Interpretability of Spatiotemporal Dynamics of the Brain Processes Followed by Mindfulness Intervention in a Brain-Inspired Spiking Neural Network Architecture

 ,
,  , ,
, ,  ,
,
Abstract
:1. Introduction
1.1. Mindfulness Training: The Neural Basis for Performance Monitoring
1.2. Computational Modelling of Data in a Brain-Inspired Spiking Neural Network Architecture
- (i)
- design a SNN architecture to model and understand brain activity patterns generated before and after mindfulness training for spatiotemporal relationships between continuous ERP data streams, while participants of two groups (experimental and waitlist) were responding to (1): target stimuli (2) and nontarget (distractor) stimuli.
- (ii)
- create computational models from ERP data to better scrutinize the temporal dynamic processes at different time points of stimulus presentation.
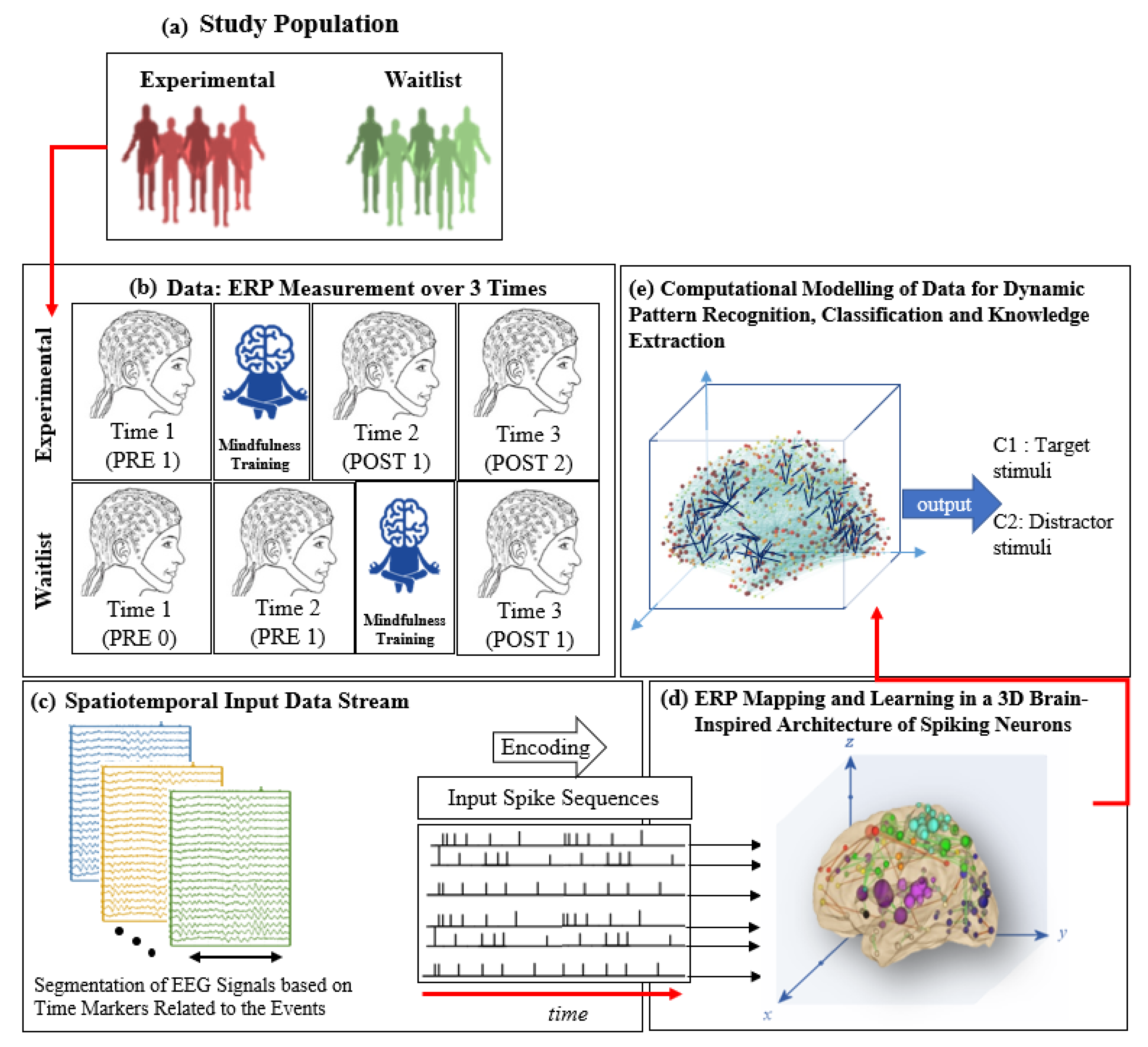
2. Materials and Methods
2.1. Ethics
2.2. Data and Software Availability
2.3. Participants
2.4. Trait Mindfulness
2.5. Mindfulness Intervention
2.6. ERP Cognitive Task
2.7. EEG Acquisition and Pre-Processing
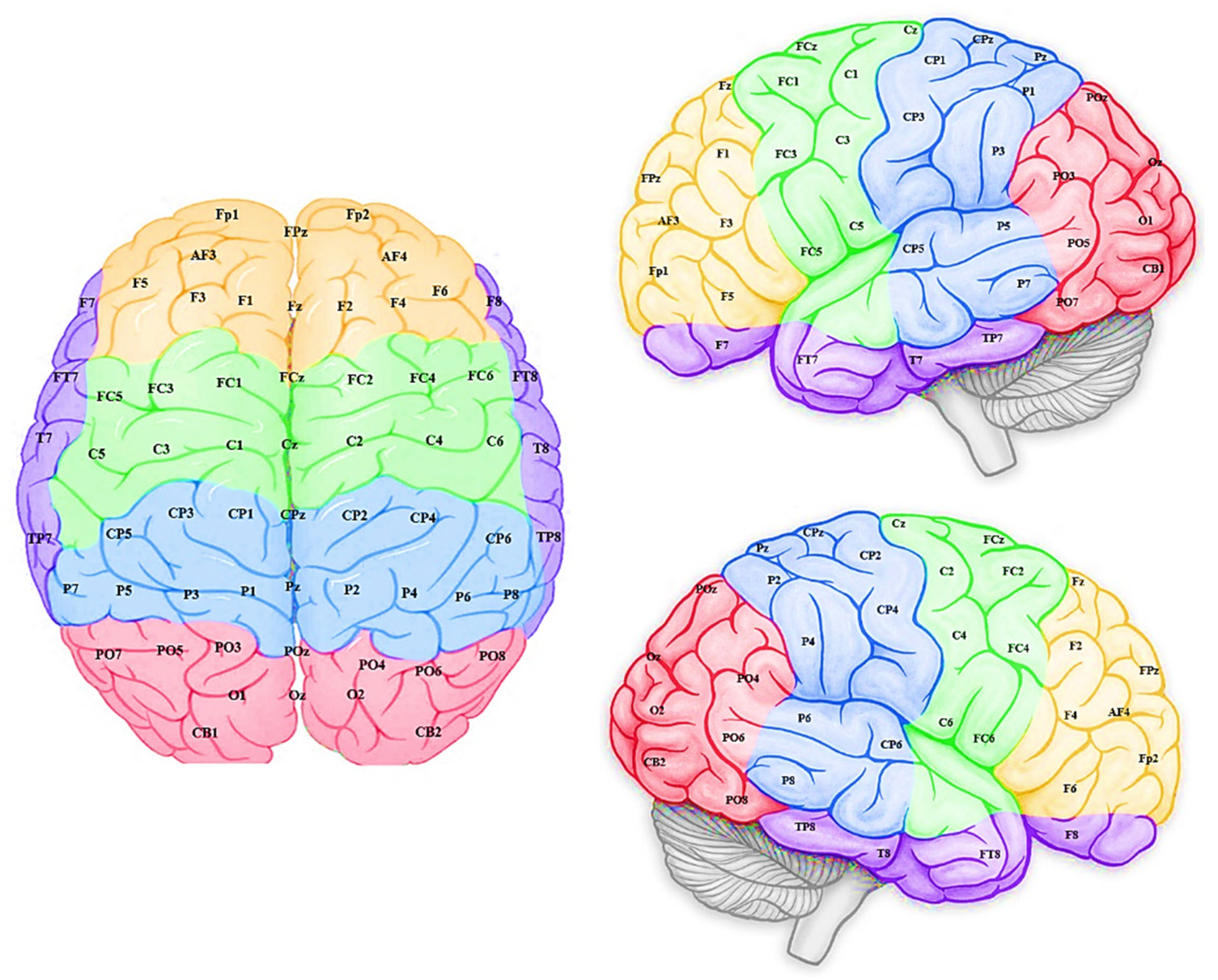
2.8. Method: Brain-Inspired Spiking Neural Networks
- -
- A brain template is utilised to build a 3-dimensional space of spiking neurons (SNN model) that reserve spatial information of brain structure. Mapping of brain data variables into the SNN model.
- -
- Input spatiotemporal data (brain signals) are converted to a ‘spike train’ (series of binary events when the brain signal reaches a threshold value).
- -
- The SNN model is initialised using the brain-inspired small-world connectivity rule.
- -
- The initialised SNN model learns through a biologically plausible learning algorithm to adapt the model’s connectivity, resulting in long chains of connections.
