Anti-Proliferative and Genotoxic Activities of the Helichrysum petiolare Hilliard & B.L. Burtt


School of Biology and Environmental Sciences, University of Mpumalanga, Private Bag X11283, Mbombela 1200, South Africa
*
Author to whom correspondence should be addressed.
Sci. Pharm. 2020, 88(4), 49; https://0-doi-org.brum.beds.ac.uk/10.3390/scipharm88040049
Submission received: 10 July 2020
/
Revised: 9 August 2020
/
Accepted: 24 August 2020
/
Published: 20 October 2020
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:Helichrysum petiolare (Asteraceae family) is part of the Helichrysum genus which comprises of an estimated 600 species. Several parts of the plant have been used traditionally for the treatment of various ailments, such as cough, infection, asthma, chest problems, diabetes and wounds. Given its various chemical constituents with anticancer properties, there has been no scientific evidence of its usage for the treatment of cancer. This study aims to investigate the anti-proliferative and genotoxic activities of H. petiolare methanol extract. The cytotoxic effect and cell cycle analysis of mouse melanoma cells (B16F10) and human melanoma cells (MeWo) were assessed using the ImageXpress Micro XLS Widefield High-Content Analysis System. The genotoxic potential of the extract towards Vero cells was also assessed using the micronucleus assay. The extract displayed cytotoxicity towards B16F10 and MeWo skin melanoma cells, thereby showing a dose-dependent decrease in cell density. This was preceded by cell cycle arrest in B16F10 cells at the S phase and MeWo cell arrest at the early M phase with a significant increase in apoptosis in both cells. Furthermore, the extract displayed genotoxic potential at the tested concentrations (12.5–200 μg/mL). Overall, the results revealed that H. petiolare extract may have the potential to eradicate skin cancer.
1. Introduction
Cancer is one of the most dreadful diseases globally and it is described by abnormal cellular proliferation. It develops due to the cellular accumulation of various epigenetic and genetic events [1,2]. Some factors such as lifestyle, environment and nutrition have also been reported to play a crucial role in the pathogenesis of cancer. In a normal cell cycle, the growth and division of cells occur in a well-organized manner, but in cancerous cells, defective caspase-mediated cell death (apoptosis) leads to increased cell proliferation [3]. Skin cancer is among the most prevalent types of cancer globally and its incidence is increasing at an alarming rate. One report showed that there were more than two million new cases of skin cancer diagnosed in 2018 [4]. This number has been growing, likely due to the earlier detection of the disease, increased sun exposure, and longer lifespans. However, the treatment of skin cancer is commonly overwhelmed by the fact that most synthetic chemotherapeutic agents are frequently associated with a variety of toxic effects, mainly due to their non-selectivity in killing cancerous cells [5,6]. These limitations and costs of modern treatments for skin cancer pose many shortfalls. Many patients plagued with this disease do not seek proper treatment, resulting in increased morbidity and mortality.
Recently, several medicinal herbs have attracted numerous researchers’ attention for cancer treatment. A number of medicinal plants and their bioactive components have been revealed to possess anti-carcinogenic and anti-proliferative effects against cancerous skin cells with fewer side effects [7,8,9]. Hence, medicinal plants may serve as potential sources for developing new anticancer drugs.
Helichrysum is a genus consist of an estimated 600 species of flowering plants in the family of Asteraceae. Out of the 600 species, 244 species including Helichrysum petiolare are used for various traditional medicine treatments in South Africa. Some of the observed pharmacological activities include antioxidant, antimicrobial, anticancer, antidiabetic and anti-inflammatory effects [10,11]. Several parts of H. petiolare have been used traditionally for the treatment of various ailments such as cough, cold fever, infection, asthma, chest problems, diabetes, hypertension and wounds [12]. Some of the biological activity of H. petiolare previously reported include anti-tyrosinase, anti-inflammatory, antifungal and antioxidant effects [13].
Although H. petiolare is not used traditionally for the treatment of cancer, reports have shown that this plant has some chemical constituents that are responsible for its anti-cancer activity, such as flavonoids, polyphenolics and saponins [12,14]. Based on this reason, in this study, we examine the anti-proliferative activities of H. petiolare methanol extract against two cancerous skin cells: mouse melanoma cells (B16F10) and human melanoma cells (MeWo). In addition, an attempt is also made to examine the genotoxic potential of the extract towards Vero cells using the micronucleus assay.
