Optimization of Macroalgal Density and Salinity for Nutrient Removal by Caulerpa lentillifera from Aquaculture Effluent

 ,
,  and
and 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Fish Collection and Acclimatization for Experiment
2.2. Macroalgae Collection and Pre-Culture
2.3. Application of Macroalgal Density
2.4. Application of Salinity and Aeration
2.5. Measurement of Nutrient Uptake
2.6. Inferential Analysis
2.7. Regression Analysis
3. Results
3.1. Macroalgal Density
3.1.1. Relationship between Nutrient Concentration and Algal Density
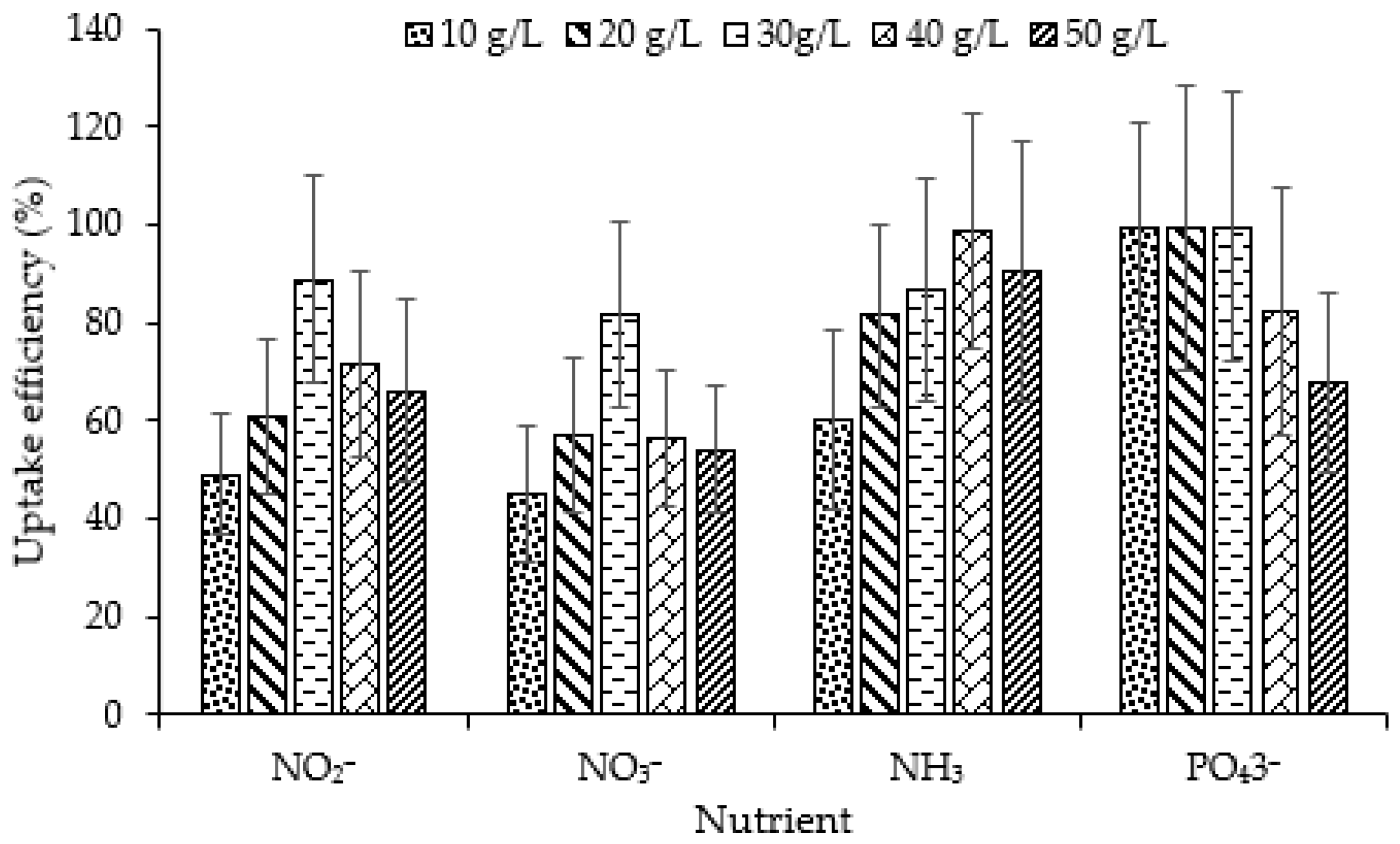
3.1.2. Relationship between Nutrient Uptake Efficiency and Macroalgal Density
3.2. Salinity and Aeration
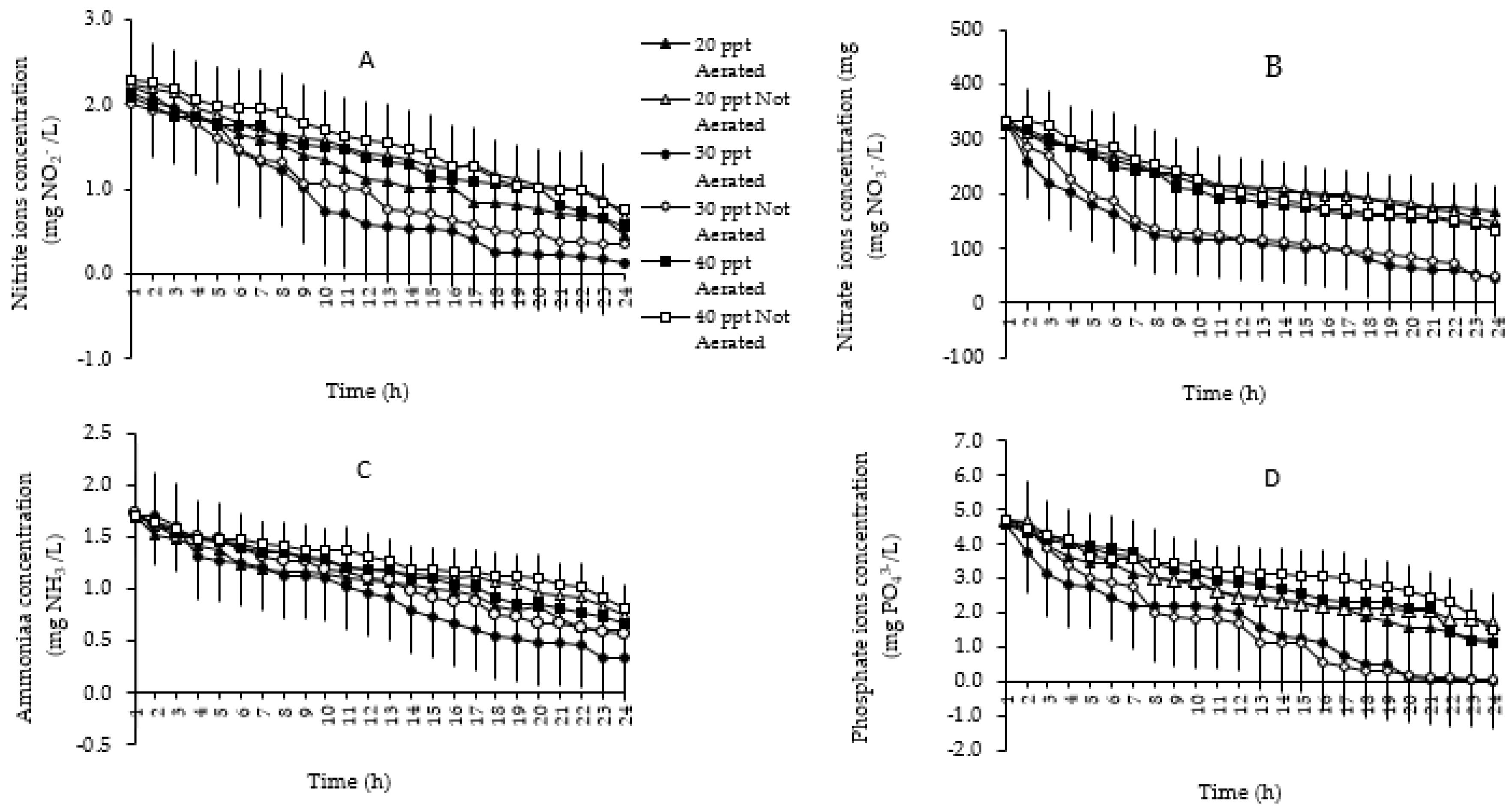
3.2.1. Relationship between Nutrient Concentration and Salinity and Aeration
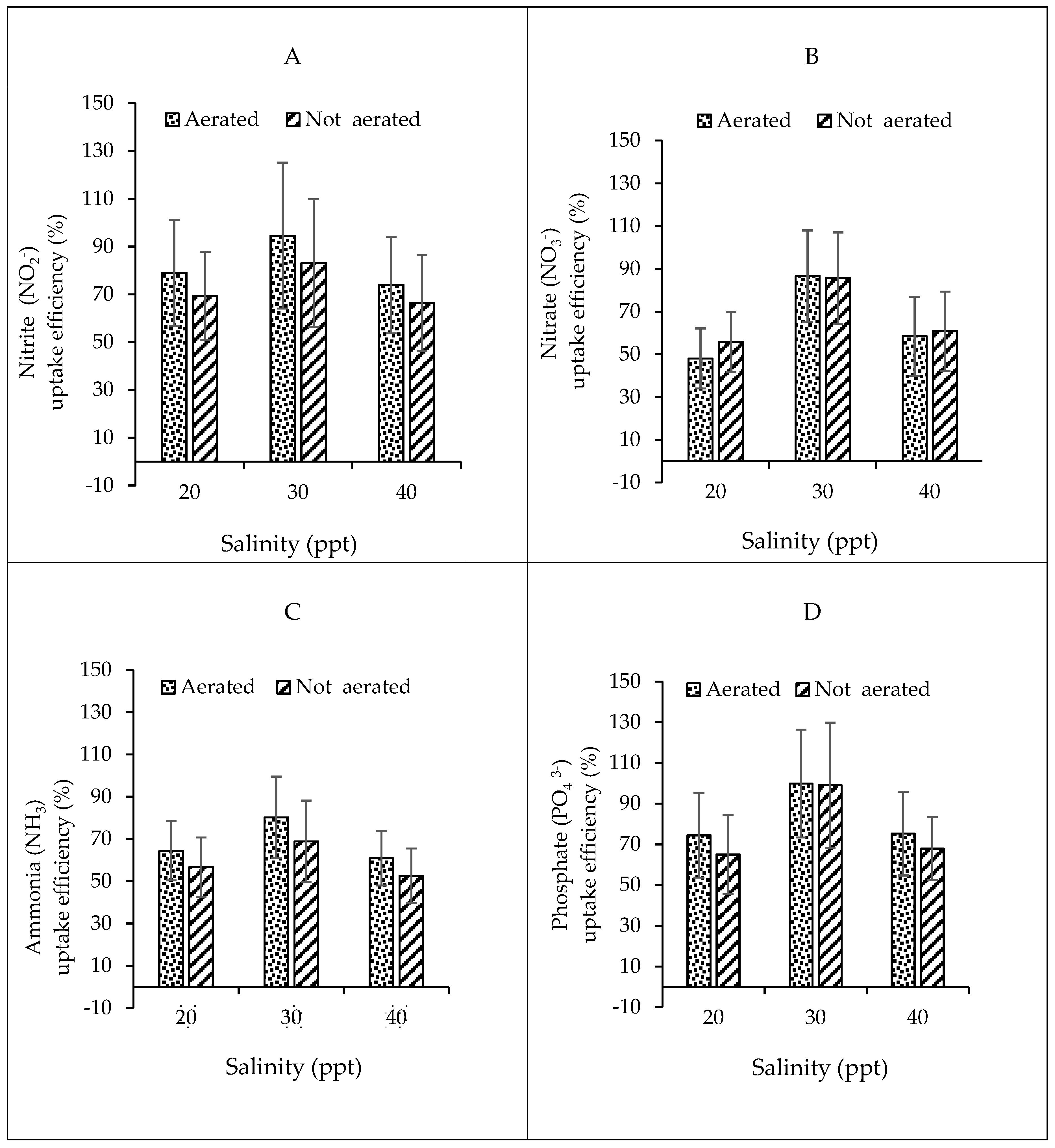
3.2.2. Relationship between Nutrient Uptake Efficiency and Salinity and Aeration
3.2.3. Relationship between Photosynthesis affected by Salinity and Aeration
3.2.4. Effects of Various Factors on Nutrient Uptake Efficiency
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Chung, I.K. The development of environmentally sound integrated polyculture system. Min. Sci. Tech. Project #M6-0203-00-0041-02-A01-00-021-00. Cultivation of Gracilaria parvispora (Rhodophyta) in shrimp-farm effluent ditches and floating cages in Hawaii: A two-phase polyculture system. Aquaculture 2002, 193, 239–248. [Google Scholar]
- Turcios, A.E.; Papenbrock, J. Sustainable treatment of aquaculture effluents-What can we learn from the past for the future? Sustainability 2014, 6, 836–856. [Google Scholar] [CrossRef]
- Chopin, T.; Buschmann, A.H.; Halling, C.; Troell, M.; Kautsky, N.; Neori, A. Integrating seaweeds into marine aquaculture systems. A key toward sustainability. J. Phycol. 2001, 37, 975–986. [Google Scholar] [CrossRef]
- Boyd, C.E.; Tucker, C.; Mcnevin, A.; Bostick, A.; Katherine, B.; Clay, J. Indicators of Resource Use Efficiency and Environmental Performance in Fish and Crustacean Aquaculture. Rev. Fish. Sci. 2007, 15, 327–360. [Google Scholar] [CrossRef]
