Human Land-Use Practices Lead to Global Long-Term Increases in Photosynthetic Capacity

Abstract
:1. Introduction
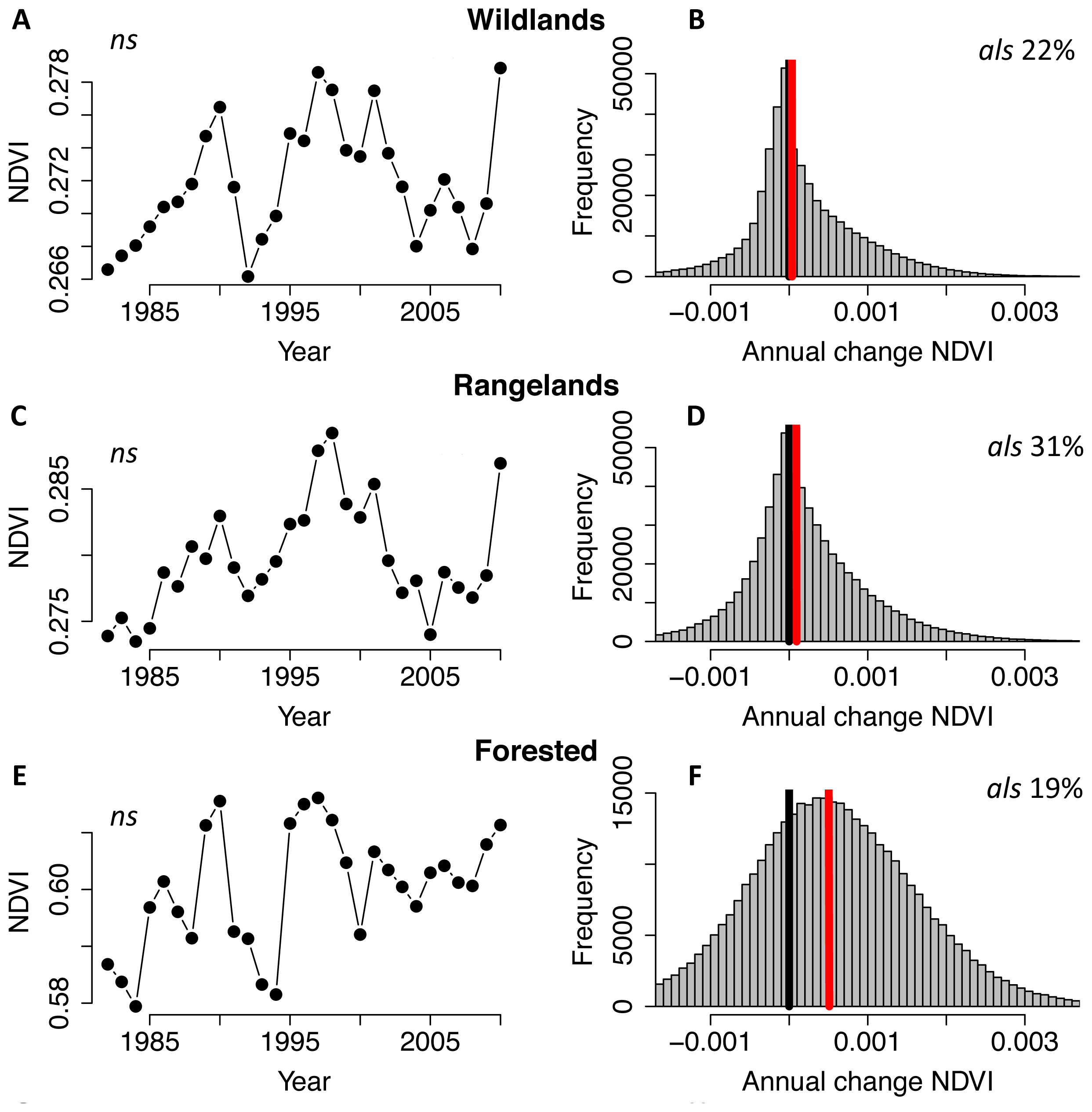
