
Influence of Scale Effect of Canopy Projection on Understory Microclimate in Three Subtropical Urban Broad-Leaved Forests

Abstract
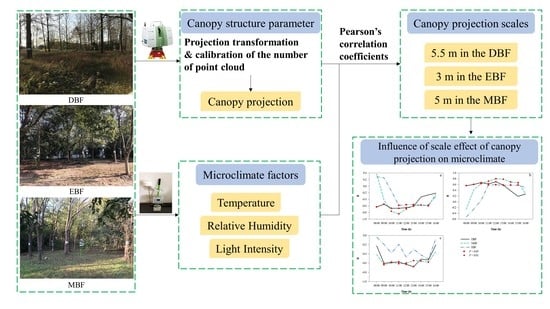
:
1. Introduction
2. Materials and Methods
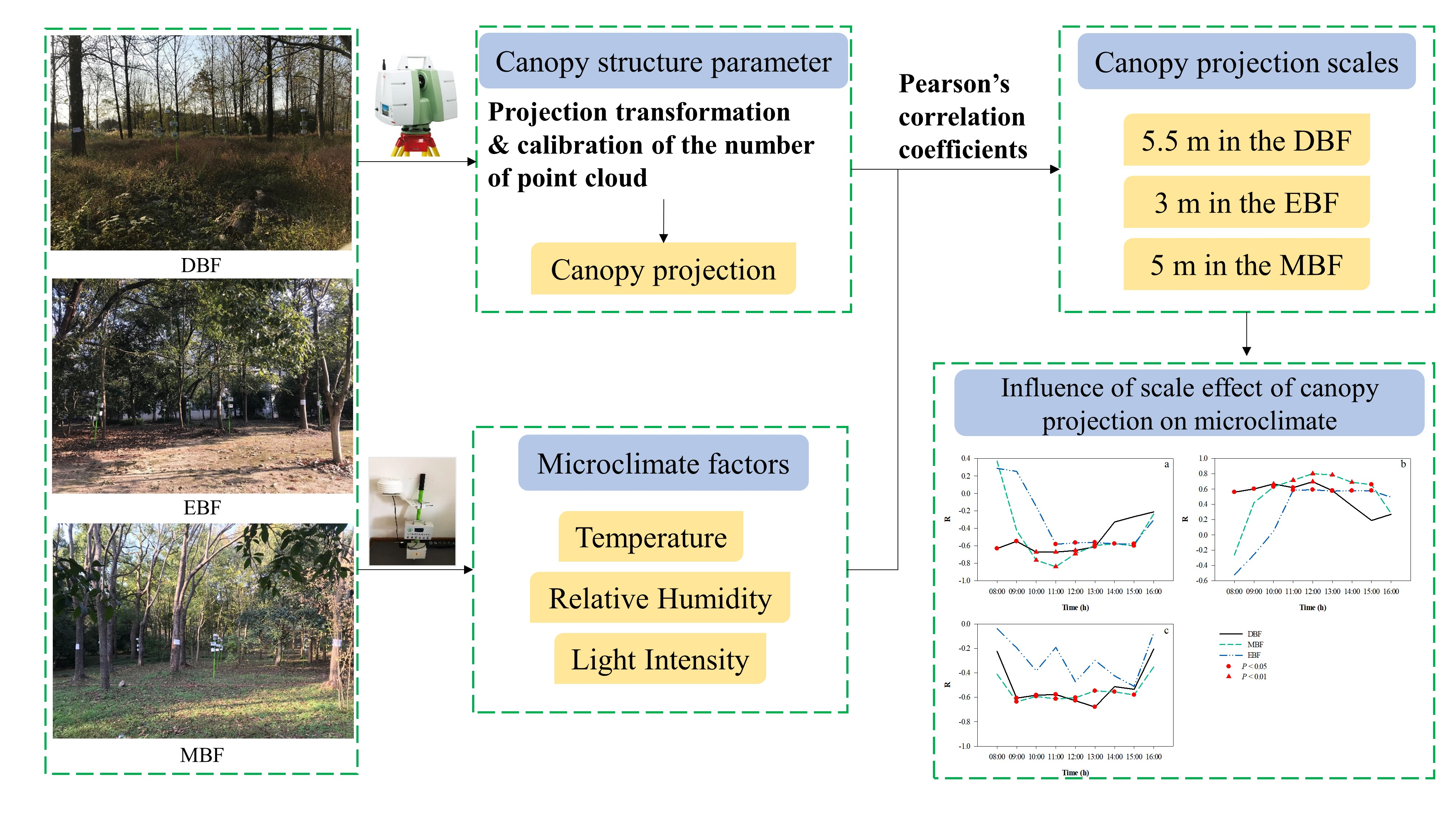
2.1. Study Area
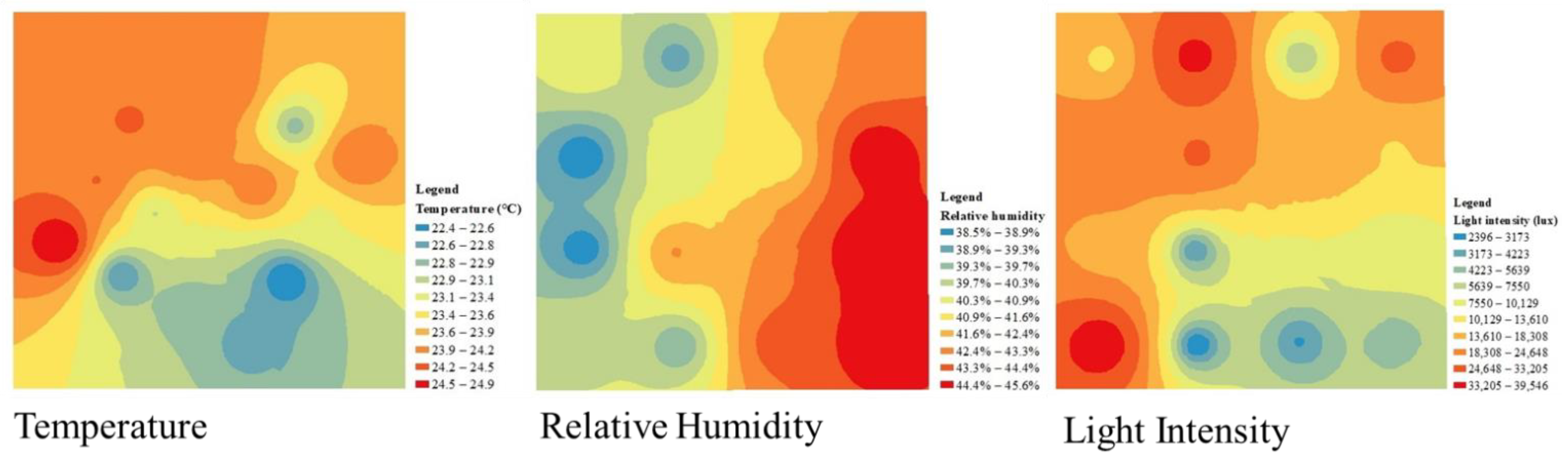
2.2. Temperature, Relative Humidity, and Light Intensity Data
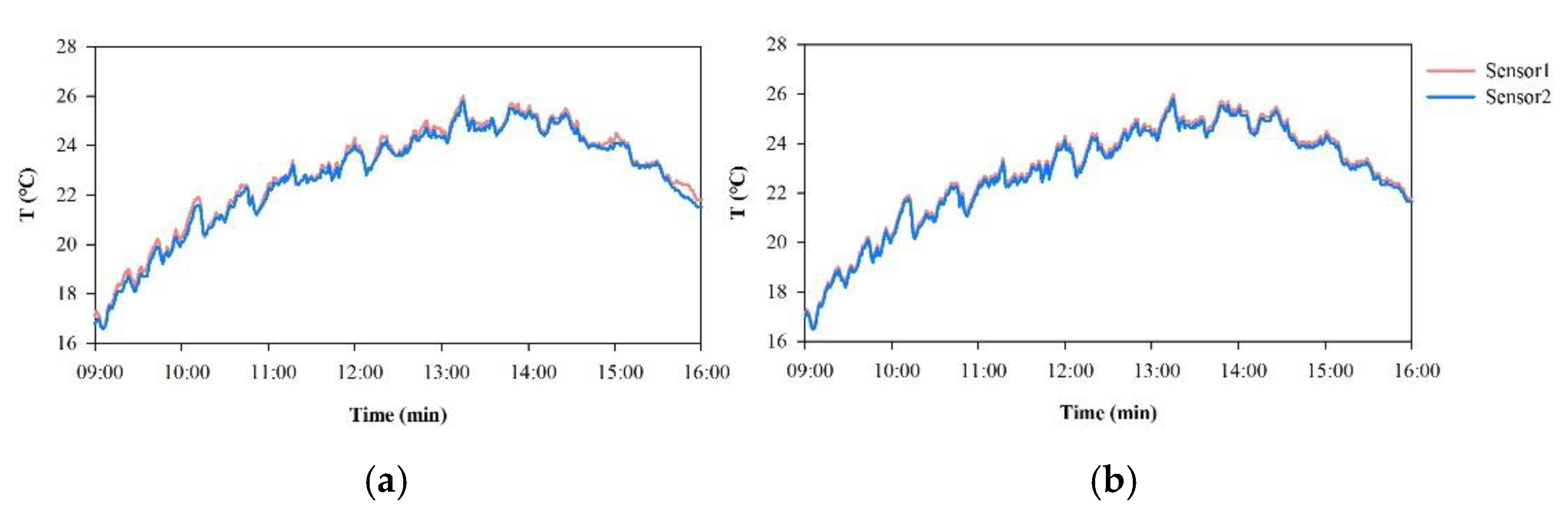
Processing of T, RH, LI Data
2.3. Point Cloud Data
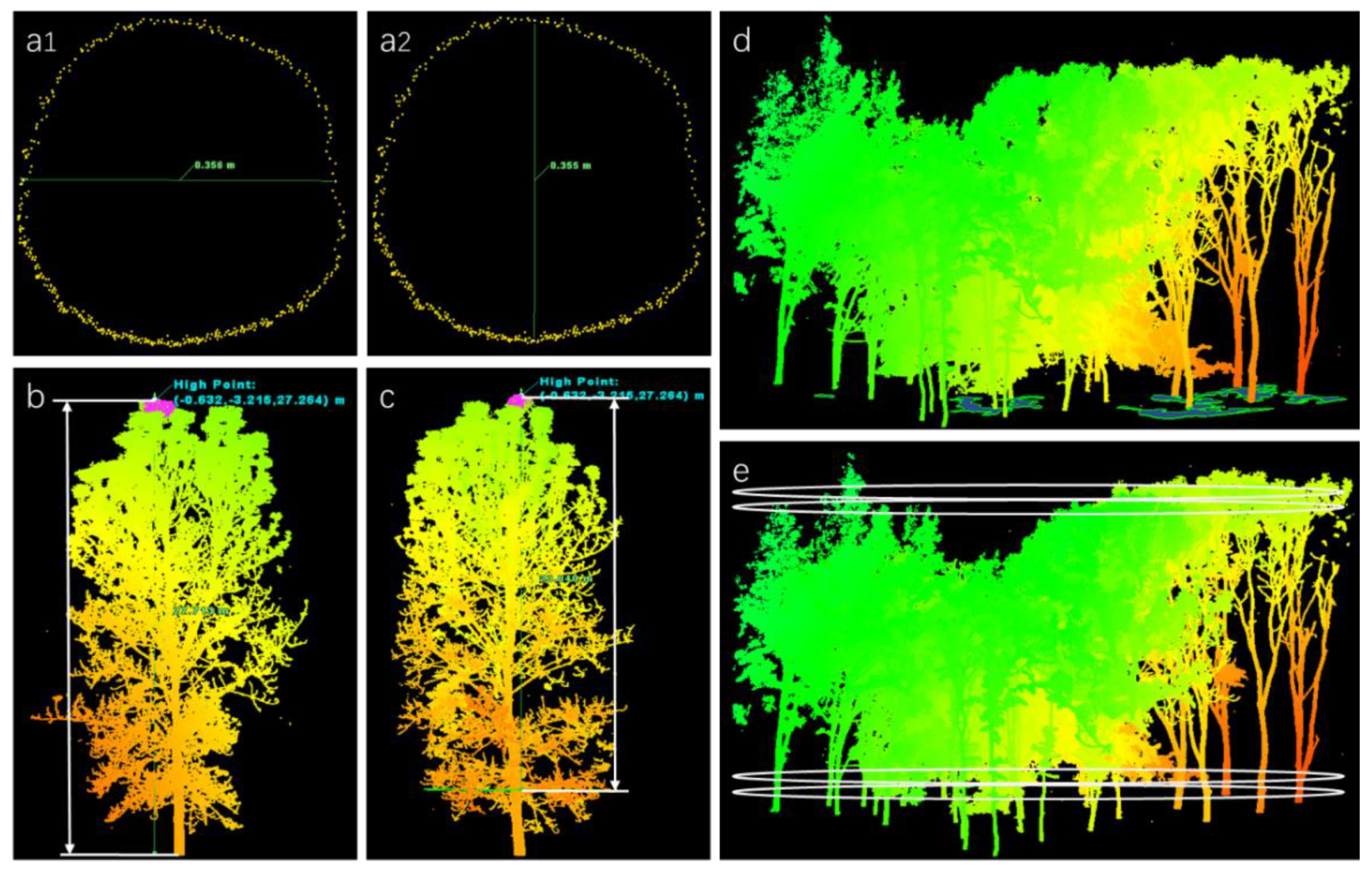
2.3.1. DBH, TH, CT, and CCA Extraction
