Comparative Study of Angiotensin I-Converting Enzyme (ACE) Inhibition of Soy Foods as Affected by Processing Methods and Protein Isolation

Abstract
:1. Introduction
2. Materials and Methods
2.1. Soybean Material and Chemicals
2.2. Preparation of Soy Foods
2.2.1. Soymilk
2.2.2. Tofu
2.2.3. Natto
2.2.4. Tempeh
2.2.5. Soy Yogurt
2.2.6. Soybean Sprout
2.3. Simulated In Vitro Digestion of Soy Foods
2.4. Characterization of Digested Soy Foods
2.5. Preparation and Hydrolysis of 7S and 11S Fractions
2.6. Determination of Angiotensin Converting Enzyme (ACE) Inhibitory Activity
2.7. Determination of Peptide Molecular Weight Distribution
2.8. Determination of Soluble Proteins
2.9. Statistical Analysis
3. Results and Discussion
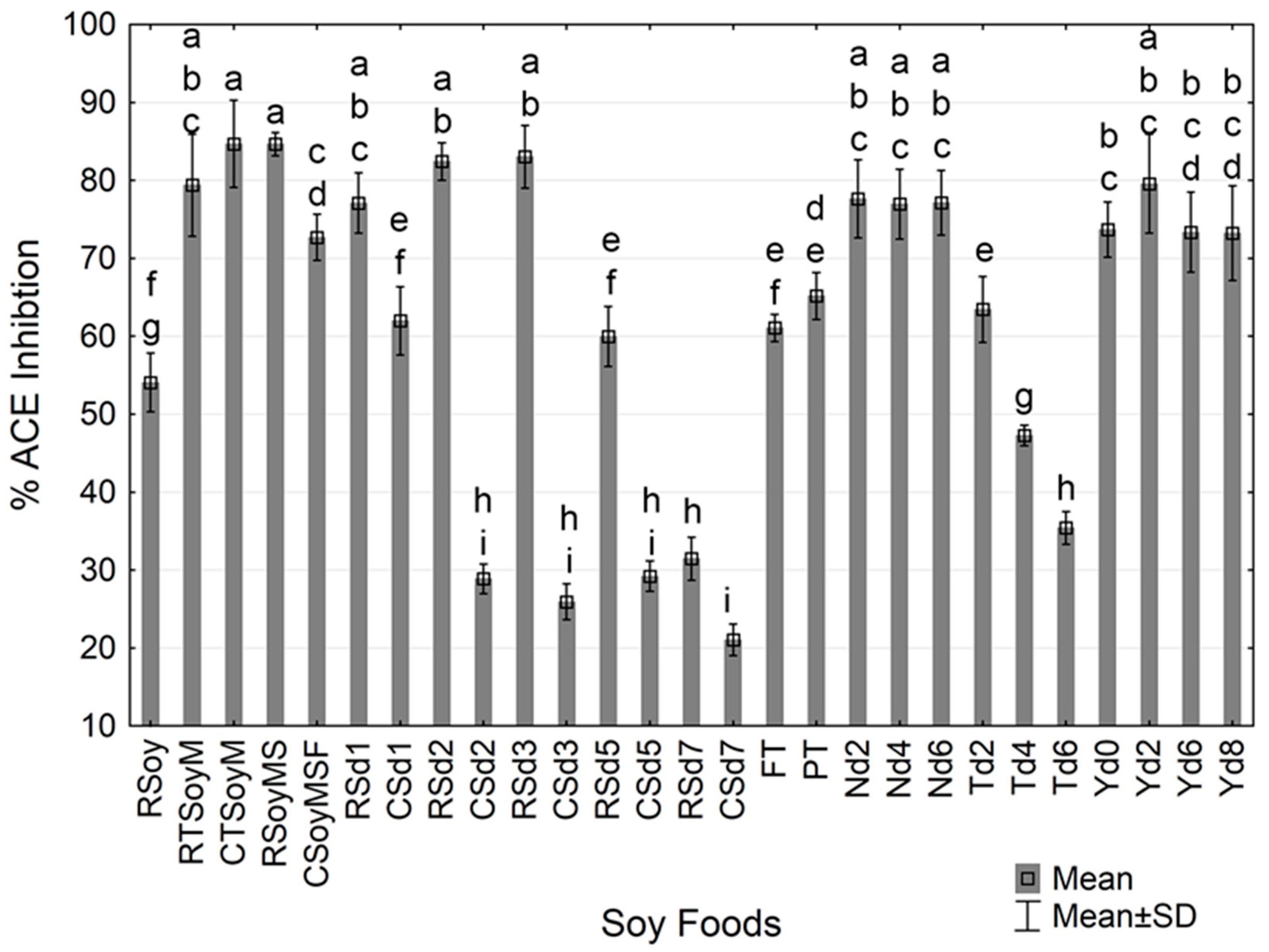
3.1. Angiotensin Converting Enzyme (ACE) Inhibitory Activity of Digested Fermented and Non-Fermented Soy Foods
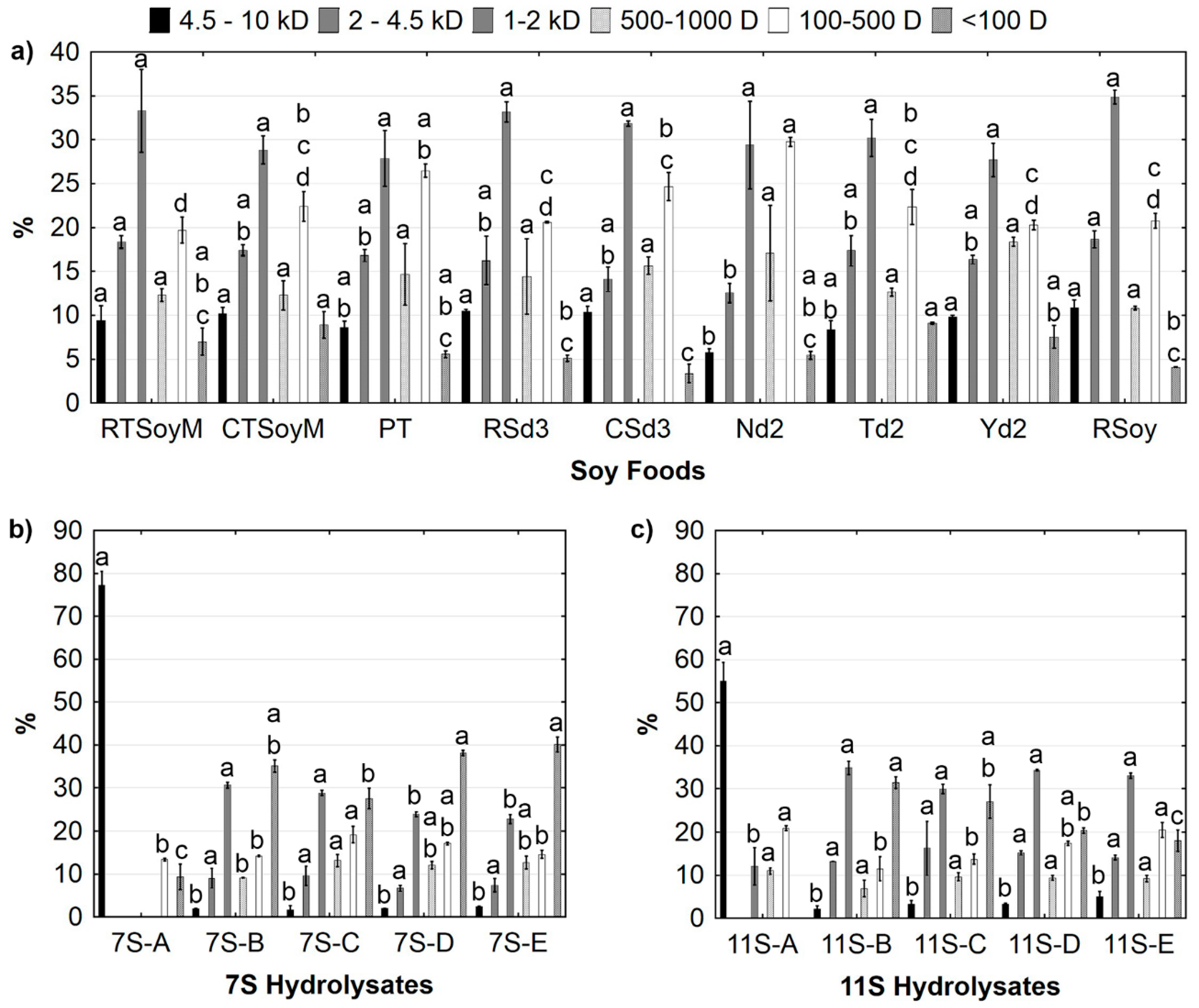
3.2. Characterization of Digested Soy Foods
3.3. Angiotensin Converting Enzyme (ACE) Inhibitory Capacity and Molecular Weight Distribution of 7S and 11S Hydrolysates
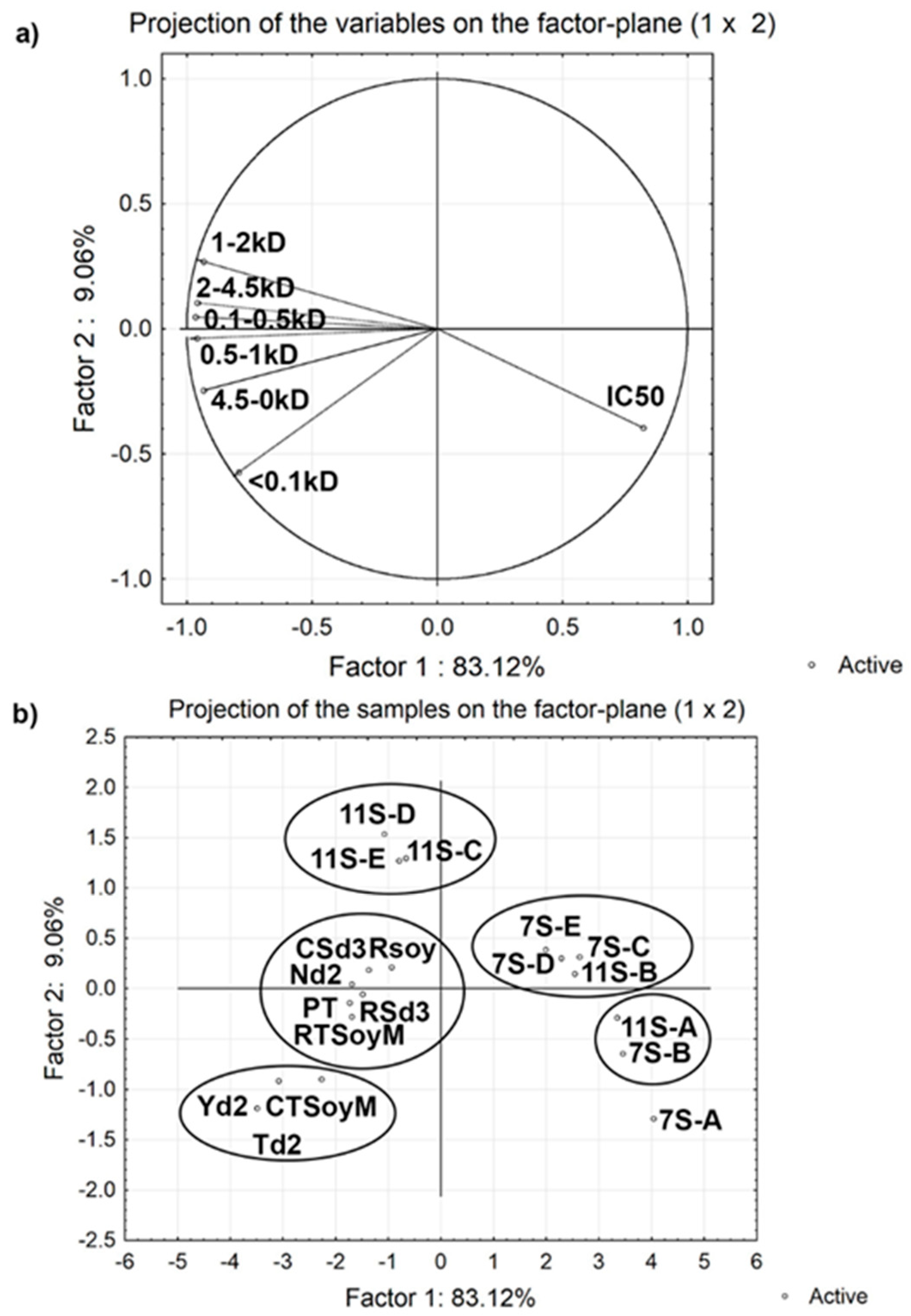
3.4. Effect of Molecular Weight of Peptides on Angiotensin Converting Enzyme (ACE) Inhibitory Activity
4. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Merai, R.; Siegel, C.; Rakotz, M.; Basch, P.; Wright, J.; Wong, B.; Thorpe, P.; Thorpe, P. CDC grand rounds: A public health approach to detect and control hypertension. MMWR Morb. Mortal. Wkly. Rep. 2016, 65, 1261–1264. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fitzgerald, R.J.; Murray, B.A. Bioactive peptides and lactic fermentations. Int. J. Dairy Technol. 2006, 59, 118–125. [Google Scholar] [CrossRef]
