Characterization of Canopy Layering in Forested Ecosystems Using Full Waveform Lidar

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Material and Methods
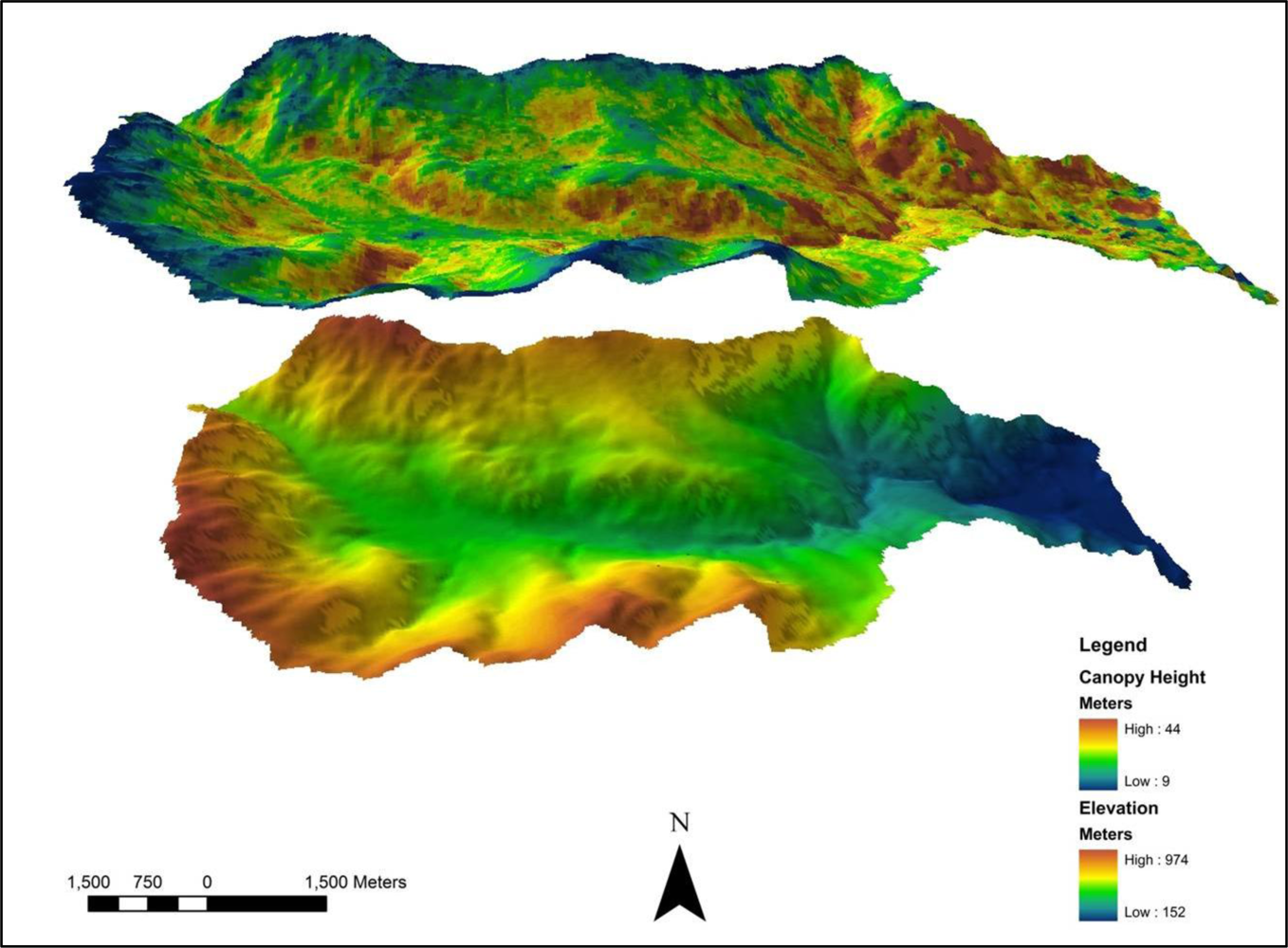
2.1. Study Area
2.2. Lidar Data
2.3. Characterization of Vertical Canopy Structure
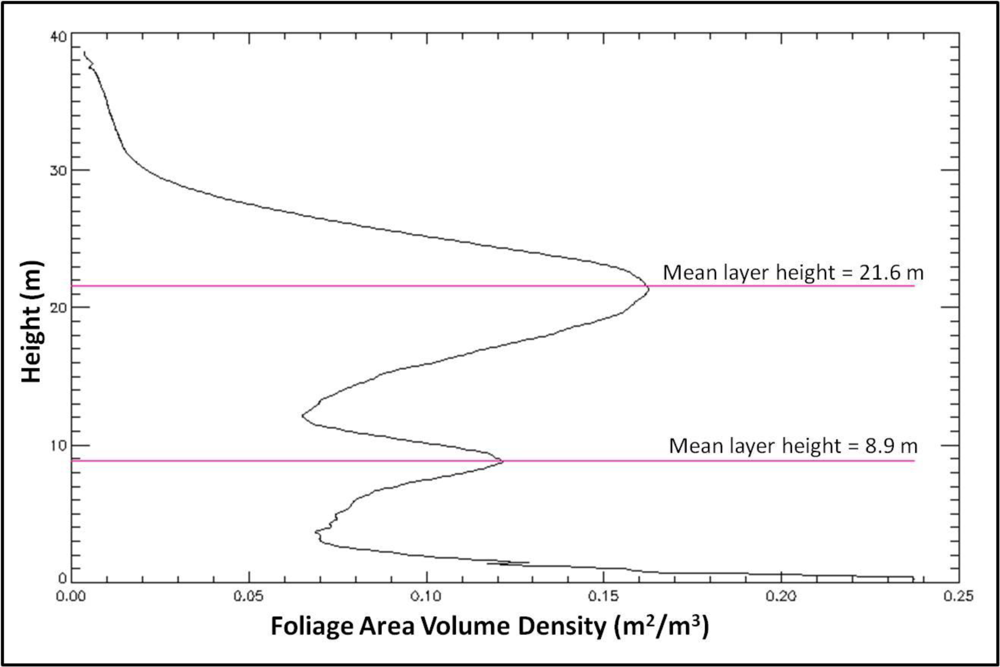
2.3.1. Foliage Area Profile
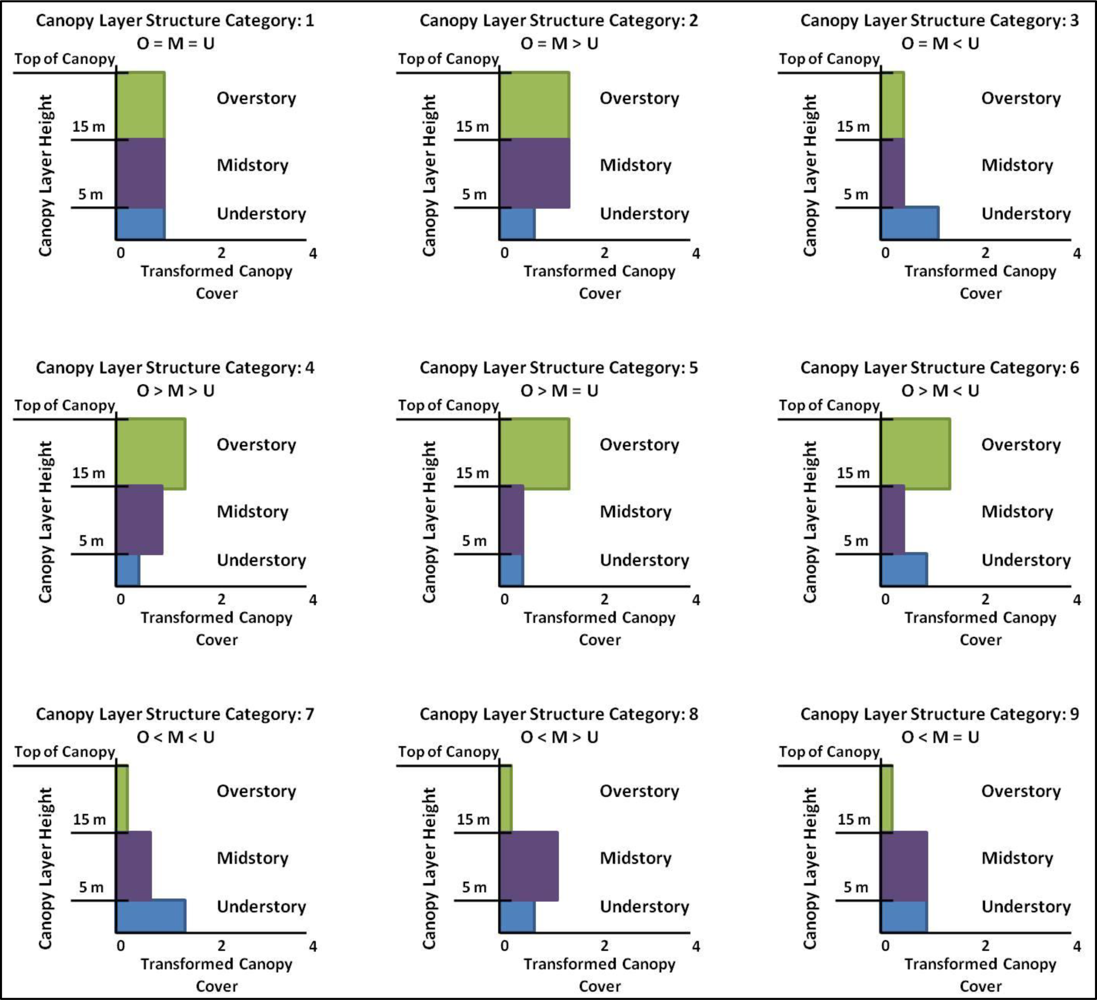
2.3.2. Canopy Layer Structure
2.3.3. Foliage Profile Layering
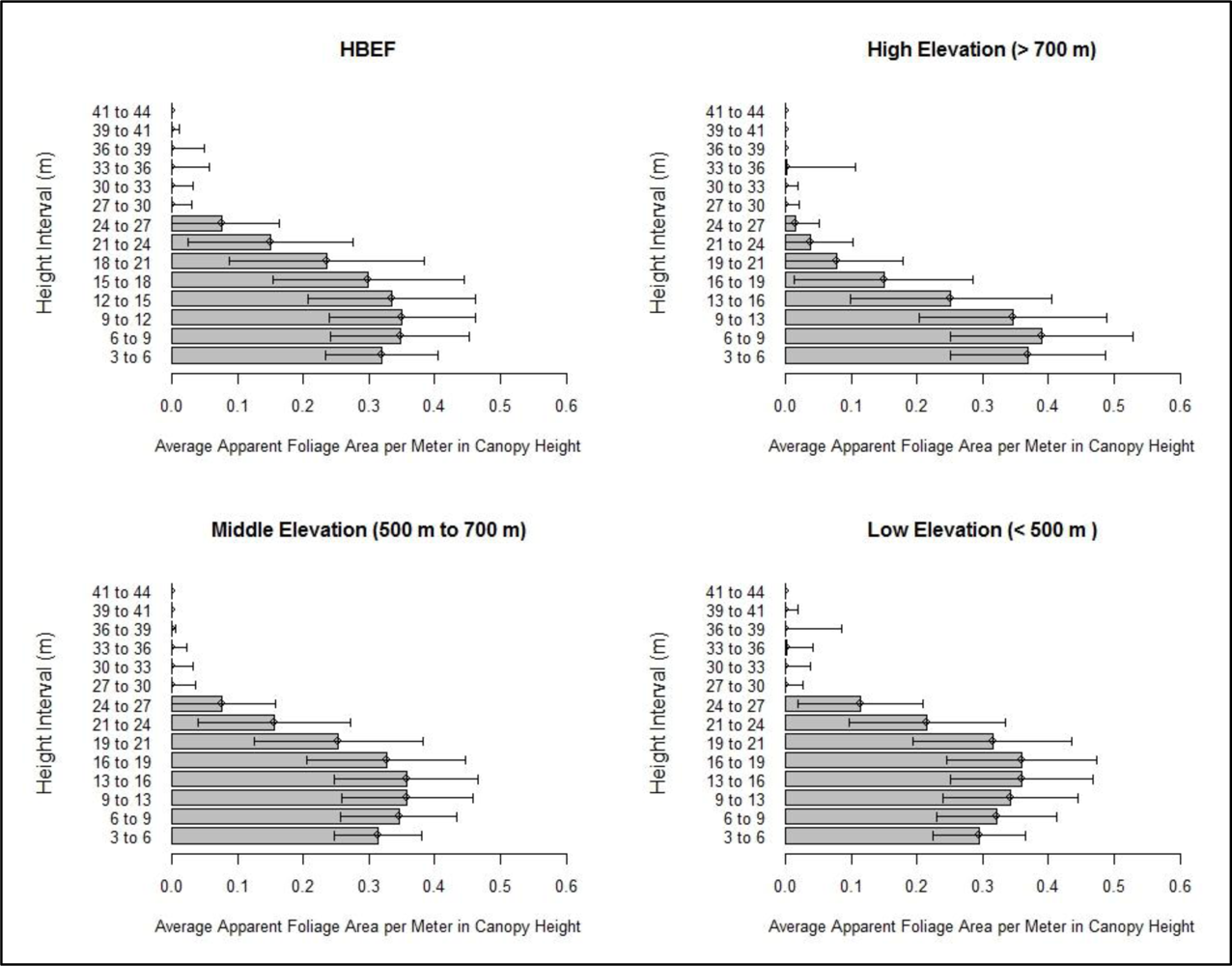
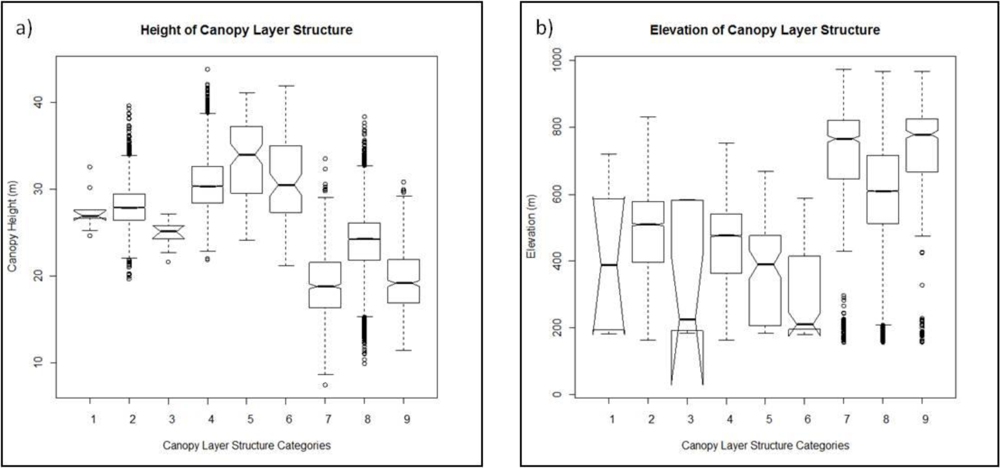
2.3.4. Height and Elevation Analyses
3. Results
3.1. Foliage Area Profile
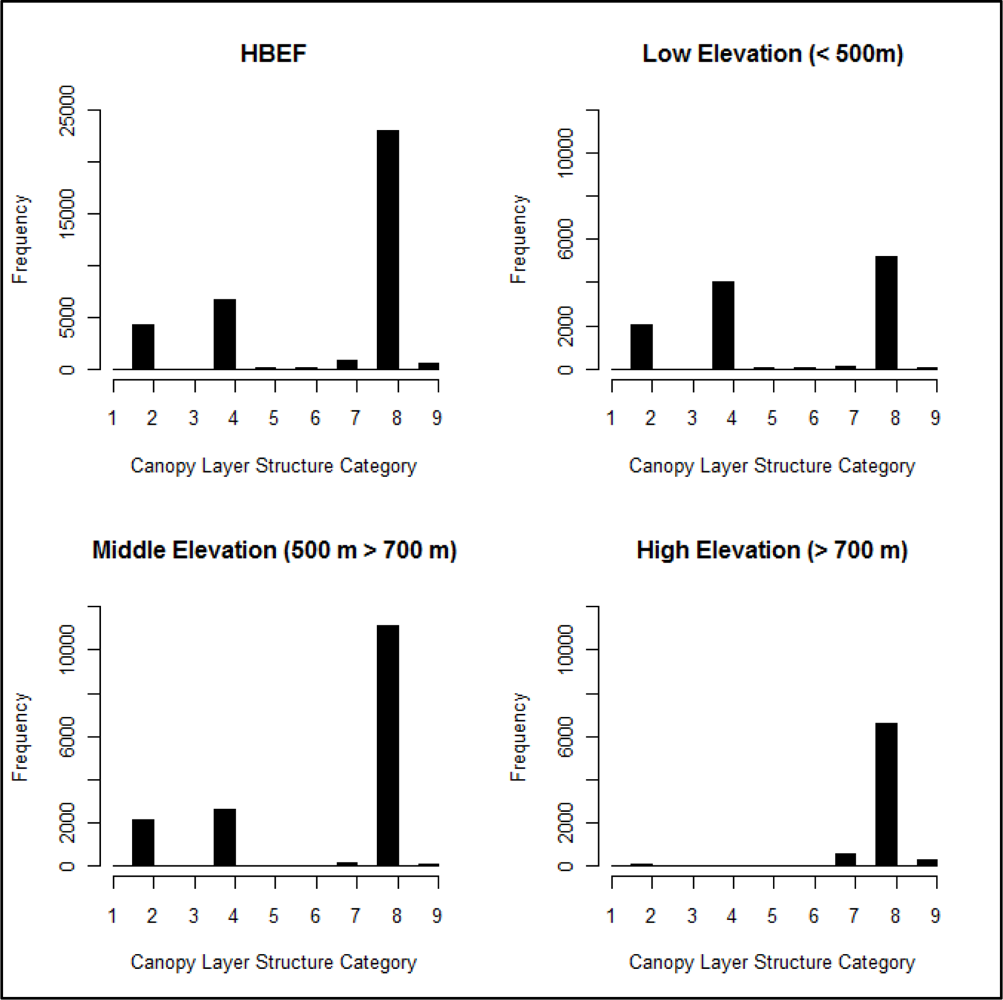
3.2. Canopy Layer Structure
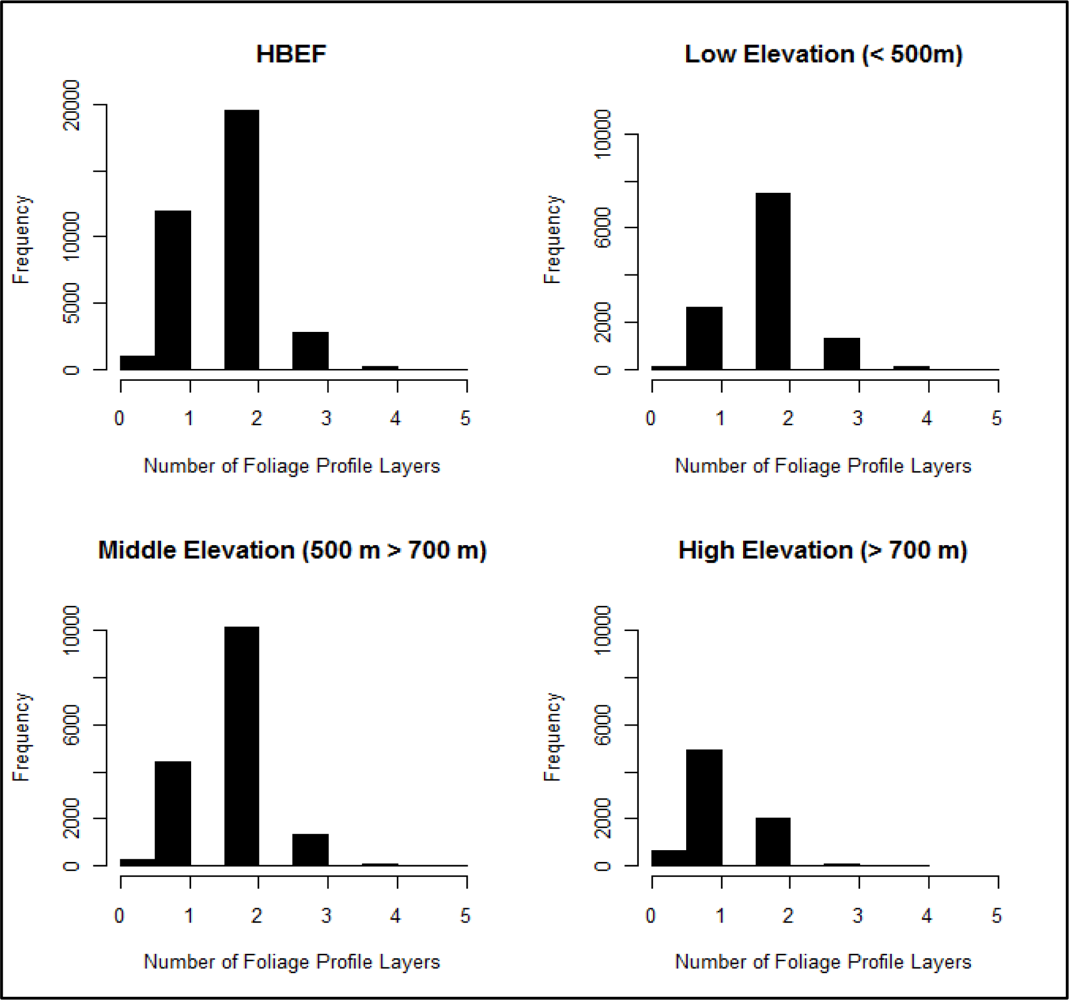
3.3. Foliage Profile Layering
4. Discussion
5. Conclusions
Acknowledgments
References
- MacArthur, R.H.; MacArthur, J.W. On bird species diversity. Ecology 1961, 42, 594–598. [Google Scholar]
- Spies, T.A. Forest structure: A key to the ecosystem. Northwest Sci 1998, 72, 34–39. [Google Scholar]
- Latham, P.A.; Zuuring, H.R.; Coble, D.W. A method for quantifying vertical forest structure. Forest Ecol. Manage 1998, 104, 157–170. [Google Scholar]
- Shaw, D.C. Vertical Organization of Canopy Biota. In Forest Canopies; Loman, M.D., Rinker, H.B., Eds.; Elsevier Academic Press: Amsterdam, The Netherlands, 2004; pp. 73–102. [Google Scholar]
- DeVries, P.J.; Murray, D.; Lande, R. Species diversity in vertical, horizontal, and temporal dimensions of a fruit-feeding butterfly community in an Ecuadorian rainforest. Biol. J. Linn. Soc 1997, 62, 343–364. [Google Scholar]
