43 | 1.10 × 10−12 |
| ESI (+) | 21 | Alkaloids | Scopoletin | 8.437 | 192.04209 | C10 H8 O4 | 2.26 | 5.20 | 5.97 × 10−12 |
| ESI (+) | 22 | Phenolics | Sinapyl alcohol | 7.064 | 210.08902 | C11 H14 O4 | 1.30 | 0.39 | 2.92 × 10−8 |
| ESI (+) | 23 | Phenolics | Eugenol | 10.704 | 164.08361 | C10 H12 O2 | 1.02 | 0.46 | 1.13 × 10−5 |
| ESI (+) | 24 | Amino acid | L-Cysteinylglycine | 6.258 | 178.04111 | C5 H10 N2 O3 S | 1.05 | 2.16 | 4.38 × 10−6 |
| ESI (+) | 25 | Sugars | D-Ribulose 5-phosphate | 1.324 | 230.01883 | C5 H11 O8 P | 1.54 | 3.05 | 2.43 × 10−7 |
| ESI (+) | 26 | Phenolics | Eriodictyol | 10.112 | 288.0633 | C15 H12 O6 | 2.38 | 0.18 | 6.33 × 10−11 |
| ESI (+) | 27 | Others | Farnesyl pyrophosphate | 11.295 | 382.12948 | C15 H28 O7 P2 | 1.10 | 2.11 | 0.000214 |
| ESI (+) | 28 | Others | trans-Zeatin | 6.178 | 219.11171 | C10 H13 N5 O | 1.15 | 2.51 | 0.002331 |
| ESI (+) | 29 | Pyrimidine | Thymine | 4.325 | 126.04292 | C5 H6 N2 O2 | 3.57 | 13.72 | 4.45 × 10−15 |
| ESI (+) | 30 | Amino acid | Fructoselysine | 11.044 | 308.15916 | C12 H24 N2 O7 | 1.45 | 2.89 | 1.29 × 10−8 |
| ESI (+) | 31 | Phenolics | Syringic acid | 6.276 | 198.05268 | C9 H10 O5 | 1.55 | 0.32 | 6.83 × 10−11 |
| ESI (+) | 32 | Phenolics | alpha-Phocaecholic acid | 1.133 | 424.28289 | C24 H40 O6 | 1.74 | 3.63 | 8.48 × 10−9 |
| ESI (+) | 33 | Phenolics | Caffeic acid | 10.025 | 180.04209 | C9 H8 O4 | 1.28 | 0.42 | 0.00074 |
| ESI (+) | 34 | Phenolics | 1,4-Dihydroxy-2-naphthoic acid | 7.005 | 204.04184 | C11 H8 O4 | 1.18 | 0.42 | 2.88 × 10−8 |
| ESI (+) | 35 | Phenolics | Kynurenic acid | 8.003 | 189.04248 | C10 H7 N O3 | 1.91 | 3.98 | 5.60 × 10−8 |
| ESI (+) | 36 | Purine | Guanine | 1.934 | 151.04938 | C5 H5 N5 O | 1.11 | 2.34 | 0.000149 |
| ESI (-) | 37 | Organic acids | L-Proline | 1.165 | 115.0632 | C5 H9 N O2 | 2.33 | 0.16 | 2.24 × 10−8 |
| ESI (-) | 38 | Organic acids | Citric acid | 7.876 | 192.027 | C6 H8 O7 | 1.03 | 2.31 | 1.13 × 10−6 |
| ESI (-) | 39 | Organic acids | Fumaric acid | 1.029 | 116.0109 | C4 H4 O4 | 2.11 | 0.20 | 2.39 × 10−6 |
| ESI (-) | 40 | Organic acids | trans-Cinnamic acid | 2.965 | 148.0523 | C9 H8 O2 | 1.45 | 0.31 | 2.96 × 10−9 |
| ESI (-) | 41 | Phenolics | Ascorbic acid | 1.118 | 176.032 | C6 H8 O6 | 2.30 | 0.16 | 9.97 × 10−8 |
| ESI (-) | 42 | Others | Reduced Glutathione | 1.066 | 307.0832 | C10 H17 N3 O6 S | 1.56 | 0.28 | 1.62 × 10−9 |