2.8.1. Initialisation and Encoding of the ERP Data in the SNN Model
2.8.2. Unsupervised Learning Mechanism in a SNN Model
2.8.3. Supervised Learning and Pattern Classification in SNN Models
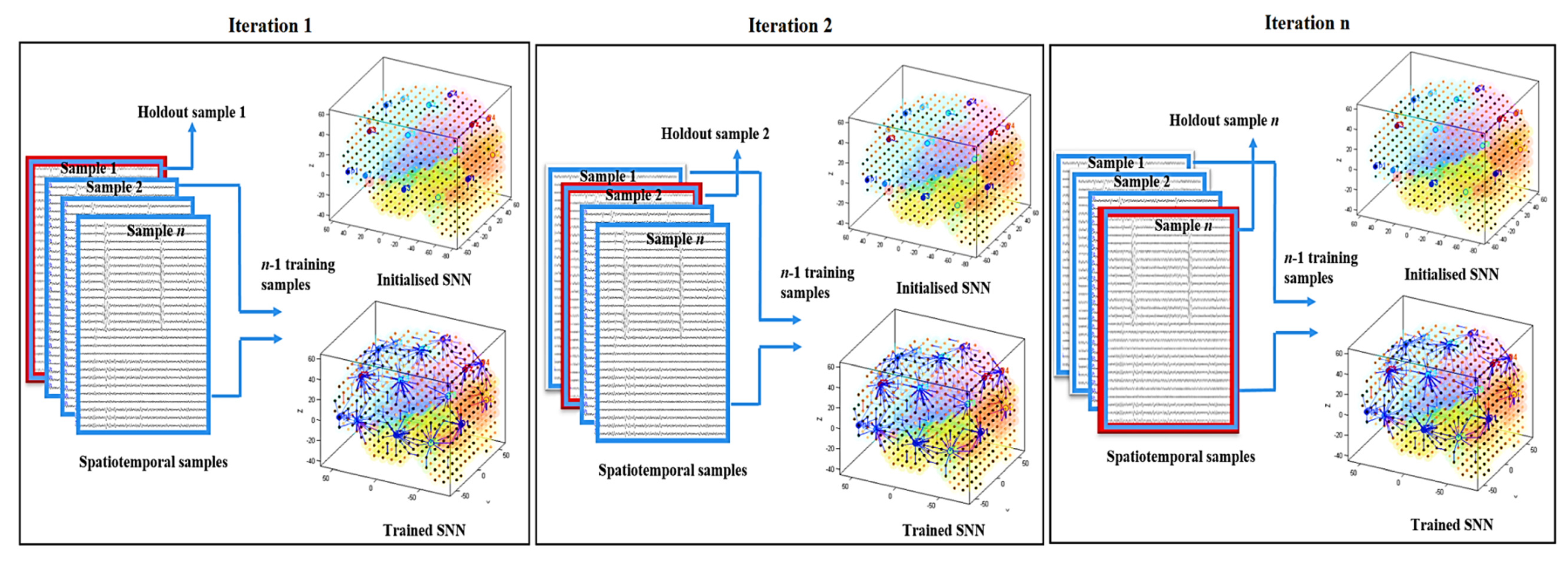
2.8.4. SNN Model Evaluation
- The SNN model is initialised.
- The initialised SNN model is trained with (n − 1) samples (one sample is excluded from the training).
