Mechanisms of Hamstring Strain Injury: Interactions between Fatigue, Muscle Activation and Function

 ,
,
Abstract
:1. Introduction
2. Mechanisms of Hamstring Strain Injury
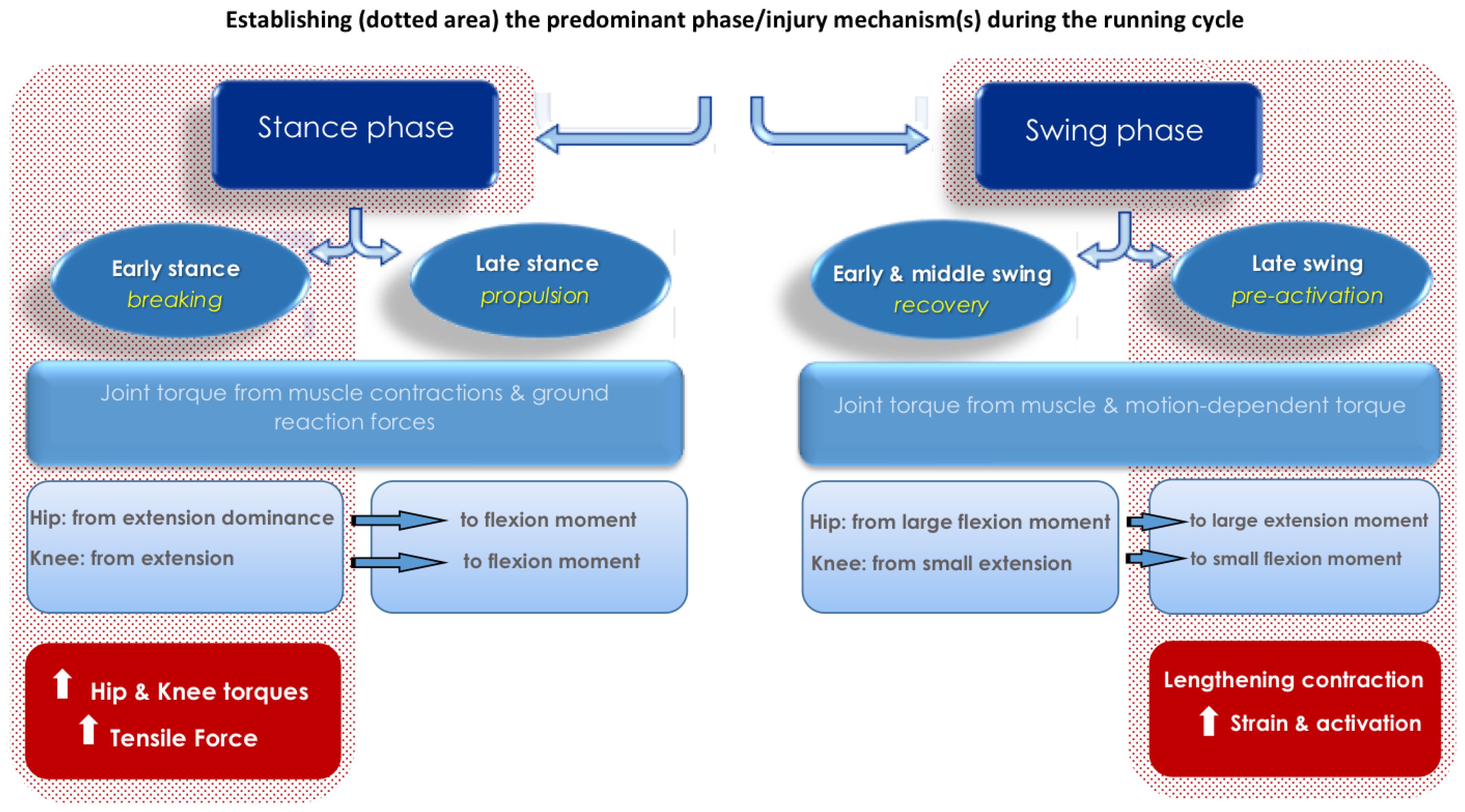
2.1. Hip, Knee and Hamstring Mechanics during High-Speed Running
2.2. The Late Swing Phase
2.3. The Early Stance Phase
2.4. The Swing-Stance Transition Period
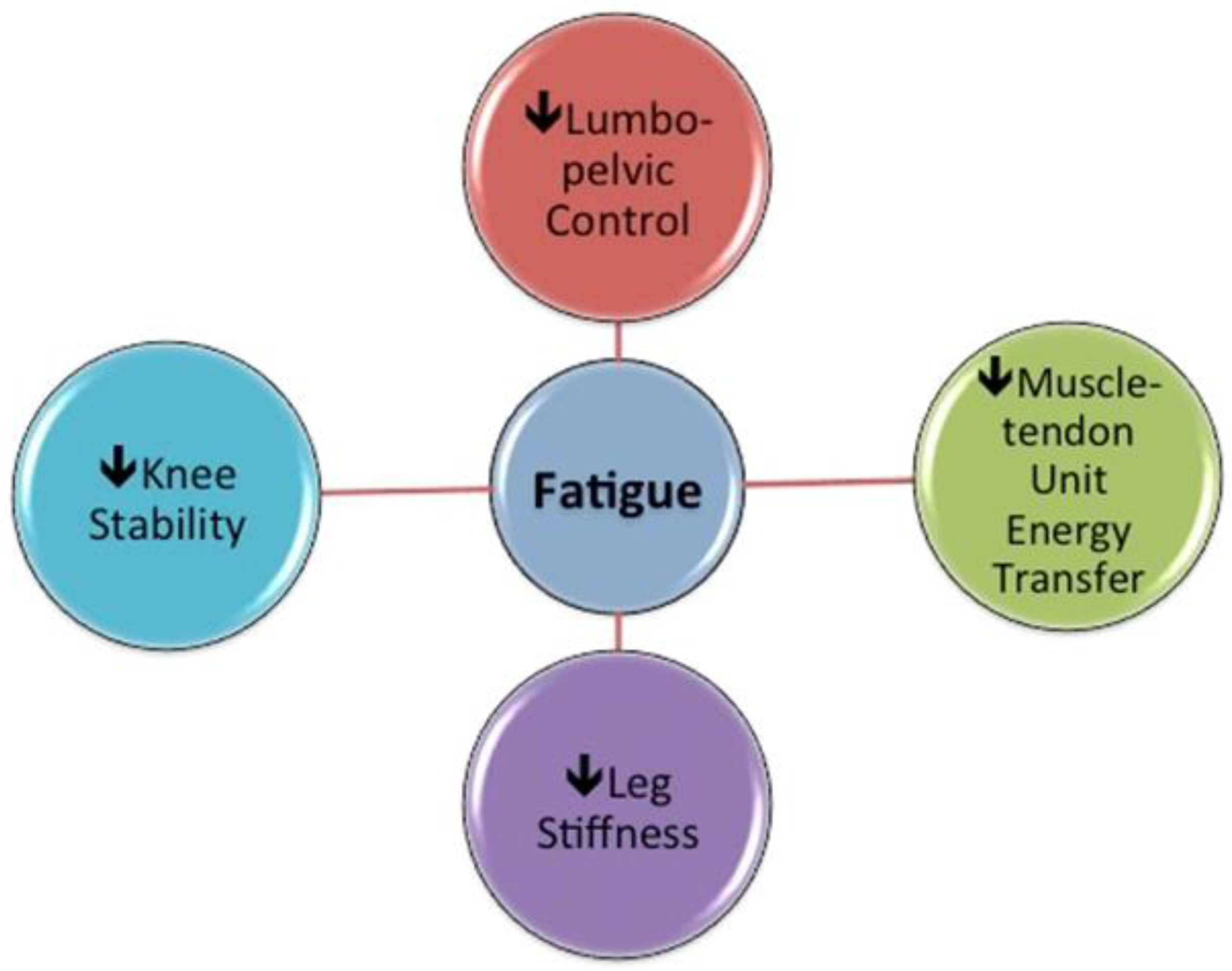
2.5. Interactions between Fatigue, Hamstring Muscle Activation and Function
3. Conclusions
4. Future Directions
Author Contributions
Funding
Conflicts of Interest
References
- Gabbe, B.J.; Bennell, K.L.; Finch, C.F.; Wajswelner, H.; Orchard, J.W. Predictors of hamstring injury at the elite level of Australian football. Scand. J. Med. Sci. Sports 2006, 16, 7–13. [Google Scholar] [CrossRef] [PubMed]
- Gabbe, B.; Finch, C.; Wajswelner, H.; Bennell, K. Australian football: Injury profile at the community level. J. Sci. Med. Sport 2002, 5, 149–160. [Google Scholar] [CrossRef]
- Orchard, J.; Wood, T.; Seward, H.; Broad, A. Comparison of injuries in elite senior and junior Australian football. J. Sci. Med. Sport 1998, 1, 83–88. [Google Scholar] [CrossRef]
- Orchard, J.; Seward, H. Epidemiology of injuries in the Australian Football League, seasons 1997–2000. Br. J. Sports Med. 2002, 36, 39–44. [Google Scholar] [CrossRef] [Green Version]
- Seward, H.; Orchard, J.; Hazard, H.; Collinson, D. Football injuries in Australia at the élite level. Med. J. Aust. 1993, 159, 298–301. [Google Scholar] [CrossRef]
- Feeley, B.T.; Kennelly, S.; Barnes, R.P.; Muller, M.S.; Kelly, B.T.; Rodeo, S.A.; Warren, R.F. Epidemiology of National Football League training camp injuries from 1998 to 2007. Am. J. Sports Med. 2008, 36, 1597–1603. [Google Scholar] [CrossRef]
- Brooks, J.H.M.; Fuller, C.W.; Kemp, S.P.T.; Reddin, D.B. Incidence, risk, and prevention of hamstring muscle injuries in professional rugby union. Am. J. Sports Med. 2006, 34, 1297–1306. [Google Scholar] [CrossRef]
- Brooks, J.H.M.; Fuller, C.W.; Kemp, S.P.T.; Reddin, D.B. Epidemiology of injuries in English professional rugby union: Part 2 training Injuries. Br. J. Sports Med. 2005, 39, 767–775. [Google Scholar] [CrossRef] [Green Version]
- Brooks, J.H.M.; Fuller, C.W.; Kemp, S.P.T.; Reddin, D.B. Epidemiology of injuries in English professional rugby union: Part 1 match injuries. Br. J. Sports Med. 2005, 39, 757–766. [Google Scholar] [CrossRef] [Green Version]
- Ekstrand, J.; Hagglund, M.; Walden, M. Injury incidence and injury patterns in professional football: The UEFA injury study. Br. J. Sports Med. 2011, 45, 553–558. [Google Scholar] [CrossRef] [Green Version]
- Hawkins, R.D.; Hulse, M.A.; Wilkinson, C.; Hodson, A.; Gibson, M. The association football medical research programme: An audit of injuries in professional football. Br. J. Sports Med. 2001, 35, 43–47. [Google Scholar] [CrossRef] [PubMed]