- Fukushima, D. Recent progress in research and technology on soybeans. Food Sci. Technol. Res. 2001, 7, 8–16. [Google Scholar] [CrossRef] [Green Version]
- Capriotti, A.; Caruso, G.; Cavaliere, C.; Samperi, R. Identification of potential bioactive generated by imulated gastrointestinal digestion of soybean seeds and soy milk proteins. J. Food Compos. Anal. 2015, 44, 205–213. [Google Scholar] [CrossRef]
- Gu, Y.; Wu, J. LC-MS/MS Coupled with QSAR modeling in characterizing of angiotensin I-converting enzyme inhibitory peptides from soybean proteins. Food Chem. 2013, 141, 2682–2690. [Google Scholar] [CrossRef] [PubMed]
- Gibbs, B.F.; Zougman, A.; Masse, R.; Mulligan, C. Production and characterization of bioactive peptides from soy hydrolysate and soy-fermented food. Food Res. Int. 2004, 37, 123–131. [Google Scholar] [CrossRef]
- Coscueta, E.R.; Campos, D.A.; Osório, H.; Nerli, B.B.; Pintado, M. Enzymatic soy protein hydrolysis: A tool for biofunctional food ingredient production. Food Chem. X 2019, 1, 100006. [Google Scholar] [CrossRef]
- Li, F.; Ohnishi-Kameyama, M.; Takahashi, Y.; Yamaki, K. Angiotensin I-converting enzyme inhibitory activities of chinese fermented soypaste and estimation of the inhibitory substances. J. Funct. Foods 2013, 5, 1991–1995. [Google Scholar] [CrossRef]
- Alauddin, M.; Shirakawa, H.; Hiwatashi, K.; Shimakage, A.; Takahashi, S.; Shinbo, M.; Komai, M. Processed soymilk effectively ameliorates blood pressure elevation in spontaneously hypertensive rats. J. Funct. Foods 2015, 14, 126–132. [Google Scholar] [CrossRef]