- The average of the quantitative information (spatiotemporal connectivity) in the trained SNN model is calculated.
3. Results
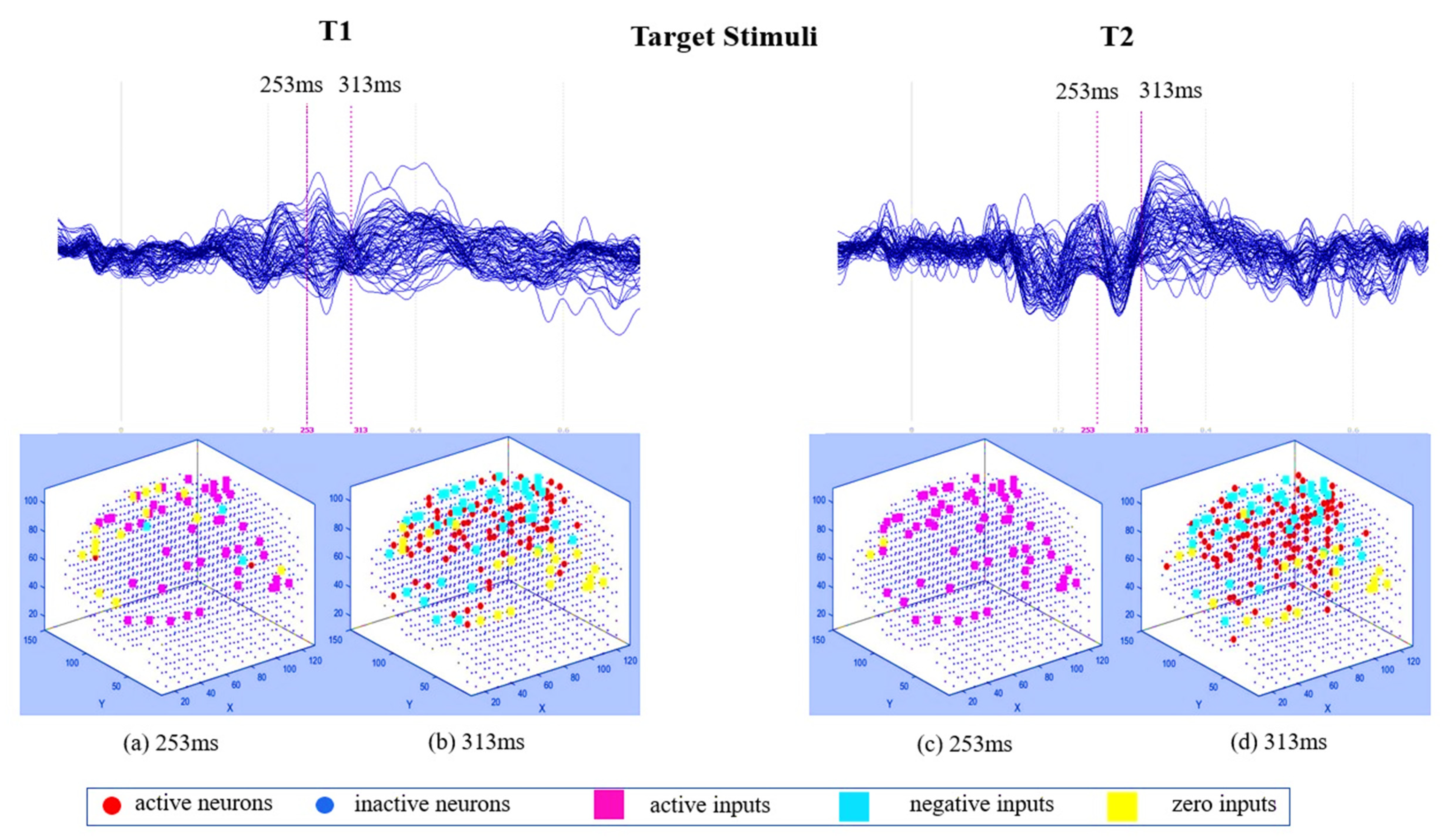
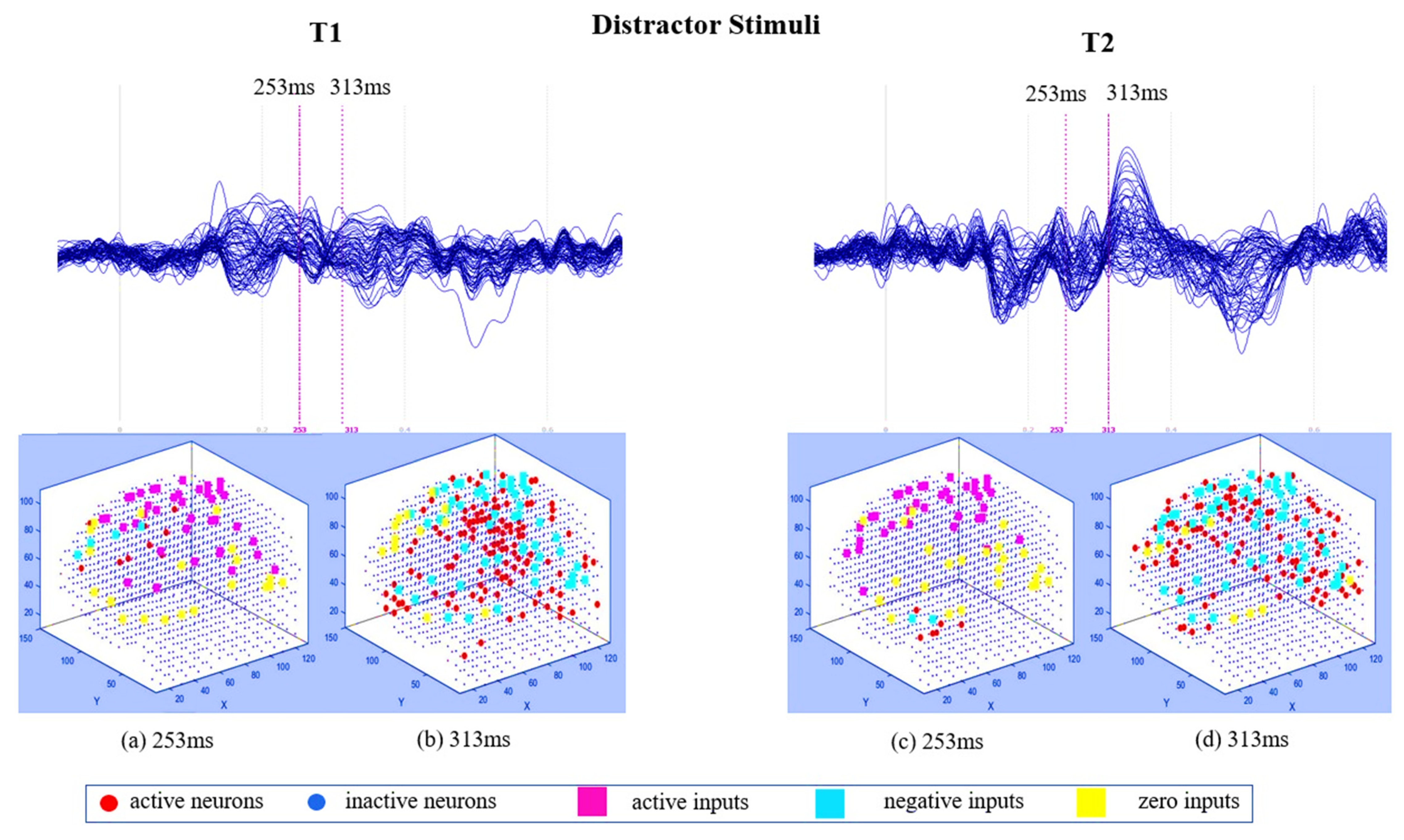
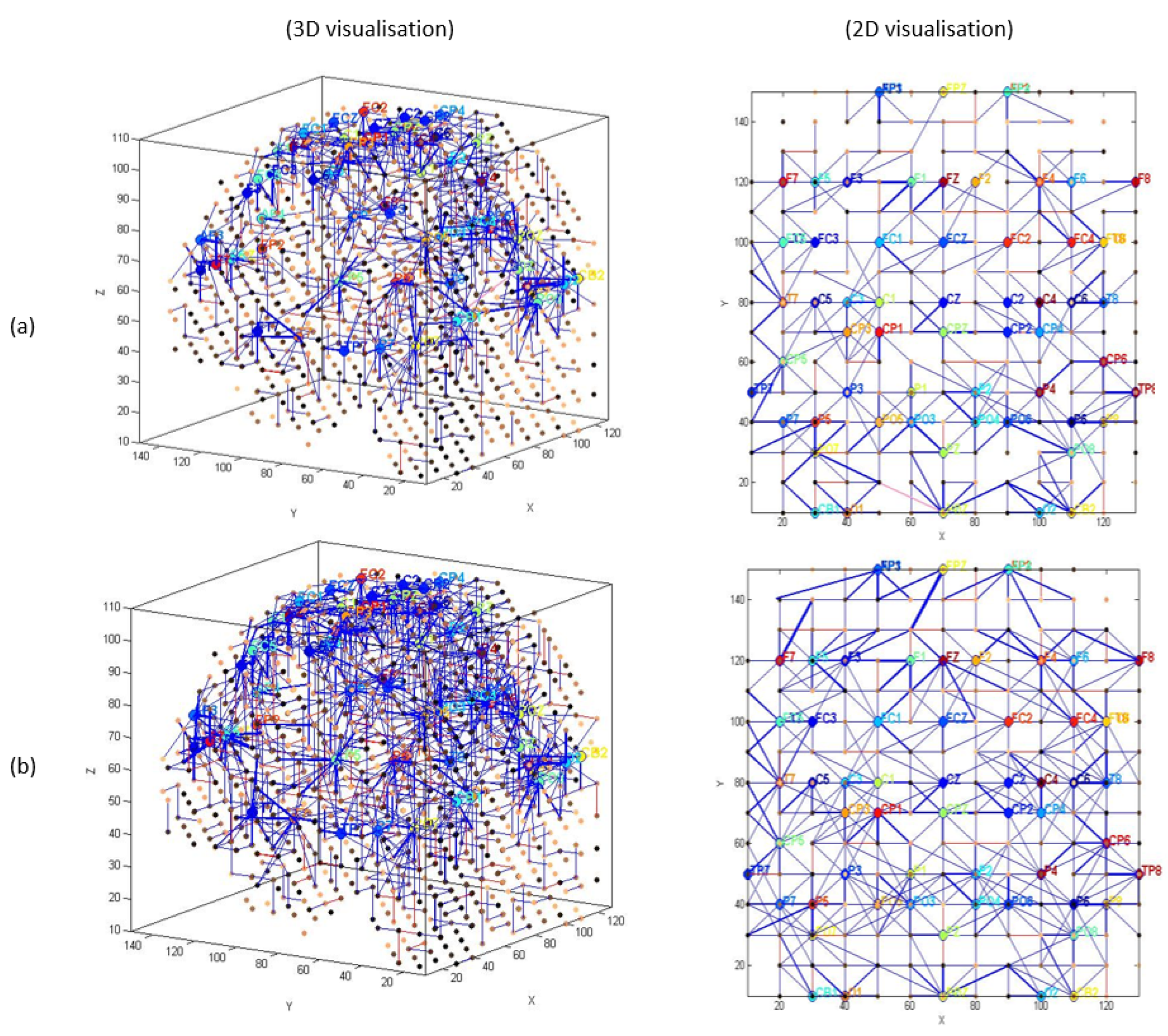
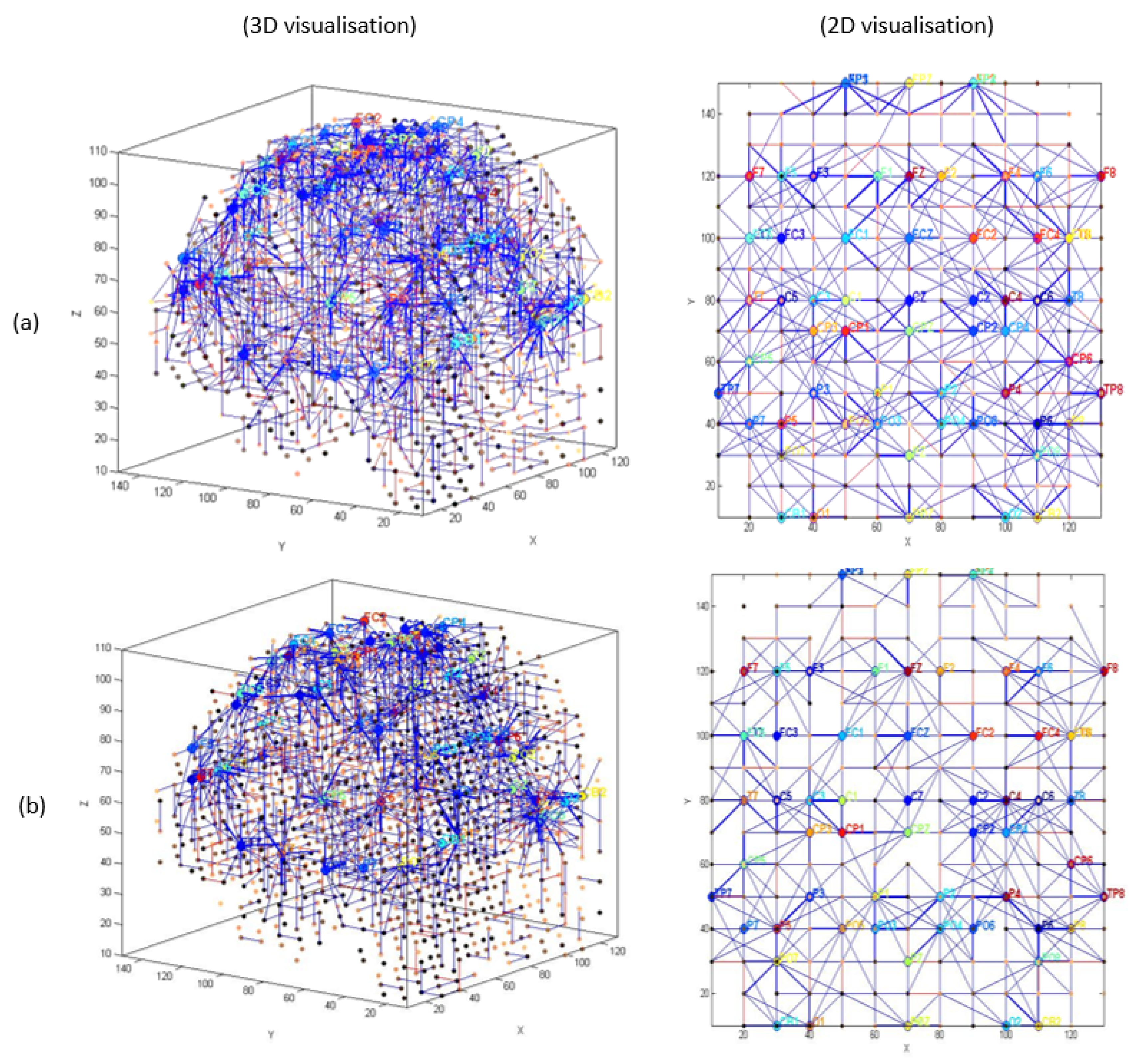
