
Muscle- and Mode-Specific Responses of the Forearm Flexors to Fatiguing, Concentric Muscle Actions


Abstract
:1. Introduction
2. Results
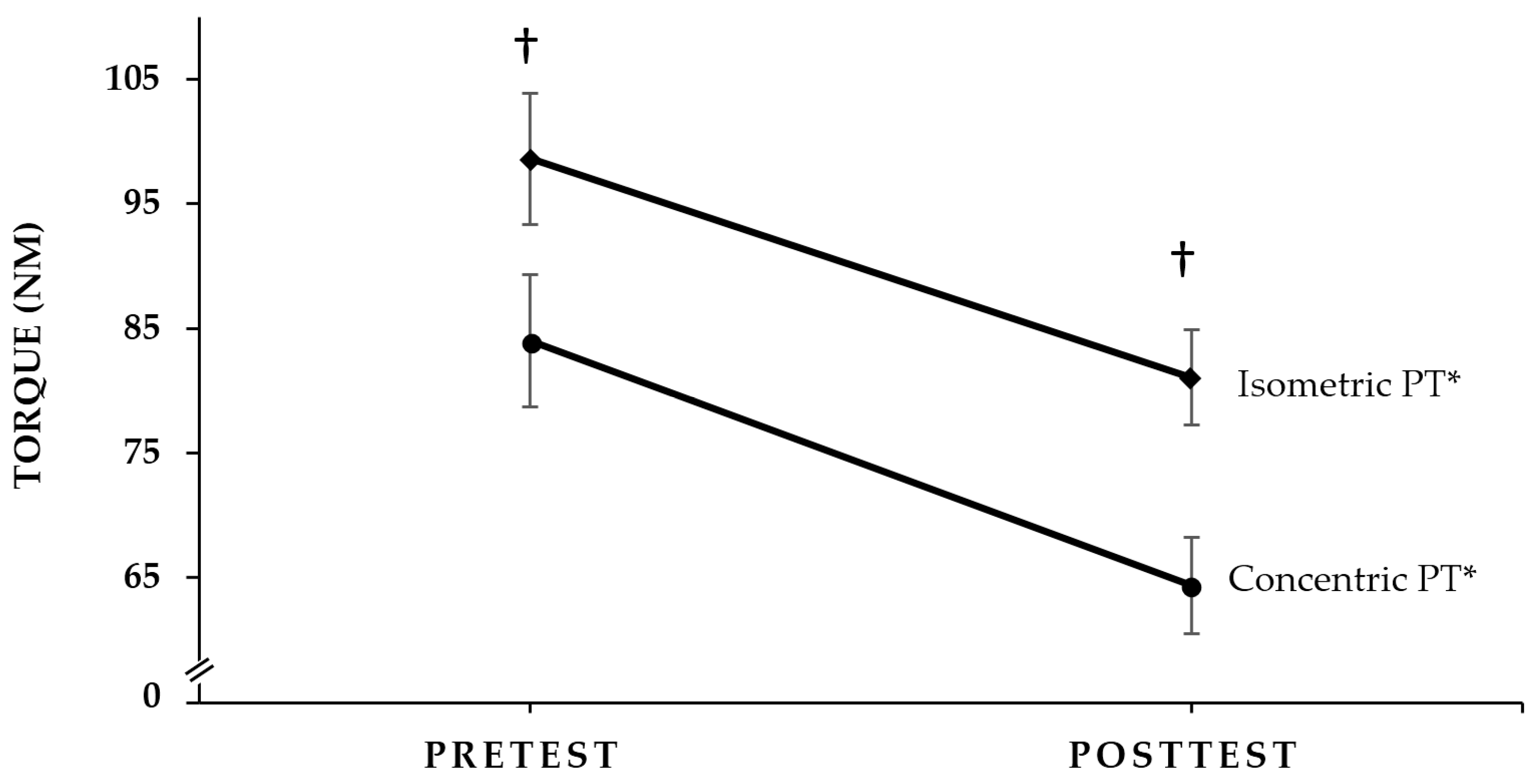
2.1. Concentric PT and Isometric PT Responses
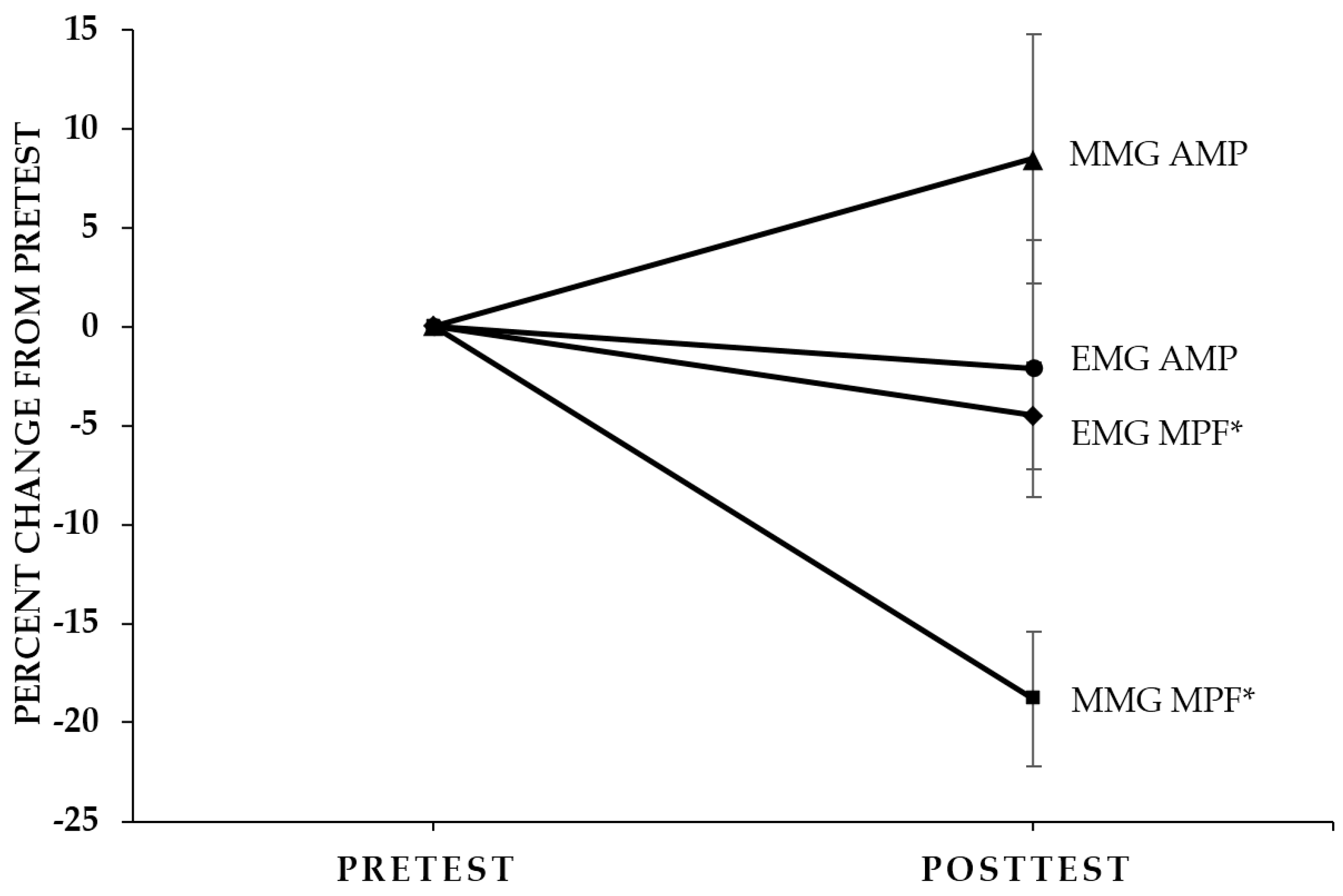
2.2. Concentric PT and Isometric PT Neuromuscular Responses
3. Discussion
3.1. The Effects of the Submaximal, Dynamic Workbout
3.2. Torque
3.3. Pretest Versus Posttest EMG and MMG Responses
4. Materials and Methods
4.1. Subjects
4.2. Procedures
4.3. Statistical Analyses
5. Conclusions
Author Contributions
Conflicts of Interest
References
- Basmajian, J.V. Muscles Alive, Their Functions Revealed by Electromyography; Williams & Wilkins: Philadelphia, PA, USA, 1978. [Google Scholar]
- De Luca, C.J. Physiology and mathematics of myoelectric signals. IEEE Trans. Biomed. Eng. 1979, 26, 313–325. [Google Scholar] [CrossRef] [PubMed]
- Lindstrom, L.; Magnusson, R.; Petersen, I. Muscular fatigue and action potential conduction velocity changes studied with frequency analysis of EMG signals. Electromyography 1970, 10, 341–356. [Google Scholar] [PubMed]
- Mills, K.R. Power spectral analysis of electromyogram and compound muscle action potential during muscle fatigue and recovery. J. Physiol. 1982, 326, 401–409. [Google Scholar] [CrossRef] [PubMed]
- Gordon, G.; Holbourn, A.H.S. The sounds from single motor units in a contracting muscle. J. Physiol. Lond. 1948, 107, 456–464. [Google Scholar] [CrossRef] [PubMed]
- Orizio, C. Muscle sound—Bases for the introduction of a mechanomyographic signal in muscle studies. Crit. Rev. Biomed. Eng. 1993, 21, 201–243. [Google Scholar] [PubMed]
- Orizio, C.; Perini, R.; Veicsteinas, A. Muscular sound and force relationship during isometric contraction in man. Eur. J. Appl. Physiol. Occup. Physiol. 1989, 58, 528–533. [Google Scholar] [CrossRef] [PubMed]
- Beck, T.W.; Housh, T.J.; Johnson, G.O.; Weir, J.P.; Cramer, J.T.; Coburn, J.W.; Malek, M.H. Mechanomyographic amplitude and mean power frequency versus torque relationships during isokinetic and isometric muscle actions of the biceps brachii. J. Electromyogr. Kinesiol. 2004, 14, 555–564. [Google Scholar] [CrossRef] [PubMed]