- Khan, F.A.; Ansari, A.A. Eutrophication: An ecological vision. Bot. Rev. 2007, 71, 449–482. [Google Scholar] [CrossRef]
- Chopin, T.; Cooper, J.A.; Reid, G.; Cross, S.; Moore, C. Open-water integrated multitrophic aquaculture: Environmental biomitigation and economic diversification of fed aquaculture by extractive aquaculture. Rev. Aquac. 2012, 4, 209–220. [Google Scholar] [CrossRef]
- Phang, S.; Chu, W.; Rabiei, R. Phycoremediation. In The Algae World; Sahoo, D., Seckbach, J., Eds.; Springer: Esrael, Israel, 2015; Volume 26, pp. 357–381. [Google Scholar]
- Haseena, M.; Malik, M.F.; Javed, A.; Arshad, S.; Asif, N.; Zulfiqar, S.; Hanif, J. Water pollution and human health. J. Environ. Risk Assess. Remediat. 2017, 3, 16–19. [Google Scholar] [CrossRef]
- Mugg, J.; Serrano, A.; Liberti, A.; Rice, M.A. Aquaculture Effluents: A Guide for Water Quality Regulators and Aquaculturists; NRCA Publication, University of Massachusetts Dartmouth: North Dartmouth, MA, USA, 2000. [Google Scholar]
- Boyd, C.E. Guidelines for aquaculture effluent management at the farm-level. Aquaculture 2003, 226, 101–112. [Google Scholar] [CrossRef]
- Rijn, J.V. The potential for integrated biological treatment systems in recirculating fish culture—A review. Aquaculture 1996, 139, 181–201. [Google Scholar] [CrossRef]
- Jones, A.B.; Dennison, W.C.; Preston, N.P. Integrated treatment of shrimp effluent by sedimentation, oyster filtration and macroalgal absorption: A laboratory scale study. Aquaculture 2001, 193, 155–178. [Google Scholar] [CrossRef]
- Devi, I.R.P.; Gowri, V.S. Biological Treatment of Aquaculture Discharge Waters by Seaweeds. J. Ind. Pollut. Control 2007, 23, 135–140. [Google Scholar]
- Lobban, C.S.; Harrison, P.J. Seaweed Ecology and Physiology; Cambridge University Press: Cambridge, UK, 1994; pp. 163–240. [Google Scholar]
- Paul, N.A.; De Nys, R. Promise and pitfalls of locally abundant sea-weeds as biofilters for integrated aquaculture. Aquaculture 2008, 281, 49–55. [Google Scholar] [CrossRef]
- Guo, H.; Yao, J.; Sun, Z.; Duan, D. Effects of salinity and nutrients on the growth and chlorophyll fluorescence of Caulerpa lentillifera. Chin. J. Oceanol. Limnol. 2015, 33, 410–418. [Google Scholar] [CrossRef]