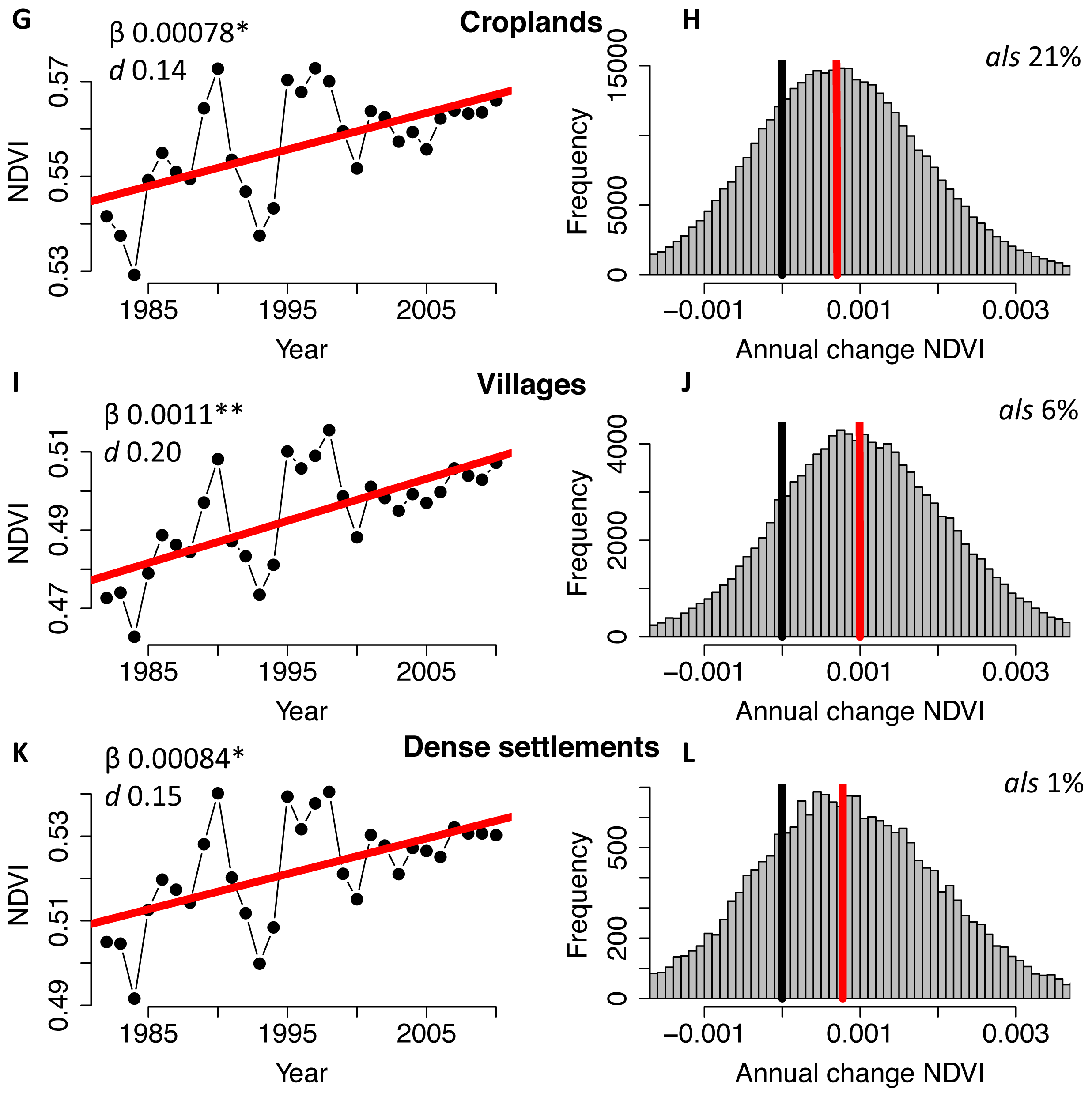
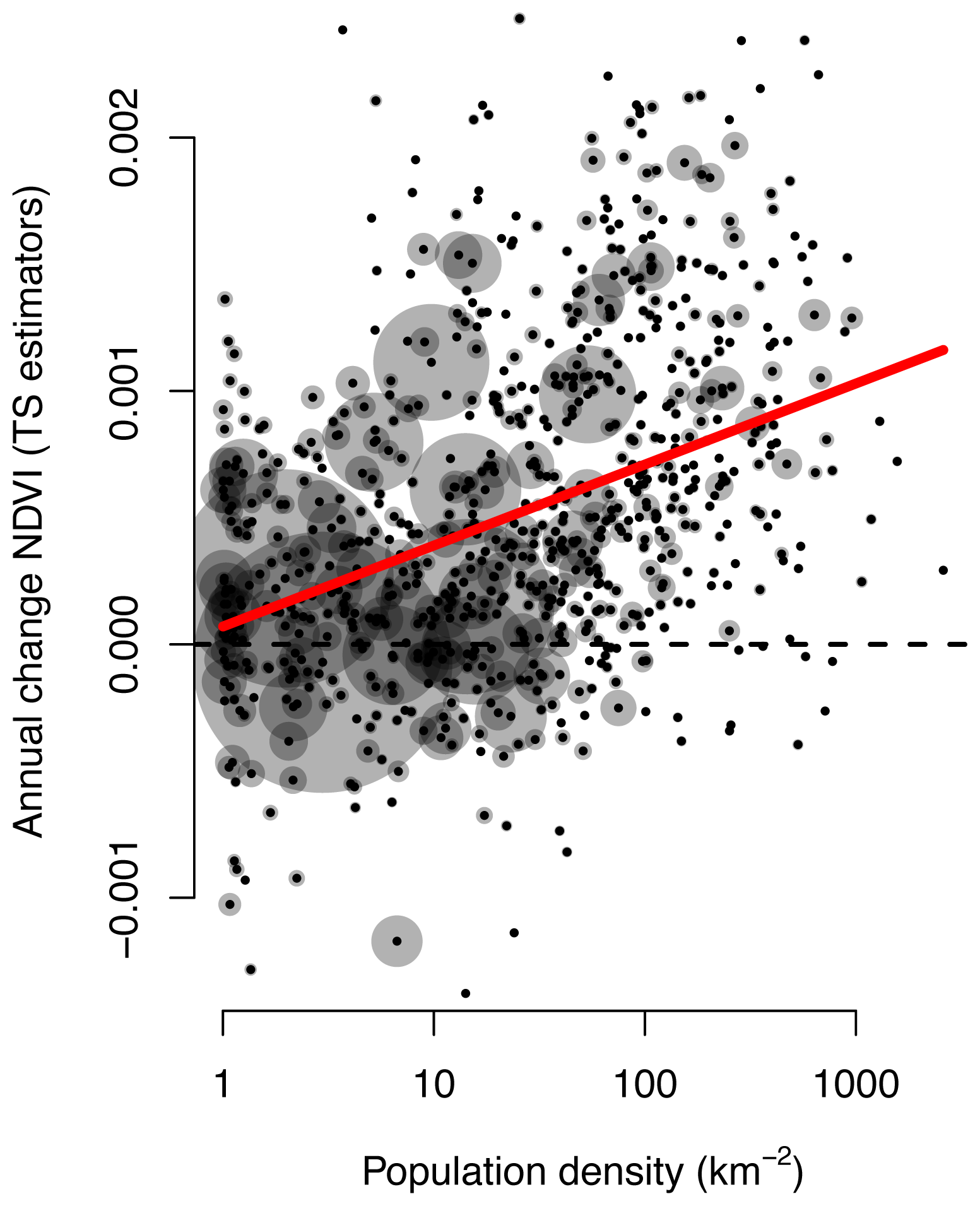
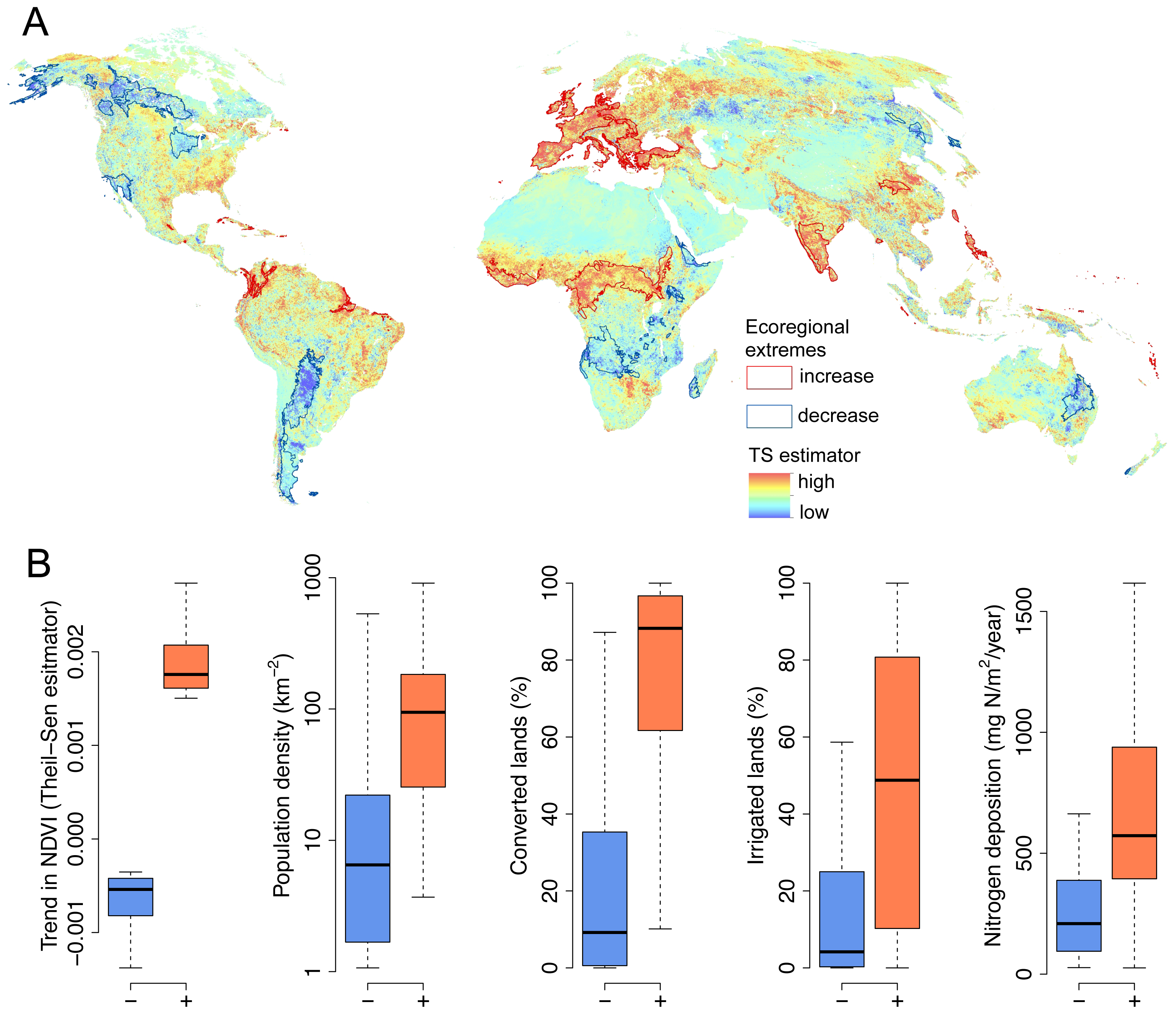
2. Results and Discussion
3. Experimental Section
3.1. NDVI Data and Pre-Processing
3.2. Trend Analysis & Anthropogenic Effects
4. Conclusions
Acknowledgments
Conflicts of Interest
- Author ContributionsThomas Mueller, Gunnar Dressler, Compton J. Tucker and William F. Fagan conceptualized the study, Thomas Mueller and Gunnar Dressler analyzed the data, and all authors contributed to the writing of the manuscript.