- Farina, D.; Merletti, R.; Enoka, R.M. The extraction of neural strategies from the surface EMG. J. Appl. Physiol. 2004, 96, 1486–1495. [Google Scholar] [CrossRef] [PubMed]
- Farina, D.; Merletti, R.; Enoka, R.M. The extraction of neural strategies from the surface EMG: An update. J. Appl. Physiol. 2014, 96, 1486–1495. [Google Scholar] [CrossRef] [PubMed]
- Keenan, K.G.; Valero-Cuevas, F.J. Epoch length to accurately estimate the amplitude of interference EMG is likely the result of unavoidable amplitude cancellation. Biomed. Signal Process. Control 2008, 3, 154–162. [Google Scholar] [CrossRef] [PubMed]
- Beck, T.W.; Housh, T.J.; Cramer, J.T.; Weir, J.P.; Johnson, G.O.; Coburn, J.W.; Malek, M.H.; Mielke, M. Mechanomyographic amplitude and frequency responses during dynamic muscle actions: A comprehensive review. Biomed. Eng. Online 2005, 4. [Google Scholar] [CrossRef] [PubMed]
- Stokes, M.J.; Dalton, P.A. Acoustic myography for investigating human skeletal muscle fatigue. J. Appl. Physiol. 1991, 71, 1422–1426. [Google Scholar] [PubMed]
- Enoka, R.M.; Stuart, D.G. Neurobiology of muscle fatigue. J. Appl. Physiol. 1992, 72, 1631–1648. [Google Scholar] [PubMed]
- Hunter, S.K. Sex Differences and Mechanisms of Task-Specific Muscle Fatigue. Exerc. Sport Sci. Rev. 2009, 37, 113–122. [Google Scholar] [CrossRef] [PubMed]
- Camic, C.L.; Housh, T.J.; Zuniga, J.M.; Hendrix, C.R.; Bergstrom, H.C.; Traylor, D.A.; Schmidt, R.J.; Johnson, G.O. Electromyographic and mechanomyographic responses across repeated maximal isometric and concentric muscle actions of the leg extensors. J. Electromyogr. Kinesiol. 2013, 23, 342–348. [Google Scholar] [CrossRef] [PubMed]
- Yeung, S.S.; Au, A.L.; Chow, C.C. Effects of fatigue on the temporal neuromuscular control of vastus medialis muscle in humans. Eur. J. Appl. Physiol. Occup. Physiol. 1999, 80, 379–385. [Google Scholar] [CrossRef] [PubMed]
- Babault, N.; Desbrosses, K.; Fabre, M.-S.; Michaut, A.; Pousson, M. Neuromuscular fatigue development during maximal concentric and isometric knee extensions. J. Appl. Physiol. 2006, 100, 780–785. [Google Scholar] [CrossRef] [PubMed]
- Crenshaw, A.G.; Karlsson, S.; Styf, J.; Backlund, T.; Friden, J. Knee extension torque and intramuscular pressure of the vastus lateralis muscle during eccentric and concentric activities. Eur. J. Appl. Physiol. Occup. Physiol. 1995, 70, 13–19. [Google Scholar] [CrossRef] [PubMed]