- Liu, H.; Wang, F.; Wang, Q.; Dong, S.; Tian, X. A comparative study of the nutrient uptake and growth capacities of seaweeds Caulerpa lentillifera and Gracilaria lichenoides. J. Appl. Phycol. 2016, 28, 3083–3089. [Google Scholar] [CrossRef]
- Manríquez-Hernández, J.A.; Duston, J.; Garbary, D.J. Effect of irradiance on bioremediation capacity of the red algae Chondrus crispus and Palmaria palmata. Aquac. Int. 2016, 24, 39–55. [Google Scholar] [CrossRef]
- Wedchaparn, O.; Ayisi, C.L.; Huo, Y.; He, P. Effects of different light intensity fluctuations on growth Rate, Nutrient Uptake and Photosynthetic Efficiency of Gracilaria asiatica. J. Fish. Aquat. Sci. 2015, 10, 533–542. [Google Scholar]
- Manríquez-Hernández, J.A. Interaction of Irradiance and Stocking Density on Nutrient Uptake by Red Macroalgae. Implications for Bioremediation of Fish Farm Effluents. Master’s Thesis, Dalhousie University Halifax, Halifax, NS, Canada, 2013. [Google Scholar]
- Barr, N.G.; Kloeppel, A.; Rees, T.A.V.; Scherer, C.; Taylor, R.B.; Wenzel, A. Wave surge increases rates of growth and nutrient uptake in the green seaweed Ulva pertusa maintained at low bulk flow velocities. Aquat. Biol. 2008, 3, 179–186. [Google Scholar] [CrossRef]
- Caines, S.; Manríquez-Hernández, J.A.; Duston, J.; Corey, P.; Garbary, D.J. Intermittent aeration affects the bioremediation potential of two red algae cultured in finfish effluent. J. Appl. Phycol. 2014, 26, 2173–2181. [Google Scholar] [CrossRef]
- Msuya, F.E.; Neori, A. Effect of water aeration and nutrient load level on biomass yield, N uptake and protein content of the seaweed Ulva lactuca cultured in seawater tanks. J. Appl. Phycol. 2008, 20, 1021–1031. [Google Scholar] [CrossRef]
- Lapointe, B.E.; Tenore, K.R. Experimental outdoor studies with Ulva fasciata Delile. I. Interaction of light and nitrogen on nutrient uptake, growth, and biochemical composition. J. Exp. Mar. Biol. Ecol. 1981, 53, 135–152. [Google Scholar] [CrossRef]