2. Materials and Methods
2.1. Reagents
Skin melanoma cell lines (B16F10 and MeWo) were acquired from Highveld Biological, Johannesburg, South Africa, while the Vero cells were purchased from ATCC (Manassas, VA, USA). Roswell Park Memorial Institute (RPMI) 1640, Dulbecco’s phosphate-buffered saline (DPBS) with Ca2+ and Mg2+ and DPBS without Ca2+ and Mg2+, trypsin-EDTA and Dulbecco’s modified Eagle’s medium (DMEM) low-glucose cell culture medium were purchased from HyClone, Longa, UT, USA. Fetal bovine serum (FBS) was purchased from Biowest (Logan, UT, USA). BisBenzamide H 33342 trihydrochloride (Hoechst 33342), propidium iodide, griseofulvin and etoposide were purchased from Sigma-Aldrich (St. Louis, MO, USA). An annexin V-FITC kit was obtained from MACS Miltenyi Biotec, Cologne, Germany. NucRed was purchased from Molecular Probes®, Life Technologies—Thermo Fisher Scientific (Logan, UT, USA).
2.2. Plant Material and Extract Preparation
Fresh, mature leaves of H. petiolare were collected in December 2019 from the town of Alice, Amatole District, Eastern Cape, South Africa. The voucher specimen (Voucher no. HEL-1340) was authenticated by a botanist (Prof. Chris Cupido) and preserved in the Giffen Herbarium of the University of Fort Hare. For the extraction process, the milled plant (60 g) was mixed using methanol (1000 mL) at room temperature for 24 h. The resulting mixture was filtered and then concentrated to dryness by means of a rotary evaporator (RVO 004; Ignos, Prague, Czech Republic) to obtain a dried extract (14.43% dry extract).
2.3. Cell Culture Conditions
MeWo and B16F10 skin melanoma cells were separately maintained in 10-cm culture dishes in complete DMEM (low-glucose culture medium) supplemented with FBS (10%) and incubated at 37 °C in a humidified incubator with 5% CO2 [15].
2.4. Imaging and Analysis
The ImageXpress Micro XLS Widefield High-Content Analysis System (Molecular Devices®, San Jose, CA, USA) with the MetaXpress® High-Content Image Acquisition and Analysis Software (Molecular Devices®, San Jose, CA, USA) was used to acquire and analyze all images [15]. Nine image sites per well were acquired for all imaging experiments.
2.5. Cytotoxicity Assay and Cell Cycle Analysis
Briefly, B16F10 and MeWo skin melanoma cells were seeded separately in 96-well plates at densities of 3000 cells/well using 100 μL aliquots and left to attach overnight. For treatment, cells were treated by adding 100 μL of H. petiolare (0–100 µg/mL) and then incubated for 48 h at 37°C in a humidified 5% CO2 incubator. Melphalan was used as a positive control. The treatment medium was removed and replaced with a 50 µL staining solution (5 mL binding buffer containing 50 µL Annexin V-FITC reagent (Milteny Biotec Annexin V Kit: Cat no 130-092-052) and a 2 µL Hoechst 33342 solution (10 mg/mL in DMSO)) and then incubated for 15 min at 37 °C. Thereafter, 50 μL of propidium iodide (PI) (2 μg/mL in binding buffer) was added and then incubated for an additional 5 min, after which the images were acquired. The cell cycle analysis was then carried out using a cell cycle module.
2.6. Genotoxicity (Micronucleus Assay)
The micronucleus assay was performed as described previously [16]. Vero cells were seeded in a 96-well plate at a density of 3000 cells/well using 100 μL aliquots and left overnight to attach. For the treatment, an additional 100 μL of the plant extract at different concentrations (12.5–200 μg/mL) or positive controls, griseofulvin, (35 μg/mL) and etoposide (29 μg/mL), was added. The treated cells were incubated for 48 h at 37 °C in a humidified 5% CO2 incubator. The spent culture medium was removed by aspiration and cells were fixed using 4% formaldehyde for 15 min at room temperature, then 100 μL of NucRedTM Live 647 stain dye (50 μL NucRed in 10 mL of PBS, according to the manufacturer’s instructions) was added. Thereafter, cells were further incubated for an additional 15 min at room temperature prior to imaging.
2.7. Statistical Analysis
Three replicates of each test sample (methanol extract) were used. The data were statistically analyzed by Student’s t-test (two-tailed paired).