- Henderson, G.; Barnes, C.A.; Portas, M.D. Factors associated with increased propensity for hamstring injury in English Premier League soccer players. J. Sci. Med. Sport 2010, 13, 397–402. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Woods, C.; Hawkins, R.D.; Maltby, S.; Hulse, M.; Thomas, A.; Hodson, A. The football association medical research programme: An audit of injuries in professional football—Analysis of hamstring injuries. Br. J. Sports Med. 2004, 38, 36–41. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, H.; Garrett, W.E.; Moorman, C.T.; Yu, B. Injury rate, mechanism, and risk factors of hamstring strain injuries in sports: A review of the literature. J. Sport Health Sci. 2012, 1, 92–101. [Google Scholar] [CrossRef] [Green Version]
- Opar, D.A.; Williams, M.D.; Shield, A.J. Hamstring strain injuries: Factors that lead to injury and re-injury. Sports Med. 2012, 42, 209–226. [Google Scholar] [CrossRef]
- Ekstrand, J.; Hagglund, M.; Walden, M. Epidemiology of muscle injuries in professional football (soccer). Am. J. Sports Med. 2011, 39, 1226–1232. [Google Scholar] [CrossRef] [Green Version]
- Hagglund, M.; Walden, M.; Ekstrand, J. Risk factors for lower extremity muscle injury in professional soccer the UEFA injury study. Am. J. Sports Med. 2013, 41, 327–335. [Google Scholar] [CrossRef] [Green Version]
- Heiderscheit, B.C.; Sherry, M.A.; Silder, A.; Chumanov, E.S.; Thelen, D.G. Hamstring strain injuries: Recommendations for diagnosis, rehabilitation, and injury prevention. J. Orthop. Sports Phys. Ther. 2010, 40, 67–81. [Google Scholar] [CrossRef] [Green Version]
- Buckthorpe, M.; Wright, S.; Bruce-Low, S.; Nanni, G.; Sturdy, T.; Gross, A.S.; Bowen, L.; Styles, B.; Della Villa, S.; Davison, M.; et al. Recommendations for hamstring injury prevention in elite football: Translating research into practice. Br. J. Sports Med. 2019, 53, 449–456. [Google Scholar] [CrossRef]
- Sun, Y.; Wei, S.; Zhong, Y.; Fu, W.; Li, L.; Liu, Y. How joint torques affect hamstring injury risk in sprinting swing–stance transition. Med. Sci. Sports Exerc. 2015, 47, 373–380. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ekstrand, J.; Waldén, M.; Hägglund, M. Hamstring injuries have increased by 4% annually in men’s professional football, since 2001: A 13-year longitudinal analysis of the UEFA Elite Club injury study. Br. J. Sports Med. 2016, 50, 731–737. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hägglund, M.; Waldén, M.; Magnusson, H.; Kristenson, K.; Bengtsson, H.; Ekstrand, J. Injuries affect team performance negatively in professional football: An 11-year follow-up of the UEFA Champions League injury study. Br. J. Sports Med. 2013, 47, 738–742. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Askling, C.M.; Tengvart, M.; Saartok, T.; Thorstensson, A. Acute first-time hamstring strains during high-speed running—A longitudinal study including clinical and magnetic resonance imaging findings. Am. J. Sports Med. 2007, 35, 197–206. [Google Scholar] [CrossRef] [PubMed]
- Ekstrand, J. Keeping your top players on the pitch: The key to football medicine at a professional level. Br. J. Sports Med. 2013, 47, 723–724. [Google Scholar] [CrossRef] [Green Version]