- Puchalska, P.; Concepción García, M.; Luisa Marina, M. Identification of native angiotensin-I converting enzyme inhibitory peptides in commercial soybean based infant formulas using HPLC-Q-ToF-MS. Food Chem. 2014, 157, 62–69. [Google Scholar] [CrossRef]
- Hernández-Ledesma, B.; García-Nebot, M.J.; Fernández-Tomé, S.; Amigo, L.; Recio, I. Dairy protein hydrolysates: Peptides for health benefits. Int. Dairy J. 2014, 38, 82–100. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Y.; Guo, S.; Liu, Z.; Chang, S.K.C. Off-flavor related volatiles in soymilk as affected by soybean variety, grinding, and heat-processing methods. J. Agric. Food Chem. 2012, 60, 7457–7462. [Google Scholar] [CrossRef] [PubMed]
- Meng, S.; Chang, S.; Gillen, A.M.; Zhang, Y. Protein and quality analyses of accessions from the USDA soybean germplasm collection for tofu production. Food Chem. 2016, 213, 31–39. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wei, Q.; Wolf-Hall, C.; Chang, K.C. Natto characteristics as affected by steaming time, bacillus strain, and fermentation time. J. Food Sci. 2001, 66, 167–173. [Google Scholar] [CrossRef]
- Kumari, S.; Chang, S.K.C. Effect of cooking on isoflavones, phenolic acids, and antioxidant activity in sprouts of prosoy soybean (glycine max). J. Food Sci. 2016, 81, C1679–C1691. [Google Scholar] [CrossRef]
- Liu, C.; Wang, H.; Cui, Z.; He, X.; Wang, X.; Zeng, X.; Ma, H. Optimization of extraction and isolation for 11S and 7S globulins of soybean seed storage protein. Food Chem. 2007, 102, 1310–1316. [Google Scholar] [CrossRef]
- Cushman, D.W.; Cheung, H.S. Spectophotometric assay and properties of the angiotensin I-converting enzyme of rabbit lung. Biochem. Pharmacol. 1971, 20, 1637–1648. [Google Scholar] [CrossRef]