- Seghers, J.; Spaepen, A. Muscle fatigue of the elbow flexor muscles during two intermittent exercise protocols with equal mean muscle loading. Clin. Biomech. 2004, 19, 24–30. [Google Scholar] [CrossRef]
- Mathiassen, S.E. The influence of exercise/rest schedule on the physiology and psychophysiological response to isometric shoulder-neck exercise. Eur. J. Appl. Physiol. Occup. Physiol. 1993, 67, 528–539. [Google Scholar] [CrossRef] [PubMed]
- Perry-Rana, S.R.; Housh, T.J.; Johnson, G.O.; Bull, A.J.; Berning, J.M.; Cramer, J.T. MMG and EMG responses during fatiguing isokinetic muscle contractions at different velocities. Muscle Nerve 2002, 26, 367–373. [Google Scholar] [CrossRef] [PubMed]
- Camic, C.L.; Housh, T.J.; Zuniga, J.M.; Bergstrom, H.C.; Schmidt, R.J.; Johnson, G.O. Mechanomyographic and electromyographic responses during fatiguing eccentric muscle actions of the leg extensors. J. Appl. Biomech. 2014, 30, 255–261. [Google Scholar] [CrossRef] [PubMed]
- Perry-Rana, S.R.; Housh, T.J.; Johnson, G.O.; Bull, A.J.; Cramer, J.T. MMG and EMG responses during 25 maximal, eccentric, isokinetic muscle actions. Med. Sci. Sports Exerc. 2003, 35, 2048–2054. [Google Scholar] [CrossRef] [PubMed]
- Tesch, P.A.; Dudley, G.A.; Duvoisin, M.R.; Hather, B.M.; Harris, R.T. Force and EMG signal patterns during repeated bouts of concentric or eccentric muscle actions. Acta Physiol. Scand. 1990, 138, 263–271. [Google Scholar] [CrossRef] [PubMed]
- Gray, J.C.; Chandler, J.M. Percent Decline in Peak Torque Production during Repeated Concentric and Eccentric Contractions of the Quadriceps Femoris Muscle. J. Orthop. Sports Phys. Ther. 1989, 10, 309–314. [Google Scholar] [CrossRef] [PubMed]
- Hill, E.C.; Housh, T.J.; Camic, C.L.; Smith, C.M.; Cochrane, K.C.; Jenkins, N.D.; Cramer, J.T.; Schmidt, R.J.; Johnson, G.O. The Effects of Velocity on Electromyographic, Mechanomyographic, and Torque Responses to Repeated Eccentric Muscle Actions. J. Strength Cond. Res. 2015, 30, 1743–1751. [Google Scholar] [CrossRef] [PubMed]
- Hunter, S.K. Sex differences in human fatigability: Mechanisms and insight to physiological responses. Acta Physiol. Oxf. 2014, 210, 768–789. [Google Scholar] [CrossRef] [PubMed]