- Marshall, P.W.M.; Lovell, R.; Jeppesen, G.K.; Andersen, K.; Siegler, J.C. Hamstring muscle fatigue and central motor output during a simulated soccer match. PLoS ONE 2014, 9, e102753. [Google Scholar] [CrossRef] [Green Version]
- Timmins, R.G.; Opar, D.A.; Williams, M.D.; Schache, A.G.; Dear, N.M.; Shield, A.J. Reduced biceps femoris myoelectrical activity influences eccentric knee flexor weakness after repeat sprint running. Scand. J. Med. Sci. Sports 2014, 24, e299–e305. [Google Scholar] [CrossRef] [Green Version]
- Marqués-Jiménez, D.; Calleja-González, J.; Arratibel, I.; Delextrat, A.; Terrados, N. Fatigue and recovery in soccer: Evidence and challenges. Open Sports Sci. J. 2017, 10, 52–70. [Google Scholar] [CrossRef] [Green Version]
- Boyas, S.; Guével, A. Neuromuscular fatigue in healthy muscle: Underlying factors and adaptation mechanisms. Ann. Phys. Rehabil. Med. 2011, 54, 88–108. [Google Scholar] [CrossRef] [Green Version]
- Silva, J.R.; Rumpf, M.C.; Hertzog, M.; Castagna, C.; Farooq, A.; Girard, O.; Hader, K. Acute and residual soccer match-related fatigue: A systematic review and meta-analysis. Sports Med. 2018, 48, 539–583. [Google Scholar] [CrossRef]
- Clarkson, P.M.; Nosaka, K.; Braun, B. Muscle function after exercise-induced muscle damage and rapid adaptation. Med. Sci. Sports Exerc. 1992, 24, 512–520. [Google Scholar] [CrossRef] [PubMed]
- Gabbett, T.J. The training—Injury prevention paradox: Should athletes be training smarter and harder? Br. J. Sports Med. 2016, 50, 273–280. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bengtsson, H.; Ekstrand, J.; Waldén, M.; Hägglund, M. Muscle injury rate in professional football is higher in matches played within 5 days since the previous match: A 14-year prospective study with more than 130 000 match observations. Br. J. Sports Med. 2018, 52, 1116–1122. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Thorpe, R.T.; Atkinson, G.; Drust, B.; Gregson, W. Monitoring fatigue status in elite team-sport athletes: Implications for practice. Int. J. Sports Physiol. Perform. 2017, 12, S227–S234. [Google Scholar] [CrossRef] [Green Version]
- Yu, B.; Liu, H.; Garrett, W.E. Mechanism of hamstring muscle strain injury in sprinting. J. Sport Health Sci. 2017, 6, 130–132. [Google Scholar] [CrossRef]
- Askling, C.M.; Tengvar, M.; Thorstensson, A. Acute hamstring injuries in Swedish elite football: A prospective randomised controlled clinical trial comparing two rehabilitation protocols. Br. J. Sports Med. 2013, 47, 953–959. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Askling, C.; Saartok, T.; Thorstensson, A. Type of acute hamstring strain affects flexibility, strength, and time to return to pre-injury level. Br. J. Sports Med. 2006, 40, 40–44. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Askling, C.M.; Tengvar, M.; Saartok, T.; Thorstensson, A. Acute first-time hamstring strains during slow-speed stretching: Clinical, magnetic resonance imaging, and recovery characteristics. Am. J. Sports Med. 2007, 35, 1716–1724. [Google Scholar] [CrossRef] [PubMed]