3. Results and Discussion
3.1. Cytotoxicity towards B16F10 Melanoma (Hoechst/Propidium Iodide)
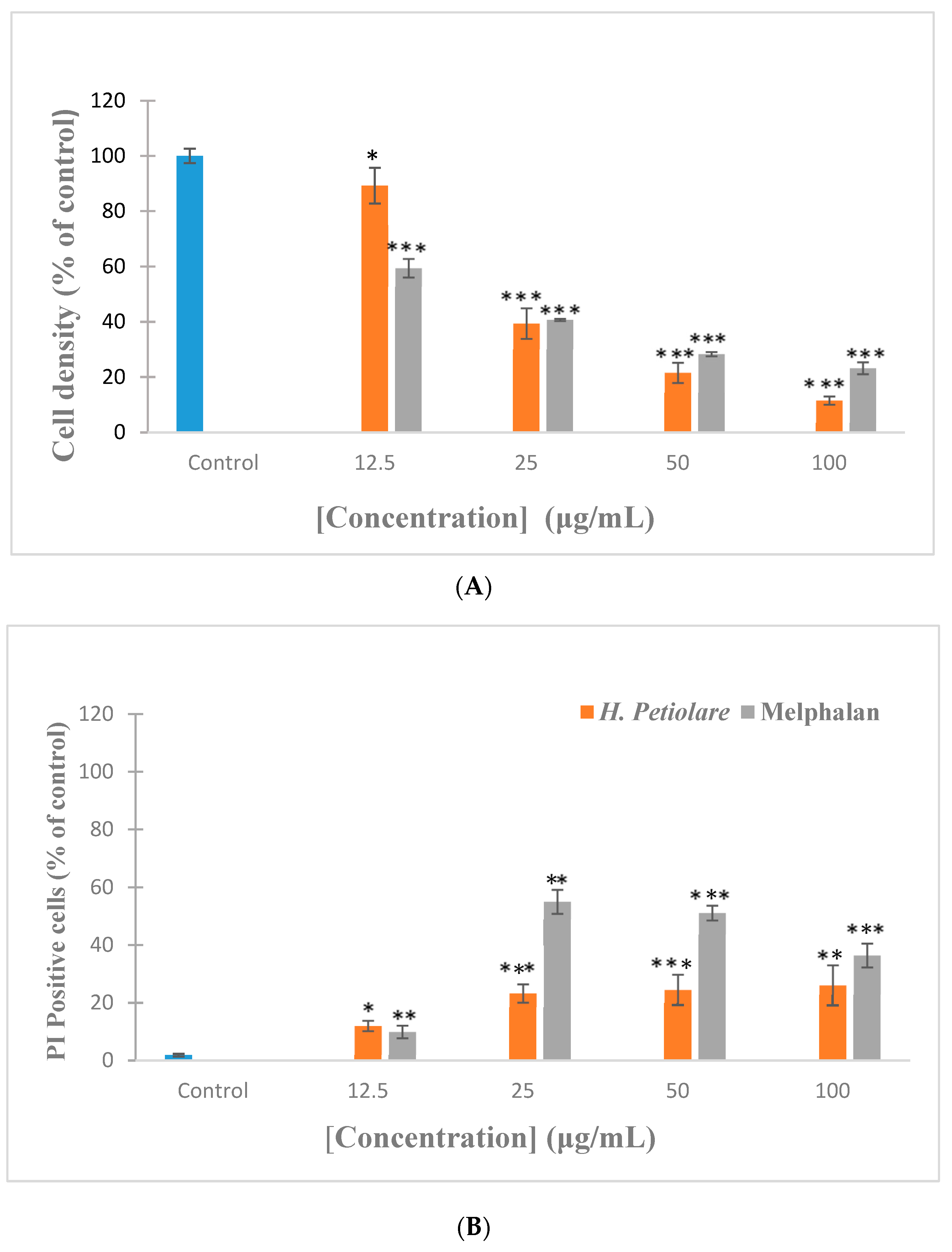
The dose-dependent anti-proliferative effect of H. petiolare extract and the positive control (Melphalan) against B16F10 cells is shown in Figure 1. The extract exhibited a strong significant (p < 0.001) decrease in cell density at the tested concentrations of 25 µg/mL and higher (50–100 μg/mL). This reduction was less marked compared to the trend seen by the control cells and the positive control, Melphalan. However, despite the sharp decline in cell density by the extract, cell death, as determined using propidium iodide (PI) staining, does not show a reciprocal response and cell death appears to remain relatively constant above 25 µg/mL. This indicates that cell death is not the only contributory mechanism responsible for the decline in cell density and suggests that the inhibition of cell proliferation is an additional mechanism. It is interesting to note that the cytotoxic effect of H. petiolare extract against B16F10 melanoma cells has never been reported in the literature. However, some studies have been conducted on the anticancer properties of Helichrysum genus. For example, Helichrysum plicatum, a prominent member of the Helicrysum genus, was found to be active against K562 and PC3 cancer cells [15]. Those results support the findings observed in this study, although the cancerous cells used in that study are quite different from the ones reported in the present study. Taken together with the cytotoxic results, it could be said that H. petiolare extract is therefore a promising candidate for the treatment of cancer.