- Fong, P.; Boyer, K.E.; Desmond, J.S.; Zedler, J.B. Salinity stress, nitrogen competition, and facilitation: What controls seasonal succession of two opportunistic green macroalgae? J. Exp. Mar. Biol. Ecol. 1996, 206, 203–221. [Google Scholar] [CrossRef]
- Choi, T.S.; Kang, E.J.; Kim, J.H.; Kim, K.Y. Effect of salinity on growth and nutrient uptake of Ulva pertusa (Chlorophyta) from an eelgrass bed. Algae 2010, 25, 17–26. [Google Scholar] [CrossRef]
- Msuya, F. Effects of stocking density and additional nutrients on growth of the commercially farmed seaweeds Eucheuma denticulatum and Kappaphycus alvarezii in Zanzibar Tanzania. TaJONAS 2013, 4, 605–612. [Google Scholar]
- Macchiavello, J.; Bulboa, C. Nutrient uptake efficiency of Gracilaria chilensis and Ulva lactuca in an IMTA system with the red abalone Haliotis rufescens. Lat. Am. J. Aquat. Res. 2014, 42, 523–533. [Google Scholar] [CrossRef]
- Madsen, J.D.; Chambers, P.A.; James, W.F.; Koch, E.W.; Westlake, D.F. The interaction between water movement, sediment dynamics and submersed macrophytes. Hydrobiologia 2001, 444, 71–84. [Google Scholar] [CrossRef]
- Ratana-Arporn, P.; Chirapart, A. Nutritional Evaluation of Tropical Green Seaweeds Caulerpa lentillifera and Ulva reticulata. Kasetsart J. (Nat. Sci.) 2006, 181, 97–106. [Google Scholar]
- Paul, N.A.; Neveux, N.; Magnusson, M.; de Nys, R. Comparative production and nutritional value of “sea grapes” the tropical green seaweeds Caulerpa lentillifera and C. racemosa. J. Appl. Phycol. 2014, 26, 1833–1844. [Google Scholar] [CrossRef]
- Rabia, M.D.S. Cultivation of Caulerpa lentillifera Using Tray and Sowing Methods in Brackishwater Pond. Environ. Sci. 2016, 4, 23–29. [Google Scholar] [CrossRef]
- McHugh, D.J. A Guide to the Seaweed Industry; Food and Agriculture Organization of United Nations: Rome, Italy, 2003. [Google Scholar]
- Chokwiwattanawanit, A. Efficiency of the Macroalgae Caulerpa lentillifera and Acanthophora spicifera for the Treatment of Nitrogen Compound from Shrimp Pond Effluent. Master’s Thesis, Environmental Science, Graduate School, Chulalongkorn University, Bangkok, Thailand, 2000. [Google Scholar]