- Barr, A.E.; Barbe, M.F. Pathophysiological Tissue Changes Associated With Repetitive Movement: A Review of the Evidence. Phys. Ther. 2002, 82, 173–187. [Google Scholar] [PubMed]
- Sjøgaard, G.; Søgaard, K. Muscle Injury in Repetitive Motion Disorders. Clin. Orthop. Relat. Res. 1998, 351, 21–31. [Google Scholar] [PubMed]
- Järvinen, T.A.H.; Kannus, P.; Maffulli, N.; Khan, K.M. Achilles Tendon Disorders: Etiology and Epidemiology. Foot Ankle Clin. 2005, 10, 255–266. [Google Scholar] [CrossRef] [PubMed]
- Rohmert, W. Problems in determining rest allowances Part 1: Use of modern methods to evaluate stress and strain in static muscular work. Appl. Ergon. 1973, 4, 91–95. [Google Scholar] [CrossRef]
- Noakes, T.D. Implications of exercise testing for prediction of athletic performance: A contemporary perspective. Med. Sci. Sports Exerc. 1988, 20, 319–330. [Google Scholar] [CrossRef] [PubMed]
- Camic, C.L. An Assessment of the Motor Control Strategies and Effects of Fatigue Specific to Isometric, Concentric, and Eccentric Muscle Actions. Available online: http://digitalcommons.unl.edu/dissertations/AAI3449431/ (accessed on 28 September 2016).
- Ettema, G.J.; Huijing, P.A. Skeletal muscle stiffness in static and dynamic contractions. J. Biomech. 1994, 27, 1361–1368. [Google Scholar] [CrossRef]
- Edwards, R.H. Human muscle function and fatigue. Ciba Found. Symp. 1981, 82, 1–18. [Google Scholar] [PubMed]
- Maclaren, D.P.; Gibson, H.; Parry-Billings, M.; Edwards, R.H. A review of metabolic and physiological factors in fatigue. Exerc. Sport Sci. Rev. 1989, 17, 29–66. [Google Scholar] [CrossRef] [PubMed]
- Fortune, E.; Lowery, M.M. Effect of Extracellular Potassium Accumulation on Muscle Fiber Conduction Velocity: A Simulation Study. Ann. Biomed. Eng. 2009, 37, 2105–2117. [Google Scholar] [CrossRef] [PubMed]
- Westerblad, H.; Allen, D.G.; Lannergren, J. Muscle fatigue: Lactic acid or inorganic phosphate the major cause? News Physiol. Sci. 2002, 17, 17–21. [Google Scholar] [PubMed]
- Malek, M.H.; Housh, T.J.; Crouch, L.D.; Johnson, G.O.; Hendrix, C.R.; Beck, T.W.; Mielke, M.; Schmidt, R.J.; Housh, D.J. Plasma ammonia concentrations and the slow component of oxygen uptake kinetics during cycle ergometry. J. Strength Cond. Res. 2008, 22, 2018–2026. [Google Scholar] [CrossRef] [PubMed]