3.2. Cytotoxicity towards MeWo Melanoma (Hoechst/PI)
A further assessment of cytotoxicity was also carried out in MeWo cells. The result (Figure 2) showed that treatment of MeWo cells with the extract caused a dose-dependent decrease in the cell density compared to the control cells at all the tested concentrations (12.5–100 μg/mL). In addition, cell death, as indicated by PI staining, was more prominent in MeWo cells with increasing concentrations (12.5–50 µg/mL) of the extract. On the other hand, the positive control, Mephalan, induced cell density and cell death better than the trend seen with the plant extract and the control cells. Indeed, the decline in cell density, as exhibited by the extract, appears to involve both cell death and the inhibition of cell proliferation. However, several constituents of the H. petiolare extract, as described by Maroyi [12], have been described in the literature for their cytotoxic activity on different cancer cell lines [16,17]. These plant constituents have been reported to affect different intracellular signaling pathways implicated in the initiation, promotion and progression of cancer [18,19]. Then, the observed cytotoxicity activity of the plant extract against MeWo melanoma cells might be attributed to the synergic effects of some of its chemical constituents that function on the cells, thereby leading to growth inhibition. The findings from this study suggest that the extract suppressed the growth of MeWo melanoma cells, therefore showing anticancer properties. However, it is imperative to perform a cell cycle analysis to assess population cell death and to examine whether the extract can induce cell cycle arrest.
3.3. Cell Cycle Analysis in B16F10 Melanoma (Annexin V/Hoechst)
Loss of control of the cell cycle plays a significant role in the growth of cancer. Thus, halting the cycle of cancer cells at any stage is an important strategy in the treatment of cancer [20]. In the present study, cell cycle analysis was determined to further confirm whether the H. petiolare extract mediated any alteration of a specific phase in cell cycle progression. As may be expected, the majority of cells were in the G0/G1 phase, a typical feature, as this represents the longest phase of the cell cycle. Significant changes in the G0/G1 and S phases of the cell cycle are apparent after treatment with the extract, suggesting an S phase arrest (Figure 3). In contrast to the extract, the positive control, Melphalan, induced a robust increase in G2 arrested cells, which is consistent with DNA crosslinking agents. To exclude apoptotic cells from the cell cycle analysis, Annexin V staining was used to identify apoptotic cells. Treatment with the extract caused a significant but small increase in apoptosis. However, it is known that cell cycle arrest is an important cause of the inhibition of cancer cell proliferation. For example, previous studies conducted by Liu et al. [21] revealed that Ophiorrhiza pumila inhibited liver cancer cell proliferation by inducing G2/M arrest. Another study conducted by Swanepoel et al. [22] also showed that Anemone nemorosa extract inhibited HeLa cervical cancer cell proliferation by inducing cell arrest at the early M phase. Thus, in this study, the extract demonstrated growth inhibitory effects in the B16F10 skin melanoma cells by inducing cell cycle arrest at the S phase. The cell cycle arrest at the S phase observed in this study is in support of other previous studies that reported plant extracts that induced an S phase arrest [23,24]. Studies have indicated that cell cycle arrest at the S phase is controlled by cyclin and cyclin-dependent kinase (CDKs) [25]. The CDKs are upregulated by cyclins and downregulated by cyclin-dependent kinase inhibitors (CDKI) such as p16INK4a and p21Cip1 [26]. However, it should be noted that gene expression analysis was not investigated in this present study. From this perspective, it is clear that the proliferation of B16F10 skin melanoma cells was halted by DNA arrest during the S phase at 50 μg/mL, therefore suggesting that H. petiolare may be a potential candidate in eradicating skin cancer.
3.4. Cell Cycle Analysis in MeWo Melanoma (Annexin V/Hoechst)
Significant changes in the G0/G1 phase of the cell cycle are apparent after treatment with the extract. The extract caused a predominant cell cycle arrest at the early M phase (Figure 4). In contrast to the extract, Melphalan induced a significant increase in both the S and G2 phases, but this still remains consistent for a DNA crosslinking agent. To exclude apoptotic cells from the cell cycle analysis, Annexin V staining was used to identify apoptotic cells. Treatment with the extract caused a significant but small increase in apoptosis. One report showed that mitotic arrest is predominantly due to the disruption of the formation of the mitotic spindle, resulting in mitotic catastrophe [27]. The observed cell cycle arrest at the early M phase, as exhibited by the extract in the MeWo melanoma cells, therefore suggests that the extract may act by triggering the disruption of the formation of the mitotic spindle, thereby causing mitotic catastrophe [27]. However, several researchers have indicated that phytochemical components play an important role in regulating the genes that control the cell cycle, proliferation and apoptosis pathway in cancerous cells [28,29,30]. Therefore, this effect of H. petiolare extract on cell cycle progression may be attributed to its secondary metabolites, which were previously reported [12,14].