2.3.2. CV Extraction
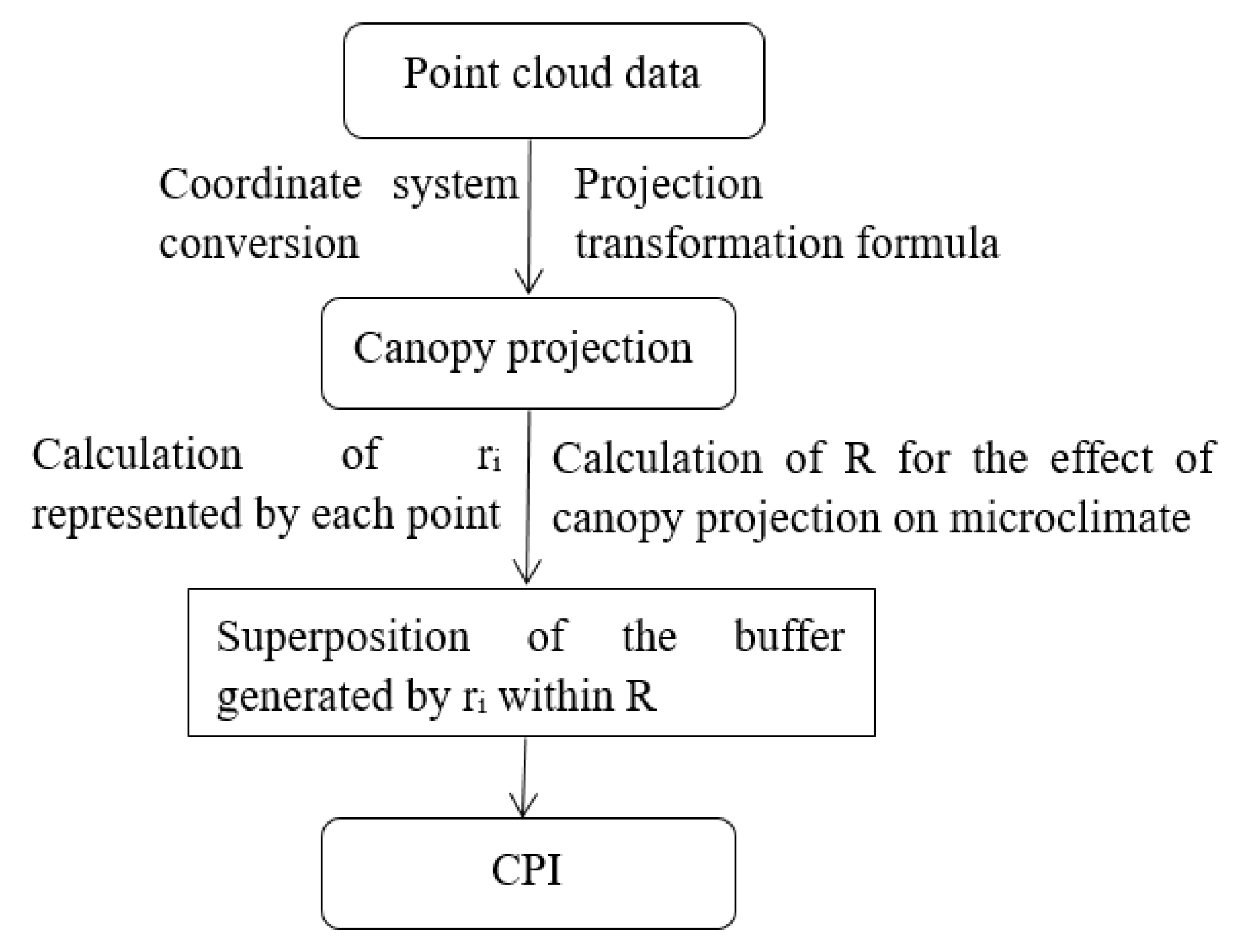
2.3.3. CPI Extraction
- The coordinates (X, Y, and Z) of each point were determined by the position of a TLS device. For the convenience of calculation, we obtained the azimuth angle of the forests through the electronic total station, and then the current coordinate system was converted to the geodetic coordinate system with the center of the sample plot as the origin.
- The point cloud data of the canopy projection in the forest were generated from 8:00 to 16:00 using the projection transformation formula (Equations (6)–(10)).