- Gladden, L.B. Lactate metabolism: A new paradigm for the third millennium. J. Physiol. 2004, 558, 5–30. [Google Scholar] [CrossRef] [PubMed]
- Robergs, R.A.; Ghiasvand, F.; Parker, D. Biochemistry of exercise-induced metabolic acidosis. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2004, 287, R502–R516. [Google Scholar] [CrossRef] [PubMed]
- Marsden, C.D.; Meadows, J.C.; Merton, P.A. “Muscular wisdom” that minimizes fatigue during prolonged effort in man: Peak rates of motoneuron discharge and slowing of discharge during fatigue. Adv. Neurol. 1983, 39, 169–211. [Google Scholar] [PubMed]
- Garland, S.J.; Gossen, E.R. The muscular wisdom hypothesis in human muscle fatigue. Exerc. Sport Sci. Rev. 2002, 30, 45–49. [Google Scholar] [CrossRef] [PubMed]
- Fuglevand, A.J. Neural Aspects of Fatigue. Neuroscientist 1996, 2, 203–206. [Google Scholar] [CrossRef]
- Fuglevand, A.J.; Keen, D.A. Re-evaluation of muscle wisdom in the human adductor pollicis using physiological rates of stimulation. J. Physiol. 2003, 549, 865–875. [Google Scholar] [CrossRef] [PubMed]
- Fuglevand, A.J.; Zackowski, K.M.; Huey, K.A.; Enoka, R.M. Impairment of neuromuscular propagation during human fatiguing contractions at submaximal forces. J. Physiol. 1993, 460, 549–572. [Google Scholar] [CrossRef] [PubMed]
- De Luca, C.J.; Lefever, R.S.; McCue, M.P.; Xenakis, A.P. Control scheme governing concurrently active human motor units during voluntary contractions. J. Physiol. (Lond.) 1982, 329, 129–142. [Google Scholar] [CrossRef]
- De Luca, C.J.; Erim, Z. Common drive of motor units in regulation of muscle force. Trends Neurosci. 1994, 17, 299–305. [Google Scholar] [CrossRef]
- Kline, J.C.; de Luca, C.J. Synchronization of motor unit firings: An epiphenomenon of firing rate characteristics not common inputs. J. Neurophysiol. 2016, 115, 178–192. [Google Scholar] [CrossRef] [PubMed]
- Nigg, B.M.; MacIntosh, B.R.; Mester, J. Biomechanics and Biology of Movement; Human Kinetics: Champaign, IL, USA, 2000. [Google Scholar]
- Defreitas, J.M.; Beck, T.W.; Ye, X.; Stock, M.S. Synchronization of low- and high-threshold motor units. Muscle Nerve 2014, 49, 575–583. [Google Scholar] [CrossRef] [PubMed]