- Kalko, E.K.V; Handley, C.O.J. Neotropical bats in the canopy: Diversity, community structure, and implications for conservation. Plant Ecol 2001, 153, 319–333. [Google Scholar]
- Paijmans, K. An analysis of four tropical rain forest sites in New Guinea. J. Ecol 1970, 58, 77–101. [Google Scholar]
- Sherry, T.W. Competitive interactions and adaptive strategies of American redstarts and least flycatchers in a Northern Hardwoods forest. The Auk 1979, 96, 265–283. [Google Scholar]
- Popma, J.; Bongers, F.; Del Castillo, J.M. Patterns in the vertical structure of the tropical lowland rain forest of los tuxtlas, Mexico. Vegetatio 1988, 74, 81–91. [Google Scholar]
- Aber, J.D.; Pastor, J.; Melillo, J.M. The university of notre dame changes in forest canopy structure along a site quality gradient in Southern Wisconsin. Am. Midl. Nat 1982, 108, 256–265. [Google Scholar]
- Baker, P.J.; Wilson, J.S. A quantitative technique for the identification of canopy stratification in tropical and temperate forests. Forest Ecol. Manage 2000, 127, 77–86. [Google Scholar]
- Weltz, M.A.; Ritchie, C.; Fox, H.D. Comparison of laser and field measurements of vegetation height and canopy cover watershed precision vegetation properties height and canopy. Water Resour. Res 1994, 30, 1311–1319. [Google Scholar]
- Parker, G.G.; Brown, M.J. Forest canopy stratification—Is it useful? Am. Nat 2000, 155, 473–484. [Google Scholar]