3.5. Genotoxicity (Micronucleus Assay)
Micronuclei refer to small nuclei that are formed whenever a chromosome or a fragment of a chromosome is not incorporated into one of the daughter nuclei during cell division [31]. This is commonly a sign of genotoxic events and chromosomal instability (DNA strand breaks and chromosome loss or damage). In this study, a significant cytotoxicity towards Vero cells at the highest three concentrations tested (50–200 µg/mL) was seen following exposure compared to the trend seen in control cells (Figure 5). In addition, a significant increase in the formation of micronuclei cells was also observed with increasing concentrations (50–200 µg/mL) of the extract. Furthermore, the extract caused a dose-dependent decrease in the ratio of 2n to 4n and a dose-dependent increase in the nuclear size of the cells at the highest concentrations. On the other hand, etoposide, used as a positive control, exhibited a better increase in nuclear size compared to the trend seen in extract and control cells. It is clear that all the parameters observed in this study support the meaningful genotoxic effect of the plant extract. This is the first study to investigate the genotoxic potential of H. petiolare using a micronucleus assay. However, the genotoxic effect of plants from the Helichrysum genus has been reported. The study conducted by Eroglu et al. [32] revealed that methanol extracts of H. pamphylicum and H. sanguineum induced the formation of micronuclei in human lymphocyte cultures, thereby suggesting their genotoxic potential. This previous report is in agreement with the findings of this present study. It can be deduced from this study that H. petiolare extract exhibits genotoxic potential, therefore supporting its ability as an anticancer agent.
4. Conclusions
In conclusion, the treatment of melanoma cells with this plant extract is characterized by a strong decline in cell density; however, cell death does not appear to fully explain the decline in cell density and suggests that proliferation is strongly impeded by the extract. To make a more definitive conclusion, additional experiments to confirm the mechanism of cell death is recommended. Data from the cell cycle analysis, however, provide some evidence that the mechanism involves aberrant cell division. In B16F10 cells, the extract caused cell arrest in the S phase, while in MeWo cells, extract treatment was characterized by arrest in the early M phase. This dissimilarity between the B16F10 and MeWo cells probably represents species differences. Data derived from all the parameters of the micronucleus assay in Vero cells support the meaningful genotoxic effect of the H. petiolare extract. Furthermore, a dose-dependent decrease in the ratio of 2n to 4n observed in the micronucleus assay also supports a mechanism that involves aberrant cell division. Overall, the results of this study show that methanol extract from H. petiolare may have the ability to eradicate skin cancer.
Author Contributions
Investigation, I.J.S., W.O.-M.; supervision, I.J.S., W.O.-M.; writing—original draft, I.J.S.; writing—review and editing, I.J.S., W.O.-M., I.J.S. All authors have read and agreed to the published version of the manuscript.
Funding
This work was funded by the National Research Foundation (NRF) (grant no: 105161).
Acknowledgments
The authors are grateful to the University of Mpumalanga and the National Research Foundation, South Africa, for their financial support.
Conflicts of Interest
The authors declare that there are no conflict of interest regarding the publication of this paper.
References
- Giri, B.; Gomes, A.; Debnath, A.; Saha, A.; Biswas, A.K.; Dasgupta, S.C. Antiproliferative, cytotoxic and apoptogenic activity of Indian toad (Bufo melanostictus, Schneider) skin extract on U937 and K562 cells. Toxicon 2006, 48, 388–400. [Google Scholar] [CrossRef] [PubMed]