- Koulouris, G.; Connell, D. Evaluation of the hamstring muscle complex following acute injury. Skelet. Radiol. 2003, 32, 582–589. [Google Scholar] [CrossRef]
- Silder, A.; Thelen, D.G.; Heiderscheit, B.C. Effects of prior hamstring strain injury on strength, flexibility, and running mechanics. Clin. Biomech. 2010, 25, 681–686. [Google Scholar] [CrossRef] [Green Version]
- Van der Made, A.D.; Wieldraaijer, T.; Kerkhoffs, G.M.; Kleipool, R.P.; Engebretsen, L.; van Dijk, C.N.; Golanó, P. The hamstring muscle complex. Knee Surg. Sports Traumatol. Arthrosc. Off. J. ESSKA 2015, 23, 2115–2122. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Sun, Y.; Zhu, W.; Yu, J. The late swing and early stance of sprinting are most hazardous for hamstring injuries. J. Sport Health Sci. 2017, 6, 133–136. [Google Scholar] [CrossRef] [PubMed]
- Howard, R.M.; Conway, R.; Harrison, A.J. Muscle activity in sprinting: A review. Sports Biomech. 2018, 17, 1–17. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kenneally-Dabrowski, C.J.B.; Brown, N.A.T.; Lai, A.K.M.; Perriman, D.; Spratford, W.; Serpell, B.G. Late swing or early stance? A narrative review of hamstring injury mechanisms during high-speed running. Scand. J. Med. Sci. Sports 2019, 29, 1083–1091. [Google Scholar] [CrossRef] [PubMed]
- Schache, A.G.; Blanch, P.D.; Dorn, T.W.; Brown, N.A.T.; Rosemond, D.; Pandy, M.G. Effect of running speed on lower limb joint kinetics. Med. Sci. Sports Exerc. 2011, 43, 1260–1271. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schache, A.G.; Dorn, T.W.; Blanch, P.D.; Brown, N.A.T.; Pandy, M.G. Mechanics of the human hamstring muscles during sprinting. Med. Sci. Sports Exerc. 2012, 44, 647–658. [Google Scholar] [CrossRef] [Green Version]
- Nagano, Y.; Higashihara, A.; Takahashi, K.; Fukubayashi, T. Mechanics of the muscles crossing the hip joint during sprint running. J. Sports Sci. 2014, 32, 1722–1728. [Google Scholar] [CrossRef]
- Thelen, D.; Chumanov, E.S.; Best, T.M.; Swanson, S.C.; Heiderscheit, B.C. Simulation of biceps femoris musculotendon mechanics during the swing phase of sprinting. Med. Sci. Sports Exerc. 2005, 37, 1931–1938. [Google Scholar] [CrossRef] [Green Version]
- Higashihara, A.; Nagano, Y.; Takahashi, K.; Fukubayashi, T. Effects of forward trunk lean on hamstring muscle kinematics during sprinting. J. Sports Sci. 2015, 33, 1366–1375. [Google Scholar] [CrossRef]
- Yu, B.; Queen, R.M.; Abbey, A.N.; Liu, Y.; Moorman, C.T.; Garrett, W.E. Hamstring muscle kinematics and activation during overground sprinting. J. Biomech. 2008, 41, 3121–3126. [Google Scholar] [CrossRef]
- Beltran, L.; Ghazikhanian, V.; Padron, M.; Beltran, J. The proximal hamstring muscle-tendon-bone unit: A review of the normal anatomy, biomechanics, and pathophysiology. Eur. J. Radiol. 2012, 81, 3772–3779. [Google Scholar] [CrossRef] [PubMed]
- Schuermans, J.; Van Tiggelen, D.; Palmans, T.; Danneels, L.; Witvrouw, E. Deviating running kinematics and hamstring injury susceptibility in male soccer players: Cause or consequence? Gait Posture 2017, 57, 270–277. [Google Scholar] [CrossRef] [PubMed]