- McElhinny, C.; Gibbons, P.; Brack, C. Forest and woodland stand structural complexity: Its definition and measurement. Forest Ecol. Manage 2005, 218, 1–24. [Google Scholar]
- Moffett, M.W. What’s “Up”? A critical loolc at the basic terms of canopy biology. Biotropica 2000, 32, 569–596. [Google Scholar]
- Smith, A.P. Stratification of temperature and tropical forests. Am. Nat 1973, 107, 671–683. [Google Scholar]
- Hitimana, J.; Kiyiapi, J.L.; Njunge, J.T. Forest structure characteristics in disturbed and undisturbed sites of Mt. Elgon Moist Lower Montane Forest, western Kenya. Forest Ecol. Manage 2004, 194, 269–291. [Google Scholar]
- Maltamo, M.; Packalen, P.; Yu, X.; Eerikainen, K.; Hyyppa, J.; Pitkanen, J. Identifying and quantifying structural characteristics of heterogeneous boreal forests using laser scanner data. Forest Ecol. Manage 2005, 216, 41–50. [Google Scholar]
- Dubayah, R.O.; Drake, J.B. Lidar remote sensing for forestry applications. J. Forest 2000, 98, 44–46. [Google Scholar]
- Lefsky, M.A.; Cohen, W.B.; Parker, G.G.; Harding, D.J. Lidar remote sensing for ecosystem studies. BioScience 2002, 52, 19–30. [Google Scholar]
- Vierling, K.T.; Vierling, L.A.; Gould, W.A.; Martinuzzi, S.; Clawges, R.M. Lidar: Shedding new light on habitat characterization and modeling. Front. Ecol. Environ 2008, 6, 90–98. [Google Scholar]