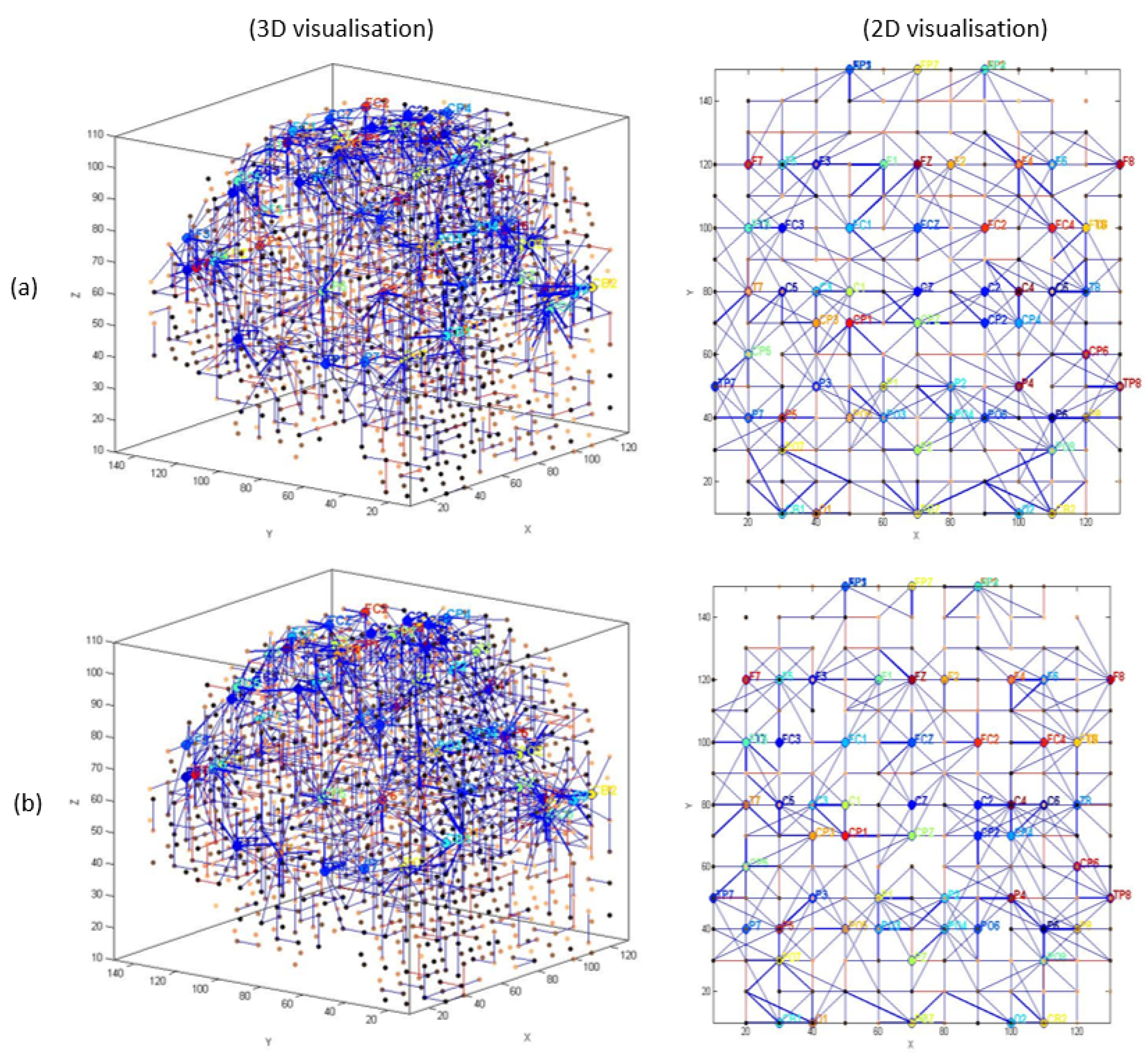
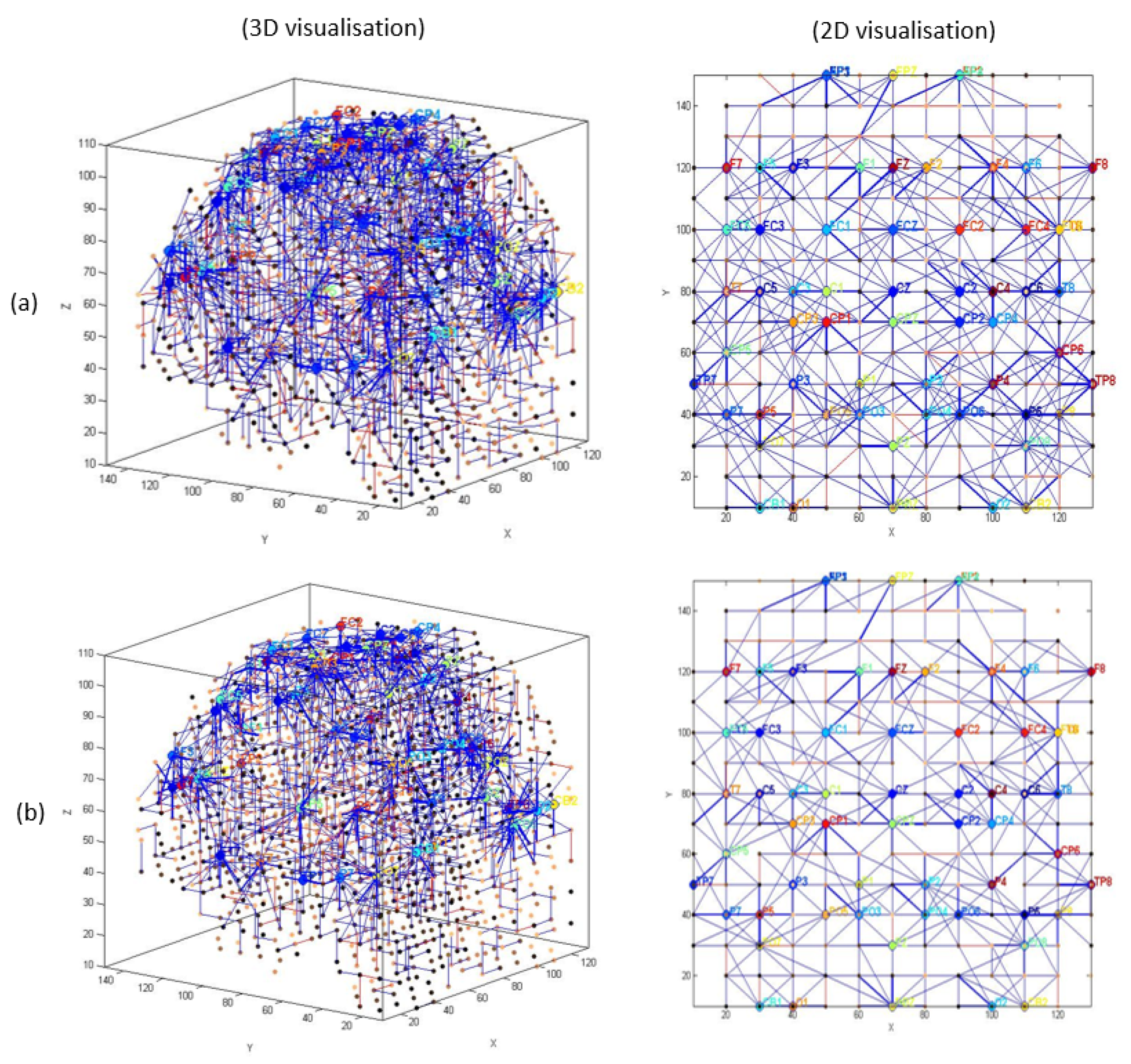
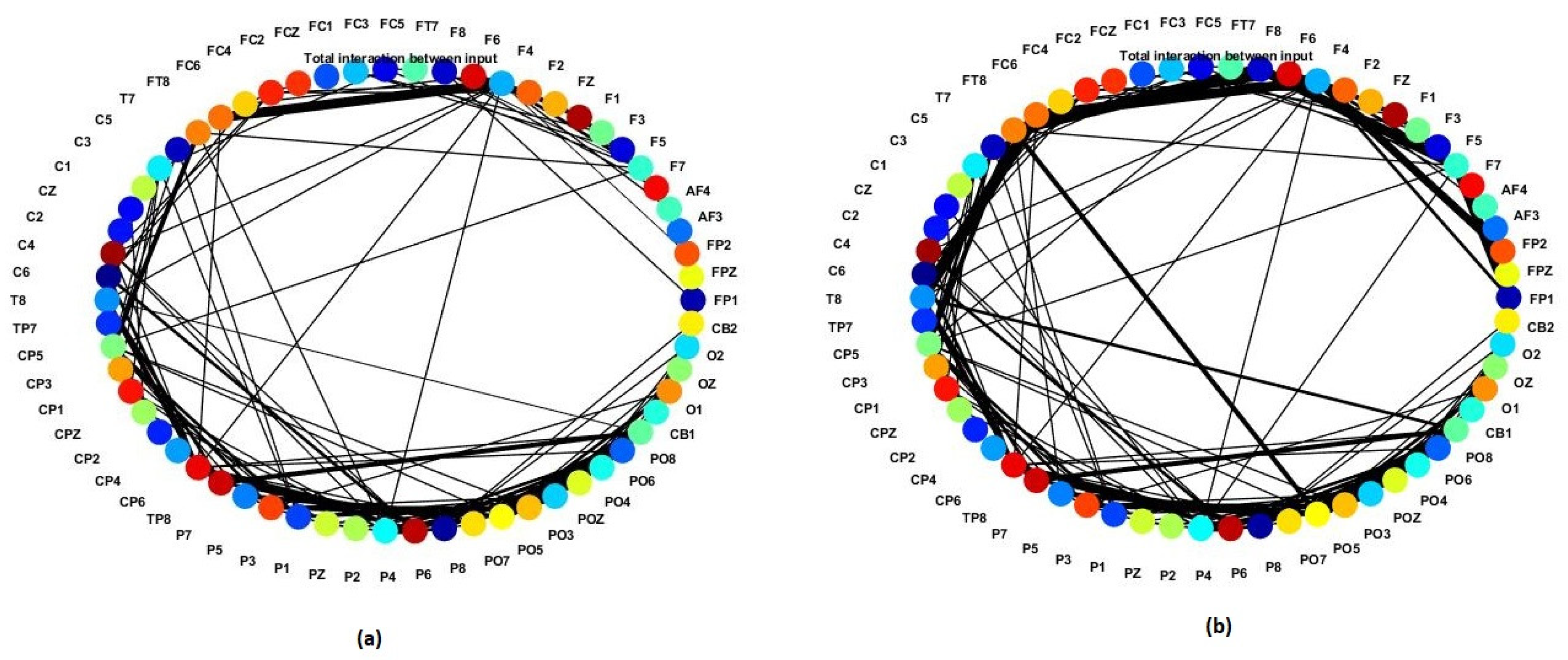
3.1. Pattern Recognition, Visualisation, and Mapping
- (1)
- Mapping, modelling, learning, classifying, and understanding of ERP data.
- (2)
- Statistical and quantitative analysis was performed on the SNN models to assess the model significance.
