Frequency and Spectrum of Radiation-Induced Mutations Revealed by Whole-Genome Sequencing Analyses of Plants


Radiation Breeding Team, Advanced Radiation Technology Institute, Korea Atomic Energy Research Institute, Jeongeup 56212, Korea
*
Author to whom correspondence should be addressed.
Quantum Beam Sci. 2019, 3(2), 7; https://0-doi-org.brum.beds.ac.uk/10.3390/qubs3020007
Submission received: 29 March 2019
/
Revised: 26 April 2019
/
Accepted: 26 April 2019
/
Published: 30 April 2019
(This article belongs to the Special Issue Ion Beams in Biology and Medicine)
Abstract
:Mutation breeding and functional genomics studies of mutant populations have made important contributions to plant research involving the application of radiation. The frequency and spectrum of induced mutations have long been regarded as the crucial determinants of the efficiency of the development and use of mutant populations. Systematic studies regarding the mutation frequency and spectrum, including genetic and genomic analyses, have recently resulted in considerable advances. These studies have consistently shown that the mutation frequency and spectrum are affected by diverse factors, including radiation type, linear energy transfer, and radiation dose, as well as the plant tissue type and condition. Moreover, the whole-genome sequencing of mutant individuals based on next-generation sequencing technologies has enabled the genome-wide quantification of mutation frequencies according to DNA mutation types as well as the elucidation of mutation mechanisms based on sequence characteristics. These studies will contribute to the development of a highly efficient and more controlled mutagenesis method relevant for the customized research of plants. We herein review the characteristics of radiation-induced mutations in plants, mainly focusing on recent whole-genome sequencing analyses as well as factors affecting the mutation frequency and spectrum.
1. Introduction
Radiation has been widely applied to develop new cultivars used for crop production and as genetic resources for functional genomics studies in the 90 years since Stadler [1,2] first published the results of mutations induced by irradiation in crops. Initially, X-rays and gamma rays were primarily used for these purposes, but neutrons and ion beams have subsequently been applied for systematic characterizations and utilizations [3,4]. Although treatments with chemical mutagens, such as ethyl methanesulfonate (EMS), may result in more frequent mutations, the advantages of radiation include the fact that it induces a higher proportion of mutations, which substantially affects gene functions (e.g., the deletion of a complete gene), thus resulting in more loss-of-function mutations related to target traits with fewer mutations per genome [5,6,7]. Accordingly, more than three quarters of the 3286 mutant varieties registered in the Mutant Variety Database [8] were developed by irradiation.
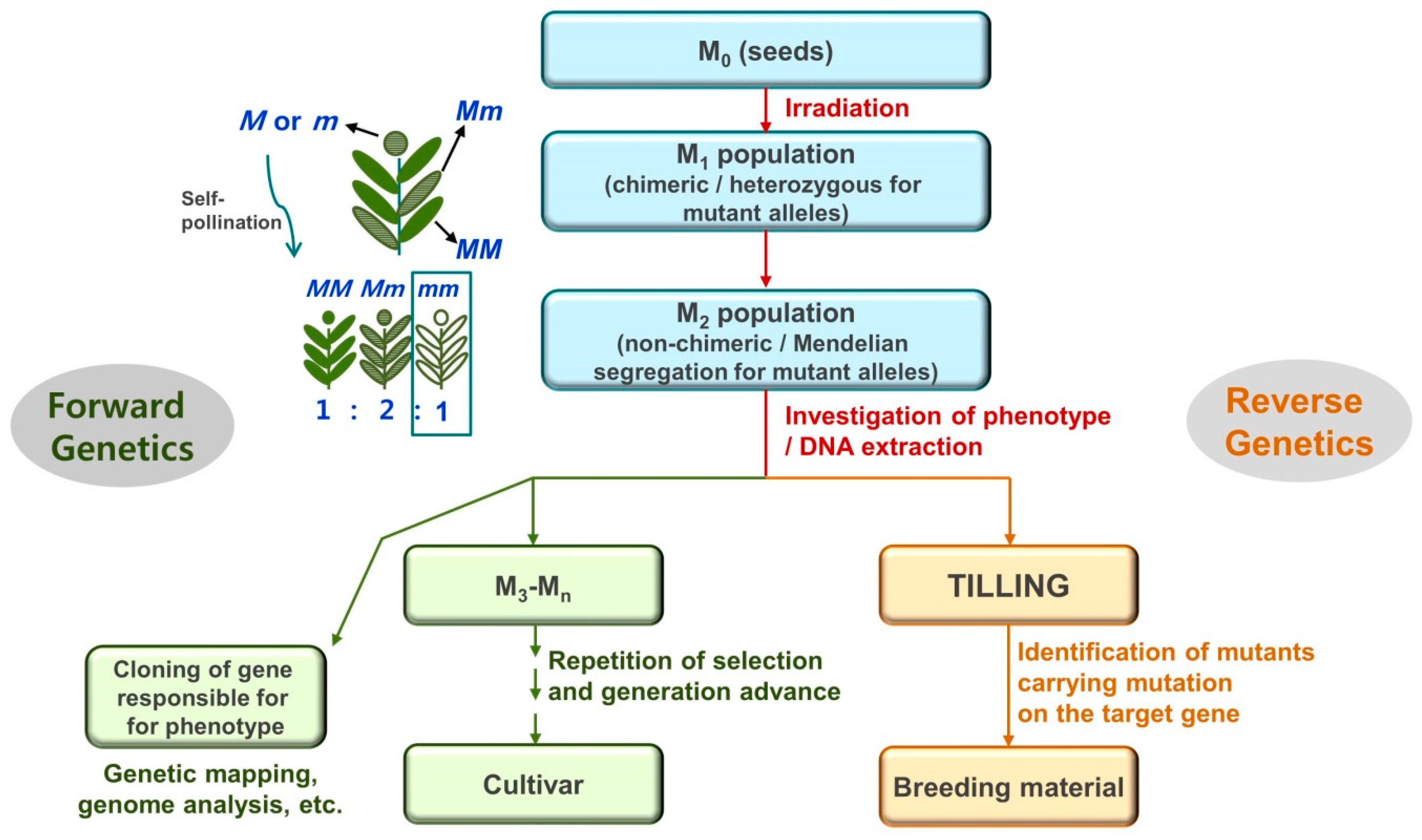
Regarding radiation-induced mutations, the frequency and spectrum of mutations are dependent on many factors, including radiation type and dose, linear energy transfer (LET; i.e., the amount of energy transferred to materials when radiation traverses a unit distance), and plant tissue type and condition [5,9,10]. Therefore, an in-depth characterization of the mutagenic effects for a specific irradiation treatment is very important for breeding and functional genomics studies. Populations with many individuals in the M2 or more advanced generations are generally developed for plant mutation studies (Figure 1). Mutant lines with target characteristics are selected in a population based on the phenotypes (forward genetics approach) and/or by screening for mutations to genes expected to be responsible for target characteristics (reverse genetics approach). The effect of a mutation on gene function as well as the mutation frequency should be considered when determining the appropriate size of a population for obtaining a certain gene coverage for knockout or knockdown studies. Additionally, the types of DNA mutations caused by irradiation should be considered. For example, if a high rate of knockout mutations for each gene is required, irradiation that results in frequent small deletions would be suitable [5]. However, if the objective is to achieve the simultaneous knockout of genes duplicated in tandem, irradiation that induces large-scale mutations, such as large deletions, is a better option [11,12]. The prevailing types of DNA mutations are also determinants of mutant screening methods in reverse genetics approaches. Single nucleotide polymorphisms and small deletions can be detected by Targeting Induced Local Lesions IN Genomes (TILLING), in which mismatches between wild-type and mutant DNA amplicons are screened based on the cleavage by endonucleases (e.g., CelI) or high-resolution melting analysis [13]. In contrast, larger deletions may be detected by De-TILLING analysis based on a specific amplification of a mutant DNA fragment with a large deletion [14,15].