- Because the surrounding trees also affected the sample plot, the point cloud data outside the plot were removed according to the boundary of the plot after the projection transformation to obtain point cloud data of the canopy projection at different times.
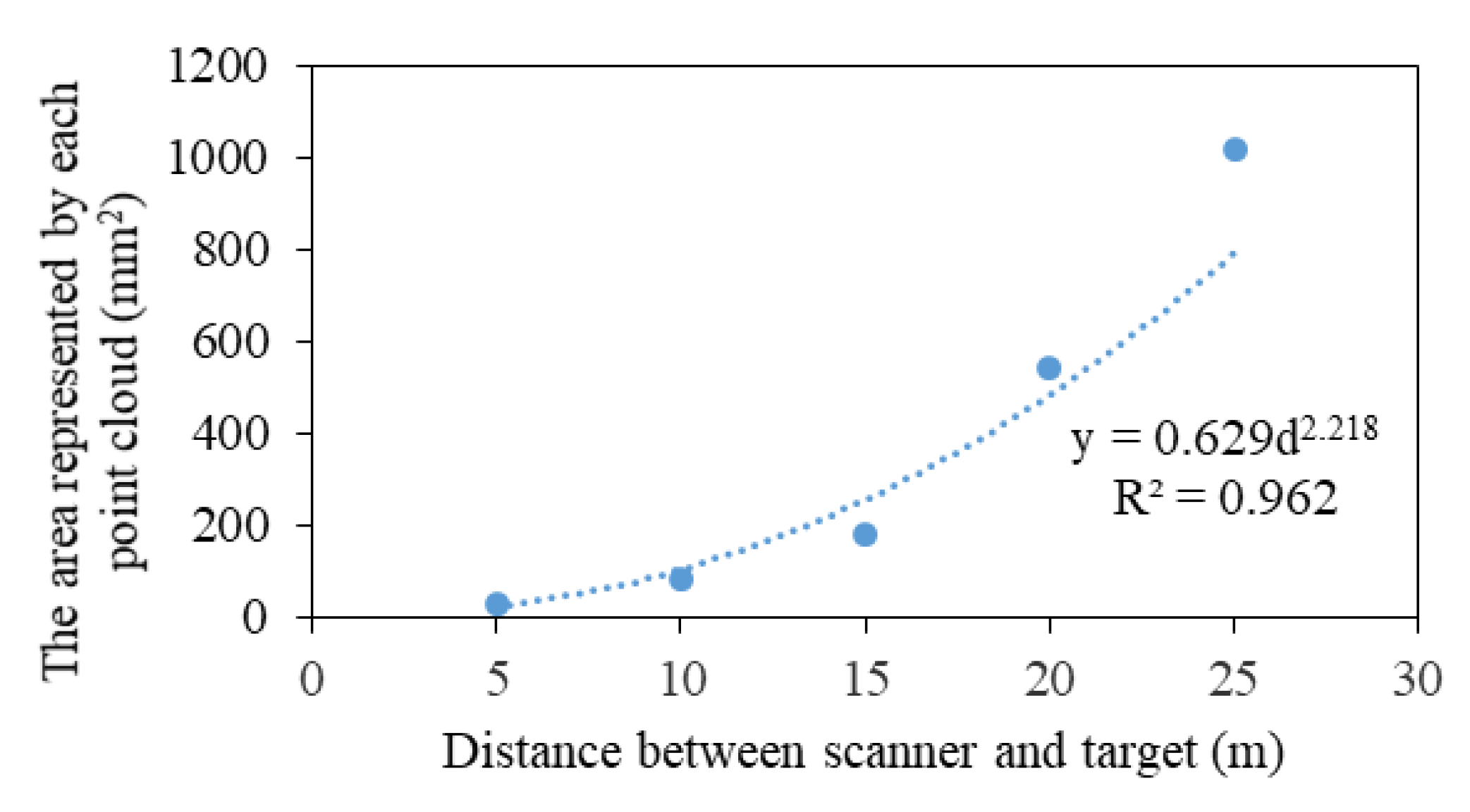
- The attribute of each point has four fields (, , , ), and the radius represented by each point was obtained according to the formula of area and radius (Equation (11)). The CPI was calculated by superimposing the buffers.