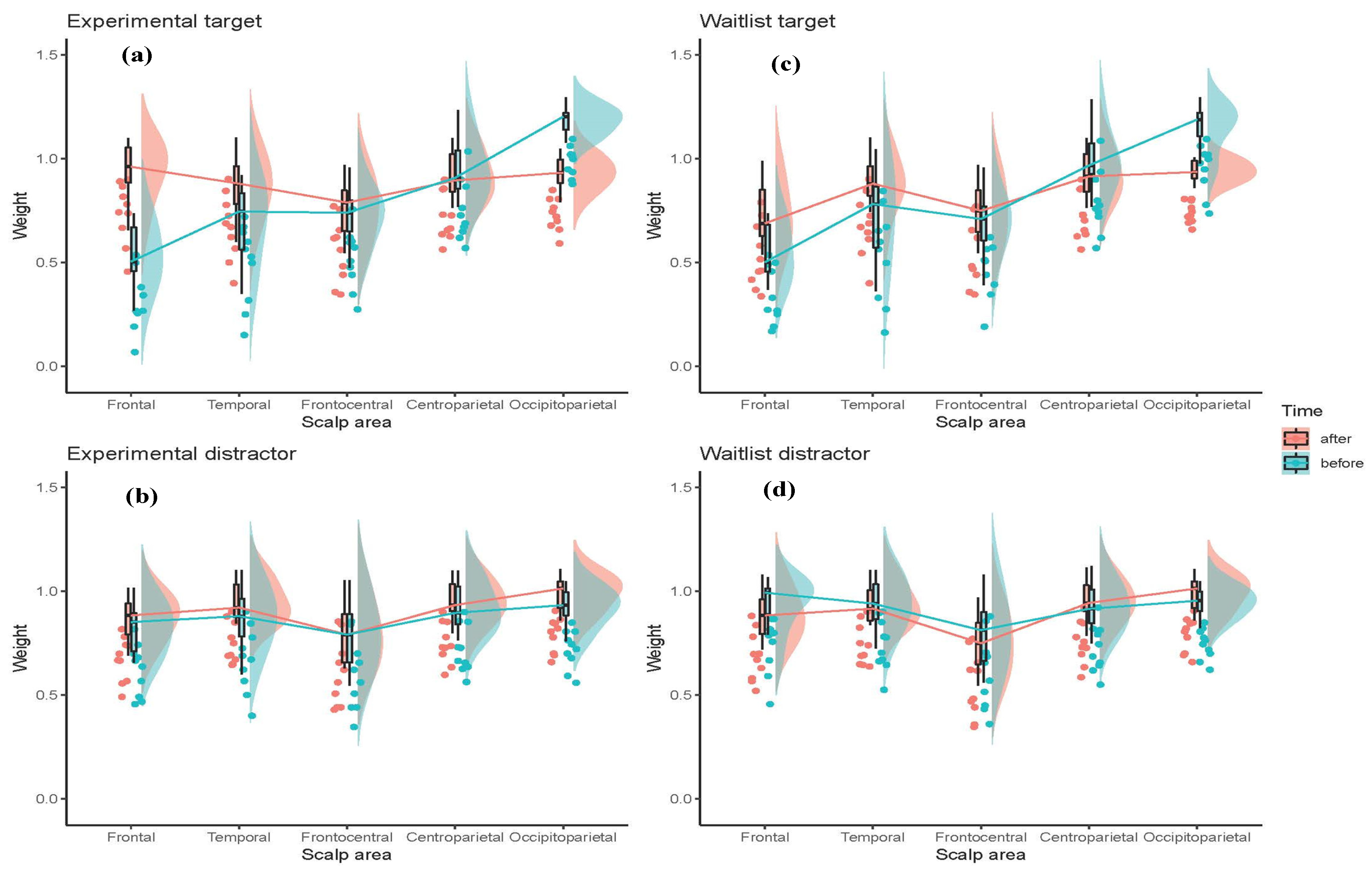
3.2. Statistical Analysis of the SNN Models
3.3. Correlations between Connections Weights, Mindfulness, and Psychometrics Data
3.4. Pattern Classification and Discrimination
4. Discussion
4.1. Comparison of Stimuli (Target vs. Distractor)
4.2. Effect of Mindfulness Training (T1 vs. T2)
4.3. Pattern Classification (T1 vs. T2 vs. Post-Assessment)
4.4. Limitations and Future Research
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Appendix A
- Brain-Inspired: brain-inspired refers to the mathematical implementation of a method that gets inspiration from neuroscience research on brain activity.
- Beck’s Depression Inventory (BDI) measures the severity, depth, and intensity of depression with the Cronbach’s alpha of 0.90.
- The DASS measures the three negative emotional states of anxiety, stress, and depression. Cronbach’s alpha values of 0.89, 0.78, and 0.81 for the subscales of depression, anxiety, and stress, respectively.
- The Five Facet Mindfulness Questionnaire (FFMQ) is a psychometric measure of mindfulness that provides a five-domain profile of various aspects that are characteristic of mindfulness. However, scores may also be interpreted as a single overall score.
- In spiking neural networks, an artificial neuron refers to a computational unit that mimics the behaviour of a biological neuron which receives the information, processes it, and produces an output.
- Connection weights refer to the temporal relationship between two neurons in terms of the time of spike. The average weight of all neural connections (every single EEG channel) in each SNN model are extracted as a metric for comparison.
Appendix B

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Participants | Id | Age | Gender | FFMQ Post-Pre |
|---|---|---|---|---|
| Experimental Group | S1 | 29 | M | 7.18 |
| S2 | 29 | F | 6.82 | |
| S3 | 40 | M | 0.81 | |
| S4 | 47 | F | 10.92 | |
| S5 | 19 | F | 2.01 | |
| S6 | 26 | F | 15.38 | |
| S7 | 23 | M | 6.37 | |
| S8 | 19 | F | 6.37 | |
| S9 | 19 | F | 2.43 | |
| S10 | 37 | F | 6.54 | |
| Mean (SD) | 28.80 (9.74) | 6.48 (4.32) | ||
| Waitlist (Control) Group | S#1 | 22 | F | 7.57 |
| S#2 | 54 | F | 5.99 | |
| S#3 | 30 | M | 10.99 | |
| S#4 | 27 | M | 10.66 | |
| S#5 | 21 | F | 1.34 | |
| S#6 | 37 | F | 0.92 | |
| S#7 | 33 | F | 7.98 | |
| S#8 | 20 | F | 3.92 | |
| S#9 | 27 | F | 2.03 | |
| S#10 | 19 | F | 4.33 | |
| Mean (SD) | 29 (10.58) | 5.57 (3.67) |

References
- Dawson, A.F.; Brown, W.W.; Anderson, J.; Datta, B.; Donald, J.N.; Hong, K.; Allan, S.; Mole, T.B.; Jones, P.B.; Galante, J. Mindfulness-based interventions for university students: A systematic review and meta-analysis of randomised controlled trials. Appl. Psychol. Health Well Being 2019, 12, 384–410. [Google Scholar] [CrossRef] [PubMed]
- Sevilla-Llewellyn-Jones, J.; Santesteban-Echarri, O.; Pryor, I.; McGorry, P.; Alvarez-Jimenez, M.; Firth, J.; Chadi, N. Web-based mindfulness interventions for mental health treatment: Systematic review and meta-analysis. JMIR Ment. Health 2018, 5, e10278. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schumer, M.C.; Lindsay, E.K.; Creswell, J.D. Brief mindfulness training for negative affectivity: A systematic review and meta-analysis. J. Consult. Clin. Psychol. 2018, 86, 569–583. [Google Scholar] [CrossRef] [PubMed]
- Goyal, M.; Singh, S.; Sibinga, E.; Gould, N.; Rowland-Seymour, A.; Sharma, R.; Berger, Z.; Sleicher, D.; Maron, D.; Shihab, H.; et al. Meditation programs for psychological stress and well-being: A systematic review and meta-analysis. JAMA Intern. Med. 2014, 174, 357–368. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rademaker, M.; Stegeman, I.; Ho-Kang-You, K.; Stokroos, R.; Smit, D. The effect of mindfulness-based interventions on tinnitus burden. A systematic review. Front. Neurol. 2019, 10, 1135. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tang, Y.Y.; Hölzel, B.K.; Posner, M.I. The neuroscience of mindfulness meditation. Nat. Rev. Neurosci. 2015, 16, 213. [Google Scholar] [CrossRef] [PubMed]
- Creswell, J.D. Mindfulness interventions. Annu. Rev. Psychol. 2017, 68, 491–516. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Teper, R.; Inzlicht, M. Meditation, mindfulness and executive control: The importance of emotional acceptance and brain-based performance monitoring. Soc. Cogn. Affect. Neurosci. 2013, 8, 85–92. [Google Scholar] [CrossRef] [PubMed]
- Wenk-Sormaz, H. Meditation can reduce habitual responding. Altern. Ther. Health Med. 2005, 11, 42–58. [Google Scholar]
- Moore, A.; Malinowski, P. Meditation, mindfulness and cognitive flexibility. Conscious. Cogn. 2009, 18, 176–186. [Google Scholar] [CrossRef]
- Chambers, R.; Lo, B.C.Y.; Allen, N.B. The impact of intensive mindfulness training on attentional control, cognitive style, and affect. Cogn. Ther. Res. 2008, 32, 303–322. [Google Scholar] [CrossRef]
- Semple, R.J. Does mindfulness meditation enhance attention? A randomized controlled trial. Mindfulness 2010, 1, 121–130. [Google Scholar] [CrossRef]
- Sahdra, B.K.; MacLean, K.A.; Ferrer, E.; Shaver, P.R.; Rosenberg, E.L.; Jacobs, T.L.; Zanesco, A.P.; King, B.G.; Aichele, S.; A Bridwell, D.; et al. Enhanced response inhibition during intensive meditation training predicts improvements in self-reported adaptive socioemotional functioning. Emotion 2011, 11, 299–312. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Young, K.S.; Van Der Velden, A.M.; Craske, M.G.; Pallesen, K.J.; Fjorback, L.; Roepstorff, A.; Parsons, C.E. The impact of mindfulness-based interventions on brain activity: A systematic review of functional magnetic resonance imaging studies. Neurosci. Biobehav. Rev. 2018, 84, 424–433. [Google Scholar] [CrossRef]
- Taren, A.A.; Gianaros, P.J.; Greco, C.M.; Lindsay, E.; Fairgrieve, A.; Brown, K.W.; Rosen, R.K.; Ferris, J.L.; Julson, E.; Marsland, A.L.; et al. Mindfulness meditation training and executive control network resting state functional connectivity: A randomized controlled trial. Psychosom. Med. 2017, 79, 674. [Google Scholar] [CrossRef]
- Lomas, T.; Ivtzan, I.; Fu, C.H. A systematic review of the neurophysiology of mindfulness on EEG oscillations. Neurosci. Biobehav. Rev. 2015, 57, 401–410. [Google Scholar] [CrossRef]
- Cacioppo, J.T.; Tassinary, L.G.; Berntson, G. Handbook of Psychophysiology; Cambridge University Press: Cambridge, UK, 2007. [Google Scholar]
- Aftanas, L.I.; Golosheykin, S. Impact of regular meditation practice on EEG activity at rest and during evoked negative emotions. Int. J. Neurosci. 2005, 115, 893–909. [Google Scholar] [CrossRef]
- Farb, N.A.S.; Anderson, A.K.; Segal, Z.V. The mindful brain and emotion regulation in mood disorders. Can. J. Psychiatry 2012, 57, 70–77. [Google Scholar] [CrossRef] [Green Version]
- Bailey, N.W.; Freedman, G.; Raj, K.; Sullivan, C.M.; Rogasch, N.C.; Chung, S.W.; Hoy, K.E.; Chambers, R.; Hassed, C.; Van Dam, N.T.; et al. Mindfulness meditators show altered distributions of early and late neural activity markers of attention in a response inhibition task. PLoS ONE 2019, 14, e0203096. [Google Scholar] [CrossRef] [Green Version]
- Shanok, N.A.; Reive, C.; Mize, K.D.; Jones, N.A. Mindfulness meditation intervention alters neurophysiological symptoms of anxiety and depression in preadolescents. Psychophysiology 2020, 34, 159–170. [Google Scholar] [CrossRef]
- Creswell, J.D.; Taren, A.A.; Lindsay, E.K.; Greco, C.M.; Gianaros, P.J.; Fairgrieve, A.; Marsland, A.L.; Brown, K.W.; Way, B.M.; Rosen, R.K.; et al. Alterations in resting-state functional connectivity link mindfulness meditation with reduced interleukin-6: A randomized controlled trial. Biol. Psychiatry 2016, 80, 53–61. [Google Scholar] [CrossRef]
- Kim, H.-C.; Tegethoff, M.; Meinlschmidt, G.; Stalujanis, E.; Belardi, A.; Jo, S.; Lee, J.; Kim, D.-Y.; Yoo, S.-S.; Lee, J.-H. Mediation analysis of triple networks revealed functional feature of mindfulness from real-time fMRI neurofeedback. NeuroImage 2019, 195, 409–432. [Google Scholar] [CrossRef]