- Pariyawathee, S.; Songsangjinda, P.; Deraxbudsarakom, S.; Tuntichodok, P.; Chiayvareesajja, S. Optimum condition of environmental factors for growth of sea grape (Caulerpa lentillifera: J. Agardh). Thai Fish Gaz 2003, 56, 443–448. (In Thai) [Google Scholar]
- Ghosh, A.; Mahapatra, B.K.; Datta, N.C. Ornamental fish farming - successful small-scale business in India. Aquacult. Asia 2003, 8, 14–16. [Google Scholar]
- Gonzalez, R.J.; Cooper, J.; Head, D. Physiological responses to hyper-saline waters in sailfin mollies (Poecilia latipinna). Comp. Biochem. Physiol. Mol. Integr. Physiol. 2005, 142, 397–403. [Google Scholar] [CrossRef] [PubMed]
- Nordlie, F.G.; Haney, D.C.; Walsh, S.J. Comparisons of salinity tolerances and osmotic regulatory capabilities in populations of sailfin molly (Poecilia latipinna) from brackish and fresh waters. Copeia 1992, 741–746. [Google Scholar] [CrossRef]
- Chen, W.Y.C.K.T. Salinity-dependent expression of the branchial Na + / K + / 2Cl2 cotransporter and Na + / K + ATPase in the sailfin molly correlates with hypoosmoregulatory endurance. J. Comp. Physiol. B 2011, 181, 953–964. [Google Scholar]
- APHA. Standard Methods for the Examination of Water and Wastewater, 21st ed.; American Public Health Association/American Water Works Association/Water Environment Federation: Washington, DC, USA, 2005. [Google Scholar]
- The Ministry of Natural Resources and Environment, Thailand. Coastal Aquaculture Effluent Standard in Thailand. B.E. 2004, 121, 2547. [Google Scholar]
- Kang, Y.H.; Shin, J.A.; Kim, M.S.; Chung, I.K. A preliminary study of the bioremediation potential of Codium fragile applied to seaweed integrated multi-trophic aquaculture (IMTA) during the summer. J. Appl. Phycol. 2008, 20, 183–190. [Google Scholar] [CrossRef]
- Kang, Y.H.; Park, S.R.; Chung, I.K. Biofiltration efficiency and biochemical composition of three seaweed species cultivated in a fish-seaweed integrated culture. Algae 2011, 26, 97–108. [Google Scholar]
- Tremblay-Gratton, A.; Boussin, J.C.; Tamigneaux, É.; Vandenberg, G.W.; Le François, N.R. Bioremediation efficiency of Palmaria palmata and Ulva lactuca for use in a fully recirculated cold-seawater naturalistic exhibit: Effect of high NO3 and PO4 concentrations and temperature on growth and nutrient uptake. J. Appl. Phycol. 2018, 30, 1295–1304. [Google Scholar] [CrossRef]