- Kleine, B.U.; Stegeman, D.F.; Mund, D.; Anders, C. Influence of motoneuron firing synchronization on SEMG characteristics in dependence of electrode position. J. Appl. Physiol. 2001, 91, 1588–1599. [Google Scholar] [PubMed]
- Yao, W.X.; Fuglevand, A.J.; Enoka, R.M. Motor-unit synchronization increases EMG amplitude and decreases force steadiness of simulated contractions. J. Neurophysiol. 2000, 83, 441–452. [Google Scholar] [PubMed]
- Semmler, J.G. Motor unit synchronization and neuromuscular performance. Exerci. Sport Sci. Rev. 2002, 30, 8–14. [Google Scholar] [CrossRef]
- Brown, L.E. Isokinetics in Human Performance; Human Kinetics: Champaign, IL, USA, 2000. [Google Scholar]
- Orizio, C.; Perini, R.; Diemont, B.; Figini, M.M.; Veicsteinas, A. Spectral analysis of muscular sound during isometric contraction of biceps brachii. J. Appl. Physiol. 1990, 68, 508–512. [Google Scholar] [PubMed]
- Orizio, C.; Gobbo, M.; Diemont, B.; Esposito, F.; Veicsteinas, A. The surface mechanomyogram as a tool to describe the influence of fatigue on biceps brachii motor unit activation strategy. Historical basis and novel evidence. Eur. J. Appl. Physiol. 2003, 90, 326–336. [Google Scholar] [CrossRef] [PubMed]
- Kang, T.; Seo, Y.; Park, J.; Dong, E.; Seo, B.; Han, D. The effects of elbow joint angle change on the elbow flexor muscle activation in pulley with weight exercise. J. Phys. Ther. Sci. 2013, 25, 1133–1136. [Google Scholar] [CrossRef] [PubMed]
- Yang, J.; Lee, J.; Lee, B.; Kim, S.; Shin, D.; Lee, Y.; Lee, J.; Han, D.; Choi, S. The Effects of Elbow Joint Angle Changes on Elbow Flexor and Extensor Muscle Strength and Activation. J. Phys. Ther. Sci. 2014, 26, 1079–1082. [Google Scholar] [CrossRef] [PubMed]
- Barbero, M.; Merletti, R.; Rainoldi, A.; Jull, G.A. Atlas of Muscle Innervation Zones: Understanding Surface Electromyography and Its Applications; Springer: Milan, Italy, 2012. [Google Scholar]
- Brown, L.E.; Whitehurst, M.; Gilbert, R.; Buchalter, D.N. The effect of velocity and gender on load range during knee extension and flexion exercise on an isokinetic device. J. Orthop. Sports Phys. Ther. 1995, 21, 107–112. [Google Scholar] [CrossRef] [PubMed]