- Brewer, J.A.; Worhunsky, P.D.; Gray, J.R.; Tang, Y.-Y.; Weber, J.; Kober, H. Meditation experience is associated with differences in default mode network activity and connectivity. Proc. Natl. Acad. Sci. USA 2011, 108, 20254–20259. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schroder, H.S.; Fisher, M.; Lin, Y.; Lo, S.L.; Danovitch, J.H.; Moser, J.S. Neural evidence for enhanced attention to mistakes among school-aged children with a growth mindset. Dev. Cogn. Neurosci. 2017, 24, 42–50. [Google Scholar] [CrossRef]
- Campos, E.; Hazlett, C.; Tan, P.; Truong, H.; Loo, S.; Distefano, C.; Jeste, S.; Şentürk, D. Principle ERP reduction and analysis: Estimating and using principle ERP waveforms underlying ERPs across tasks, subjects and electrodes. NeuroImage 2020, 212, 116630. [Google Scholar] [CrossRef]
- Luck, S.J. An Introduction to the Event-Related Potential Technique; MIT Press: Cambridge, MA, USA, 2014. [Google Scholar]
- Van Leeuwen, S.; Singer, W.; Melloni, L. Meditation increases the depth of information processing and improves the allocation of attention in space. Front. Hum. Neurosci. 2012, 6, 133. [Google Scholar] [CrossRef] [Green Version]
- Norris, C.J.; Creem, D.; Hendler, R.; Kober, H. Brief mindfulness meditation improves attention in novices: Evidence from ERPs and moderation by neuroticism. Front. Hum. Neurosci. 2018, 12, 315. [Google Scholar] [CrossRef] [Green Version]
- Klopsis, A.L. The Impact of a Single Session of Mindfulness Meditation on the Attentional Blink in Non-Meditators. Master’s Thesis, City University of New York, New York, NY, USA, 2020. [Google Scholar]
- Sanger, K.L.; Dorjee, D. Mindfulness training for adolescents: A neurodevelopmental perspective on investigating modifications in attention and emotion regulation using event-related brain potentials. Cogn. Affect. Behav. Neurosci. 2015, 15, 696–711. [Google Scholar] [CrossRef] [Green Version]
- Zhang, W.; Ouyang, Y.; Tang, F.; Chen, J.; Li, H. Breath-focused mindfulness alters early and late components during emotion regulation. Brain Cogn. 2019, 135, 103585. [Google Scholar] [CrossRef]
- Eddy, M.D.; Brunyé, T.T.; Tower-Richardi, S.; Mahoney, C.R.; Taylor, H.A. The effect of a brief mindfulness induction on processing of emotional images: An ERP study. Front. Psychol. 2015, 6, 1391. [Google Scholar] [CrossRef] [Green Version]
- Isbel, B.; Lagopoulos, J.; Hermens, D.; Stefanidis, K.; Summers, M.J. Mindfulness improves attention resource allocation during response inhibition in older adults. Mindfulness 2020, 11, 1500–1510. [Google Scholar] [CrossRef]
- Donkers, F.C.L.; Van Boxtel, G.J. The N2 in go/no-go tasks reflects conflict monitoring not response inhibition. Brain Cogn. 2004, 56, 165–176. [Google Scholar] [CrossRef]
- Falkenstein, M. Inhibition, conflict and the Nogo-N2. Clin. Neurophysiol. 2006, 117, 1638–1640. [Google Scholar] [CrossRef]
- Luck, S.J.; Kappenman, E.S. The Oxford Handbook of Event-Related Potential Components; Oxford University Press: Oxford, UK, 2011. [Google Scholar]
- Linden, D.E. The P300: Where in the brain is it produced and what does it tell us? Neuroscience 2005, 11, 563–576. [Google Scholar] [CrossRef]
- Justo-Guillén, E.; Ricardo-Garcell, J.; Rodríguez-Camacho, M.; Rodríguez-Agudelo, Y.; de Larrea-Mancera, E.S.L.; Solís-Vivanco, R. Auditory mismatch detection, distraction, and attentional reorientation (MMN-P3a-RON) in neurological and psychiatric disorders: A review. Intern. J. Psychophysiol. 2019, 146, 85–100. [Google Scholar] [CrossRef]
- Gow, R.V.; Matsudaira, T.; Taylor, E.; Rubia, K.; Crawford, M.; Ghebremeskel, K.; Ibrahimovic, A.; Vallée-Tourangeau, F.; Williams, L.M.; Sumich, A. Total red blood cell concentrations of ω-3 fatty acids are associated with emotion-elicited neural activity in adolescent boys with attention-deficit hyperactivity disorder. Prostaglandins Leukot. Essent. Fat. Acids 2009, 80, 151–156. [Google Scholar] [CrossRef]
- Polich, J. Updating P300: An integrative theory of P3a and P3b. Clin. Neurophysiol. 2007, 118, 2128–2148. [Google Scholar] [CrossRef] [Green Version]
- Wongupparaj, P.; Sumich, A.; Wickens, M.; Kumari, V.; Morris, R.G. Individual differences in working memory and general intelligence indexed by P200 and P300: A latent variable model. Biol. Psychol. 2018, 139, 96–105. [Google Scholar] [CrossRef] [Green Version]
- Sumich, A.; Kumari, V.; Dodd, P.; Ettinger, U.; Hughes, C.; Zachariah, E.; Sharma, T. N100 and P300 amplitude to Go and No–Go variants of the auditory oddball in siblings discordant for schizophrenia. Schizophr. Res. 2008, 98, 265–277. [Google Scholar] [CrossRef]
- Gow, R.V.; Rubia, K.; Taylor, E.; Vallée-Tourangeau, F.; Matsudaira, T.; Ibrahimovic, A.; Sumich, A. Abnormal centroparietal ERP response in predominantly medication-naive adolescent boys with ADHD during both response inhibition and execution. J. Clin. Neurophysiol. 2012, 29, 181–189. [Google Scholar] [CrossRef]
- Huster, R.J.; Enriquez-Geppert, S.; Lavallee, C.F.; Falkenstein, M.; Herrmann, C.S. Electroencephalography of response inhibition tasks: Functional networks and cognitive contributions. Int. J. Psychophysiol. 2013, 87, 217–233. [Google Scholar] [CrossRef]
- Cahn, B.R.; Polich, J. Meditation (Vipassana) and the P3a event-related brain potential. Int. J. Psychophysiol. 2009, 72, 51–60. [Google Scholar] [CrossRef] [Green Version]
- Moore, A.W.; Egruber, T.; Ederose, J.; Malinowski, P. Regular, brief mindfulness meditation practice improves electrophysiological markers of attentional control. Front. Hum. Neurosci. 2012, 6, 18. [Google Scholar] [CrossRef] [Green Version]
- Singh, N.; Telles, S. Neurophysiological effects of meditation based on evoked and event related potential recordings. BioMed Res. Int. 2015, 2015, 406261. [Google Scholar] [CrossRef]
- Kakumanu, R.J.; Nair, A.K.; Sasidharan, A.; John, J.P.; Mehrotra, S.; Panth, R.; Kutty, B.M. State-trait influences of Vipassana meditation practice on P3 EEG dynamics. Prog. Brain Res. 2019, 244, 115–136. [Google Scholar] [PubMed]
- Suskauer, S.J.; Simmonds, D.J.; Fotedar, S.; Blankner, J.G.; Pekar, J.J.; Denckla, M.B.; Mostofsky, S.H. Functional magnetic resonance imaging evidence for abnormalities in response selection in attention deficit hyperactivity disorder: Differences in activation associated with response inhibition but not habitual motor response. J. Cogn. Neurosci. 2008, 20, 478–493. [Google Scholar] [CrossRef] [PubMed]
- Kozasa, E.H.; Sato, J.R.; Lacerda, S.S.; Barreiros, M.A.M.; Radvany, J.; Russell, T.A.; Sanches, L.G.; Mello, L.E.A.M.; Amaro, E. Meditation training increases brain efficiency in an attention task. NeuroImage 2012, 59, 745–749. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Baerentsen, K.B.; Hartvig, N.V.; Stødkilde-Jørgensen, H.; Mammen, J. Onset of meditation explored with fMRI. NeuroImage 2001, 13, 297. [Google Scholar] [CrossRef]
- Lazar, M.; Weinstein, D.M.; Tsuruda, J.S.; Hasan, K.M.; Arfanakis, K.; Meyerand, M.; Badie, B.; Rowley, H.; Haughton, V.; Field, A.; et al. White matter tractography using diffusion tensor deflection. Hum. Brain Mapp. 2003, 18, 306–321. [Google Scholar] [CrossRef]
- Lazar, S.W.; Kerr, C.E.; Wasserman, R.H.; Gray, J.R.; Greve, D.N.; Treadway, M.T.; McGarvey, M.; Quinn, B.T.; Dusek, J.A.; Benson, H.; et al. Meditation experience is associated with increased cortical thickness. NeuroReport 2005, 16, 1893–1897. [Google Scholar] [CrossRef] [Green Version]
- Doborjeh, Z. Modelling of Spatiotemporal EEG and ERP Brain Data for Dynamic Pattern Recognition and Brain State Prediction using Spiking Neural Networks: Methods and Applications in Psychology. Ph.D. Thesis, Auckland University of Technology, Auckland, Australia, 2019. [Google Scholar]