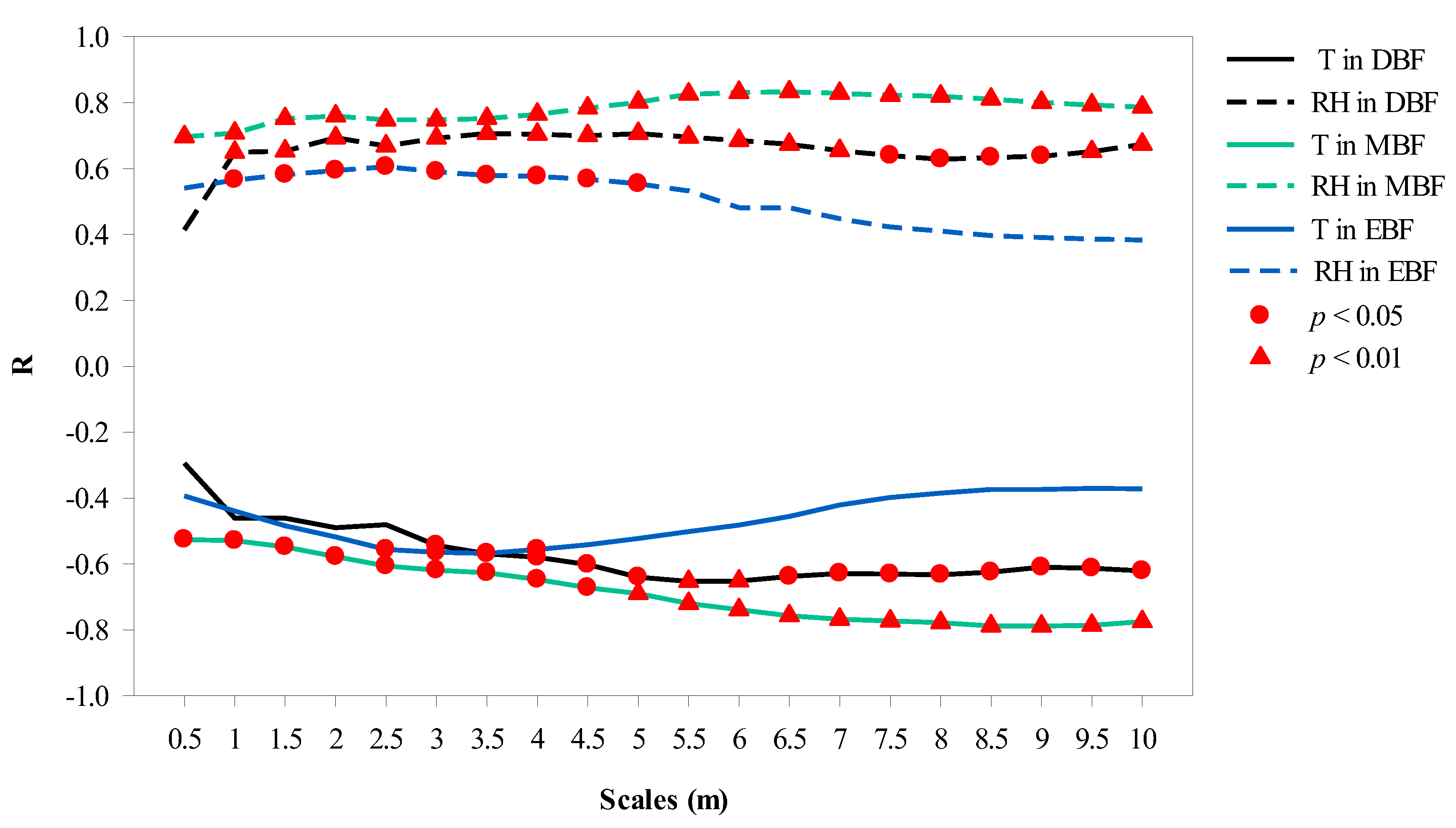
- To explore which buffer provided the most relevant information on T and RH, we compared 20 different scales ranging from 0.5 m to 10 m, with an interval of 0.5 m at 12:00, when the solar altitude angle was the largest. The radius of the sensor probe (13 mm) was used as the buffer radius to describe the effects of canopy projection on LI, which was attributed to the fact that the light sensor measures the value of the current position.
- Based on step five, the correlations between CPI and the microclimate factors in each forest were calculated from 08:00 to 16:00.
2.4. Canopy Image Data
Leaf Area Index (LAI) and CC Extraction
2.5. Statistical Analyses
3. Results
3.1. Canopy Structural Characteristics
3.2. Canopy Projection Scales
3.3. Hourly Variations in CPI and Microclimate Factors
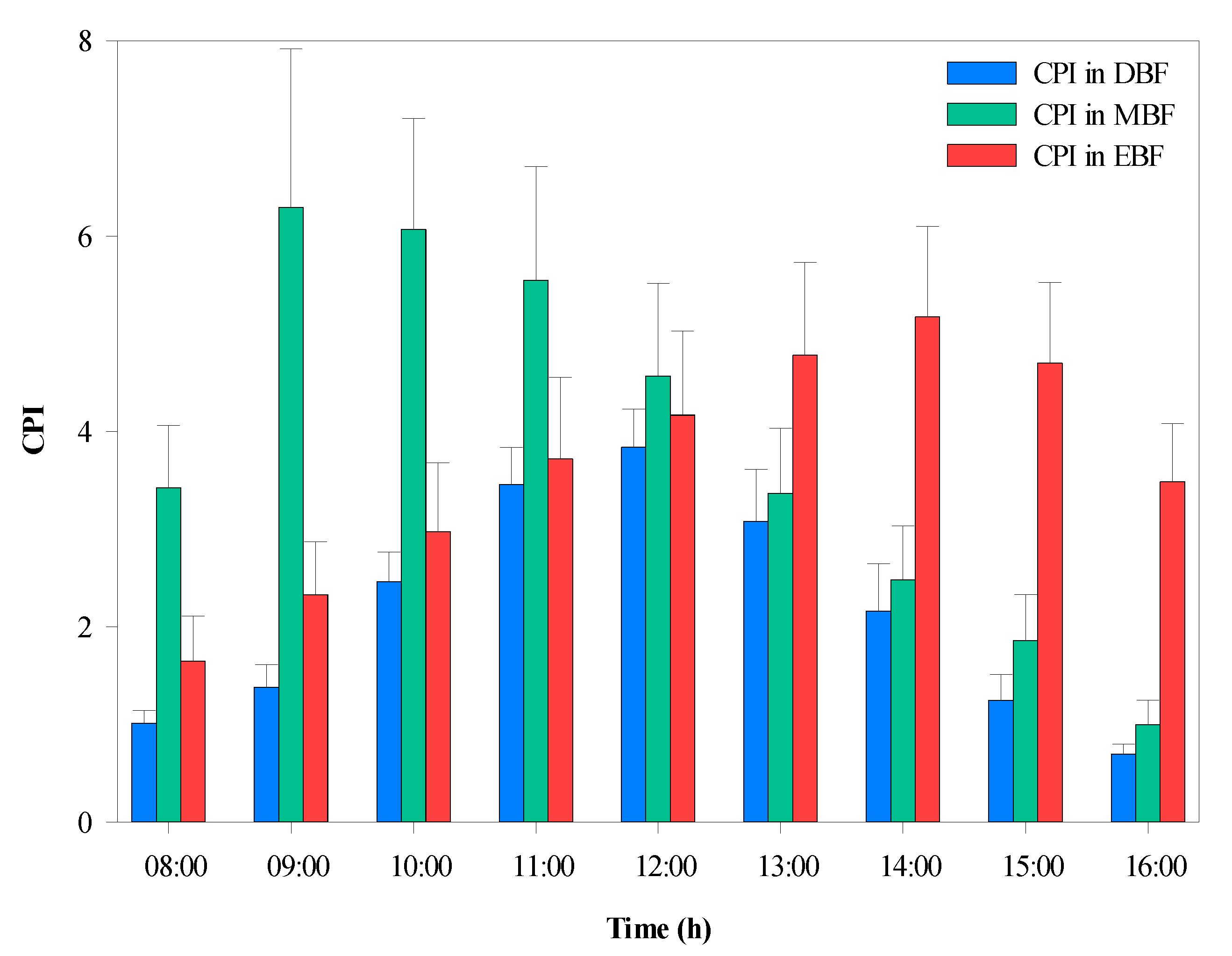
3.3.1. Hourly Variation in CPI
3.3.2. Hourly Variation in the Microclimate Factors
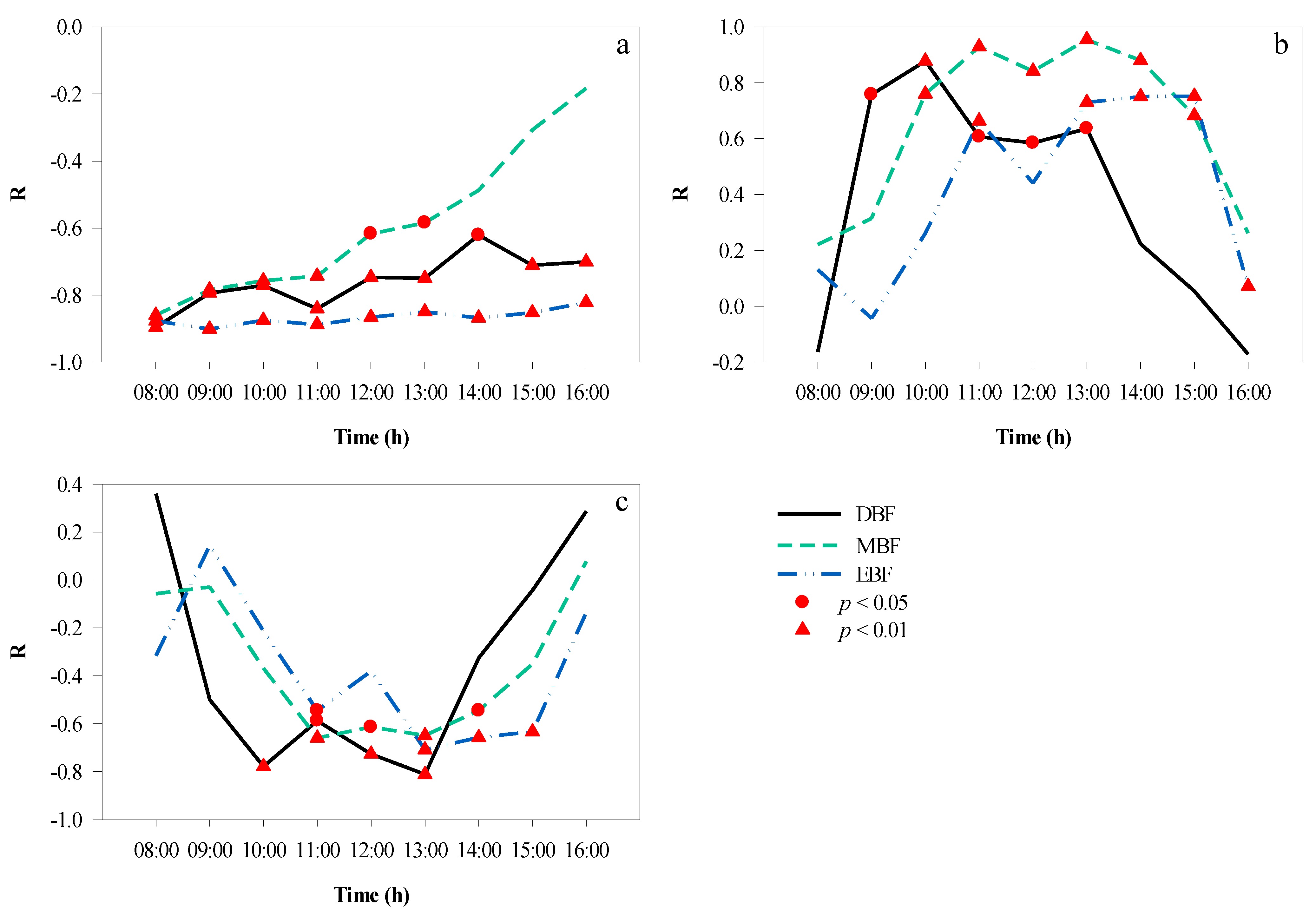
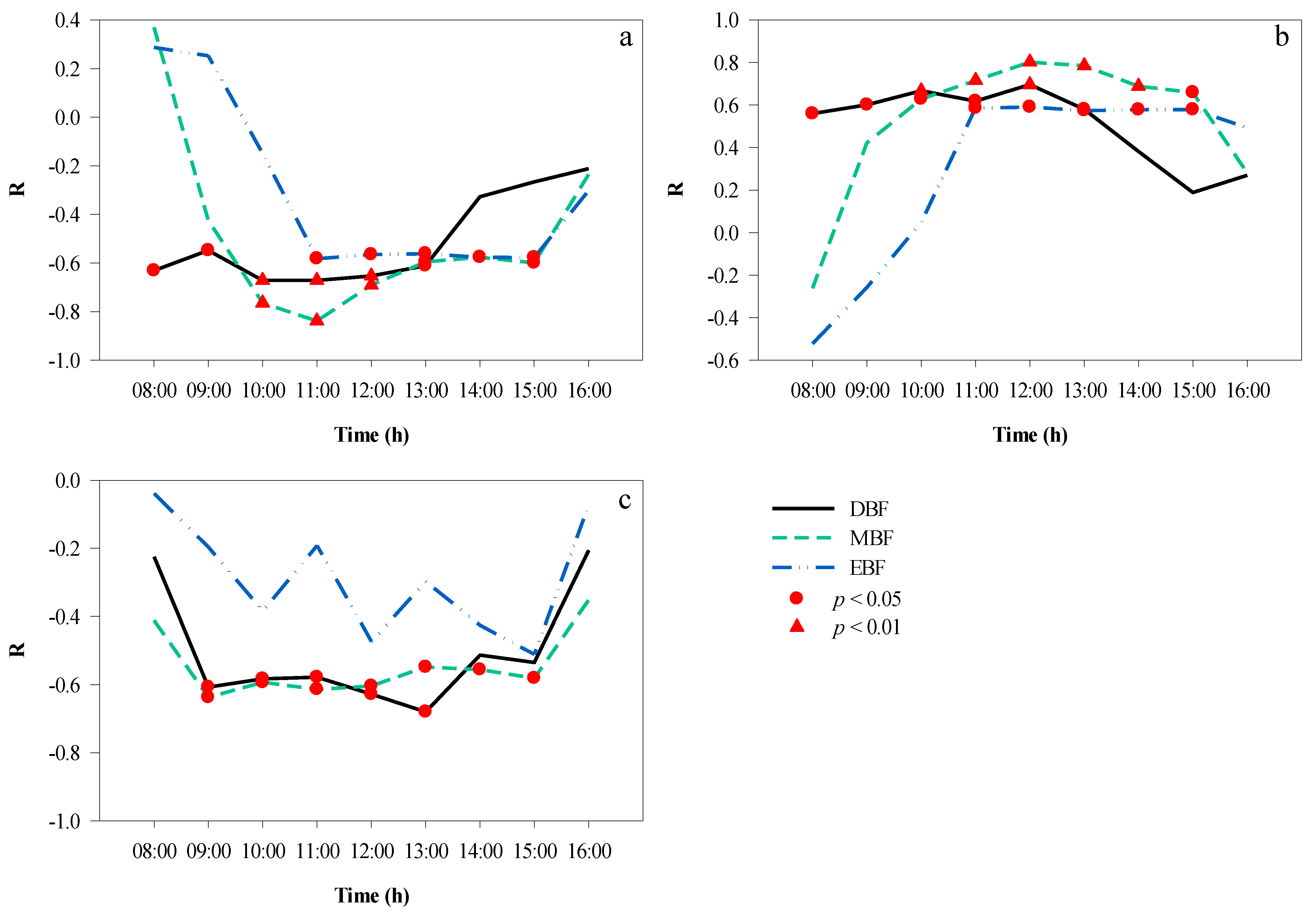
3.4. Correlation Analyses between Microclimate Factors
3.5. Correlation Analyses between the CPI and Microclimate Factors
4. Discussion
4.1. Scale Effect of Canopy Projection
4.2. Correlation of Microclimate Factors in Different Urban Forests
4.3. Effect of Canopy Projection on the Microclimate
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Gaudio, N.; Gendre, X.; Saudreau, M.; Seigner, V.; Balandier, P. Impact of tree canopy on thermal and radiative microclimates in a mixed temperate forest: A new statistical method to analyse hourly temporal dynamics. Agric. For. Meteorol. 2017, 237, 71–79. [Google Scholar] [CrossRef] [Green Version]
- Bellard, C.; Bertelsmeier, C.; Leadley, P.; Thuiller, W.; Courchamp, F. Impacts of climate change on the future of biodiversity. Ecol. Lett. 2012, 15, 365–377. [Google Scholar] [CrossRef] [Green Version]
- Hofmeister, J.; Hošek, J.; Brabec, M.; Střalková, R.; Mýlová, P.; Bouda, M.; Pettit, J.L.; Rydval, M.; Svoboda, M. Microclimate edge effect in small fragments of temperate forests in the context of climate change. For. Ecol. Manag. 2019, 448, 48–56. [Google Scholar] [CrossRef]
- Atkin, O.K.; Bloomfield, K.J.; Reich, P.B.; Tjoelker, M.G.; Asner, G.P.; Bonal, D.; Bönisch, G.; Bradford, M.G.; Cernusak, L.A.; Cosio, E.G.; et al. Global variability in leaf respiration in relation to climate, plant functional types and leaf traits. New Phytol. 2015, 206, 614–636. [Google Scholar] [CrossRef] [Green Version]
- Bertrand, R.; Lenoir, J.; Piedallu, C.; Dillon, G.R.; De Ruffray, P.; Vidal, C.; Pierrat, J.-C.; Gégout, J.-C. Changes in plant community composition lag behind climate warming in lowland forests. Nature 2011, 479, 517–520. [Google Scholar] [CrossRef]
- Ali, S.B.; Patnaik, S. Assessment of the impact of urban tree canopy on microclimate in Bhopal: A devised low-cost traverse methodology. Urban Clim. 2019, 27, 430–445. [Google Scholar] [CrossRef]
- Baker, T.P.; Moroni, M.T.; Hunt, M.A.; Worledge, D.; Mendham, D.S. Temporal, environmental and spatial changes in the effect of windbreaks on pasture microclimate. Agric. For. Meteorol. 2021, 297, 108265. [Google Scholar] [CrossRef]
- Meili, N.; Manoli, G.; Burlando, P.; Carmeliet, J.; Chow, W.T.L.; Coutts, A.M.; Roth, M.; Velasco, E.; Vivoni, E.R.; Fatichi, S. Tree effects on urban microclimate: Diurnal, seasonal, and climatic temperature differences explained by separating radiation, evapotranspiration, and roughness effects. Urban For. Urban Green. 2021, 58, 126970. [Google Scholar] [CrossRef]
- Greiser, C.; Meineri, E.; Luoto, M.; Ehrlén, J.; Hylander, K. Monthly microclimate models in a managed boreal forest landscape. Agric. For. Meteorol. 2018, 250, 147–158. [Google Scholar] [CrossRef]