In crop research, mutation frequencies have been commonly determined based on observable phenotypes, such as developmental changes or variations in leaf or flower colors, in mutant populations without the need for information regarding the mutated genes [10,16,17,18]. These studies provided useful practical information for breeding, but the mutation frequency for a single gene could not be calculated, and the types and mechanisms of DNA mutations were not elucidated. Studies involving a model plant species, Arabidopsis thaliana, overcame these limitations because of its well-characterized phenotype-to-genotype relationships [5,19,20,21,22]. Because the frequency and sequences of mutant alleles for specific genes were analyzed in these studies, the mutation frequency for a single gene and the characteristics of the mutated sequences were revealed. The recent application of high-throughput sequencing technology has provided additional information at the whole-genome level. Complete genome sequences of mutant individuals in small experimental groups have generated abundant information regarding the composition and distribution of different types of mutations (Table 1) [7,9,11,12,23,24,25]. Moreover, the reported whole-genome sequencing (WGS) analysis and construction of a mutation database for 1504 individuals in a large mutant population might mark the beginning of the era of radiation mutation breeding based on complete genome sequences [26,27].
In this review, we introduce the plant factors influencing the frequency and spectrum of mutations induced by irradiation. Additionally, mutation characteristics are described based on the results of recent WGS studies. Finally, future perspectives and objectives are discussed.
2. Factors Affecting the Mutation Frequency and Spectrum
Numerous factors influence the mutation frequency and spectrum in both independent and interdependent manners. In this section, we will review several factors that have been mainly analyzed by genetic studies.
2.1. Linear Energy Transfer
Numerous investigations have proven that the frequency and types of mutations in the advanced generations (M2 and later) as well as the relative biological effectiveness determined in the M1 generation are largely dependent on the LET of the applied radiation [4,27,28,29]. Among the radiation types frequently used for mutagenesis, heavy ion beams and fast neutrons are high-LET radiation types, whereas gamma rays and X-rays are classified as low-LET types. The complex track structure of high-LET radiation in which energetic secondary electrons are generated, leading to energy deposition near the primary particle track, results in clustered DNA damages, which is related to the close localization of complex DNA lesions [30,31]. The clustered DNA damages include double-strand breaks (DSBs) as well as single-strand breaks (SSBs) and diverse DNA lesions [31]. In contrast, low-LET radiation mainly damages DNA via an indirect mechanism in which energy deposited in water molecules produces hydroxyl radicals that target DNA molecules [32]. This process mainly results in SSBs and diverse base or sugar lesions, although some DSBs may also occur as the result of attempts to repair multiple DNA lesions induced within a small region [33]. The differences between high- and low-LET radiation regarding the mechanisms underlying DNA damages may result in unique mutation induction features due to at least three factors, namely the frequency of DSBs, the efficiency of DNA repair, and the non-homologous end joining (NHEJ) mechanism. Kikuchi et al. [34] reported that a 50-Gy neon beam produces about eight times more chromosomal breaks than irradiation by X-rays. In a comparison of carbon beams with different LETs, Hase et al. [27] demonstrated that the efficiency of DSB repair by NHEJ is much lower for high-LET radiation based on the finding that the difference in the radiation sensitivity between the wild-type and DNA ligase IV mutant (deficient in the classical NHEJ (C-NHEJ) repair pathway) is much smaller for the high-LET carbon beam than for the low-LET carbon beam. Hirano et al. [11] added a new interpretation for the same result based on the DNA rearrangement junction sequences obtained in their WGS analysis. They suggested that an alternative end-joining (A-EJ) pathway instead of the C-NHEJ pathway may be mainly responsible for the DNA repair during the application of high-LET radiation; this will be discussed in more detail later.
In terms of the mutation frequency, Shikazono et al. [21] reported that the average mutation rate per dose is about 17-fold higher for carbon beams (LET = 113 KeV μm−1) than for electron beams (LET = 0.2 KeV μm−1) in the A. thaliana M2 generation. This difference decreases to 3.4-fold if the comparison is performed for radiation doses that result in similar lethalities. Kazama et al. [28] analyzed A. thaliana phenotypic mutation frequencies for radiation types with more subdivided LETs. They determined that carbon and nitrogen ion beams with a specific LET (30 KeV μm−1), which was designated as LETmax, resulted in the highest mutation frequency when compared with radiation treatments with a higher or lower LET. For example, a carbon beam (30 KeV μm−1) irradiation resulted in 4.0- and 2.4-times more albino mutants in the M2 generation than irradiations with 22.5 KeV μm−1 and 61.5 KeV μm−1 carbon beams, respectively. In a subsequent study, the ratio between small alterations (single base substitutions (SBSs) and small insertions and deletions (InDels)) and DNA rearrangements varied depending on the LET. A carbon beam with LETmax frequently induced null mutations of genes due to small alterations rather than DNA rearrangements [5]. The mutation phenotype spectrum is also affected by the LET. A much broader range of flower color variations was detected for a carbon beam irradiation than for a gamma irradiation [35].
2.2. Irradiation Dose
Lethality and fertility in the M1 generation are usually considered when the mutation frequency due to the irradiation dose is investigated because the M2 population must be generated by reproduction in mutation breeding studies (Figure 1). Therefore, determining the lowest dose that results in the desired mutation frequency is warranted. Yamaguchi et al. [10] investigated the relationship between the survival rate of rice M1 individuals and the phenotypic mutation frequency in M2 populations during radiation treatments with increasing doses. They revealed that the mutation frequency per M2 plant increased rapidly as the survival rate decreased when the survival rate was in the range of 90–100%, but it increased gradually as the survival rate decreased below 90%. Interestingly, the highest number of M1 lines with M2 progenies exhibiting a mutant phenotype per seed sown in the M1 generation was detected following an irradiation with the shoulder dose (i.e., the dose at which the survival rate starts to decrease rapidly in the dose–survival rate curve), regardless of radiation type. Therefore, they assumed that an irradiation with the shoulder dose can efficiently induce mutations and minimize the effects on survival. The optimal dose can be decreased to 50–75% of the dose at the end of the shoulder to minimize the background mutation during breeding [36].
Kazama et al. [5] confirmed that, in addition to the LET, the particle number of ion beams is closely related to the mutation frequency. An analysis of the mutation frequency of A. thaliana plants exposed to carbon ion beams with different LETs (22.5 and 30 KeV μm−1) indicated that the mutation frequency increased as the number of particles increased whilethe maximum mutation frequency was much higher for 30 KeV μm−1 than for 22.5 KeV μm−1. Accordingly, they inferred the reason why irradiation with a LET higher than 30 KeV μm−1 (LETmax) results in a lower mutation frequency than that obtained with an irradiation with LETmax. Because increases in the irradiation dosage are limited due to plant lethality, an insufficient number of particles may be treated by high-LET radiation, which may decrease the chances of mutation. Therefore, an integrated understanding of LET, irradiation dose, and relative biological effects on plant survival or fertility is required for determining the optimal irradiation type and conditions for mutation breeding studies.