- Hyde, P.; Dubayah, R.; Peterson, B.; Blair, J.B.; Hofton, M. Mapping forest structure for wildlife habitat analysis using waveform lidar: Validation of montane ecosystems. Remote Sens. Environ 2005, 96, 427–437. [Google Scholar]
- Goetz, S.; Steinberg, D.; Dubayah, R.; Blair, B. Laser remote sensing of canopy habitat heterogeneity as a predictor of bird species richness in an eastern temperate forest, USA. Remote Sens. Environ 2007, 108, 254–263. [Google Scholar]
- Swatantran, A.; Dubayah, R.; Goetz, S.; Hofton, M.; Betts, M.G.; Sun, M.; Simard, M.; Holmes, R. Mapping migratory bird prevalence using remote sensing data fusion. PLoS One, 2012. [Google Scholar] [CrossRef]
- Hill, R.A.; Hinsley, S.A.; Gaveau, D.L.A.; Bellamy, P.E. Predicting habitat quality for Great Tits (Parus major) with airborne laser scanning data. Int. J. Remote Sens 2004, 25, 4851–4855. [Google Scholar]
- Goetz, S.J.; Steinberg, D.; Betts, M.G.; Holmes, R.T.; Doran, P.J.; Dubayah, R.; Hofton, M. Lidar remote sensing variables predict breeding habitat of a Neotropical migrant bird. Ecology 2010, 91, 1569–1576. [Google Scholar]
- Means, J.E.; Acker, S.A.; Harding, D.J.; Cohen, W.B.; Blair, J.B.; Harmon, M.E.; Mckee, W.A. Use of large-footprint scanning airborne lidar to estimate forest stand characteristics in the Western Cascades of Oregon. Remote Sens. Environ 1999, 3308, 298–308. [Google Scholar]
- Sexton, J.O.; Bax, T.; Siqueira, P.; Swenson, J.J.; Hensley, S. Forest ecology and management a comparison of lidar, radar, and field measurements of canopy height in pine and hardwood forests of southeastern North America. Forest Ecol. Manage 2009, 257, 1136–1147. [Google Scholar]