- Karki, U.; Goodman, M.S. Microclimatic differences between mature loblolly-pine silvopasture and open-pasture. Agrofor. Syst. 2015, 89, 319–325. [Google Scholar] [CrossRef]
- Von Arx, G.; Dobbertin, M.; Rebetez, M. Spatio-temporal effects of forest canopy on understory microclimate in a long-term experiment in Switzerland. Agric. For. Meteorol. 2012, 166, 144–155. [Google Scholar] [CrossRef]
- Frenne, P.D.; Zellweger, F.; Rodríguez-Sánchez, F.; Scheffers, B.R.; Lenoir, J. Global buffering of temperatures under forest canopies. Nat. Ecol. Evol. 2019, 3, 744–749. [Google Scholar] [CrossRef] [PubMed]
- Frey, S.; Hadley, A.S.; Johnson, S.L.; Schulze, M.; Jones, J.A.; Betts, M.G. Spatial models reveal the microclimatic buffering capacity of old-growth forests. Sci. Adv. 2016, 2, e1501392. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Müller, C.M. Critical forest age thresholds for the diversity of lichens, molluscs and birds in beech (Fagus sylvatica L.) dominated forests. Ecol. Indic. 2009, 9, 922–932. [Google Scholar] [CrossRef]
- Schuster, M.J.; Wragg, P.D.; Williams, L.J.; Butler, E.E.; Stefanski, A.; Reich, P.B. Phenology matters: Extended spring and autumn canopy cover increases biotic resistance of forests to invasion by common buckthorn (Rhamnus cathartica). For. Ecol. Manag. 2020, 464, 118067. [Google Scholar] [CrossRef]
- Georgi, N.J.; Zafiriadis, K. The impact of park trees on microclimate in urban areas. Urban Ecosyst. 2006, 9, 195–209. [Google Scholar] [CrossRef]
- Hardwick, S.R.; Toumi, R.; Pfeifer, M.; Turner, E.C.; Nilus, R.; Ewers, R.M. The relationship between leaf area index and microclimate in tropical forest and oil palm plantation: Forest disturbance drives changes in microclimate. Agric. For. Meteorol. 2015, 201, 187–195. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kong, F.H.; Yan, W.J.; Zheng, G.; Yin, H.W.; Cavan, G.; Zhan, W.F.; Zhang, N.; Cheng, L. Retrieval of three-dimensional tree canopy and shade using terrestrial laser scanning (TLS) data to analyze the cooling effect of vegetation. Agric. For. Meteorol. 2016, 217, 22–34. [Google Scholar] [CrossRef]
- Jung, M.C.; Dyson, K.; Alberti, M. Urban Landscape Heterogeneity Influences the Relationship between Tree Canopy and Land Surface Temperature. Urban For. Urban Green. 2021, 57, 126930. [Google Scholar] [CrossRef]
- Andrade, A.J.; Tomback, D.F.; Seastedt, T.R.; Mellmann-Brown, S. Soil moisture regime and canopy closure structure subalpine understory development during the first three decades following fire. For. Ecol. Manag. 2021, 483, 118783. [Google Scholar] [CrossRef]
- Kašpar, V.; Hederová, L.; Macek, M.; Müllerová, J.; Prošek, J.; Surový, P.; Wild, J.; Kopecký, M. Temperature buffering in temperate forests: Comparing microclimate models based on ground measurements with active and passive remote sensing. Remote Sens. Environ. 2021, 263, 112522. [Google Scholar] [CrossRef]
- Yurtseven, H.; Akgul, M.; Coban, S.; Gulci, S. Determination and accuracy analysis of individual tree crown parameters using UAV based imagery and OBIA techniques. Measurement 2019, 145, 651–664. [Google Scholar] [CrossRef]
- De Almeida, D.R.A.; Broadbent, E.N.; Ferreira, M.P.; Meli, P.; Zambrano, A.M.A.; Gorgens, E.B.; Resende, A.F.; de Almeida, C.T.; do Amaral, C.H.; Corte, A.P.D.; et al. Monitoring restored tropical forest diversity and structure through UAV-borne hyperspectral and lidar fusion. Remote Sens. Environ. 2021, 264, 112582. [Google Scholar] [CrossRef]
- Martin-Ducup, O.; Mofack, G., II; Wang, D.; Raumonen, P.; Ploton, P.; Sonké, B.; Barbier, N.; Couteron, P.; Pélissier, R. Evaluation of automated pipelines for tree and plot metric estimation from TLS data in tropical forest areas. Ann. Bot. 2021, in press. [Google Scholar] [CrossRef]
- Baker, T.P.; Jordan, G.J.; Baker, S.C. Microclimatic edge effects in a recently harvested forest: Do remnant forest patches create the same impact as large forest areas? For. Ecol. Manag. 2016, 365, 128–136. [Google Scholar] [CrossRef]