2.3. Effects of Plant Tissue Type and Condition
The hydration state of a plant sample can influence the mutagenic effect of irradiation because indirect DNA damages are mainly caused by hydroxyl radicals generated from water molecules. Therefore, the water content may affect the mutation frequency or the proportion of direct and indirect DNA damages. Water contents below 14% reportedly increase the sensitivity of barley seeds to gamma rays and X-rays, although the underlying mechanism has not been characterized [3]. In contrast, according to Vochita et al. [37], who analyzed the cytological effects of low-LET radiation (X-ray and electron beams) on dry and hydrated seeds, the mitotic index is lower and the rate of chromosomal aberrations is higher in irradiated hydrated samples.
Regarding irradiated tissues, Hase et al. [9] proved that the mutation frequency induced by carbon beam irradiation was higher for dry seeds than for seedlings in A. thaliana. The difference in the mutation rates was mainly due to the high rate of InDel mutations in the irradiated seeds (described later). They suggested the following three reasons for this finding: the higher water content in seedlings may lead to fewer direct-to-indirect DNA damages, the cells carrying genetic material with abundant InDels may be selectively eliminated during the cell proliferation of seedling meristems, and the chromatin in seeds is highly condensed, which may be related to the repair process. Meanwhile, Kowyama et al. [38] reported that the mutation frequency in rice was higher for microspores irradiated at the late tetrad stage than for irradiated seeds or egg cells. Consequently, the irradiation of microspores at this stage may be an efficient method for generating a large number of mutations in rice.
Finally, a specific pretreatment of samples before irradiation can increase the frequency of the targeted mutation. For example, a pretreatment with sucrose, which upregulates the expression of anthocyanin biosynthesis genes, increases the number of flower-color mutants generated by carbon beam irradiation in petunia, although the frequency of chlorophyll mutations is almost unchanged [39]. Therefore, modulating the gene expression pattern may lead to increased controlled mutation, although the responsible mechanism will need to clarified [40].
3. Characteristics of DNA Mutations Revealed by Whole-Genome Sequencing
Additional complete genome sequences of plant mutants generated by irradiation have been reported since Belfield et al. [23] first published the sequence of A. thaliana mutants obtained by fast-neutron irradiation. These sequences have been used for the comparative analysis of DNA mutations between various types of radiation treatments or target samples and also for constructing a genomic database for mutant populations (Table 1). In this section, we will describe the characteristics of DNA mutations caused by irradiation under diverse conditions (mainly radiation type and LET) based on WGS studies. The results of EMS treatments and spontaneous mutations were also included for comparison. The data from earlier investigations were reinterpreted for an integrated description. Thus, the values for several parameters presented herein may differ somewhat from the values in the original articles because of differences in categorization. In addition, we cannot rule out the possibility that characteristics of mutations may be different according to plant species, irradiation conditions, and NGS analysis methods. Thus, we recommend that readers refer to the data in this section to assess the overall features of DNA mutations.
3.1. Frequency and Distribution of Mutations
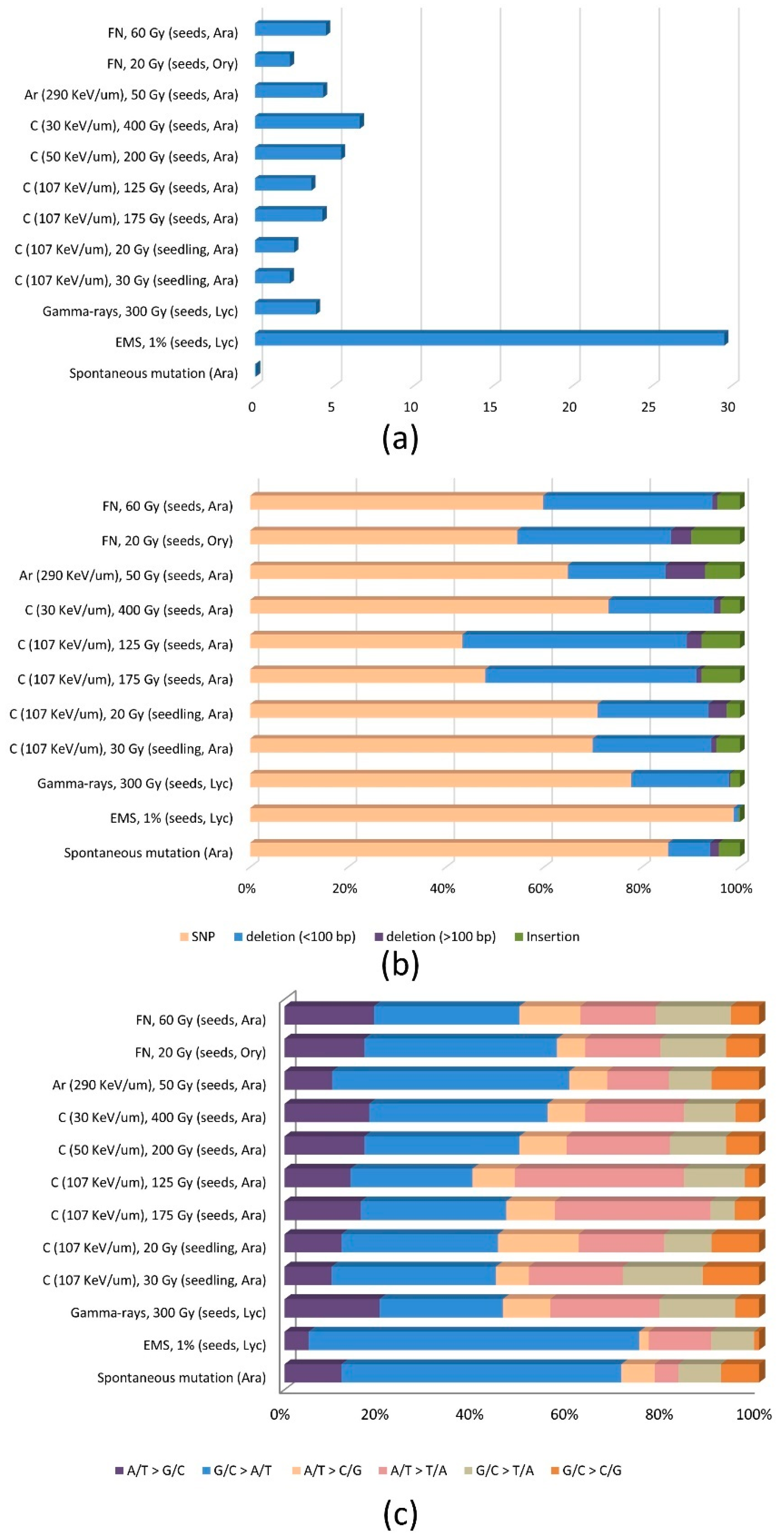
Mutation frequencies in the M1 generation were estimated from those of the M2 or M3 generations, assuming a Mendelian segregation of mutant alleles. These values were used for comparing the mutation frequencies of different generations (M2 or M3) and may differ substantially from the actual mutation frequency of the M1 generation because many of the large deletions induced in the M1 generation are not transmitted to the next generation [41]. A carbon beam with LETmax resulted in a high mutation frequency (6.6 × 10−7/bp) in A. thaliana (Figure 2a). This mutation frequency was similar to or higher than that resulting from an irradiation of A. thaliana by a carbon beam with a higher LET (107 KeV μm−1; 4.3 × 10−7/bp at 175 Gy and 3.6 × 10−7/bp at 125 Gy) or an argon beam (290 KeV μm−1; 4.3 × 10−7/bp). The irradiation of seedlings produced the lowest mutation frequency. The mutation frequency due to fast neutrons was similar to that obtained with carbon ion beam irradiation (107 KeV μm−1) and an argon beam at 60 Gy. Additionally, the mutation frequency in gamma-irradiated tomato plants was not much lower than that resulting from ion beams, which was inconsistent with the findings of other studies that indicated significantly higher mutation frequencies were induced by ion beam irradiations [10,21]. The mutation frequency range among irradiations was 2.2–6.6 × 10−7/bp, which was 31 to 93-times higher than the frequency of spontaneous mutations estimated for A. thaliana (7.1 × 10−9/bp), and 4.5 to 13.4-times lower than the mutation frequency in EMS-treated tomato (29.6 × 10−7/bp).