References
- Anyamba, A.C.; Tucker, J. Analysis of Sahelian vegetation dynamics using NOAA-AVHRR NDVI data from 1981–2003. J. Arid Environ 2007, 63, 596–614. [Google Scholar]
- Jeyaseelan, T.; Roy, P.S.; Young, S.S. Persistent changes in NDVI between 1982 and 2003 over India using AVHRR GIMMS (Global Inventory Modeling and Mapping Studies) data. Int. J. Remote Sens 2007, 28, 4927–4946. [Google Scholar]
- Olsson, L.; Eklundh, L.; Ardö, J. A recent greening of the Sahel—Trends, patterns and potential causes. J. Arid Environ 2005, 63, 556–566. [Google Scholar]
- Myneni, R.B.; Nemani, R.R.; Running, S.W. Estimation of global leaf area index and absorbed par using radiative transfer models. IEEE Trans. Geosci. Remote Sens 1997, 35, 1380–1393. [Google Scholar]
- Nemani, R.R.; Keeling, C.D.; Hashimoto, H.; Jolly, W.M.; Piper, S.C.; Tucker, C.J.; Myneni, R.B.; Running, S.W. Climate-driven increases in global terrestrial net primary production from 1982 to 1999. Science 2003, 300, 1560–1563. [Google Scholar]
- Tucker, C.J.; Sellers, P.J. Satellite remote-sensing of primary production. Int. J. Remote Sens 1986, 7, 1395–1416. [Google Scholar]
- Hickler, T.; Eklundh, L.; Seaquist, J.W.; Smith, B.; Ardo, J.; Olsson, L.; Sykes, M.T.; Sjostrom, M. Precipitation controls Sahel greening trend. Geophys. Res. Lett 2005, 32. [Google Scholar] [CrossRef]
- Cao, M.K.; Prince, S.D.; Small, J.; Goetz, S.J. Remotely sensed interannual variations and trends in terrestrial net primary productivity 1981–2000. Ecosystems 2004, 7, 233–242. [Google Scholar]
- Sanderson, E.W.; Jaiteh, M.; Levy, M.A.; Redford, K.H.; Wannebo, A.V.; Woolmer, G. The human footprint and the last of the wild. Bioscience 2002, 52, 891–904. [Google Scholar]
- Mittermeier, R.A.; Mittermeier, C.G.; Brooks, T.M.; Pilgrim, J.D.; Konstant, W.R.; da Fonseca, G.A.B.; Kormos, C. Wilderness and biodiversity conservation. Proc. Natl. Acad. Sci. USA 2003, 100, 10309–10313. [Google Scholar]
- Foley, J.A.; DeFries, R.; Asner, G.P.; Barford, C.; Bonan, G.; Carpenter, S.R.; Chapin, F.S.; Coe, M.T.; Daily, G.C.; Gibbs, H.K.; et al. Global consequences of land use. Science 2005, 309, 570–574. [Google Scholar]
- Hurtt, G.C.; Chini, L.P.; Frolking, S.; Betts, R.A.; Feddema, J.; Fischer, G.; Fisk, J.P.; Hibbard, K.; Houghton, R.A.; Janetos, A.; et al. Harmonization of land-use scenarios for the period 1500–2100: 600 years of global gridded annual land-use transitions, wood harvest, and resulting secondary lands. Clim. Chang 2011, 109, 117–161. [Google Scholar]
- Ellis, E.C.; Ramankutty, N. Putting people in the map: Anthropogenic biomes of the world. Front. Ecol. Environ 2008, 6, 439–447. [Google Scholar]
- Baldi, G.; Nosetto, M.D.; Aragon, R.; Aversa, F.; Paruelo, J.M.; Jobbagy, E.G. Long-term satellite NDVI data sets: Evaluating their ability to detect ecosystem functional changes in South America. Sensors 2008, 8, 5397–5425. [Google Scholar]
- Bradley, B.A.; Mustard, J.F. Comparison of phenology trends by land cover class: A case study in the Great Basin, USA. Glob. Chang. Biol 2008, 14, 334–346. [Google Scholar]
- Neigh, C.S.R.; Tucker, C.J.; Townshend, J.R.G. North American vegetation dynamics observed with multi-resolution satellite data. Remote Sens. Environ 2008, 112, 1749–1772. [Google Scholar]