- Mbaveng, A.T.; Kuete, V.; Mapunya, B.M.; Beng, V.P.; Nkengfack, A.E.; Meyer, J.J.M.; Lall, N. Evaluation of four cameroonian medicinal plants for anticancer, antigonorrheal and antireverse transcriptase activities. Environ.Toxicol. Pharmacol. 2011, 32, 162–167. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zimmerman, M.A.; Huang, Q.; Li, F.; Liu, X.; Li, C.-Y. Cell death–stimulated cell proliferation: A tissue regeneration mechanism usurped by tumors during radiotherapy. Semin. Radiat. Oncol. 2013, 23, 288–295. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bray, F.; Ferlay, J.; Soerjomataram, I.; Siegel, R.L.; Torre, L.A.; Jemal, A. Global Cancer Statistics 2018: GLOBOCAN estimates of incidence and mortality worldwide for 36 cancers in 185 countries. CA A Cancer J. Clin. 2018, 68, 394–424. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Soura, E.; Chasapi, V.; Stratigos, A.J. Pharmacologic treatment options for advanced epithelial skin cancer. Expert Opin.Pharmacotherl. 2015, 16, 1479–1493. [Google Scholar] [CrossRef]
- Rudnick, E.W.; Thareja, S.; Cherpelis, B.B. Oral therapy for non melanoma skin cancer in patients with advanced disease and large tumor burden: A review of the literature with focus on a new generation of targeted therapies. Int. J. Dermatol. 2016, 55, 249–258. [Google Scholar] [CrossRef]
- Cruz, J.F.D.; Kim, Y.S.; Lumbera, W.M.L.; Hwang, S.G. Viscum Album Var Hot Water Extract Mediates Anti-cancer Effects through G1 Phase Cell Cycle Arrest in SK-Hep1 Human Hepatocarcinoma cells. Asian Pac. J. Cancer Prev. 2015, 16, 6417–6421. [Google Scholar] [CrossRef] [Green Version]
- Esmaeilbeig, M.; Kouhpayeh, S.A.; Amirghofran, Z. An Investigation of the Growth Inhibitory Capacity of Several Medicinal Plants From Iran on Tumor Cell Lines. Iran. J. Cancer Prev. 2015, 8, 4032. [Google Scholar] [CrossRef] [Green Version]
- Vijaybabu, K.; Punnagai, K. In-Vitro Anti-Proliferative Effects of Ethanolic Extract of Vanilla Planifolia Leaf Extract Against A431 Human Epidermoid Carcinoma Cells. Biomed. Pharmacol. J. 2019, 12, 1141–1146. [Google Scholar] [CrossRef]
- Mancini, E.; De Martino, L.; Marandino, A.; Scognamiglio, M.R.; De Feo, V. Chemical Composition and Possible in Vitro Phytotoxic Activity of Helichrsyum italicum (Roth) Don ssp. italicum. Molecules 2011, 16, 7725–7735. [Google Scholar] [CrossRef] [Green Version]
- Akaberia, M.; Sahebkar, A.; Azizid, N.; Emamiae, S.A. Everlasting flowers: Phytochemistry and pharmacology of the genus Helichrysum. Ind. Crop. Prod. 2019, 138, 111471. [Google Scholar] [CrossRef]
- Maroyi, A. Helicrysum Petiolare Hilland and B.L. Burtt: A review of its medicinal uses, phytochemistry, and biological activities. Asian J. Pharm. Clin. Res. 2019, 12, 69–71. [Google Scholar]
- Lourens, A.C.U.; Reddy, D.; Baser, K.H.C.; Viljoen, A.M.; Van Vuuren, S. In vitro biological activity and essential oil composition of four indigenous South African Helichrysum species. J. Ethnopharmacol. 2004, 95, 253–258. [Google Scholar] [CrossRef] [PubMed]
- Otang-mbeng, W.; Sagbo, I.J. Gas Chromatography—Mass Spectrometry Analysis of the Volatile Compounds from the Ethanol Extracts of Bulbine asphodeloides and Helichrysum petiolare. Pharmacogn. Rev. 2019, 11, 219–223. [Google Scholar] [CrossRef]
- Bigovica, D.; Savikina, K.; Jankovica, T.; Menkovica, N.; Zdunica, G.; Tatjana, S.; Djuric, Z. Antiradical and Cytotoxic Activity of Different Helichrysum plicatum Flower Extracts. Nat. Prod. Commun. 2011, 6, 819–822. [Google Scholar]
- Andrade, L.N.; Lima, T.C.; Amaral, R.G.; Pessoa, C.D.; Filho, M.O.M.; Soares, B.M.; Nascimento, L.G.; Carvalho, A.A.; De Sousa, D.P. Evaluation of the Cytotoxicity of Structurally Correlatedp-Menthane Derivatives. Molecules 2015, 20, 13264–13280. [Google Scholar] [CrossRef] [Green Version]
- Afoulous, S.; Ferhout, H.; Raoelison, E.G.; Valentin, A.; Moukarzel, B.; Couderc, F.; Bouajila, J. Helichrysum gymnocephalum Essential Oil: Chemical Composition and Cytotoxic, Antimalarial and Antioxidant Activities, Attribution of the Activity Origin by Correlations. Molecules 2011, 16, 8273–8291. [Google Scholar] [CrossRef]
- Neergheen, V.S.; Bahorun, T.; Taylor, E.W.; Jen, L.S.; Aruoma, O.I. Targeting specific cell signaling transduction pathways by dietary and medicinal phytochemicals in cancer chemoprevention. Toxicology 2010, 278, 229–241. [Google Scholar] [CrossRef]