Several WGS investigations revealed an even distribution of mutations in the genome. Irradiations by fast neutrons (20 Gy) and a carbon beam (50 KeV μm−1) induced mutations that were evenly distributed across chromosomes [24,25,27]. Additionally, mutations were not preferentially located in genic or intergenic regions following fast neutron irradiation (60 Gy) [23]. Similarly, the distribution of mutations was not biased toward euchromatin or heterochromatin in tomato irradiated with gamma rays or treated with EMS [7].
3.2. Mutation Spectrum and Mechanism
DNA mutations are often classified as SBSs, small InDels, or large-scale DNA rearrangements, including large deletions, duplications, inversions, and translocations. Although radiation treatments are believed to mainly induce large-scale mutations, sequencing analyses have indicated that SBSs and small InDels represent a considerable portion of the total mutations in terms of the number of mutation events. On the basis of WGS analyses (Figure 2b), we determined that range of the ratio of SBSs to InDels was 0.77 (carbon beam, 107 KeV μm−1, 125 Gy) to 3.5 (gamma rays). These ratios were much smaller than those resulting from spontaneous mutations (5.8) and EMS treatments (75.8). The high-LET irradiation of seeds (e.g., carbon beam (107 KeV μm−1), argon beam (290 KeV μm−1), and fast neutrons) generally led to a larger proportion of InDels compared with the effects of low-LET irradiation. The irradiation of seedlings reportedly induced relatively few InDels, which may contribute to the lower mutation frequency of seedlings than of seeds following a carbon beam irradiation [9]. Interestingly, only a carbon beam irradiation (107 KeV μm−1) of A. thaliana seeds resulted in more InDels than single nucleotide polymorphisms although the LET was lower than that of argon beams, implying that a high LET is not the only requirement for a higher frequency of InDels.
The main substitution in radiation-induced mutations was G/C to A/T, which is similar to the results of spontaneous or EMS-induced mutations, but its proportion of the total SBSs was much lower (Figure 2c). Many of the substituted bases in this type of mutation involved pyrimidine dinucleotides in fast neutron and carbon beam-induced mutants, implying that the mechanism underlying UV-induced mutations may be applicable [23,24]. Irradiation with UV light can induce the formation of cyclobutane pyrimidine dimers at pyrimidine dinucleotide sites. The C and 5-methyl-C in cyclobutane pyrimidine dimers are unstable and deaminate to U and T, leading to a C-to-T mutation [42]. This is different from the well-known mechanism responsible for the G/C to A/T substitution following an EMS treatment, which involves the alkylation of G, leading to the mispairing with T. Other substitutions such as A/T to T/A, A/T to G/C, G/C to T/A, and A/T to C/G are generally more frequent in SBSs induced by radiation than by spontaneous mutations or EMS, although the relative proportion differs according to the radiation type and irradiation conditions (Figure 2c). The mechanisms for these mutations remain largely unknown. However, Belfield et al. [23] proposed that the G/C to T/A substitution might be caused by the guanine oxidation due to the effects of reactive oxygen species and hydroxyl radicals produced by irradiation. Hase et al. [9] disagreed with this proposal because they did not observe an increase in the frequency of this type of substitution following the irradiation of seedlings, which is expected to increase the concentration of hydroxyl radicals. The relatively even composition of radiation-induced SBSs leads to less-biased amino acid changes compared with those induced by EMS if the SBSs are located in genes [27].
Although it is difficult to generalize the distribution pattern of deletions according to their length because of the relatively few large deletions detected in a limited number of samples, the proportion of large deletions (>100 bp) is highest following irradiations with an argon beam (290 KeV μm−1 at 50 Gy; 28.6%) and fast neutrons (20 Gy; 11.6%) with a high LET. In several studies, a major portion of the irradiation-induced single nucleotide deletions was detected in homopolymeric sequences, implying they might be generated by the DNA replication slippage induced by irradiation [9,23,24]. However, significant portions of larger deletions were shown to involve microhomologous sequences at their junctions [9,11,12]. The error-prone A-EJ pathway, which usually involves microhomologous sequences at the DNA ends, might be the responsible mechanism because this pathway is associated with deletions in mammals [43], and the expression of A-EJ pathway-related genes in rice is upregulated as the LET increases during irradiations [44]. Similar relationships between length and junction sequences were reported for insertions caused by carbon beam irradiation [9]. Some of the insertions resulting from argon beam irradiation were identified as filler DNA sequences connecting DNA rearrangement junctions to each other [12]. None of the filler DNA sequences were significantly similar to any sequences in the A. thaliana reference genome. These filler DNA sequences were suggested to be synthesized by the repair pathway mediated by DNA polymerase theta, which is known to be involved in the A-EJ pathway of mammals [12].
Large-scale DNA rearrangements have been analyzed in only a few studies [11,12,25,26] because of the difficulties in detecting these mutations using the short reads generated by NGS. In terms of the number of mutations, large DNA rearrangements represented 9.0% of the total mutations following the fast neutron irradiation (20 Gy) of rice [26] and 26.7% and 3.8% of the total mutations induced by argon beam (290 KeV μm−1, 50 Gy) and carbon beam (30 KeV μm−1, 400 Gy) irradiations of A. thaliana, respectively. Hirano et al. [11] revealed that 62% of the DNA rearrangement junctions contained microhomologous sequences, suggesting the A-EJ pathway was involved in repairing the DSBs caused by argon beam irradiation. A subsequent study by Kazama et al. [12] demonstrated that the DNA rearrangement characteristics vary according to the radiation LET. Argon beam irradiations lead to more complicated rearrangements involving multiple events and many filler DNA sequences (discussed above) compared with carbon beam radiation treatments. The higher frequency of DNA rearrangements associated with argon beam irradiations suggests that high-LET irradiation leads to complex and clustered DNA damages that are difficult to repair.
3.3. Effects on Gene Function
Gene mutations that can be inherited by the next generation are usually used for mutation breeding or functional genomics studies. Naito et al. (2005) observed that most large deletions induced by gamma rays or carbon beams in the M1 generation cannot be inherited by the subsequent generation. Additionally, Kazama et al. [12] confirmed that five of six large deletions that were heterozygous in the M2 generation could not exist in the homozygous state in the M3 generation. These observations may indicate that genes essential for survival were located in the deleted regions. Therefore, the zygosity of gene mutations as well as the effects of the mutations on gene functions must be analyzed. Only three published studies provide enough information to satisfy these requirements [11,12,23]. Although two independent investigations [11,12] generated information regarding argon beam irradiations, only the gene function data from Hirano et al. [11] were considered for this review because one mutant line in the Kazama et al. [12] study contained extraordinarily long deleted sequences and included 852 genes.