- You, S.J.; Udenigwe, C.C.; Aluko, R.E.; Wu, J. Multifunctional peptides from egg white lysozyme. Food Res. Int. 2010, 43, 848–855. [Google Scholar] [CrossRef]
- Smith, P.K.; Krohn, R.I.; Hermanson, G.T.; Mallia, A.K.; Gartner, F.H.; Provenzano, M.D.; Fujimoto, E.K.; Goeke, N.M.; Olson, B.J.; Klenk, D. Measurement of protein using bicinchoninic acid. Anal. Chem. 1985, 150, 76–85. [Google Scholar] [CrossRef]
- Lourenço da Costa, E.; Antonio da Rocha Gontijo, J.; Netto, F.M. Effect of heat and enzymatic treatment on the antihypertensive activity of whey protein hydrolysates. Int. Dairy J. 2007, 17, 632–640. [Google Scholar] [CrossRef]
- Zakharov, A.; Carchilan, M.; Stepurina, T.; Rotari, V.; Wilson, K.; Vaintraub, I. A comparative study of the role of the major proteinases of germinated common bean (Phaseolus vulgaris L.) and soybean (Glycine max (L.) merrill) seeds in the degradation of their storage proteins. J. Exp. Bot. 2004, 55, 2241–2249. [Google Scholar] [CrossRef] [PubMed]
- Yang, M.; Li, L. Physicochemical, textural and sensory characteristics of probiotic soy yogurt prepared from germinated soybean. Food Technol. Biotechnol. 2010, 9862, 490–496. [Google Scholar]
- Kirk, T.K.; Gifford, O.; Drive, P.; Farrell, R.L. Enzymatic “combustion”: The microbial degradation of lignin. Microbiology 1987, 465–505. [Google Scholar] [CrossRef] [PubMed]
- Barbana, C.; Boye, J.I. Angiotensin I-converting enzyme inhibitory activity of chickpea and pea protein hydrolysates. Food Res. Int. 2010, 43, 1642–1649. [Google Scholar] [CrossRef]
- Lahogue, V.; Réhel, K.; Taupin, L.; Haras, D.; Allaume, P. A HPLC-UV method for the determination of angiotensin I-converting enzyme (ACE) inhibitory activity. Food Chem. 2010, 118, 870–875. [Google Scholar] [CrossRef]