- Tsusaka, T.W.; Msere, H.W.; Gondwe, L.; Madzonga, O.; Clarke, S.; Siambi, M. Assessing the Post-harvest Constraints in Smallholders’ Groundnut Production: A Survey in Central Malawi. Res. J. Agric. Sci. 2016, 6, 213–226. [Google Scholar]
- Tsusaka, T.W.; Otsuka, K. The changing effects of agro-climate on cereal crop yields during the green revolution in India, 1972 to 2002. J. Sustain. Dev. 2013, 6, 11–36. [Google Scholar] [CrossRef]
- Tsusaka, T.W.; Otsuka, K. The changes in the effects of temperature and rainfall on cereal crop yields in sub-saharan africa, a country level panel data study, 1989 to 2004. Environ. Econ. 2013, 4, 70–80. [Google Scholar]
- Seetha, A.; Tsusaka, T.W.; Munthali, T.W.; Musukwa, M.; Mwangwela, A.; Kalumikiza, Z.; Manani, T.; Kachulu, L.; Kumwenda, N.; Musoke, M.; et al. How immediate and significant is the outcome of training on diversified diets, hygiene and food safety? An effort to mitigate child undernutrition in rural Malawi. Public Health Nutr. 2018, 21, 1156–1166. [Google Scholar] [CrossRef] [Green Version]
- Vossler, C.A. Panel Data with Serial Correlation (38862) 2012. Available online: https://mpra.ub.uni-muenchen.de/38862/1/MPRA_paper_38862.pdf (accessed on 15 June 2018).
- StataCorp. Stata: Release 15. Statistical Software; StataCorp LLC: College Station, TX, USA, 2017. [Google Scholar]
- Matos, J.; Costa, S.; Rodrigues, A.; Pereira, R.; Sousa Pinto, I. Experimental integrated aquaculture of fish and red seaweeds in Northern Portugal. Aquaculture 2006, 252, 31–42. [Google Scholar] [CrossRef]
- Prathumchai, N.; Polprasert, C.; Englande, A.J. Phosphorus leakage from fisheries sector. A case study in Thailand. Environ. Pollut. 2016, 219, 967–975. [Google Scholar] [CrossRef]
- UN-Water, Consolidated Technical Input from UN Agencies on Water and Sanitation Related Indicators. 2015. Available online: https://sustainabledevelopment.un.org/content/documents/2076Consolidated%20Technical%20inputs.pdf (accessed on 21 April 2019).