- Chumanov, E.S.; Heiderscheit, B.C.; Thelen, D.G. Hamstring musculotendon dynamics during stance and swing phases of high-speed running. Med. Sci. Sports Exerc. 2011, 43, 525–532. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fiorentino, N.M.; Rehorn, M.R.; Chumanov, E.S.; Thelen, D.G.; Blemker, S.S. Computational models predict larger muscle tissue strains at faster sprinting speeds. Med. Sci. Sports Exerc. 2014, 46, 776–786. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Heiderscheit, B.C.; Hoerth, D.M.; Chumanov, E.S.; Swanson, S.C.; Thelen, B.J.; Thelen, D.G. Identifying the time of occurrence of a hamstring strain injury during treadmill running: A case study. Clin. Biomech. 2005, 20, 1072–1078. [Google Scholar] [CrossRef]
- Higashihara, A.; Nagano, Y.; Ono, T.; Fukubayashi, T. Relationship between the peak time of hamstring stretch and activation during sprinting. Eur. J. Sport Sci. 2014, 16, 36–41. [Google Scholar] [CrossRef]
- Schache, A.G.; Wrigley, T.V.; Baker, R.; Pandy, M.G. Biomechanical response to hamstring muscle strain injury. Gait Posture 2009, 29, 332–338. [Google Scholar] [CrossRef]
- Higashihara, A.; Ono, T.; Kubota, J.; Okuwaki, T.; Fukubayashi, T. Functional differences in the activity of the hamstring muscles with increasing running speed. J. Sports Sci. 2010, 28, 1085–1092. [Google Scholar] [CrossRef]
- Askling, C. Hamstring Muscle Strain; Karolinska Institutet: Stockholm, Sweden, 2008. [Google Scholar]
- Van Hooren, B.; Bosch, F. Is there really an eccentric action of the hamstrings during the swing phase of high-speed running? Part I: A critical review of the literature. J. Sports Sci. 2017, 35, 2313–2321. [Google Scholar] [CrossRef]
- Van Hooren, B.; Bosch, F. Is there really an eccentric action of the hamstrings during the swing phase of high-speed running? Part II: Implications for exercise. J. Sports Sci. 2017, 35, 2322–2333. [Google Scholar] [CrossRef]
- Van Hooren, B.; Bosch, F. Influence of muscle slack on high-intensity sport performance: A review. Strength Cond. J. 2016, 38, 75–87. [Google Scholar] [CrossRef]
- Mair, S.D.; Seaber, A.V.; Glisson, R.R.; Garrett, W.E. The role of fatigue in susceptibility to acute muscle strain injury. Am. J. Sports Med. 1996, 24, 137–143. [Google Scholar] [CrossRef] [PubMed]
- Thelen, D.G.; Chumanov, E.S.; Hoerth, D.M.; Best, T.M.; Swanson, S.C.; Li, L.; Young, M.; Heiderscheit, B.C. Hamstring muscle kinematics during treadmill sprinting. Med. Sci. Sports Exerc. 2005, 37, 108–114. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mann, R.V. A kinetic analysis of sprinting. Med. Sci. Sports Exerc. 1981, 13, 325–328. [Google Scholar] [CrossRef] [PubMed]
- Mann, R.; Sprague, P. A kinetic analysis of the ground leg during sprint running. Res. Q. Exerc. Sport 1980, 51, 334–348. [Google Scholar] [CrossRef] [PubMed]