- Rahman, M.A.; Hartmann, C.; Moser-Reischl, A.; von Strachwitz, M.F.; Paeth, H.; Pretzsch, H.; Pauleit, S.; Rötzer, T. Tree cooling effects and human thermal comfort under contrasting species and sites. Agric. For. Meteorol. 2020, 287, 107947. [Google Scholar] [CrossRef]
- Shashua-Bar, L.; Potchter, O.; Bitan, A.; Boltansky, D.; Yaakov, Y. Microclimate modelling of street tree species effects within the varied urban morphology in the Mediterranean city of Tel Aviv, Israel. Int. J. Climatol. 2010, 30, 44–57. [Google Scholar] [CrossRef]
- Gómez-Muñoz, V.M.; Porta-Gándara, M.A.; Fernández, J.L. Effect of tree shades in urban planning in hot-arid climatic regions. Landsc. Urban Plan. 2010, 94, 149–157. [Google Scholar] [CrossRef]
- Akbari, H.; Kurn, D.M.; Bretz, S.E.; Hanford, J.W. Peak power and cooling energy savings of shade trees. Energy Build. 1997, 25, 139–148. [Google Scholar] [CrossRef] [Green Version]
- Berry, R.; Livesley, S.J.; Aye, L. Tree canopy shade impacts on solar irradiance received by building walls and their surface temperature. Build. Environ. 2013, 69, 91–100. [Google Scholar] [CrossRef]
- Gál, T.; Unger, J. A new software tool for SVF calculations using building and tree-crown databases. Urban Clim. 2014, 10, 594–606. [Google Scholar] [CrossRef] [Green Version]
- An, S.M.; Kim, B.S.; Lee, H.Y.; Kim, C.H.; Yi, C.Y.; Eum, J.H.; Woo, J.H. Three-dimensional point cloud based sky view factor analysis in complex urban settings. Int. J. Climatol. 2014, 34, 2685–2701. [Google Scholar] [CrossRef]
- Bramer, I.; Anderson, B.J.; Bennie, J.; Bladon, A.J.; De Frenne, P.; Hemming, D.; Hill, R.A.; Kearney, M.R.; Körner, C.; Korstjens, A.H.; et al. Advances in monitoring and modelling climate at ecologically relevant scales. Adv. Ecol. Res. 2018, 58, 101–161. [Google Scholar]
- Yokohari, M.; Brown, R.D.; Kato, Y.; Yamamoto, S. The cooling effect of paddy fields on summertime air temperature in residential Tokyo, Japan. Landsc. Urban Plan. 2001, 53, 17–27. [Google Scholar] [CrossRef]
- Maclean, I.M.D.; Mosedale, J.R.; Bennie, J.J. Microclima: An r package for modelling meso- and microclimate. Methods Ecol. Evol. 2019, 10, 280–290. [Google Scholar] [CrossRef] [Green Version]
- Rahman, M.A.; Moser, A.; Rötzer, T.; Pauleit, S. Within canopy temperature differences and cooling ability of Tilia cordata trees grown in urban conditions. Build. Environ. 2017, 114, 118–128. [Google Scholar] [CrossRef]
- Zhao, K.; Zhang, L.; Dong, J.; Wu, J.; Ye, Z.; Zhao, W.; Ding, L.; Fu, W. Risk assessment, spatial patterns and source apportionment of soil heavy metals in a typical Chinese hickory plantation region of southeastern China. Geoderma 2020, 360, 114011. [Google Scholar] [CrossRef]
- Wu, J.; Lin, H.; Meng, C.; Jiang, P.; Fu, W. Effects of intercropping grasses on soil organic carbon and microbial community functional diversity under Chinese hickory (Carya cathayensis Sarg.) stands. Soil Res. 2014, 52, 575–583. [Google Scholar] [CrossRef]
- Li, C.; Cai, Y.; Xiao, L.D.; Gao, X.Y.; Shi, Y.J.; Du, H.Q.; Zhou, Y.F.; Zhou, G.M. Effects of different planting approaches and site conditions on aboveground carbon storage along a 10-year chronosequence after moso bamboo reforestation. For. Ecol. Manag. 2021, 482, 118867. [Google Scholar] [CrossRef]
- Hangzhou Municipal Statistics Bureau. Hangzhou Statistical Yearbook; China Statistics Press: Beijing, China, 2017. (In Chinese) [Google Scholar]
- Chen, J.R. Research on 3-D Reconstruction from Point Cloud. MA Thesis, Wuhan University of Technology, Wuhan, China, 2011. (In Chinese). [Google Scholar]