- Doborjeh, Z.; Doborjeh, M.; Taylor, T.; Kasabov, N.; Wang, G.Y.; Siegert, R.; Sumich, A. Spiking neural network modelling approach reveals how mindfulness training rewires the brain. Sci. Rep. 2019, 9, 1–15. [Google Scholar] [CrossRef] [PubMed]
- Doborjeh, Z.; Kasabov, N.; Doborjeh, M.G.; Sumich, A. Modelling peri-perceptual brain processes in a deep learning spiking neural network architecture. Sci. Rep. 2018, 8, 1–13. [Google Scholar]
- Durai, M.; Sanders, P.; Doborjeh, Z.; Wendt, A.; Kasabov, N.; Searchfield, G.D. Prediction of tinnitus masking benefit within a case series using a spiking neural network model. In Progress in Brain Research; Waxman, S., Stein, D.G., Swaab, D., Fields, H., Eds.; Elsevier: Amsterdam, The Netherlands, 2020. [Google Scholar]
- Kasabov, N.; Doborjeh, M.G.; Doborjeh, Z. Mapping, learning, visualization, classification, and understanding of fMRI data in the NeuCube evolving spatiotemporal data machine of spiking neural networks. IEEE Trans. Neural Netw. Learn. Syst. 2016, 28, 887–899. [Google Scholar] [CrossRef] [PubMed]
- Doborjeh, Z.G.; Doborjeh, M.G.; Kasabov, N. Attentional bias pattern recognition in spiking neural networks from spatio-temporal EEG data. Cogn. Comput. 2018, 10, 35–48. [Google Scholar] [CrossRef]
- Moore, M.; Shafer, A.T.; Bakhtiari, R.; Dolcos, F.; Singhal, A. Integration of spatio-temporal dynamics in emotion-cognition interactions: A simultaneous fMRI-ERP investigation using the emotional oddball task. NeuroImage 2019, 202, 116078. [Google Scholar] [CrossRef] [PubMed]
- Holzinger, A. Machine Learning for Health Informatics: State-of-the-Art and Future Challenges; Springer: Berlin/Heidelberg, Germany, 2016. [Google Scholar]
- Ratsch, G.; Sonnenburg, S.; Schäfer, C. Learning interpretable SVMs for biological sequence classification. BMC Bioinform. 2006, 7, 1–14. [Google Scholar] [CrossRef]
- Ieracitano, C.; Adeel, A.; Morabito, F.C.; Hussain, A. A novel statistical analysis and autoencoder driven intelligent intrusion detection approach. Neurocomputing 2020, 387, 51–62. [Google Scholar] [CrossRef]
- Mammone, N.; Ieracitano, C.; Morabito, F.C. A deep CNN approach to decode motor preparation of upper limbs from time–frequency maps of EEG signals at source level. Neural Netw. 2020, 124, 357–372. [Google Scholar] [CrossRef]
- Capecci, E.; Doborjeh, Z.G.; Mammone, N.; La Foresta, F.; Morabito, F.C.; Kasabov, N. Longitudinal study of alzheimer’s disease degeneration through EEG data analysis with a NeuCube spiking neural network model. In Proceedings of the 2016 International Joint Conference on Neural Networks (IJCNN), Vancouver, BC, Canada, 24–29 July 2016; pp. 1360–1366. [Google Scholar]
- Lotte, F.; Congedo, M.; Lecuyer, A.; Lamarche, F.; Arnaldi, B. A review of classification algorithms for EEG-based brain–computer interfaces. J. Neural Eng. 2007, 4, R1–R13. [Google Scholar] [CrossRef]
- Izhikevich, E.M. Polychronization: Computation with spikes. Neural Comput. 2006, 18, 245–282. [Google Scholar] [CrossRef]
- Kasabov, N.; Dhoble, K.; Nuntalid, N.; Indiveri, G. Dynamic evolving spiking neural networks for on-line spatio- and spectro-temporal pattern recognition. Neural Netw. 2013, 41, 188–201. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kasabov, N.K. Time-Space, Spiking Neural Networks and Brain-Inspired Artificial Intelligence; Springer: Berlin/Heidelberg, Germany, 2018. [Google Scholar]
- Bharatharaj, J.; Medvedev, O.N.; Taylor, T.; Wrapson, W.; Rix, G.; Sumich, A.; Wang, G.Y.; Csako, R.; Anstiss, D.; Ranta, J.T.; et al. A Pilot Randomized Controlled Trial for a Videoconference-Delivered Mindfulness-Based Group Intervention in a Nonclinical Setting. Mindfulness 2018, 10, 700–711. [Google Scholar] [CrossRef] [Green Version]
- Lee, E.H.; Lee, S.J.; Hwang, S.T.; Hong, S.H.; Kim, J.M. Reliability and validity of the Beck depression inventory-II among Korean adolescents. Psychiatry Investig. 2017, 14, 30. [Google Scholar] [CrossRef] [Green Version]
- Tran, T.D.; Tran, T.; Fisher, J. Validation of the depression anxiety stress scales (DASS) 21 as a screening instrument for depression and anxiety in a rural community-based cohort of northern Vietnamese women. BMC Psychiatry 2013, 13, 24. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gu, J.; Strauss, C.; Crane, C.; Barnhofer, T.; Karl, A.; Cavanagh, K.; Kuyken, W. Examining the factor structure of the 39-item and 15-item versions of the five facet mindfulness questionnaire before and after mindfulness-based cognitive therapy for people with recurrent depression. Psychol. Assess. 2016, 28, 791. [Google Scholar] [CrossRef] [Green Version]
- Heym, N.; Heasman, B.C.; Hunter, K.; Blanco, S.R.; Wang, G.Y.; Siegert, R.; Cleare, A.; Gibson, G.R.; Kumari, V.; Sumich, A.L. The role of microbiota and inflammation in self-judgement and empathy: Implications for understanding the brain-gut-microbiome axis in depression. Psychopharmacology 2019, 236, 1459–1470. [Google Scholar] [CrossRef] [Green Version]
- Baer, R.A.; Smith, G.T.; Hopkins, J.; Krietemeyer, J.; Toney, L. Using self-report assessment methods to explore facets of mindfulness. Assessment 2006, 13, 27–45. [Google Scholar] [CrossRef] [Green Version]
- Medvedev, O.N.; Krägeloh, C.U.; Narayanan, A.; Siegert, R.J. Measuring mindfulness: Applying generalizability theory to distinguish between state and trait. Mindfulness 2017, 8, 1036–1046. [Google Scholar] [CrossRef]
- Devcich, D.A.; Rix, G.; Bernay, R.; Graham, E. Effectiveness of a mindfulness-based program on school children’s self-reported well-being: A pilot study comparing effects with an emotional literacy program. J. Appl. Sch. Psychol. 2017, 33, 309–330. [Google Scholar] [CrossRef]
- Kasabov, N. NeuCube: A spiking neural network architecture for mapping, learning and understanding of spatio-temporal brain data. Neural Netw. 2014, 52, 62–76. [Google Scholar] [CrossRef]
- Hansen, S.T.; Hansen, L.K. Spatio-temporal reconstruction of brain dynamics from EEG with a Markov prior. NeuroImage 2017, 148, 274–283. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Murtagh, F. Multilayer perceptrons for classification and regression. Neurocomputing 1991, 2, 183–197. [Google Scholar] [CrossRef]
- Yao, X.J.; Panaye, A.; Doucet, J.P.; Zhang, R.S.; Chen, H.F.; Liu, M.C.; Hu, Z.D.; Fan, B.T. Comparative study of QSAR/QSPR correlations using support vector machines, radial basis function neural networks, and multiple linear regression. J. Chem. Inf. Comput. Sci. 2004, 44, 1257–1266. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bullmore, E.T.; Sporns, O. Complex brain networks: Graph theoretical analysis of structural and functional systems. Nat. Rev. Neurosci. 2009, 10, 186–198. [Google Scholar] [CrossRef]
- Tu, E.; Kasabov, N.; Yang, J. Mapping temporal variables into the NeuCube for improved pattern recognition, predictive modeling, and understanding of stream data. IEEE Trans. Neural Netw. Learn. Syst. 2016, 28, 1305–1317. [Google Scholar] [CrossRef] [Green Version]
- Talairach, J.; Tournoux, P. Co-Planar Stereotaxic Atlas of the Human Brain: 3-Dimensional Proportional System: An Approach to Cerebral Imaging; Thieme: New York, NY, USA, 1988. [Google Scholar]
- Liao, X.; Vasilakos, A.V.; He, Y. Small-world human brain networks: Perspectives and challenges. Neurosci. Biobehav. Rev. 2017, 77, 286–300. [Google Scholar] [CrossRef]
- Masquelier, T.; Guyonneau, R.; Thorpe, S.J. Competitive STDP-based spike pattern learning. Neural Comput. 2009, 21, 1259–1276. [Google Scholar] [CrossRef]
- Casula, E.P.; Pellicciari, M.C.; Picazio, S.; Caltagirone, C.; Koch, G. Spike-timing-dependent plasticity in the human dorso-lateral prefrontal cortex. NeuroImage 2016, 143, 204–213. [Google Scholar] [CrossRef]
- Van Schaik, A.; Liu, S.-C. AER EAR: A Matched Silicon Cochlea Pair with Address Event Representation Interface. IEEE Trans. Circuits Syst. I Regul. Pap. 2005, 54, 48–59. [Google Scholar] [CrossRef]