- Smart, L.S.; Swenson, J.J.; Christensen, N.L.; Sexton, J.O. Three-dimensional characterization of pine forest type and red-cockaded woodpecker habitat by small-footprint, discrete-return lidar. Forest Ecol. Manage 2012, 281, 100–110. [Google Scholar]
- Trainor, A.M.; Walters, J.R.; Morris, W.F.; Weiss, J.; Sexton, J.O. Environmental and conspecific cues influencing Red-Cockaded Woodpecker (Picoides borealis) prospecting movements during dispersal behavior. Landscape Ecol 2013, 28, 755–767. [Google Scholar]
- Coops, N.C.; Hilker, T.; Wulder, M.A.; St-Onge, B.; Newnham, G.; Siggins, A.; Trofymow, J.A. Estimating canopy structure of Douglas-fir forest stands from discrete-return LiDAR. Trees 2007, 21, 295–310. [Google Scholar]
- Jupp, D.L.B.; Culvenor, D.S.; Lovell, J.L.; Newnham, G.J.; Strahler, A.H.; Woodcock, C.E. Estimating forest LAI profiles and structural parameters using a ground-based laser called “Echidna”. Tree Physiol 2008, 29, 171–181. [Google Scholar]
- Zimble, D.A.; Evans, D.L.; Carlson, G.C.; Parker, R.C.; Grado, S.C.; Gerard, P.D. Characterizing vertical forest structure using small-footprint airborne LiDAR. Remote Sens. Environ 2003, 87, 171–182. [Google Scholar]
- Clawges, R.; Vierling, K.; Vierling, L.; Rowell, E. The use of airborne lidar to assess avian species diversity, density, and occurrence in a pine/aspen forest. Remote Sens. Environ 2008, 112, 2064–2073. [Google Scholar]