- Hayes, M.; Tiwari, B.K. Bioactive carbohydrates and peptides in foods: An overview of sources, downstream processing steps and associated bioactivities. Int. J. Mol. Sci. 2015, 16, 22485–22508. [Google Scholar] [CrossRef] [Green Version]
- Nik, A.M.; Tosh, S.M.; Poysa, V.; Woodrow, L.; Corredig, M. Protein recovery in soymilk and various soluble fractions as a function of genotype differences, changes during heating, and homogenization. J. Agric. Food Chem. 2008, 56, 10893–10900. [Google Scholar] [CrossRef]
- Thanh, V.H.; Shibasaki, K. Major proteins of soybean seeds. subunit structure of beta.-conglycinin. J. Agric. Food Chem. 1978, 26, 692–695. [Google Scholar] [CrossRef]
- Wu, J.; Aluko, R.E.; Nakai, S. Structural requirements of angiotensin I-converting enzyme inhibitory peptides: Quantitative structure—Activity relationship study of Di- and tripeptides. J. Agric. Food Chem. 2006, 54, 732–738. [Google Scholar] [CrossRef]
- Wu, J.; Aluko, R.E.; Nakai, S. Structural requirements of angiotensin I-converting enzyme inhibitory peptides: Quantitative structure-activity relationship modeling of peptides containing 4-10 amino acid residues. QSAR Comb. Sci. 2006, 25, 873–880. [Google Scholar] [CrossRef]
- Zhang, Y.; Pechan, T.; Chang, S.K.C. Antioxidant and angiotensin-I converting enzyme inhibitory activities of phenolic extracts and fractions derived from three phenolic-rich legume varieties. J. Funct. Foods 2018, 42, 289–297. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
| Digested Soy Foods | IC50 (mg/mL) | Soluble Protein (µg/mg) |
|---|---|---|