- Beck, A.T.; Alford, B.A. Depression: Causes and Treatment; University of Pennsylvania Press: Philadelphia, PA, USA, 2009. [Google Scholar]
- Beaufort, I.N.; Oene, G.H.D.W.-V.; Buwalda, V.A.; De Leeuw, J.R.J.; Goudriaan, A.E. The depression, anxiety and stress scale (DASS-21) as a screener for depression in substance use disorder in patients: A pilot study. Eur. Addict. Res. 2017, 23, 260–268. [Google Scholar] [CrossRef]
- Gardner, M.W.; Dorling, S.R. Artificial neural networks (the multilayer perceptron)—A review of applications in the atmospheric sciences. Atmos. Environ. 1998, 32, 2627–2636. [Google Scholar] [CrossRef]
- Costantini, G.; Todisco, M.; Casali, D.; Carota, M.; Saggio, G.; Bianchi, L.; Abbafati, M.; Quitadamo, L.R. SVM Classification of EEG Signals for Brain Computer Interface. In Proceedings of the 19th Italian Workshop on Neural Nets, Vietri sul Mare, Salerno, Italy, 28–30 May 2009; pp. 229–233. [Google Scholar]
- Singh-Curry, V.; Husain, M. The functional role of the inferior parietal lobe in the dorsal and ventral stream dichotomy. Neuropsychologia 2009, 47, 1434–1448. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bledowski, C. Localizing P300 generators in visual target and distractor processing: A combined event-related potential and functional magnetic resonance imaging study. J. Neurosci. 2004, 24, 9353–9360. [Google Scholar] [CrossRef] [PubMed]
- Salisbury, D.F.; Griggs, C.B.; Shenton, M.E.; McCarley, R.W. The NoGo P300 ‘anteriorization’effect and response inhibition. Clin. Neurophysiol. 2004, 115, 1550–1558. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Goldstein, A.; Spencer, K.M.; Donchin, E. The influence of stimulus deviance and novelty on the P300 and novelty P3. Psychophysiology 2002, 39, 781–790. [Google Scholar] [CrossRef] [PubMed]
- Courchesne, E.; A Hillyard, S.; Galambos, R. Stimulus novelty, task relevance and the visual evoked potential in man. Electroencephalogr. Clin. Neurophysiol. 1975, 39, 131–143. [Google Scholar] [CrossRef] [Green Version]
- Li, B.-J.; Friston, K.; Mody, M.; Wang, H.-N.; Lu, H.; Hu, D.-W. A brain network model for depression: From symptom understanding to disease intervention. CNS Neurosci. Ther. 2018, 24, 1004–1019. [Google Scholar] [CrossRef]
- Lao, S.-A.; Kissane, D.; Meadows, G. Cognitive effects of MBSR/MBCT: A systematic review of neuropsychological outcomes. Conscious. Cogn. 2016, 45, 109–123. [Google Scholar] [CrossRef]
- Jensen, C.G.; Vangkilde, S.; Frokjaer, V.; Hasselbalch, S.G. Mindfulness training affects attention—Or is it attentional effort? J. Exp. Psychol. Gen. 2012, 141, 106–123. [Google Scholar] [CrossRef] [Green Version]
- Gill, L.-N.; Renault, R.; Campbell, E.; Rainville, P.; Khoury, B. Mindfulness induction and cognition: A systematic review and meta-analysis. Conscious. Cogn. 2020, 84, 102991. [Google Scholar] [CrossRef]
- Sevinc, G.; Hölzel, B.K.; Hashmi, J.; Greenberg, J.; McCallister, A.; Treadway, M.; Schneider, M.L.; Dusek, J.A.; Carmody, J.; Lazar, S.W. Common and dissociable neural activity after mindfulness-based stress reduction and relaxation response programs. Psychosom. Med. 2018, 80, 439–451. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Anderson, N.D.; Lau, M.A.; Segal, Z.V.; Bishop, S.R. Mindfulness-based stress reduction and attentional control. Clin. Psychol. Psychother. 2007, 14, 449–463. [Google Scholar] [CrossRef]
- Vega, B.R.; Melero, J.; Pérez, C.B.; Cebolla, S.; Mira, J.; Valverde, C.; Fernández-Liria, A. Impact of mindfulness training on attentional control and anger regulation processes for psychotherapists in training. Psychother. Res. 2013, 24, 202–213. [Google Scholar] [CrossRef] [PubMed]
- Delgado-Pastor, L.C.; Perakakis, P.; Subramanya, P.; Telles, S.; Vila, J. Mindfulness (Vipassana) meditation: Effects on P3b event-related potential and heart rate variability. Int. J. Psychophysiol. 2013, 90, 207–214. [Google Scholar] [CrossRef] [PubMed]
- Garrison, K.A.; Scheinost, D.; Constable, R.T.; Brewer, J.A. BOLD signal and functional connectivity associated with loving kindness meditation. Brain Behav. 2014, 4, 337–347. [Google Scholar] [CrossRef]
- Zeidan, F.; Baumgartner, J.N.; Coghill, R. The neural mechanisms of mindfulness-based pain relief: A functional magnetic resonance imaging-based review and primer. Pain Rep. 2019, 4, e759. [Google Scholar] [CrossRef]










| The Averaged Connection Weight of SNN Models | |||||
|---|---|---|---|---|---|
| Group | Site | Time Point | |||
| T1 | T2 | ||||
| Type of Stimuli | |||||
| Target | Distractor | Target | Distractor | ||
| Experimental | Frontal | 0.53 | 0.82 | 0.94 | 0.87 |
| Temporal | 0.69 | 0.87 | 0.87 | 0.96 | |
| Frontocentral | 0.72 | 0.80 | 0.77 | 0.81 | |
| Centroparietal | 0.95 | 0.92 | 0.92 | 0.95 | |
| Occipitoparietal | 1.18 | 0.92 | 0.93 | 0.99 | |
| Total weight | 0.81 | 0.86 | 0.86 | 0.91 | |
| Waitlist | Frontal | 0.55 | 0.94 | 0.73 | 0.87 |
| Temporal | 0.74 | 0.94 | 0.88 | 0.95 | |
| Frontocentral | 0.7 | 0.81 | 0.75 | 0.85 | |
| Centroparietal | 0.99 | 0.92 | 0.92 | 0.95 | |
| Occipitoparietal | 1.15 | 0.95 | 0.94 | 0.99 | |
| Total weight | 0.82 | 0.91 | 0.84 | 0.92 | |
| Variables | F-Value | Degrees of Freedom | p-Value | Partial-eta2 (ηp2) |
|---|---|---|---|---|
| Site | 18.432 | 4,72 | <0.001 | 0.506 |
| Time | 24.468 | 1,18 | <0.001 | 0.576 |
| Stimuli | 38.405 | 1,18 | <0.001 | 0.681 |
| Time * Group | 14.962 | 1,18 | 0.001 | 0.454 |
| Hemisphere * Stimuli | 9.815 | 1,18 | 0.006 | 0.353 |
| Site * Time † | 44.631 | 2.761,49.702 | <0.001 | 0.713 |
| Site * Stimuli | 40.842 | 2.135,38.434 | <0.001 | 0.694 |
| Time * Stimuli | 5.786 | 1,18 | 0.027 | 0.243 |
| Site * Time * Group | 5.931 | 2.761,49.702 | 0.002 | 202 |
| SNN-Based Methodology (Experimental Group) | ||||||
| ERP Data Classes | Target Stimuli at T1(C1) | Target Stimuli at T2 (C2) | Target Stimuli at T3 (C3) | Accuracy (%) | Total Accuracy (%) | F-Score (%) |
| Target Stimuli at T1(C1) | 7 | 1 | 2 | 70.00 | 73.00 | 79.00 |
| Target Stimuli at T2 (C2) | 1 | 9 | 0 | 90.00 | ||
| Target Stimuli at T3 (C3) | 2 | 2 | 6 | 60.00 | ||
| Traditional Machine Learning Methods | ||||||
| Methods Accuracy in % | MLP 50 | SVM 58.50 | MLR 48.50 | |||
| SNN-based methodology (waitlist Group) | ||||||
| ERP data Classes | Target Stimuli at T1(C1) | Target Stimuli at T2 (C2) | Target Stimuli at T3 (C3) | Accuracy (%) | Total accuracy % | F-Score (%) |
| Target Stimuli at T1(C1) | 7 | 2 | 1 | 70.00 | 73.00 | 79.00 |
| Target Stimuli at T2 (C2) | 1 | 7 | 2 | 70.00 | ||
| Target Stimuli at T3 (C3) | 1 | 1 | 8 | 80.00 | ||
| Traditional Machine Learning Methods | ||||||
| Methods Accuracy in % | MLP 59.50 | SVM 51.50 | MLR 55.50 | |||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Doborjeh, Z.; Doborjeh, M.; Crook-Rumsey, M.; Taylor, T.; Wang, G.Y.; Moreau, D.; Krägeloh, C.; Wrapson, W.; Siegert, R.J.; Kasabov, N.; et al. Interpretability of Spatiotemporal Dynamics of the Brain Processes Followed by Mindfulness Intervention in a Brain-Inspired Spiking Neural Network Architecture. Sensors 2020, 20, 7354. https://0-doi-org.brum.beds.ac.uk/10.3390/s20247354
Doborjeh Z, Doborjeh M, Crook-Rumsey M, Taylor T, Wang GY, Moreau D, Krägeloh C, Wrapson W, Siegert RJ, Kasabov N, et al. Interpretability of Spatiotemporal Dynamics of the Brain Processes Followed by Mindfulness Intervention in a Brain-Inspired Spiking Neural Network Architecture. Sensors. 2020; 20(24):7354. https://0-doi-org.brum.beds.ac.uk/10.3390/s20247354
Chicago/Turabian StyleDoborjeh, Zohreh, Maryam Doborjeh, Mark Crook-Rumsey, Tamasin Taylor, Grace Y. Wang, David Moreau, Christian Krägeloh, Wendy Wrapson, Richard J. Siegert, Nikola Kasabov, and et al. 2020. "Interpretability of Spatiotemporal Dynamics of the Brain Processes Followed by Mindfulness Intervention in a Brain-Inspired Spiking Neural Network Architecture" Sensors 20, no. 24: 7354. https://0-doi-org.brum.beds.ac.uk/10.3390/s20247354