- Ono, T.; Higashihara, A.; Shinohara, J.; Hirose, N.; Fukubayashi, T. Estimation of tensile force in the hamstring muscles during overground sprinting. Int. J. Sports Med. 2015, 36, 163–168. [Google Scholar] [CrossRef]
- Ono, T.; Okuwaki, T.; Fukubayashi, T. Differences in activation patterns of knee flexor muscles during concentric and eccentric exercises. Res. Sports Med. Print 2010, 18, 188–198. [Google Scholar] [CrossRef]
- Ono, T.; Higashihara, A.; Fukubayashi, T. Hamstring functions during hip-extension exercise assessed with electromyography and magnetic resonance imaging. Res. Sports Med. Print 2011, 19, 42–52. [Google Scholar] [CrossRef]
- Kumazaki, T.; Ehara, Y.; Sakai, T. Anatomy and physiology of hamstring injury. Int. J. Sports Med. 2012, 33, 950–954. [Google Scholar] [CrossRef]
- Avrillon, S.; Guilhem, G.; Barthelemy, A.; Hug, F. Coordination of hamstrings is individual specific and is related to motor performance. J. Appl. Physiol. 2018, 125, 1069–1079. [Google Scholar] [CrossRef]
- Pinniger, G.J.; Steele, J.R.; Groeller, H. Does fatigue induced by repeated dynamic efforts affect hamstring muscle function? Med. Sci. Sports Exerc. 2000, 32, 647–653. [Google Scholar] [CrossRef] [PubMed]
- Schuermans, J.; Van Tiggelen, D.; Danneels, L.; Witvrouw, E. Biceps femoris and semitendinosus--teammates or competitors? New insights into hamstring injury mechanisms in male football players: A muscle functional MRI study. Br. J. Sports Med. 2014, 48, 1599–1606. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Small, K.; McNaughton, L.R.; Greig, M.; Lohkamp, M.; Lovell, R. Soccer fatigue, sprinting and hamstring injury risk. Int. J. Sports Med. 2009, 30, 573–578. [Google Scholar] [CrossRef] [PubMed]
- Franettovich Smith, M.M.; Bonacci, J.; Mendis, M.D.; Christie, C.; Rotstein, A.; Hides, J.A. Gluteus medius activation during running is a risk factor for season hamstring injuries in elite footballers. J. Sci. Med. Sport 2017, 20, 159–163. [Google Scholar] [CrossRef]
- Small, K.; McNaughton, L.; Greig, M.; Lovell, R. The effects of multidirectional soccer-specific fatigue on markers of hamstring injury risk. J. Sci. Med. Sport 2010, 13, 120–125. [Google Scholar] [CrossRef]
- Cohen, D.D.; Zhao, B.; Okwera, B.; Matthews, M.J.; Delextrat, A. Angle-specific eccentric hamstring fatigue after simulated soccer. Int. J. Sports Physiol. Perform. 2015, 10, 325–331. [Google Scholar] [CrossRef]
- Mero, A.; Komi, P.V.; Gregor, R.J. Biomechanics of sprint running. A review. Sports Med. 1992, 13, 376–392. [Google Scholar] [CrossRef]
- Hader, K.; Mendez-Villanueva, A.; Ahmaidi, S.; Williams, B.K.; Buchheit, M. Changes of direction during high-intensity intermittent runs: Neuromuscular and metabolic responses. BMC Sports Sci. Med. Rehabil. 2014, 6, 2. [Google Scholar] [CrossRef] [Green Version]
- Hannah, R.; Minshull, C.; Smith, S.L.; Folland, J.P. Longer electromechanical delay impairs hamstrings explosive force versus quadriceps. Med. Sci. Sports Exerc. 2014, 46, 963–972. [Google Scholar] [CrossRef]
- Zebis, M.K.; Andersen, L.L.; Ellingsgaard, H.; Aagaard, P. Rapid hamstring/quadriceps force capacity in male vs. female elite soccer players. J. Strength Cond. Res. 2011, 25, 1989–1993. [Google Scholar] [CrossRef]