- Falkowski, M.J.; Evans, J.S.; Martinuzzi, S.; Gessler, P.E.; Hudak, A.T. Characterizing forest succession with lidar data: An evaluation for the Inland Northwest, USA. Remote Sens. Environ 2009, 113, 946–956. [Google Scholar]
- Müller, J.; Stadler, J.; Brandl, R. Composition versus physiognomy of vegetation as predictors of bird assemblages: The role of lidar. Remote Sens. Environ 2010, 114, 490–495. [Google Scholar]
- Schwarz, P.A.; Fahey, T.J.; Martin, C.W.; Siccama, T.G.; Bailey, A. Structure and composition of three northern hardwood-conifer forests with differing disturbance histories. Forest Ecol. Manage 2001, 144, 201–212. [Google Scholar]
- Blair, J.B.; Rabine, D.L.; Hofton, M.A. The laser vegetation imaging sensor: A medium-altitude, digitisation-only, airborne laser altimeter for mapping vegetation and topography. ISPRS J. Photogramm 1999, 54, 115–122. [Google Scholar]
- Hofton, M.A.; Rocchio, L.E.; Blair, J.B.; Dubayah, R. Validation of vegetation canopy lidar sub-canopy topography measurements for a dense tropical forest. J. Geodynamics 2002, 34, 491–502. [Google Scholar]
- Dubayah, R.O.; Sheldon, S.L.; Clark, D.B.; Hofton, M.A.; Blair, J.B.; Hurtt, G.C.; Chazdon, R.L. Estimation of tropical forest height and biomass dynamics using lidar remote sensing at La Selva, Costa Rica. J. Geophys. Res 2010, 115, 1–17. [Google Scholar]