- Amin, A.R.; Kucuk, O.; Khuri, F.R.; Shin, D.M. Perspectives for cancer prevention with natural compounds. J. Clin. Oncol. 2009, 27, 2712–2725. [Google Scholar] [CrossRef] [Green Version]
- Alimbetov, D.; Askarova, S.; Umbayev, B.; Davis, T.; Kipling, D. Pharmacological Targeting of Cell Cycle, Apoptotic and Cell Adhesion Signaling Pathways Implicated in Chemoresistance of Cancer Cells. Int. J. Mol. Sci. 2018, 19, 1690. [Google Scholar] [CrossRef] [Green Version]
- Liu, H.; Liao, W.; Fan, L.; Zheng, Z.; Liu, D.; Zhang, Q.; Yang, A.; Liu, F. Ethanol extract of Ophiorrhiza pumila suppresses liver cancer cell proliferation and migration. . Chin. Med. 2020, 15, 1–11. [Google Scholar]
- Swanepoel, B.; Venables, L.; Olaru, O.T.; Nitulescu, G.M.; van de Venter, M. In Vitro Anti-proliferative Activity and Mechanism of Action of Anemone nemorosa. Int. J. Mol. Sci. 2019, 20, 1217. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pereira, J.M.; Lopes-rodrigues, V.; Xavier, C.P.R.; Lima, M.J.; Lima, R.T.; Ferreira, I.C.F.R.; Vasconcelos, M.H. An Aqueous Extract of Tuberaria lignosa Inhibits Cell Growth, Alters the Cell Cycle Profile and Induces Apoptosis of NCI-H460 Tumor Cells. Molecules 2016, 21, 595. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Khazaei, S.; Hamid, R.A.; Ramachandran, V.; Esa, N.M.; Pandurangan, A.K.; Danazadeh, F.; Ismail, P. Cytotoxicity and Proapoptotic Effects of Allium atroviolaceum Flower Extract by Modulating Cell Cycle Arrest and Caspase-Dependent and p53-Independent Pathway in Breast Cancer Cell Lines. Evid. Complement. Altern. Med. 2017, 2017, 1–16. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Singh, R.P.; Agarwal, R. Natural flavonoids targeting deregulated cell cycle progression in cancer cells. Curr. Drug Targets 2006, 7, 345–354. [Google Scholar] [CrossRef]
- Sherr, C.J.; Roberts, J.M. CDK inhibitors: Positive and negative regulators of G1-phase progression. Genes Dev. 1999, 13, 1501–1512. [Google Scholar] [CrossRef] [Green Version]
- Busino, L.; Chiesa, M.; Draetta, G.F.; Donzelli, M. Cdc25A phosphatase: Combinatorial phosphorylation, ubiquitylation and proteolysis. Oncogene 2004, 23, 2050–2056. [Google Scholar] [CrossRef] [Green Version]
- Konoshima, T.; Konishi, T.; Takasaki, M.; Yamazoe, K.; Tokuda, H. Anti-tumor-promoting activity of the diterpene from Excoecaria agallocha. Biol. Pharm. Bull. 2001, 24, 1440–1442. [Google Scholar] [CrossRef] [Green Version]
- Zou, J.H.; Dai, J.; Chen, X.; Yuan, J.Q. Pentacyclic triterpenoids from leaves of Excoecaria agallocha. Chem. Pharm. Bull. 2006, 54, 920–921. [Google Scholar] [CrossRef] [Green Version]
- Reddy, K.P.R.; Durairaj, P.; Thiruvanavukkarasu, P.; Hari, R. Effect of ethanolic extract of Excoecaria agallocha leaves on the cytotoxic activity and cell cycle arrest of human breast cancer cell lines—MCF-7. Pharmacogn. Mag. 2019, 15, 346–351. [Google Scholar]
- Imreh, G.; Norberg, H.V.; Imreh, S.; Zhivotovsky, B. Chromosomal breaks during mitotic catastrophe trigger H2AX–ATM–p53-mediated apoptosis. J. Cell Sci. 2011, 124, 2951–2963. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Eroğlu1, E.H.; Hamzaoğlu1, E.; Aksoy, A.; Budak, U.; Özkul, Y. In Vitro Genotoxic Effects of Four Helichrysum Species in Human Lymphocytes Cultures. Biol. Res. 2010, 43, 177–182. [Google Scholar] [CrossRef]
Figure 1.
Cytotoxicity of H. petiolare methanol extract towards mouse melanoma cells (B16F10). (A) Cell density represents the number of nuclei stained with Hoechst and is expressed as a percentage of the untreated control. (B) Cell death is quantified as the percentage cells which stain positive for propidium iodide (PI). Melphalan was included as a positive control. Data are expressed as the mean ± SD, n = 3. * p < 0.05, ** p < 0.01 and *** p < 0.001 compared to control.
Figure 1.
Cytotoxicity of H. petiolare methanol extract towards mouse melanoma cells (B16F10). (A) Cell density represents the number of nuclei stained with Hoechst and is expressed as a percentage of the untreated control. (B) Cell death is quantified as the percentage cells which stain positive for propidium iodide (PI). Melphalan was included as a positive control. Data are expressed as the mean ± SD, n = 3. * p < 0.05, ** p < 0.01 and *** p < 0.001 compared to control.