- Sanchez, A.R.; Sugalski, M.T.; LaPrade, R.F. Anatomy and biomechanics of the lateral side of the knee. Sports Med. Arthrosc. Rev. 2006, 14, 2–11. [Google Scholar] [CrossRef] [PubMed]
- Cleather, D.J. An important role of the biarticular hamstrings is to exert internal/external rotation moments on the tibia during vertical jumping. J. Theor. Biol. 2018, 455, 101–108. [Google Scholar] [CrossRef] [PubMed]
- Turner, A.N.; Jeffreys, I. The Stretch-shortening Cycle: Proposed Mechanisms and Methods for Enhancement. Strength Cond. J. 2010, 32, 87–99. [Google Scholar] [CrossRef] [Green Version]
- Lehnert, M.; De Ste Croix, M.; Zaatar, A.; Hughes, J.; Varekova, R.; Lastovicka, O. Muscular and neuromuscular control following soccer-specific exercise in male youth: Changes in injury risk mechanisms. Scand. J. Med. Sci. Sports 2017, 27, 975–982. [Google Scholar] [CrossRef] [PubMed]
- Cormack, S.J.; Mooney, M.G.; Morgan, W.; McGuigan, M.R. Influence of neuromuscular fatigue on accelerometer load in elite Australian football players. Int. J. Sports Physiol. Perform. 2013, 8, 373–378. [Google Scholar] [CrossRef]
- McMahon, T.A.; Valiant, G.; Frederick, E.C. Groucho running. J. Appl. Physiol. 1987, 62, 2326–2337. [Google Scholar] [CrossRef]
- Hanon, C.; Thépaut-Mathieu, C.; Vandewalle, H. Determination of muscular fatigue in elite runners. Eur. J. Appl. Physiol. 2005, 94, 118–125. [Google Scholar] [CrossRef]
- Morin, J.-B.; Gimenez, P.; Edouard, P.; Arnal, P.; Jimenez-Reyes, P.; Samozino, P.; Brughelli, M.; Mendiguchia, J. Sprint acceleration mechanics: The major role of hamstrings in horizontal force production. Front. Physiol. 2015, 6, 404. [Google Scholar] [CrossRef]
- Edouard, P.; Mendiguchia, J.; Lahti, J.; Arnal, P.J.; Gimenez, P.; Jiménez-Reyes, P.; Brughelli, M.; Samozino, P.; Morin, J.-B. Sprint acceleration mechanics in fatigue conditions: Compensatory role of gluteal muscles in horizontal force production and potential protection of hamstring muscles. Front. Physiol. 2018, 9, 1706. [Google Scholar] [CrossRef] [Green Version]
- Schuermans, J.; Danneels, L.; Van Tiggelen, D.; Palmans, T.; Witvrouw, E. Proximal neuromuscular control protects against hamstring injuries in male soccer players: A prospective study with electromyography time-series analysis during maximal sprinting. Am. J. Sports Med. 2017, 45, 1315–1325. [Google Scholar] [CrossRef]
- Schuermans, J.; Van Tiggelen, D.; Danneels, L.; Witvrouw, E. Susceptibility to hamstring injuries in soccer: A prospective study using muscle functional magnetic resonance imaging. Am. J. Sports Med. 2016, 44, 1276–1285. [Google Scholar] [CrossRef] [PubMed]
- Bourne, M.N.; Timmins, R.G.; Opar, D.A.; Pizzari, T.; Ruddy, J.D.; Sims, C.; Williams, M.D.; Shield, A.J. An evidence-based framework for strengthening exercises to prevent hamstring injury. Sports Med. 2018, 48, 251–267. [Google Scholar] [CrossRef] [PubMed]