- Drake, J.B.; Dubayah, R.O.; Clark, D.B.; Knox, R.G.; Blair, J.B.; Hofton, M.A.; Chazdon, R.L.; Weishampel, J.F.; Prince, S.D. Estimation of tropical forest structural characteristics using large-footprint lidar. Remote Sens. Environ 2002, 79, 305–319. [Google Scholar]
- Ni-Meister, W.; Jupp, D.L.B.; Dubayah, R. Modeling lidar waveforms in heterogeneous and discrete canopies. IEEE Trans. Geosci. Remote Sens 2001, 39, 1943–1958. [Google Scholar]
- Means, J.E.; Acker, S.A.; Harding, D.J.; Blair, J.B.; Lefsky, M.A.; Cohen, W.B.; Harmon, M.E.; Mckee, W.A. Use of large-footprint scanning airborne lidar to estimate forest stand characteristics in the Western Cascades of Oregon. Remote Sens. Environ 1999, 67, 298–308. [Google Scholar]
- Macarthur, R.H.; Horn, H.S. Foliage profile by vertical measurements. Ecology 1969, 50, 802–804. [Google Scholar]
- Lefsky, M.A.; Harding, D.; Cohen, W.B.; Parker, G.; Shugart, H.H. Surface lidar remote sensing of basal area and biomass in deciduous forests of eastern Maryland, USA. Remote Sens. Environ 1999, 67, 83–98. [Google Scholar]
- Lefsky, M.A.; Cohen, W.B.; Acker, S.A.; Parker, G.G. Lidar remote sensing of the canopy structure and biophysical properties of douglas-fir western hemlock forests. Remote Sens. Environ 1999, 70, 339–361. [Google Scholar]
- Harding, D.J.; Lefsky, M.A.; Parker, G.G.; Blair, J.B. Laser altimeter canopy height profiles: Methods and validation for closed-canopy, broadleaf forests. Remote Sens. Environ 2001, 76, 283–297. [Google Scholar]
- Helms, J.A. (Ed.) The Dictionary of Forestry; Society of American Foresters: Bethesda, MD, USA, 1998.