- Eastman, J.R.; Sangermano, F.; Machado, E.A.; Rogan, J.; Anyamba, A. Global trends in seasonality of Normalized Difference Vegetation Index (NDVI), 1982–2011. Remote Sens 2013, 5, 4799–4818. [Google Scholar]
- Niyogi, D.; Kishtawal, C.; Tripathi, S.; Govindaraju, R.S. Observational evidence that agricultural intensification and land use change may be reducing the Indian summer monsoon rainfall. Water Resour. Res 2010. [Google Scholar] [CrossRef]
- Olson, D.M.; Dinerstein, E.; Wikramanayake, E.D.; Burgess, N.D.; Powell, G.V.N.; Underwood, E.C.; D’Amico, J.A.; Itoua, I.; Strand, H.E.; Morrison, J.C.; et al. Terrestrial ecoregions of the worlds: A new map of life on earth. Bioscience 2001, 51, 933–938. [Google Scholar]
- Fensholt, R.; Proud, S.R. Evaluation of earth observation based global long term vegetation trends—Comparing GIMMS and MODIS global NDVI time series. Remote Sens. Environ 2012, 119, 131–147. [Google Scholar]
- De Jong, R.; de Bruin, S.; de Wit, A.; Schaepman, M.E.; Dent, D.L. Analysis of monotonic greening and browning trends from global NDVI time-series. Remote Sens. Environ 2011, 115, 692–702. [Google Scholar] [Green Version]
- Donohue, R.J.; McVicar, T.R.; Roderick, M.L. Climate-related trends in Australian vegetation cover as inferred from satellite observations, 1981–2006. Glob. Chang. Biol 2009, 15, 1025–1039. [Google Scholar]
- Verbyla, D. Browning boreal forests of western North America. Environ. Res. Lett 2011, 6. [Google Scholar] [CrossRef]
- Beck, P.S.A.; Juday, G.P.; Alix, C.; Barber, V.A.; Winslow, S.E.; Sousa, E.E.; Heiser, P.; Herriges, J.D.; Goetz, S.J. Changes in forest productivity across Alaska consistent with biome shift. Ecol. Lett 2011, 14, 373–379. [Google Scholar]
- Julien, Y.; Sobrino, J.A.; Verhoef, W. Changes in land surface temperatures and NDVI values over Europe between 1982 and 1999. Remote Sens. Environ 2006, 103, 43–55. [Google Scholar]
- Stockli, R.; Vidale, P.L. European plant phenology and climate as seen in a 20-year AVHRR land-surface parameter dataset. Int. J. Remote Sens 2004, 25, 3303–3330. [Google Scholar]
- Piao, S.L.; Fang, J.Y.; Zhou, L.M.; Guo, Q.H.; Henderson, M.; Ji, W.; Li, Y.; Tao, S. Interannual variations of monthly and seasonal Normalized Difference Vegetation index (NDVI) in China from 1982 to 1999. J. Geophys. Res.: Atmos 2003. [Google Scholar] [CrossRef]
- Heumann, B.W.; Seaquist, J.W.; Eklundh, L.; Jonsson, P. AVHRR derived phenological change in the Sahel and Soudan, Africa, 1982–2005. Remote Sens. Environ 2007, 108, 385–392. [Google Scholar]
- Begue, A.; Vintrou, E.; Ruelland, D.; Claden, M.; Dessay, N. Can a 25-year trend in Soudano-Sahelian vegetation dynamics be interpreted in terms of land use change? A remote sensing approach. Glob. Environ. Chang 2011, 21, 413–420. [Google Scholar]
- De Vries, W.; Posch, M. Modelling the impact of nitrogen deposition, climate change and nutrient limitations on tree carbon sequestration in Europe for the period 1900–2050. Environ. Pollut 2011, 159, 2289–2299. [Google Scholar]
- The State of the World’s Forests; Food and Agriculture Organization of the United Nations: Rome, Italy, 2011.