- Bogdanovich, E.; Perez-Priego, O.; El-Madany, T.S.; Guderle, M.; Pacheco-Labrador, J.; Levick, S.R.; Moreno, G.; Carrara, A.; Pilar Martín, M.; Migliavacca, M. Using terrestrial laser scanning for characterizing tree structural parameters and their changes under different management in a Mediterranean open woodland. For. Ecol. Manag. 2021, 486, 118945. [Google Scholar] [CrossRef]
- Liu, K.; Shen, X.; Cao, L.; Wang, G.; Cao, F. Estimating forest structural attributes using UAV-LiDAR data in Ginkgo plantations. ISPRS J. Photogramm. Remote Sens. 2018, 146, 465–482. [Google Scholar] [CrossRef]
- Li, X.J.; Mao, F.J.; Du, H.Q.; Zhou, G.M.; Xu, X.J.; Han, N.; Sun, S.B.; Gao, G.L.; Chen, L. Assimilating leaf area index of three typical types of subtropical forest in China from MODIS time series data based on the integrated ensemble Kalman filter and PROSAIL model. ISPRS J. Photogramm. Remote Sens. 2017, 126, 68–78. [Google Scholar] [CrossRef]
- Zhao, Y.; Li, M.; Deng, J.Y.; Wang, B.T. Afforestation affects soil seed banks by altering soil properties and understory plants on the eastern Loess Plateau, China. Ecol. Indic. 2021, 126, 107670. [Google Scholar] [CrossRef]
- Hamada, S.; Ohta, T. Seasonal variations in the cooling effect of urban green areas on surrounding urban areas. Urban For. Urban Green. 2010, 9, 15–24. [Google Scholar] [CrossRef]
- Ma, S.; Concilio, A.; Oakley, B.; North, M.; Chen, J. Spatial variability in microclimate in a mixed-conifer forest before and after thinning and burning treatments. For. Ecol. Manag. 2010, 259, 904–915. [Google Scholar] [CrossRef]
- Sun, C.Y. A street thermal environment study in summer by the mobile transect technique. Theor. Appl. Climatol. 2011, 106, 433–442. [Google Scholar] [CrossRef]
- Ziter, C.D.; Pedersen, E.J.; Kucharik, C.; Turner, M.G. Scale-dependent interactions between tree canopy cover and impervious surfaces reduce daytime urban heat during summer. Proc. Natl. Acad. Sci. USA 2019, 116, 7575–7580. [Google Scholar] [CrossRef] [Green Version]
- Kovács, B.; Tinya, F.; Ódor, P. Stand structural drivers of microclimate in mature temperate mixed forests. Agric. For. Meteorol. 2017, 234, 11–21. [Google Scholar] [CrossRef] [Green Version]
- Wen, Q.; Han, W.; Ma, X.H.; Dang, Y.L.; Cai, Y.C.; Kong, K.K. Diurnal Variation of Net Photosynthetic Rate of Quercus in Autumn in Urumqi City and its Relationship with Physio-ecological Factors. J. Xinjiang Norm. Univ. Sci. Ed. 2019, 38, 6–12. (In Chinese) [Google Scholar]
- Anderson, D.B. Relative Humidity or Vapor Pressure Deficit. Ecology 1936, 17, 277–282. [Google Scholar] [CrossRef]
- Qin, Z. Cooling and Humidifying Effects and Driving Mechanisms of Beijing Olympic Forest Park in Summer. Ph.D. Thesis, Beijing Forestry University, Beijing, China, 2016. (In Chinese). [Google Scholar]
- Chakraborty, S.; Belekar, A.R.; Datye, A.; Sinha, N. Isotopic study of intraseasonal variations of plant transpiration: An alternative means to characterise the dry phases of monsoon. Sci. Rep. 2018, 8, 8647. [Google Scholar] [CrossRef] [Green Version]
- Madsen, T.V.; Breinholt, M. Effects of Air Contact on Growth, Inorganic Carbon Sources, and Nitrogen Uptake by an Amphibious Freshwater Macrophyte. Plant Physiol. 1995, 107, 149–154. [Google Scholar] [CrossRef] [Green Version]
- Barradas, V.L.; Tejeda-Martínez, A.; Jáuregui, E. Energy balance measurements in a suburban vegetated area in Mexico City. Atmos. Environ. 1999, 33, 4109–4113. [Google Scholar] [CrossRef]
- Lakhal, E. Effect of Harvest of Air Relative Humidity on Water and Heat Transfer in Soil with Crops under Arid Climatic Conditions. Int. J. Eng. Res. Appl. 2015, 5, 55–62. [Google Scholar]
- Gómez-Navarro, C.; Pataki, D.E.; Pardyjak, E.R.; Bowling, D.R. Effects of vegetation on the spatial and temporal variation of microclimate in the urbanized Salt Lake Valley. Agric. For. Meteorol. 2021, 296, 108211. [Google Scholar] [CrossRef]