| RSoy | 0.19 ± 0.01 d | 368.64 ± 4.25 d |
| RTSoyM | 0.10 ± 0.01 a | 546.36 ± 2.04 a,b |
| CTSoyM | 0.12 ± 0.01 a,b | 482.38 ± 18.82 a,b,c,d |
| PT | 0.20 ± 0.00 d | 432.00 ± 15.31 b,c,d |
| RSd3 | 0.15 ± 0.00 b,c | 562.46 ± 10.91 a |
| CSd3 | 0.20 ± 0.02 d | 381.32 ± 15.81 c,d |
| Nd2 | 0.16 ± 0.01 c,d | 569.60 ± 8.98 a |
| Td2 | 0.26 ± 0.00 e | 487.44 ± 2.57 a,b,c |
| Yd2 | 0.17 ± 0.01 c,d | 418.19 ± 8.07 c,d |
| Samples | IC50 (mg/mL) | Samples | IC50 (mg/mL) |
|---|---|---|---|
| 7S-A | 5.20 ± 0.13 a,A | 11S-A | 2.41 ± 0.38 a,B |
| 7S-B | 3.55 ± 0.03 b,A | 11S-B | 1.84 ± 0.06 b,B |
| 7S-C | 0.99 ± 0.01 c,A | 11S-C | 0.40 ± 0.04 c,B |
| 7S-D | 1.20 ± 0.01 c,A | 11S-D | 0.21 ± 0.02 c,d,B |
| 7S-E | 1.04 ± 0.01 c,A | 11S-E | 0.15 ± 0.07 d,B |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Handa, C.L.; Zhang, Y.; Kumari, S.; Xu, J.; Ida, E.I.; Chang, S.K.C. Comparative Study of Angiotensin I-Converting Enzyme (ACE) Inhibition of Soy Foods as Affected by Processing Methods and Protein Isolation. Processes 2020, 8, 978. https://0-doi-org.brum.beds.ac.uk/10.3390/pr8080978
Handa CL, Zhang Y, Kumari S, Xu J, Ida EI, Chang SKC. Comparative Study of Angiotensin I-Converting Enzyme (ACE) Inhibition of Soy Foods as Affected by Processing Methods and Protein Isolation. Processes. 2020; 8(8):978. https://0-doi-org.brum.beds.ac.uk/10.3390/pr8080978
Chicago/Turabian StyleHanda, Cíntia L., Yan Zhang, Shweta Kumari, Jing Xu, Elza I. Ida, and Sam K. C. Chang. 2020. "Comparative Study of Angiotensin I-Converting Enzyme (ACE) Inhibition of Soy Foods as Affected by Processing Methods and Protein Isolation" Processes 8, no. 8: 978. https://0-doi-org.brum.beds.ac.uk/10.3390/pr8080978