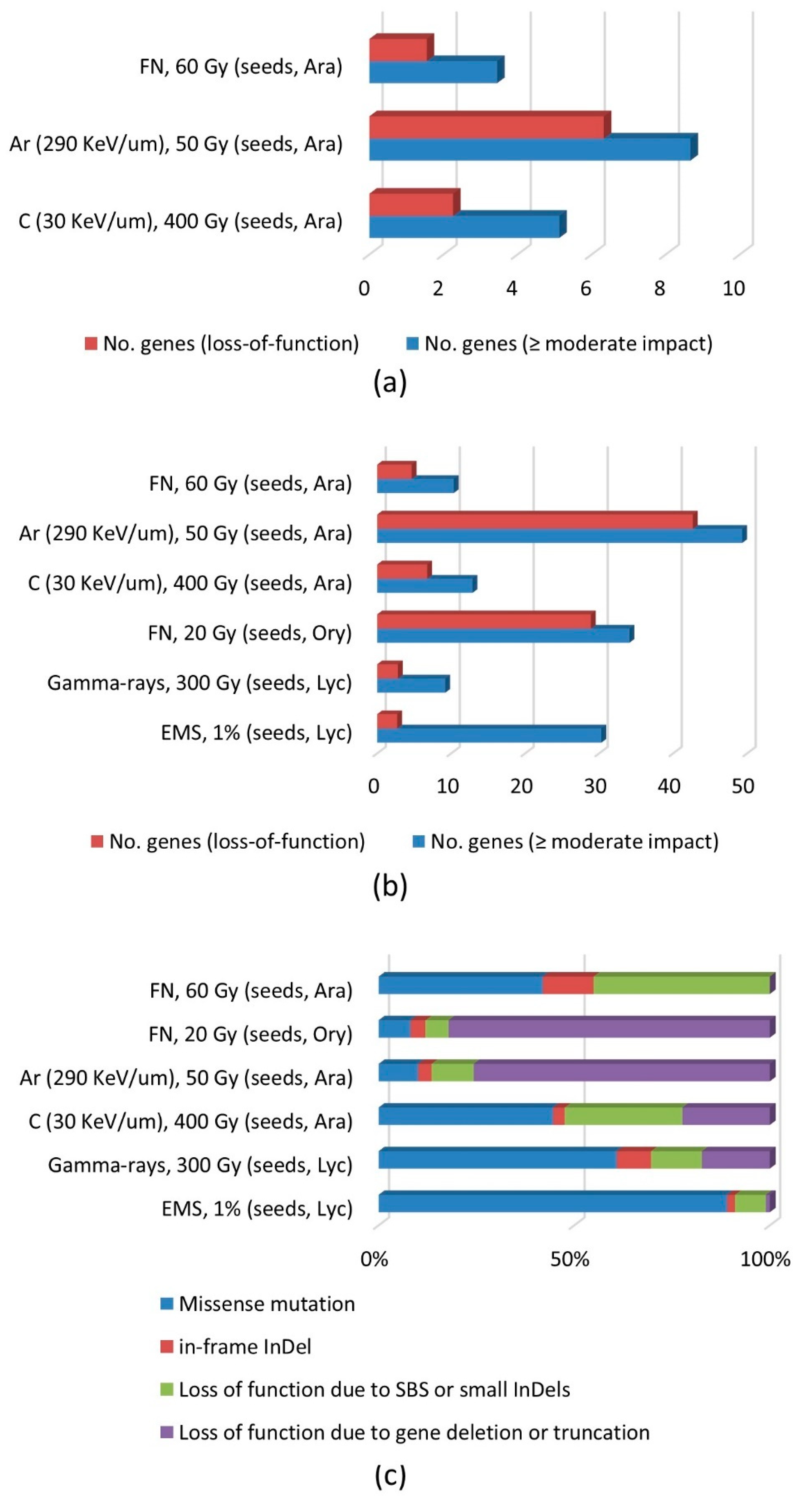
When the number of genes affected by homozygous mutations was calculated for the M2 generation, a loss-of-function mutation (i.e., nonsense mutation, loss of the start codon or splicing site, truncation, and gene loss) was predicted for 1.6, 2.3, and 6.3 genes following fast neutron (60 Gy), carbon beam (30 KeV μm−1, 400 Gy), and argon beam (290 KeV μm−1, 50 Gy) irradiations, respectively (Figure 3a). If the frequency of mutations with a moderate or high impact was calculated by adding the number of genes carrying missense mutations and in-frame InDels, the numbers of affected genes were 3.5, 5.1, and 8.7, respectively. The proportion of loss-of-function mutations was higher for the argon beam irradiation because a large portion of these mutations were caused by the loss of complete genes due to large deletions (discussed below).
If heterozygous mutations are included in analyses, the number of genes with loss-of-function mutations 2.8 to 42.7 (Figure 3b). Regarding the argon beam irradiation, the number of genes with heterozygous or homozygous mutations was 5.7-times higher than the number of genes with homozygous mutations. In contrast, it was only 2.5-times higher for the carbon beam irradiation (30 KeV μm−1, 400 Gy). These results may be explained by the fact that many of the large deletions induced by high-LET radiation can exist only in the heterozygous state, probably because genes essential for survival are present in the deleted regions. In contrast, small-scale mutations caused by lower-LET irradiation likely affect only a single gene without simultaneously influencing other genes. This assumption was supported by the spectrum of mutations affecting gene functions (Figure 3c). During irradiations by fast neutrons (20 Gy) and argon beams (290 KeV μm−1, 50 Gy), about 90% of the mutations affecting gene functions led to a loss of function, and most of them were truncations or deletions of genes due to large deletions or DNA rearrangements. Irradiations with a lower-LET carbon beam (30 KeV μm−1, 400 Gy) or gamma rays resulted in a smaller proportion of loss-of-function mutations, with SBSs or small InDels representing a significant portion of the mutations. The largest proportion of loss-of-function mutations due to SBSs or small InDels was observed for a carbon beam irradiation (30 KeV μm−1, 400 Gy). Kazama et al. [12] demonstrated that this treatment is very efficient for inducing small deletions affecting gene functions. In contrast, an EMS treatment resulted in considerably more missense mutations and fewer gene deletions or truncations. Although an EMS treatment led to a very high frequency of mutations affecting amino acid sequences, the frequency of high-impact mutations was similar to that induced by gamma-irradiation and was lower than that resulting from higher-LET irradiations when both homozygous and heterozygous mutations were included for analysis (Figure 3b).
On the basis of the frequency of mutated genes, we calculated the number of individuals in the M2 population required to ensure that at least one individual has a mutation in a gene of interest according to a certain probability. If only the homozygous mutations were considered (for a forward-genetics approach), 17,723, 12,210, and 7224 M2 individuals were required for a 90% probability following the fast neutron (60 Gy), carbon beam (30 KeV μm−1, 400 Gy), and argon beam (290 KeV μm−1, 50 Gy) irradiations, respectively. When the analysis included heterozygous mutations (for a reverse-genetics approach), only 1268 and 3781 M2 individuals were needed for a 90% probability with the argon beam (290 KeV μm−1, 50 Gy) and fast neutron (20 Gy) irradiations, respectively.
Our analyses indicated that special precautions are required when characterizing the effects of high-LET irradiation on gene functions because this radiation type usually results in large deletions of many genes. An analysis that includes only a small number of samples may lead to biased results because fewer than three deletions typically exist in a sample and even a single deletion may substantially affect the mutation frequency and spectrum of the affected genes. In fact, Belfield et al. [23] analyzed three individuals irradiated by fast neutrons (60 Gy), and gene losses due to deletions were not detected. However, gene losses were frequently detected in another study involving an analysis of 1504 individuals irradiated by fast neutrons (20 Gy) (Li et al., 2017). Therefore, a sufficient number of individuals is critical for analyses of large-scale structural variations. Additionally, as discussed earlier, a significant portion of large deletions in the heterozygous state might not be able to exist in the homozygous state. Thus, it may not be possible to evaluate the phenotypic consequences of a gene deletion. Calculations based on the frequency of homozygous mutations may be more useful for planning radiation breeding experiments or functional genomics studies.