- Delextrat, A.; Piquet, J.; Matthews, M.J.; Cohen, D.D. Strength-endurance training reduces the hamstrings strength decline following simulated football competition in female players. Front. Physiol. 2018, 9, 1059. [Google Scholar] [CrossRef] [PubMed]
- Lord, C.; Ma’ayah, F.; Blazevich, A.J. Change in knee flexor torque after fatiguing exercise identifies previous hamstring injury in football players. Scand. J. Med. Sci. Sports 2018, 28, 1235–1243. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Freckleton, G.; Cook, J.; Pizzari, T. The predictive validity of a single leg bridge test for hamstring injuries in Australian Rules Football Players. Br. J. Sports Med. 2014, 48, 713–717. [Google Scholar] [CrossRef]
- Orchard, J.W. Hamstrings are most susceptible to injury during the early stance phase of sprinting. Br. J. Sports Med. 2012, 46, 88–89. [Google Scholar] [CrossRef] [Green Version]



{kind=link}
{kind=link}
| Reference (Year) | Participants | Late Swing Phase | Early Stance Phase | Late Stance Phase |
|---|---|---|---|---|
| Schache et al. (2012) [45] | Sprinters (5 males, 2 females) | X | ||
| Chumanov et al. (2011) [52] | Recreational athletes (9 males, 3 females) | X | ||
| Fiorentino et al. (2014) [53] | Track and field athletes (7 males, 7 females) | X | ||
| Higashihara et al. (2014) [55] | Track and field athletes (13 males) | X | ||
| Higashihara et al. (2010) [57] | Track and field athletes (8 males) | X | ||
| Thelen et al. (2005) [63] | Recreational athletes (9 males, 6 females) | X | ||
| Thelen et al. (2005) [47] | Recreational athlete (1 male) | X | ||
| Yu et al. (2008) [49] | Sprinters or middle-distance runners (20 males) | X | X | |
| Mann and Sprague (1980) [65] | Sprinters (15 males) | X | ||
| Mann (1981) [64] | Sprinters (15 males) | X | ||
| Ono et al. (2015) [66] | Track and field, rugby and soccer players (12 males) | X | ||
| Sun et al. (2015) [20] | Sprinters (8 males) | X | X | |
| Liu et al. (2017) [41] | Sprinters (8 males) | X | X | |
| * Schache et al. (2009) [56] | Australian Rules Football player (1 male) | X | ||
| * Heiderscheit et al. (2005) [54] | Skier (1 male) | X |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Huygaerts, S.; Cos, F.; Cohen, D.D.; Calleja-González, J.; Guitart, M.; Blazevich, A.J.; Alcaraz, P.E. Mechanisms of Hamstring Strain Injury: Interactions between Fatigue, Muscle Activation and Function. Sports 2020, 8, 65. https://0-doi-org.brum.beds.ac.uk/10.3390/sports8050065
Huygaerts S, Cos F, Cohen DD, Calleja-González J, Guitart M, Blazevich AJ, Alcaraz PE. Mechanisms of Hamstring Strain Injury: Interactions between Fatigue, Muscle Activation and Function. Sports. 2020; 8(5):65. https://0-doi-org.brum.beds.ac.uk/10.3390/sports8050065
Chicago/Turabian StyleHuygaerts, Shaun, Francesc Cos, Daniel D. Cohen, Julio Calleja-González, Marc Guitart, Anthony J. Blazevich, and Pedro E. Alcaraz. 2020. "Mechanisms of Hamstring Strain Injury: Interactions between Fatigue, Muscle Activation and Function" Sports 8, no. 5: 65. https://0-doi-org.brum.beds.ac.uk/10.3390/sports8050065