- Clark, M.L.; Aide, T.M.; Grau, H.R.; Riner, G. A scalable approach to mapping annual land cover at 250 m using MODIS time series data: A case study in the Dry Chaco ecoregion of South America. Remote Sens. Environ 2010, 114, 2816–2832. [Google Scholar]
- Pinzon, J.E.; Tucker, C.J. A non-stationary 1981–2012 AVHRR NDVI3g time series. Remote Sens 2014. submitted. [Google Scholar]
- Tucker, C.J.; Pinzon, J.E.; Brown, M.E.; Slayback, D.A.; Pak, E.W.; Mahoney, R.; Vermote, E.F.; El Saleous, N. An extended AVHRR 8-km NDVI dataset compatible with MODIS and SPOT vegetation NDVI data. Int. J. Remote Sens 2005, 26, 4485–4498. [Google Scholar]
- Zhou, L.M.; Tucker, C.J.; Kaufmann, R.K.; Slayback, D.; Shabanov, N.V.; Myneni, R.B. Variations in northern vegetation activity inferred from satellite data of vegetation index during 1981 to 1999. J. Geophys. Res.: Atmos 2001, 106, 20069–20083. [Google Scholar]
- Sobrino, J.A.; Julien, Y. Global trends in NDVI-derived parameters obtained from GIMMS data. Int. J. Remote Sens 2011, 32, 4267–4279. [Google Scholar]
- Beck, H.E.; McVicar, T.R.; van Dijk, A.; Schellekens, J.; de Jeu, R.A.M.; Bruijnzeel, L.A. Global evaluation of four AVHRR-NDVI data sets: Intercomparison and assessment against Landsat imagery. Remote Sens. Environ 2011, 115, 2547–2563. [Google Scholar]
- Justice, C.O.; Townshend, J.R.G.; Vermote, E.F.; Masuoka, E.; Wolfe, R.E.; Saleous, N.; Roy, D.P.; Morisette, J.T. An overview of MODIS Land data processing and product status. Remote Sens. Environ 2002, 83, 3–15. [Google Scholar]
- Natural Earth Data Sets. Available online: http://www.naturalearthdata.com/ (accessed on 30 May 2014).
- Slayback, D.A.; Pinzon, J.E.; Los, S.O.; Tucker, C.J. Northern hemisphere photosynthetic trends 1982–99. Glob. Chang. Biol 2003, 9, 1–15. [Google Scholar]
- De Beurs, K.M.; Henebry, G.M. A statistical framework for the analysis of long image time series. Int. J. Remote Sens 2005, 26, 1551–1573. [Google Scholar]
- Alcaraz-Segura, D.; Chuvieco, E.; Epstein, H.E.; Kasischke, E.S.; Trishchenko, A. Debating the greening vs. Browning of the North American boreal forest: Differences between satellite datasets. Glob. Chang. Biol 2010, 16, 760–770. [Google Scholar]
- Nagelkerke, N.J.D. A note on a general definition of the coefficient of determination. Biometrika 1991, 78, 691–692. [Google Scholar]
- Socioeconomic Data and Applications Center, Center for International Earth Science Information Network (CIESIN), Columbia University and Centro Internacional de Agricultura Tropical. Gridded Population of the World, Version 3, SEDAC. 2005. Available online: http://sedac.ciesin.columbia.edu/gpw (accessed on 30 May 2014).
- Global Maps of Atmospheric Nitrogen Deposition, 1860, 1993 and 2050. Available online: http://daac.ornl.gov//CLIMATE/guides/global_N_deposition_maps.html (accessed on 30 May 2014).
- Global Map of Irrigation Areas. Available online: http://www.fao.org/nr/water/aquastat/irrigationmap/index10.stm (accessed on 30 May 2014).