4. Future Perspectives
Recent WGS analyses of irradiation-induced mutants have greatly enhanced our understanding of the mutation frequency and spectrum. The results of these studies may be useful for selecting the appropriate radiation type and irradiation conditions to generate mutations with specific characteristics. Therefore, inducing mutations in a way that is optimal for specific purposes may be achieved by selecting an appropriate radiation type and optimizing the irradiation conditions. Additionally, in a more active approach, the desired mutations may be induced following the pre-treatment or genetic engineering of irradiated samples (e.g., the silencing of a specific DNA repair pathway). Accordingly, the generation of mutants via diverse irradiation methods followed by the characterization of the DNA mutations by sophisticated NGS pipelines will likely become more popular in future studies. The use of long-read sequencing technologies (e.g., PacBio sequencing and Oxford Nanopore sequencing) will further improve the accuracy and throughput of analyses of large structural variations. Moreover, the application of sequencing technologies will provide opportunities for the efficient use of genetic resources developed by irradiation. As mentioned earlier, WGS was first completed for radiation-induced rice mutant populations [26]. Although sequencing the complete genome is likely not currently feasible for crops with large genomes, the sequencing of selected regions (e.g., exome sequencing) [45,46] may be an alternative way to develop a genome database or determine mutation frequency for mutation populations of important crops. Combining radiation and DNA sequencing technologies will help to maximize mutagenesis efficiencies and enable the systemic use of the developed genetic materials for plant breeding and functional genomics investigations.
Author Contributions
Conceptualization, Y.D.J.; Data Analysis, Y.D.J.; Writing—Original Draft Preparation, Y.D.J.; Writing—Review & Editing, J.-B.K.
Funding
This work was supported by grants from the Nuclear R&D Program of the Ministry of Science and ICT (MSIT) and the research program of KAERI, Republic of Korea.
Acknowledgments
We thank Edanz Group (www.edanzediting.com/ac) for editing a draft of this manuscript.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Stadler, L.J. Mutations in barley induced by X-rays and radium. Science 1928, 68, 186–187. [Google Scholar] [CrossRef] [PubMed]
- Stadler, L.J. Genetic effects of X-rays in maize. Proc. Natl. Acad. Sci. USA 1928, 14, 69–75. [Google Scholar] [CrossRef] [PubMed]
- Shu, Q.Y.; Forster, B.P.; Nakagawa, H. Plant Mutation Breeding and Biotechnology; CAB International: Wallingford, UK, 2012; ISBN 9789251070222. [Google Scholar]
- Tanaka, A.; Shikazono, N.; Hase, Y. Studies on Biological Effects of Ion Beams on Lethality, Molecular Nature of Mutation, Mutation Rate, and Spectrum of Mutation Phenotype for Mutation Breeding in Higher Plants. J. Radiat. Res. 2010, 233, 223–233. [Google Scholar] [CrossRef]
- Kazama, Y.; Hirano, T.; Saito, H.; Liu, Y.; Ohbu, S.; Hayashi, Y.; Abe, T. Characterization of highly efficient heavy-ion mutagenesis in Arabidopsis thaliana. BMC Plant Biol. 2011, 11, 161. [Google Scholar] [CrossRef] [PubMed]
- Koornneef, M.; Rolff, E.; Spruit, C.J.P. Genetic control of light-inhibited hypocotyl elongation in Arabidopsis thaliana (L) Heynh. Z. Pflanzenphysiol. 1980, 100, 147–160. [Google Scholar] [CrossRef]
- Shirasawa, K.; Hirakawa, H.; Nunome, T.; Tabata, S.; Isobe, S. Genome-wide survey of artificial mutations induced by ethyl methanesulfonate and gamma rays in tomato. Plant Biotechnol. J. 2016, 14, 51–60. [Google Scholar] [CrossRef]
- IAEA Mutant Variety Database. Available online: http://mvd.iaea.org (accessed on 13 March 2019).
- Hase, Y.; Satoh, K.; Kitamu, S.; Oono, Y. Physiological status of plant tissue affects the frequency and types of mutations induced by carbon-ion irradiation in Arabidopsis. Sci. Rep. 2018, 8, 1394. [Google Scholar] [CrossRef] [Green Version]
- Yamaguchi, H.; Hase, Y.; Tanaka, A.; Shikazono, N.; Degi, K.; Shimizu, A.; Morishita, T. Mutagenic effects of ion beam irradiation on rice. Breed. Sci. 2009, 177, 169–177. [Google Scholar] [CrossRef]
- Hirano, T.; Kazama, Y.; Ishii, K.; Ohbu, S.; Shirakawa, Y.; Abe, T. Comprehensive identification of mutations induced by heavy-ion beam irradiation in Arabidopsis thaliana. Plant J. 2015, 82, 93–104. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kazama, Y.; Ishii, K.; Hirano, T.; Wakana, T.; Yamada, M.; Ohbu, S.; Abe, T. Different mutational function of low- and high-linear energy transfer heavy-ion irradiation demonstrated by whole-genome resequencing of Arabidopsis mutants. Plant J. 2017, 92, 1020–1030. [Google Scholar] [CrossRef] [PubMed]
- Taheri, S.; Abdullah, T.L. TILLING, high-resolution melting (HRM), and next-generation sequencing (NGS) techniques in plant mutation breeding. Mol. Breed. 2017, 37, 40. [Google Scholar] [CrossRef]
- Li, X.; Song, Y.; Century, K.; Straight, S.; Ronald, P.; Dong, X.; Lassner, M. A fast neutron deletion mutagenesis-based reverse genetics system for plants. Plant J. 2001, 27, 235–242. [Google Scholar] [CrossRef] [Green Version]
- Rodgers, C.; Wen, J.; Chen, R.; Oldroyd, G. Deletion-based reverse genetics in Medicago truncatula. Plant Physiol. 2009, 151, 1077–1086. [Google Scholar] [CrossRef]
- Arase, S.; Hase, Y.; Abe, J.; Kasai, M.; Yamada, T.; Kitamura, K.; Narumi, I.; Tanaka, A.; Kanazawa, A. Optimization of ion-beam irradiation for mutagenesis in soybean: Effects on plant growth and production of visibly altered mutants. Plant Biotechnol. 2011, 329, 323–329. [Google Scholar] [CrossRef]
- Jo, Y.D.; Kim, S.H.; Hwang, J.E.; Kim, Y.S.; Kang, H.S.; Kim, S.W.; Kwon, S.J.; Ryu, J.; Kim, J.B.; Kang, S.Y. Construction of mutation populations by gamma-ray and carbon beam irradiation in chili pepper (Capsicum annuum L.). Hortic. Environ. Biotechnol. 2016, 57, 606–614. [Google Scholar] [CrossRef]
- Yamaguchi, H.; Shimizu, A.; Degi, K.; Morishita, T. Effects of dose and dose rate of gamma ray irradiation on mutation induction and nuclear DNA content in chrysanthemum. Breed. Sci. 2008, 335, 331–335. [Google Scholar] [CrossRef]