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Nutrient | Macroalgal Density | Salinity | Aeration | |||||||
|---|---|---|---|---|---|---|---|---|---|---|
| 20 ppt Salinity | 30 ppt Salinity | 40 ppt Salinity | ||||||||
| F Statistic (df, df) | p Value | F Statistic (df, df) | p Value | t Statistic (df) | p Value | t Statistic (df) | p Value | t Statistic (df) | p Value | |
| NO2− | 49.445 (4, 115) | 0.000 *** | 4.714 (2, 69) | 0.000 *** | 2.625 (46) | 0.011 ** | 1.082 (46) | 0.361 | 0.551 (46) | 0.989 |
| NO3− | 34.431 (4, 115) | 0.000 *** | 1.919 (2, 69) | 0.000 *** | −0.655 (46) | 0.893 | 0.633 (46) | 0.608 | 0.353 (46) | 0.591 |
| NH3− | 7.453 (4, 115) | 0.000 *** | 5.738 (2, 69) | 0.000 *** | 1.772 (46) | 0.511 | 0.851 (46) | 0.245 | 1.273 (46) | 0.129 |
| PO43− | 49.445 (4, 115) | 0.000 *** | 12.484 (2, 69) | 0.000 *** | 0.693 (46) | 0.941 | −0.020 (46) | 0.483 | 0.892 (46) | 0.087* |
| Macroalgal Density | Mean Difference (p Value) | ||||
|---|---|---|---|---|---|
| NO2− | NO3− | NH3 | PO43− | ||
| 10 g/L | 20 g/L | −12.623 * | −11.930 ** | −6.697 | −0.441 |
| (0.052) | (0.040) | (0.725) | (1.000) | ||
| 30 g/L | −50.237 *** | −55.618 *** | −36.917 *** | −0.506 | |
| (0.000) | (0.000) | (0.000) | (1.000) | ||
| 40 g/L | −30.209 *** | −20.047 *** | −69.377 *** | 48.751 *** | |
| (0.000) | (0.000) | (0.000) | (0.000) | ||
| 50 g/L | −19.542 *** | −18.030 *** | −26.623 *** | 84.751 *** | |
| (0.000) | (0.000) | (0.000) | (0.000) | ||
| 20 g/L | 30 g/L | −37.613 *** | −43.687 *** | −29.949 *** | −0.065 |
| (0.000) | (0.000) | (0.000) | (1.000) | ||
| 40 g/L | −17.585 *** | −8.116 | −62.409*** | 49.192 *** | |
| (0.002) | (0.302) | (0.000) | (0.000) | ||
| 50 g/L | −6.918 | −6.099 | −19.655 *** | 85.192 *** | |
| (0.557) | (0.592) | (0.006) | (0.000) | ||
| 30 g/L | 40 g/L | 20.028 *** | 35.571 *** | −32.460*** | 49.258 *** |
| (0.000) | (0.000) | (0.000) | (0.000) | ||
| 50 g/L | 30.695 *** | 37.588 *** | 10.2930 | 85.258 *** | |
| (0.000) | (0.000) | (0.356) | (0.000) | ||
| 40 g/L | 50 g/L | 10.666 (0.142) | 2.016 (0.989) | 42.754 *** (0.000) | 35.999 *** (0.000) |
| Salinity (ppt) | Mean Difference in Nutrient Uptake Efficiency (NUE) (%) (p Value) | ||||
|---|---|---|---|---|---|
| NO2− | NO3− | NH3 | PO43− | ||
| 20 | 30 | −21.252 *** | −24.943 *** | −10.854 * | −24.157 *** |
| (0.003) | (0.000) | (0.056) | (0.002) | ||
| 40 | −3.100 | −0.773 | 4.364 | 8.269 | |
| (0.871) | (0.989) | (0.615) | (0.442) | ||
| 30 | 40 | 18.152 ** | 24.170 *** | 15.218 *** | 32.427 *** |
| (0.012) | (0.000) | (0.004) | (0.000) | ||
| Explanatory Variable | Estimated Coefficients (p-Value) | |||||||
|---|---|---|---|---|---|---|---|---|
| NO2− | NO3− | NH3 | PO43− | |||||
| Random Effect | OLS Robust | Random Effect | OLS Robust | Random Effect | OLS Robust | Random Effect | OLS Robust | |