- Dormann, C.F.; Elith, J.; Bacher, S.; Buchmann, C.; Carl, G.; Carré, G.; Marquéz, J.R.G.; Gruber, B.; Lafourcade, B.; Leitão, P.J.; et al. Collinearity: A review of methods to deal with it and a simulation study evaluating their performance. Ecography 2012, 36, 27–46. [Google Scholar]
- Saleska, S.R.; Didan, K.; Huete, A.R.; da Rocha, H.R. Amazon forests green-up during 2005 drought. Science 2007. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| β | p-Value | |
|---|---|---|
| Intercept | 0.00017 | <0.0001 |
| Converted lands (% area) | 0.00040 | <0.0001 |
| Nitrogen (mg N/m2/year) | 0.00000025 | 0.0001 |
| Irrigation (% area) | 0.00025 | 0.013 |
| A. Models Relating Trends in NDVI to Population Density | ||
|---|---|---|
| Excluding the Moist Tropical Broadleaf Biome | ||
| β | p-Value | |
| Intercept | −0.000054 | 0.25 |
| Log10 Population (m−2) | 0.00036 | <0.0001 |
| Using Standardized TS Estimators | ||
| Coefficient of determination: 0.10, Δ AIC: 80, overall significance: <0.0001 | ||
| β | p-Value | |
| Intercept | 0.000043 | 0.68 |
| Log10 Population (m−2) | 0.00069 | <0.0001 |
| Using Fishnet as Spatial Unit | ||
| Coefficient of determination: 0.059, Δ AIC: 49, overall significance: <0.0001. | ||
| β | p-Value | |
| Intercept | 0.00024 | <0.0001 |
| Log10 Population (m−2) | 0.00023 | <0.0001 |
| B. Models Relating Trends in NDVI to Nitrogen Deposition, Irrigation, and Converted Lands | ||
| Excluding the Moist Tropical Broadleaf Biome | ||
| Coefficient of determination: 0.26, Δ AIC: 164, overall significance: <0.0001. | ||
| β | p-Value | |
| Intercept | 0.000068 | 0.044 |
| Converted lands (% area) | 0.00046 | <0.0001 |
| Nitrogen (mg N/m2/year) | 0.00000023 | 0.0022 |
| Irrigation (% area) | 0.00035 | 0.0028 |
| Using Standardized TS Estimators | ||
| Coefficient of determination: 0.15, Δ AIC: 122, overall significance: <0.0001. | ||
| β | p-Value | |
| Intercept | 0.00029 | 0.0002 |
| Converted lands (% area) | 0.00053 | 0.024 |
| Nitrogen (mg N/m2/year) | 0.00000043 | 0.0075 |
| Irrigation (% area) | 0.0010 | 0.0001 |
| Using Fishnet as Spatial Unit | ||
| Coefficient of determination: 0.11, Δ AIC: 88, overall significance: <0.0001 | ||
| β | p-Value | |
| Intercept | 0.00022 | <0.0001 |
| Converted lands (% area) | 0.00037 | 0.0003 |
| Nitrogen (mg N/m2/year) | 0.00000015 | 0.020 |
| Irrigation (% area) | 0.00031 | 0.0064 |
© 2014 by the authors; licensee MDPI, Basel, Switzerland This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Mueller, T.; Dressler, G.; Tucker, C.J.; Pinzon, J.E.; Leimgruber, P.; Dubayah, R.O.; Hurtt, G.C.; Böhning-Gaese, K.; Fagan, W.F. Human Land-Use Practices Lead to Global Long-Term Increases in Photosynthetic Capacity. Remote Sens. 2014, 6, 5717-5731. https://0-doi-org.brum.beds.ac.uk/10.3390/rs6065717
Mueller T, Dressler G, Tucker CJ, Pinzon JE, Leimgruber P, Dubayah RO, Hurtt GC, Böhning-Gaese K, Fagan WF. Human Land-Use Practices Lead to Global Long-Term Increases in Photosynthetic Capacity. Remote Sensing. 2014; 6(6):5717-5731. https://0-doi-org.brum.beds.ac.uk/10.3390/rs6065717
Chicago/Turabian StyleMueller, Thomas, Gunnar Dressler, Compton J. Tucker, Jorge E. Pinzon, Peter Leimgruber, Ralph O. Dubayah, George C. Hurtt, Katrin Böhning-Gaese, and William F. Fagan. 2014. "Human Land-Use Practices Lead to Global Long-Term Increases in Photosynthetic Capacity" Remote Sensing 6, no. 6: 5717-5731. https://0-doi-org.brum.beds.ac.uk/10.3390/rs6065717