- Hirano, T.; Kazama, Y.; Ohbu, S.; Shirakawa, Y.; Liu, Y.; Kambara, T. Fundamental and molecular mechanisms of mutagenesis molecular nature of mutations induced by high-LET irradiation with argon and carbon ions in Arabidopsis thaliana. Mutat. Res./Fundam. Mol. Mech. Mutagen. 2012, 735, 19–31. [Google Scholar] [CrossRef]
- Shikazono, N.; Tanaka, A.; Watanabe, H.; Tano, S. Rearrangements of the DNA in Carbon Ion-Induced Mutants of Arabidopsis thaliana. Genetics 2001, 157, 379–387. [Google Scholar]
- Shikazono, N.; Yokota, Y.; Kitamura, S.; Suzuki, C.; Watanabe, H.; Tano, S.; Tanaka, A. Mutation Rate and Novel tt Mutants of Arabidopsis thaliana Induced by Carbon Ions. Genetics 2003, 1455, 1449–1455. [Google Scholar]
- Shikazono, N.; Suzuki, C.; Kitamura, S.; Watanabe, H.; Tano, S. Analysis of mutations induced by carbon ions in Arabidopsis thaliana. J. Exp. Bot. 2005, 56, 587–596. [Google Scholar] [CrossRef] [PubMed]
- Belfield, E.J.; Gan, X.; Mithani, A.; Brown, C.; Jiang, C.; Franklin, K.; Alvey, E.; Wibowo, A.; Jung, M.; Bailey, K.; et al. Genome-wide analysis of mutations in mutant lineages selected following fast-neutron irradiation mutagenesis of Arabidopsis thaliana. Genome Res. 2012, 22, 1306–1315. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Du, Y.; Luo, S.; Li, X.; Yang, J.; Cui, T.; Li, W.; Yu, L. Identification of Substitutions and Small Insertion-Deletions Induced by Carbon-Ion Beam Irradiation in Arabidopsis thaliana. Front. Plant Sci. 2017, 8, 1851. [Google Scholar] [CrossRef] [Green Version]
- Li, G.; Chern, M.; Jain, R.; Martin, J.A.; Schackwitz, W.S.; Jiang, L.; Vega-Sánchez, M.E.; Lipzen, A.M.; Barry, K.W.; Schmutz, J.; et al. Genome-wide sequencing of 41 rice (Oryza sativa L.) mutated lines reveals diverse mutations induced by fast neutron irradiation. Mol. Plant 2016, 9, 1078–1081. [Google Scholar] [CrossRef]
- Li, G.; Jain, R.; Chern, M.; Pham, N.T.; Martin, J.A.; Wei, T. The sequences of 1504 mutants in the model rice variety Kitaake facilitate rapid functional genomic studies. Plant Cell 2017, 29, 1218–1231. [Google Scholar]
- Hase, Y.; Yoshihara, R.; Nozawa, S.; Narumi, I. Fundamental and Molecular Mechanisms of Mutagenesis Mutagenic effects of carbon ions near the range end in plants. Mutat. Res./Fundam. Mol. Mech. Mutagen. 2012, 731, 41–47. [Google Scholar] [CrossRef]
- Kazama, Y.; Saito, H.; Yamamoto, Y.Y.; Hayashi, Y.; Ichida, H.; Ryuto, H.; Fukunishi, N.; Abe, T. LET-dependent effects of heavy-ion beam irradiation in Arabidopsis thaliana. Plant Biotechnol. 2008, 117, 113–117. [Google Scholar] [CrossRef]
- Yoshihara, R.; Nozawa, S.; Hase, Y.; Narumi, I.; Hidema, J.; Sakamoto, A.N. Mutational effects of γ-rays and carbon ion beams on Arabidopsis seedlings. J. Radiat. Res. 2013, 54, 1050–1056. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ballarini, F.; Alloni, D.; Facoetti, A.; Ottolenghi, A. Heavy-ion effects: From track structure to DNA and chromosome damage. New J. Phys. 2008, 10, 075008. [Google Scholar] [CrossRef]
- Hada, M.; Georgakilas, A.G. Formation of clustered DNA damage after high-LET irradiation: A Review. J. Radiat. Res. 2008, 49, 203–210. [Google Scholar] [CrossRef]
- Roots, R.; Okada, S. Estimation of Life times and diffusion distances of radicals DNA strand breaks involved in X-ray-induced or killing of mammalian cells. Radiat. Res. 1975, 320, 306–320. [Google Scholar] [CrossRef]
- Wallace, S.S. Enzymatic processing of radiation-induced free radical damage in DNA1. Radiat. Res. 1998, 150, S60–S79. [Google Scholar] [CrossRef]
- Kikuchi, S.; Saito, Y.; Ryuto, H.; Fukunishi, N.; Abe, T.; Tanaka, H.; Tsujimoto, H. Mechanisms of mutagenesis effects of heavy-ion beams on chromosomes of common wheat, Triticum aestivum. Mutagenes. Res. 2009, 669, 63–66. [Google Scholar] [CrossRef]
- Okamura, M. Wide variety of flower-color and -shape mutants regenerated from leaf cultures irradiated with ion beams. Nucl. Instrum. Methods Phys. Res. 2003, 206, 574–578. [Google Scholar] [CrossRef]
- Tanaka, A.; Kansai Photon Science Institute Kyoto, Japan. Personal communication, 2016.
- Vochita, G.; Focea-ghioc, R.; Creanga, D. Direct versus indirect radiation action in irradiated vegetal embryos. Cent. Eur. J. Biol. 2014, 9, 993–1003. [Google Scholar] [CrossRef]
- Kowyama, Y.; Saba, T.; Tsuji, T.; Kawase, T. Specific developmental stages of gametogenesis for radiosensitivity and mutagenesis in rice. Euphytica 1994, 80, 27–38. [Google Scholar] [CrossRef]
- Hase, Y.; Okamura, M.; Takeshita, D.; Narumi, I. Efficient induction of flower-color mutants by ion beam irradiation in petunia seedlings treated with high sucrose. Plant Biotechnol. 2010, 103, 99–103. [Google Scholar] [CrossRef]
- Hase, Y.; Akita, Y.; Kitamura, S.; Narumi, I.; Tanaka, A. Development of an efficient mutagenesis technique using ion beams: Toward more controlled mutation breeding. Plant Biotechnol. 2012, 200, 193–200. [Google Scholar] [CrossRef]
- Naito, K.; Kusaba, M.; Shikazono, N.; Takano, T.; Tanaka, A.; Tanisaka, T.; Nishimura, M. Transmissible and nontransmissible mutations induced by irradiating Arabidopsis thaliana pollen With λ-rays and carbon ions. Genetics 2005, 169, 881–889. [Google Scholar] [CrossRef]
- Song, Q.; Cannistraro, V.J.; Taylor, J. Synergistic modulation of cyclobutane pyrimidine dimer photoproduct formation and deamination at a TmCG site over a full helical DNA turn in a nucleosome core particle. Nucleic Acids Res. 2014, 42, 13122–13133. [Google Scholar] [CrossRef]
- Decottignies, A. Alternative end-joining mechanisms: A historical perspective. Front. Genet. 2013, 4, 48. [Google Scholar] [CrossRef]
- Ishii, K.; Kazama, Y.; Morita, R.; Hirano, T.; Ikeda, T. Linear energy transfer-dependent change in rice gene expression profile after heavy-ion beam irradiation. PLoS ONE 2016, 11, e0160061. [Google Scholar] [CrossRef]
- Henry, I.M.; Nagalakshm, U.; Lieberman, M.C.; Ngo, K.J.; Krasileva, K.V.; Vasquez-Gross, H.; Akhunova, A.; Akhunov, E.; Dubcovsky, J.; Tai, T.H.; et al. Efficient genome-wide detection and cataloging of EMS-induced mutations using exome capture and next-generation sequencing. Plant Cell 2014, 26, 1382–1397. [Google Scholar] [CrossRef] [PubMed]
- Kim, W.J.; Ryu, J.; Im, J.; Kim, S.H.; Ikeda, T. Molecular characterization of proton beam-induced mutations in soybean using genotyping-by-sequencing. Mol. Genet. Genom. 2018, 293, 1169–1180. [Google Scholar] [CrossRef]
Figure 1.