| Macroalgal Density (g/L) | 3.7873 *** (0.000) | 3.7690 *** (0.000) | 3.9996 *** (0.000) | 4.1216 *** (0.000) | 1.5823 ** (0.024) | 1.7576 ** (0.020) | 1.6914 *** (0.008) | 1.6931 *** (0.023) |
| Macroalgal Density2 | −0.0590 *** (0.000) | −0.0587 *** (0.000) | −0.0616 *** (0.000) | −0.0636 *** (0.000) | −0.0143 (0.214) | −0.0172 (0.135) | −0.0422 *** (0.000) | −0.0422 *** (0.002) |
| Salinity (ppt) | 9.4015 *** (0.000) | 9.4348 *** (0.003) | 11.1777 *** (0.000) | 11.9808 *** (0.001) | 14.6871 *** (0.000) | 11.9196 *** (0.000) | 18.3915 *** (0.000) | 18.3613 *** (0.000) |
| Salinity2 | −0.1616 *** (0.000) | −0.1620 *** (0.003) | −0.1778 *** (0.000) | −0.1953 *** (0.001) | −0.2487 *** (0.006) | −0.2031 *** (0.000) | −0.3119 *** (0.000) | −0.3114 *** (0.000) |
| Aeration (1 = aerated, 0 = not aerated) | 5.8334 (0.147) | 6.2539 (0.140) | 9.2832 * (0.067) | 6.4778 (0.121) | 8.1866 (0.168) | 4.1534 (0.259) | 0.9241 (0.780) | 0.8847 (0.795) |
| Constant | −1.4759 *** (0.000) | −1.4850 *** (0.003) | −1.8919 *** (0.000) | −1.9868 *** (0.001) | −2.0492 *** (0.000) | −1.6456 *** (0.001) | −2.1547 *** (0.000) | −2.1498 *** (0.000) |
| Num. of observations | 257 | 257 | 268 | 268 | 270 | 270 | 261 | 261 |
| Goodness of fit | Wald = 65.64 *** (p = 0.000) | F-statistic = 12.37 *** (p = 0.005) R2 = 0.2148 | Wald = 65.64 *** (p = 0.000) | F-statistic = 11.19 *** (p = 0.0005) R2 = 0.2597 | Wald = 31.48 *** (p = 0.000) | F-statistic = 9.30 *** (p = 0.008) R2 = 0.2258 | Wald = 1648.73 *** (p = 0.000) | F-statistic = 325.89 *** (p = 0.000) R2 = 0.2874 |
| Parameters | Random Effect | OLS Robust * | ||||||
|---|---|---|---|---|---|---|---|---|
| NO2− | NO3− | NH3 | PO43− | NO2− | NO3− | NH3 | PO43− | |
| Critical values of Macroalgal density (g/L) | 31.58 | 33.25 | 50.00 1 | 20.03 | 31.42 | 34.33 | 51.12 2 | 20.04 |
| Critical values of Salinity (ppt) | 29.09 | 30.68 | 29.53 | 29.48 | 29.11 | 30.68 | 29.35 | 29.48 |
| Predicted NUE (%) | 56.10 | 60.33 | 98.16 | 75.05 | 56.38 | 67.23 | 51.17 3 | 74.66 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bambaranda, B.V.A.S.M.; Sasaki, N.; Chirapart, A.; Salin, K.R.; Tsusaka, T.W. Optimization of Macroalgal Density and Salinity for Nutrient Removal by Caulerpa lentillifera from Aquaculture Effluent. Processes 2019, 7, 303. https://0-doi-org.brum.beds.ac.uk/10.3390/pr7050303
Bambaranda BVASM, Sasaki N, Chirapart A, Salin KR, Tsusaka TW. Optimization of Macroalgal Density and Salinity for Nutrient Removal by Caulerpa lentillifera from Aquaculture Effluent. Processes. 2019; 7(5):303. https://0-doi-org.brum.beds.ac.uk/10.3390/pr7050303
Chicago/Turabian StyleBambaranda, B.V.A.S. Manori, Nophea Sasaki, Anong Chirapart, Krishna R. Salin, and Takuji W. Tsusaka. 2019. "Optimization of Macroalgal Density and Salinity for Nutrient Removal by Caulerpa lentillifera from Aquaculture Effluent" Processes 7, no. 5: 303. https://0-doi-org.brum.beds.ac.uk/10.3390/pr7050303