- Carey, A.B.; Kershner, J.; Biswell, B.; de Toledo, L.D. Ecological scale and forest development: Squirrels, dietary fungi, and vascular plants in managed and unmanaged forests. Wildlife Monogr 1999, 142, 3–71. [Google Scholar]
- Globe Graminoid, Tree and Shrub Height Field Guide. Avaiable online: http://www.globe.gov/documents/355050/355097/lc_fg_treeheight.pdf (accessed on 21 September 2010).
- Missouri Department of Conservation Forestry Terms and Definitions. Available online: mdc4.mdc.mo.gov/Documents/13183.doc (accessed on 22 January 2013).
- Koike, A.F.; Tabata, H.; Malla, S.B.; Koike, F.; Arthur, M. Canopy structures and its effect on shoot growth and flowering in subalpine forests. Vegetatio 1990, 86, 101–113. [Google Scholar]
- Austin, M.P. Searching for a model for use in vegetation analysis. Vegetatio 1980, 42, 11–21. [Google Scholar]
- Whittaker, R.H.; Bormann, F.H.; Likens, G.E.; Siccama, T.G. The hubbard brook ecosystem study: Forest biomass and production. Ecol. Monogr 1974, 44, 233–254. [Google Scholar]
- Kaufmann, M.R.; Ryan, M.G. Physiographic, stand, and environmental effects on individual tree growth and growth efficiency in subalpine forests. Tree Physiol 1986, 2, 47–59. [Google Scholar]
- Ryan, M.G.; Yoder, B.J. Limits to tree height hydraulic and tree growth. BioScience 1997, 47, 235–242. [Google Scholar]
- Tardif, J.; Camarero, J.J.; Ribas, M.; Gutierrez, E. Spatiotemporal variability in tree growth in the central pyrenees: Climatic and site influences. Ecol. Monogr 2003, 73, 241–257. [Google Scholar]
- Huang, C.; Goward, S.N.; Schleeweis, K.; Thomas, N.; Masek, J.G.; Zhu, Z. Remote sensing of environment dynamics of national forests assessed using the Landsat record: Case studies in eastern United States. Remote Sens. Environ 2009, 113, 1430–1442. [Google Scholar]
- Ford, E.D. Competiton and stand structure in some even-aged plant monocultures. J. Ecol 1975, 63, 311–333. [Google Scholar]
- Perry, D.A. The Competition Process in Forest Stands. In Attributes of Trees as Crop Plants; Cannell, M.G., Jackson, J.E., Eds.; Institute of Terrestrial Ecology: Abbots Ripton, Hunts, UK, 1985; pp. 481–506. [Google Scholar]
- Maguire, D.A.; Brissette, J.C.; Gu, L. Crown structure and growth efficiency of red spruce in uneven-aged, mixed-species stands in Maine. Can. J. For. Res 1998, 28, 1233–1240. [Google Scholar]
- Hurtt, G.C.; Dubayah, R.; Drake, J.; Moorcroft, P.R.; Pacala, S.W.; Blair, J.B.; Fearon, M.G. Beyond potential vegetation: Combining lidar data and a height-structured model for carbon studies. Ecol. Appl 2004, 14, 873–883. [Google Scholar]
- Thomas, R.; Hurtt, G.C.; Dubayah, R.; Schilz, M.H. Using lidar data and a height-structured ecosystem model to estimate forest carbon stocks adn fluxes over mountainous terrain. Can. J. Remote Sens 2008, 34, 351–363. [Google Scholar]
















Share and Cite
Whitehurst, A.S.; Swatantran, A.; Blair, J.B.; Hofton, M.A.; Dubayah, R. Characterization of Canopy Layering in Forested Ecosystems Using Full Waveform Lidar. Remote Sens. 2013, 5, 2014-2036. https://0-doi-org.brum.beds.ac.uk/10.3390/rs5042014
Whitehurst AS, Swatantran A, Blair JB, Hofton MA, Dubayah R. Characterization of Canopy Layering in Forested Ecosystems Using Full Waveform Lidar. Remote Sensing. 2013; 5(4):2014-2036. https://0-doi-org.brum.beds.ac.uk/10.3390/rs5042014
Chicago/Turabian StyleWhitehurst, Amanda S., Anu Swatantran, J. Bryan Blair, Michelle A. Hofton, and Ralph Dubayah. 2013. "Characterization of Canopy Layering in Forested Ecosystems Using Full Waveform Lidar" Remote Sensing 5, no. 4: 2014-2036. https://0-doi-org.brum.beds.ac.uk/10.3390/rs5042014