| ESI (-) | 43 | Phenolics | Homovanillic acid | 6.58 | 182.0578 | C9 H10 O4 | 1.10 | 2.42 | 2.62 × 10−11 |
| ESI (-) | 44 | Phenolics | Gallic acid | 2.486 | 170.0215 | C7 H6 O5 | 1.42 | 3.15 | 1.87E × 10−8 |
| ESI (-) | 45 | Phenolics | Coniferyl aldehyde | 8.86 | 178.0628 | C10 H10 O3 | 1.57 | 3.55 | 2.55 × 10−9 |
| ESI (-) | 46 | Amino acid metabolic intermediates | Saccharopine | 0.965 | 276.1314 | C11 H20 N2 O6 | 1.23 | 0.39 | 3.55 × 10−5 |
| ESI (-) | 47 | Phenolics | Gentisic acid | 6.882 | 154.0265 | C7 H6 O4 | 2.80 | 0.10 | 1.87 × 10−9 |
| ESI (-) | 48 | Phenolics | Vanillyl alcohol | 5.27 | 154.0629 | C8 H10 O3 | 1.54 | 3.62 | 2.76 × 10−5 |
| ESI (-) | 49 | Phenolics | 4-Nitrophenol | 8.546 | 139.0268 | C6 H5 N O3 | 1.20 | 2.63 | 2.73 × 10−9 |
| ESI (-) | 50 | Phenolics | Pyridoxal | 1.622 | 167.058 | C8 H9 N O3 | 1.11 | 2.44 | 1.41 × 10−8 |
| ESI (-) | 51 | Flavonoids | Pumiloside | 8.868 | 512.1788 | C26 H28 N2 O9 | 1.29 | 0.37 | 1.59 × 10−5 |
| ESI (-) | 52 | Flavonoids | Genistein | 11.175 | 270.0521 | C15 H10 O5 | 1.15 | 2.66 | 0.000446 |
| ESI (-) | 53 | Flavonoids | Astragalin | 8.489 | 448.1001 | C21 H20 O11 | 1.53 | 0.31 | 2.36 × 10−5 |
| ESI (-) | 54 | Flavonoids | Quercitrin | 7.573 | 448.0997 | C21 H20 O11 | 1.89 | 0.22 | 1.35 × 10−9 |
| ESI (-) | 55 | Flavonoids | Quercetin 3-O-(beta-D-xylosyl-(1-> 2)-beta-D-glucoside) | 6.96 | 596.1377 | C26 H28 O16 | 1.79 | 0.24 | 2.16 × 10−10 |
| ESI (-) | 56 | Phenolics | Picolinic acid | 7.886 | 123.032 | C6 H5 N O2 | 1.17 | 2.55 | 5.31 × 10−11 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Guo, X.; Xin, Z.; Yang, T.; Ma, X.; Zhang, Y.; Wang, Z.; Ren, Y.; Lin, T. Metabolomics Response for Drought Stress Tolerance in Chinese Wheat Genotypes (Triticum aestivum). Plants 2020, 9, 520. https://0-doi-org.brum.beds.ac.uk/10.3390/plants9040520
Guo X, Xin Z, Yang T, Ma X, Zhang Y, Wang Z, Ren Y, Lin T. Metabolomics Response for Drought Stress Tolerance in Chinese Wheat Genotypes (Triticum aestivum). Plants. 2020; 9(4):520. https://0-doi-org.brum.beds.ac.uk/10.3390/plants9040520
Chicago/Turabian StyleGuo, Xiaoyang, Zeyu Xin, Tiegang Yang, Xingli Ma, Yang Zhang, Zhiqiang Wang, Yongzhe Ren, and Tongbao Lin. 2020. "Metabolomics Response for Drought Stress Tolerance in Chinese Wheat Genotypes (Triticum aestivum)" Plants 9, no. 4: 520. https://0-doi-org.brum.beds.ac.uk/10.3390/plants9040520