Figure 2.
Cytotoxicity of H. petiolare methanol extract towards human melanoma cells (MeWo). (A) Cell density represents the number of nuclei stained with Hoechst and expressed as a percentage of the untreated control. (B) Cell death is quantified as the percentage cells which stain positive for PI. Melphalan included as a positive control. Data are expressed as the mean ± SD, n = 3. ** p < 0.01 and *** p < 0.001 compared to control.
Figure 2.
Cytotoxicity of H. petiolare methanol extract towards human melanoma cells (MeWo). (A) Cell density represents the number of nuclei stained with Hoechst and expressed as a percentage of the untreated control. (B) Cell death is quantified as the percentage cells which stain positive for PI. Melphalan included as a positive control. Data are expressed as the mean ± SD, n = 3. ** p < 0.01 and *** p < 0.001 compared to control.

Figure 3.
Cell cycle analysis in B16F10 cells after exposure of H. petiolare extract (50 µg/mL) and Melphalan (50 µg/mL). Cell cycle analysis was determined by the NucRed TM Live 647 stain dye. The results are presented as the percentage of cells detected in each phase. Data are expressed as the mean ± SD, n = 3. * p < 0.05, ** p < 0.01 and *** p < 0.001 compared to control.
Figure 3.
Cell cycle analysis in B16F10 cells after exposure of H. petiolare extract (50 µg/mL) and Melphalan (50 µg/mL). Cell cycle analysis was determined by the NucRed TM Live 647 stain dye. The results are presented as the percentage of cells detected in each phase. Data are expressed as the mean ± SD, n = 3. * p < 0.05, ** p < 0.01 and *** p < 0.001 compared to control.

Figure 4.
Cell cycle analysis in MeWo cells after exposure to H. petiolare extract (50 µg/mL) and Melphalan (50 µg/mL). NucRed TM Live 647 stain dye. The results are presented as the percentage of cells detected in each phase. Data are expressed as the mean ± SD, n = 3. * p < 0.05, ** p < 0.01 and *** p < 0.001 compared to control.
Figure 4.
Cell cycle analysis in MeWo cells after exposure to H. petiolare extract (50 µg/mL) and Melphalan (50 µg/mL). NucRed TM Live 647 stain dye. The results are presented as the percentage of cells detected in each phase. Data are expressed as the mean ± SD, n = 3. * p < 0.05, ** p < 0.01 and *** p < 0.001 compared to control.

Figure 5.
Micronucleus formation and nuclear parameters of Vero cells treated with different concentrations of H. petiolare methanol extract. Cytotoxicity is measured as decrease in the number of cells relative to the untreated control (also referred to as % Relative Cell Count). Data are expressed as the mean ± SD, n = 3. * p < 0.05, ** p < 0.01 and *** p < 0.001 compared to control. Griseofulvin (Grs). (A) Cytotoxicity; (B) micronucleus; (C) cell ploidy; (D) nuclear size; (E) micrographs (10× magnification) illustrating the nuclear size after treatment compared to the control.
Figure 5.
Micronucleus formation and nuclear parameters of Vero cells treated with different concentrations of H. petiolare methanol extract. Cytotoxicity is measured as decrease in the number of cells relative to the untreated control (also referred to as % Relative Cell Count). Data are expressed as the mean ± SD, n = 3. * p < 0.05, ** p < 0.01 and *** p < 0.001 compared to control. Griseofulvin (Grs). (A) Cytotoxicity; (B) micronucleus; (C) cell ploidy; (D) nuclear size; (E) micrographs (10× magnification) illustrating the nuclear size after treatment compared to the control.

Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Sagbo, I.J.; Otang-Mbeng, W. Anti-Proliferative and Genotoxic Activities of the Helichrysum petiolare Hilliard & B.L. Burtt. Sci. Pharm. 2020, 88, 49. https://0-doi-org.brum.beds.ac.uk/10.3390/scipharm88040049
AMA Style
Sagbo IJ, Otang-Mbeng W. Anti-Proliferative and Genotoxic Activities of the Helichrysum petiolare Hilliard & B.L. Burtt. Scientia Pharmaceutica. 2020; 88(4):49. https://0-doi-org.brum.beds.ac.uk/10.3390/scipharm88040049
Chicago/Turabian StyleSagbo, Idowu Jonas, and Wilfred Otang-Mbeng. 2020. "Anti-Proliferative and Genotoxic Activities of the Helichrysum petiolare Hilliard & B.L. Burtt" Scientia Pharmaceutica 88, no. 4: 49. https://0-doi-org.brum.beds.ac.uk/10.3390/scipharm88040049