- Shahidan, M.F.; Shariff, M.; Jones, P.; Salleh, E.; Abdullah, A.M. A comparison of Mesua ferrea L. and Hura crepitans L. for shade creation and radiation modification in improving thermal comfort. Landsc. Urban Plan. 2010, 97, 168–181. [Google Scholar] [CrossRef]
- Jiao, M.; Zhou, W.; Zheng, Z.; Wang, J.; Qian, Y. Patch size of trees affects its cooling effectiveness: A perspective from shading and transpiration processes. Agric. For. Meteorol. 2017, 247, 293–299. [Google Scholar] [CrossRef]
- Han, Y.J. Characteristics and diversity of understory vegetation communities in Shanghai urban forests. East China For. Manag. 2020, 34, 25–29. (In Chinese) [Google Scholar]
- Qin, Z.; Li, Z.; Cheng, F.; Chen, J.; Liang, B. Influence of canopy structural characteristics on cooling and humidifying effects of Populus tomentosa community on calm sunny summer days. Landsc. Urban Plan. 2014, 127, 75–82. [Google Scholar] [CrossRef]
- Tiţă, G.C.; Marcu, M.V.; Ignea, G.; Borz, S.A. Near the forest road: Small changes in air temperature and relative humidity in mixed temperate mountainous forests. Transp. Res. Part D Transp. Environ. 2019, 74, 82–92. [Google Scholar] [CrossRef]
- Zhao, W.J.; Shu, D.Y.; Li, C.L.; Cui, Y.C.; Liu, Y.H.; Wu, P.; Hou, Y.J.; Ding, F.J.; Academy, G.F. Relationships among transpiration, water consumption and environmental factors of Machilus ichangensis in karst forest. J. Cent. South Univ. For. Technol. 2019, 39, 108–155. (In Chinese) [Google Scholar] [CrossRef]
- Porté, A.; Huard, F.; Dreyfus, P. Microclimate beneath pine plantation, semi-mature pine plantation and mixed broadleaved-pine forest. Agric. For. Meteorol. 2004, 126, 175–182. [Google Scholar] [CrossRef]













{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Characteristics | EBF | DBF | MBF |
|---|---|---|---|
| Tree species | Cinnamomum camphora, Osmanthus fragrans, Castanopsis sclerophylla | Populus simonii, Yulania liliiflora | Koelreuteria paniculata, Cinnamomum camphora, Osmanthus fragrans, Yulania liliiflora |
| Shrub species | Ilex chinensis Sims, Symplocos sumuntia | Broussonetia papyrifera | Pittosporum tobira |
| Herbaceous plant species | Reineckea carnea | Gynostemma pentaphyllum, Pennisetum alopecuroides | Imperata cylindrica |
| Numbers | 28 | 8 | 33 |
| Average DBH/cm | 16.2 | 26.95 | 15.5 |
| Average TH/m | 10.9 | 16.4 | 13.2 |
| CCA/m2 | 357.28 | 201.22 | 364.39 |
| CT/m | 8.51 | 14.3 | 11.07 |
| CV/m3 | 1.13 | 1.06 | 1.58 |
| CC | 0.79 | 0.32 | 0.75 |
| LAI | 2.67 | 0.32 | 2.4 |
| Parameters | Temperature | Relative Humidity | Light Intensity |
|---|---|---|---|
| Range | −40–80 °C | 0–100% | 0–200,000 Lux |
| Accuracy | 0.1 °C | 0.3% | ±5% (25 °C) |
| Resolution | 0.1 °C | 0.1% | 1 Lux |
| Parameters | Value |
|---|---|
| Field-of-view | 360° × 270° |
| Scan rate | 25,000 pts/s |
| Range | 35 m @ ≥ 5% albedo |
| Accuracy of position | 6 mm |
| Accuracy of distance | 4 mm |
| Spot size | 4.5 mm (FWHH-based); 7 mm (Gaussian-based) |
| Minimum point spacing | <1 mm |
| Operating temperature | 0–40 °C |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gao, X.; Li, C.; Cai, Y.; Ye, L.; Xiao, L.; Zhou, G.; Zhou, Y. Influence of Scale Effect of Canopy Projection on Understory Microclimate in Three Subtropical Urban Broad-Leaved Forests. Remote Sens. 2021, 13, 3786. https://0-doi-org.brum.beds.ac.uk/10.3390/rs13183786
Gao X, Li C, Cai Y, Ye L, Xiao L, Zhou G, Zhou Y. Influence of Scale Effect of Canopy Projection on Understory Microclimate in Three Subtropical Urban Broad-Leaved Forests. Remote Sensing. 2021; 13(18):3786. https://0-doi-org.brum.beds.ac.uk/10.3390/rs13183786
Chicago/Turabian StyleGao, Xueyan, Chong Li, Yue Cai, Lei Ye, Longdong Xiao, Guomo Zhou, and Yufeng Zhou. 2021. "Influence of Scale Effect of Canopy Projection on Understory Microclimate in Three Subtropical Urban Broad-Leaved Forests" Remote Sensing 13, no. 18: 3786. https://0-doi-org.brum.beds.ac.uk/10.3390/rs13183786