- Diemont, B.; Figini, M.M.; Orizio, C.; Perini, R.; Veicsteinas, A. Spectral analysis of muscular sound at low and high contraction level. Int. J. Biomed. Comput. 1988, 23, 161–175. [Google Scholar] [CrossRef]
- Kwatny, E.; Thomas, D.H.; Kwatny, H.G. An application of signal processing techniques to the study of myoelectric signals. IEEE Trans. Biomed. Eng. 1970, 17, 303–313. [Google Scholar] [CrossRef] [PubMed]
- Hermens, H.J.; Freriks, B.; Disselhorst-Klug, C.; Rau, G. Development of recommendations for SEMG sensors and sensor placement procedures. J. Electromyogr. Kinesiol. 2000, 10, 361–374. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| Neuromuscular Parameters | Concentric PT | Isometric PT | |||
|---|---|---|---|---|---|
| BB | BR | BB | BR | ||
| EMG AMP (μV) | Pretest | 1.08 ± 0.20 | 1.04 ± 0.16 | 1.00 ± 0.00 | 1.00 ± 0.00 |
| Posttest | 1.12 ± 0.40 | 0.90 ± 0.16 | 1.05 ± 0.37 | 0.97 ± 0.17 | |
| EMG MPF (Hz) | Pretest | 1.05 ± 0.27 | 1.01 ± 0.12 | 1.00 ± 0.00 | 1.00 ± 0.00 |
| Posttest | 0.93 ± 0.24 | 0.97 ± 0.13 | 1.03 ± 0.27 | 0.96 ± 0.15 | |
| MMG AMP (m·s−2) | Pretest | 1.23 ± 0.42 | 1.21 ± 0.32 | 1.00 ± 0.00 | 1.00 ± 0.00 |
| Posttest | 1.42 ± 0.46 | 1.43 ± 0.38 | 1.12 ± 0.27 | 0.81 ± 0.22 | |
| MMG MPF (Hz) | Pretest | 1.23 ± 0.49 | 1.13 ± 0.38 | 1.00 ± 0.00 | 1.00 ± 0.00 |
| Posttest | 1.12 ± 0.35 | 1.19 ± 0.27 | 0.98 ± 0.33 | 0.95 ± 0.31 | |
© 2016 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC-BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hill, E.; Housh, T.; Smith, C.; Schmidt, R.; Johnson, G. Muscle- and Mode-Specific Responses of the Forearm Flexors to Fatiguing, Concentric Muscle Actions. Sports 2016, 4, 47. https://0-doi-org.brum.beds.ac.uk/10.3390/sports4040047
Hill E, Housh T, Smith C, Schmidt R, Johnson G. Muscle- and Mode-Specific Responses of the Forearm Flexors to Fatiguing, Concentric Muscle Actions. Sports. 2016; 4(4):47. https://0-doi-org.brum.beds.ac.uk/10.3390/sports4040047
Chicago/Turabian StyleHill, Ethan, Terry Housh, Cory Smith, Richard Schmidt, and Glen Johnson. 2016. "Muscle- and Mode-Specific Responses of the Forearm Flexors to Fatiguing, Concentric Muscle Actions" Sports 4, no. 4: 47. https://0-doi-org.brum.beds.ac.uk/10.3390/sports4040047