General procedure for the mutation breeding of seed-propagated crops. The M1 plant is chimeric for each induced mutation because it originates from the irradiated seeds and comprises multiple cells with a diverse mutation spectrum. Selections based on the phenotype or DNA sequences are usually performed with M2 populations, which do not contain any chimeric individuals. In a forward genetics approach, individuals with a desired phenotype are selected in a population, after which the gene responsible for the phenotype is studied. In reverse genetics approaches, lines with mutated targeted genes are selected, after which the resulting phenotype is investigated.
Figure 1.
General procedure for the mutation breeding of seed-propagated crops. The M1 plant is chimeric for each induced mutation because it originates from the irradiated seeds and comprises multiple cells with a diverse mutation spectrum. Selections based on the phenotype or DNA sequences are usually performed with M2 populations, which do not contain any chimeric individuals. In a forward genetics approach, individuals with a desired phenotype are selected in a population, after which the gene responsible for the phenotype is studied. In reverse genetics approaches, lines with mutated targeted genes are selected, after which the resulting phenotype is investigated.

Figure 2.
Frequency and spectrum of DNA mutations in mutant lines derived by irradiation. (a) Mutation frequency (× 10−7/bp) estimated for the M1 generation. (b) Proportion of single base substitutions (SBSs) and InDels. (c) Spectrum of SBSs. The genome information from the studies listed in Table 1 was analyzed. For fast-neutron-irradiated rice, the genome information from Li et al. (2017) [27] was used, but that from Li et al. (2016) [25] was not because the materials in Li et al. (2016) [25] were included in Li et al. (2017) [27]. The results of Du et al (2017) [24] were omitted from (b) because large InDels were not analyzed. Regarding the argon beam irradiation (290 KeV μm−1), the genome information from Kazama et al. (2017) was used, but that from Hirano et al. (2015) [11] was not because Kazama et al. (2017) [12] analyzed more samples; the irradiation conditions were identical in both studies.
Figure 2.
Frequency and spectrum of DNA mutations in mutant lines derived by irradiation. (a) Mutation frequency (× 10−7/bp) estimated for the M1 generation. (b) Proportion of single base substitutions (SBSs) and InDels. (c) Spectrum of SBSs. The genome information from the studies listed in Table 1 was analyzed. For fast-neutron-irradiated rice, the genome information from Li et al. (2017) [27] was used, but that from Li et al. (2016) [25] was not because the materials in Li et al. (2016) [25] were included in Li et al. (2017) [27]. The results of Du et al (2017) [24] were omitted from (b) because large InDels were not analyzed. Regarding the argon beam irradiation (290 KeV μm−1), the genome information from Kazama et al. (2017) was used, but that from Hirano et al. (2015) [11] was not because Kazama et al. (2017) [12] analyzed more samples; the irradiation conditions were identical in both studies.

Figure 3.
Effects of DNA mutations on gene function. (a) Average number of genes with homozygous mutations estimated for the M2 generation. (b) Average number of genes with homozygous or heterozygous mutations estimated for the M2 generation. (c) Classification of mutations according to their effects on gene function. Only studies that provided enough information for analyses were included. Regarding the argon beam irradiation (290 KeV μm−1), the data from Hirano et al. [11] were used in the analysis because one mutant line included in the study by Kazama et al. [12] contained extraordinarily long deleted sequences and included 852 genes.
Figure 3.
Effects of DNA mutations on gene function. (a) Average number of genes with homozygous mutations estimated for the M2 generation. (b) Average number of genes with homozygous or heterozygous mutations estimated for the M2 generation. (c) Classification of mutations according to their effects on gene function. Only studies that provided enough information for analyses were included. Regarding the argon beam irradiation (290 KeV μm−1), the data from Hirano et al. [11] were used in the analysis because one mutant line included in the study by Kazama et al. [12] contained extraordinarily long deleted sequences and included 852 genes.


{kind=link}
{kind=link}
{kind=link}
Table 1.
Investigations in which mutagen characteristics were analyzed based on the whole-genome sequencing of mutant samples. EMS: ethyl methanesulfonate.
Table 1.
Investigations in which mutagen characteristics were analyzed based on the whole-genome sequencing of mutant samples. EMS: ethyl methanesulfonate.
| Mutagen | Dosage | Species and Tissue | Biological Effect in M1 | Generation of the Sequenced Samples | Number of Sequenced Samples | Selection of Samples by Phenotype w | Reference | |
|---|---|---|---|---|---|---|---|---|
| Fast neutrons | 60 Gy | Arabidopsis thaliana (Ler), seeds | - z | M3 | 6 | Yes | [23] | |
| 20 Gy | Oryza sativa (cv. X. Kitaake), seeds | 81% fertility | M2 | 1370 | No | [27] | ||
| M3 | 134 | |||||||
| Ion beams | Argon ion beams (290 KeV μm−1) | 50 Gy | Arabidopsis thaliana (Col-0), seeds | 95% survival | M2 (pooled M3) y | 3 | Yes | [11] |
| 8 | ||||||||
| Carbon ion beams (30 KeV μm−1) | 400 Gy | 95% survival | M2 (pooled M3) | 8 | Yes | [12] | ||
| Carbon ion beams (50 KeV μm−1) | 200 Gy | - | M3 | 11 | Yes/No v | [24] | ||
| Carbon ion beams (107 KeV μm−1) | 125 Gy | 50% Dq x | M2 | 6 | No | [27] | ||
| 175 Gy | 75% Dq | 6 | ||||||
| 20 Gy | Arabidopsis thaliana (Col-0), 7-day-old seedlings | 50% Dq | 6 | |||||
| 30 Gy | 75% Dq | 6 | ||||||
| Gamma rays | 300 Gy | Solanum lycopersicum (cv. Micro- Tom), seeds | - | M3 | 3 | No | [7] | |
| EMS | 1.0% | - | 4 | |||||
Z Biological effect in the M1 generation was not described in the original articles. y M3 individuals from a single M2 line were pooled for a sequencing analysis. The sequence of the M3 pool was used to determine the M2 genotype. x Dose for the shoulder in the dose-survival rate curve. W The cases in which samples selected based on specific mutant phenotypes or selected randomly were used for whole-genome sequencing (WGS) were indicated as ‘Yes’ or ‘No’, respectively. v Both samples that showed visible or inconspicuous mutant phenotypes were used for WGS.
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Jo, Y.D.; Kim, J.-B. Frequency and Spectrum of Radiation-Induced Mutations Revealed by Whole-Genome Sequencing Analyses of Plants. Quantum Beam Sci. 2019, 3, 7. https://0-doi-org.brum.beds.ac.uk/10.3390/qubs3020007
AMA Style
Jo YD, Kim J-B. Frequency and Spectrum of Radiation-Induced Mutations Revealed by Whole-Genome Sequencing Analyses of Plants. Quantum Beam Science. 2019; 3(2):7. https://0-doi-org.brum.beds.ac.uk/10.3390/qubs3020007
Chicago/Turabian StyleJo, Yeong Deuk, and Jin-Baek Kim. 2019. "Frequency and Spectrum of Radiation-Induced Mutations Revealed by Whole-Genome Sequencing Analyses of Plants" Quantum Beam Science 3, no. 2: 7. https://0-doi-org.brum.beds.ac.uk/10.3390/qubs3020007