
Mucosal-Associated Invariant T Cell Response to Acute Exercise and Exercise Training in Older Obese Women

 ,
,  , , , and
, , , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Participants
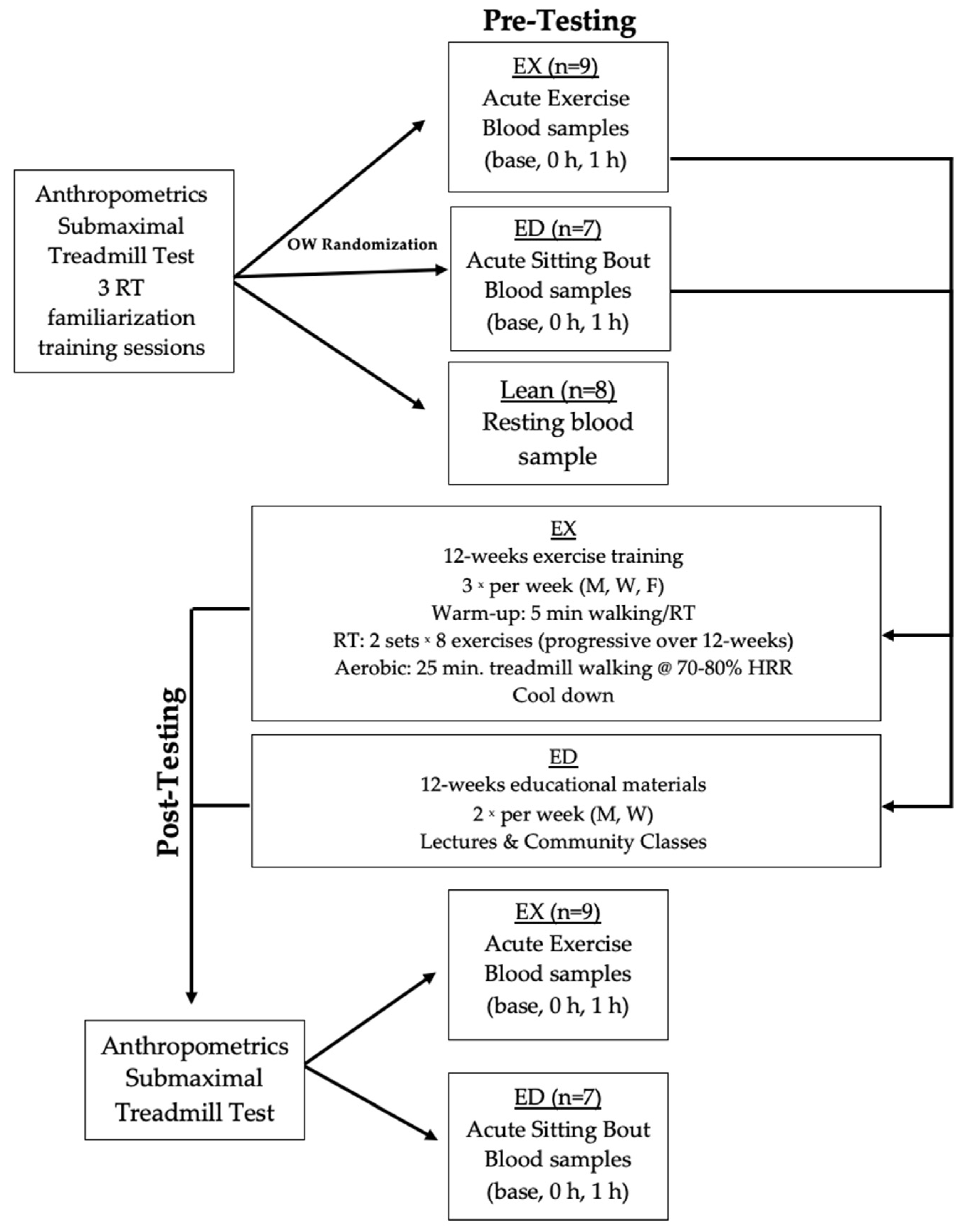
2.2. Experimental Protocol
2.3. Experimental Design
2.4. Acute Exercise Testing
2.5. Lean Acute Exercise Testing
2.6. Exercise Training
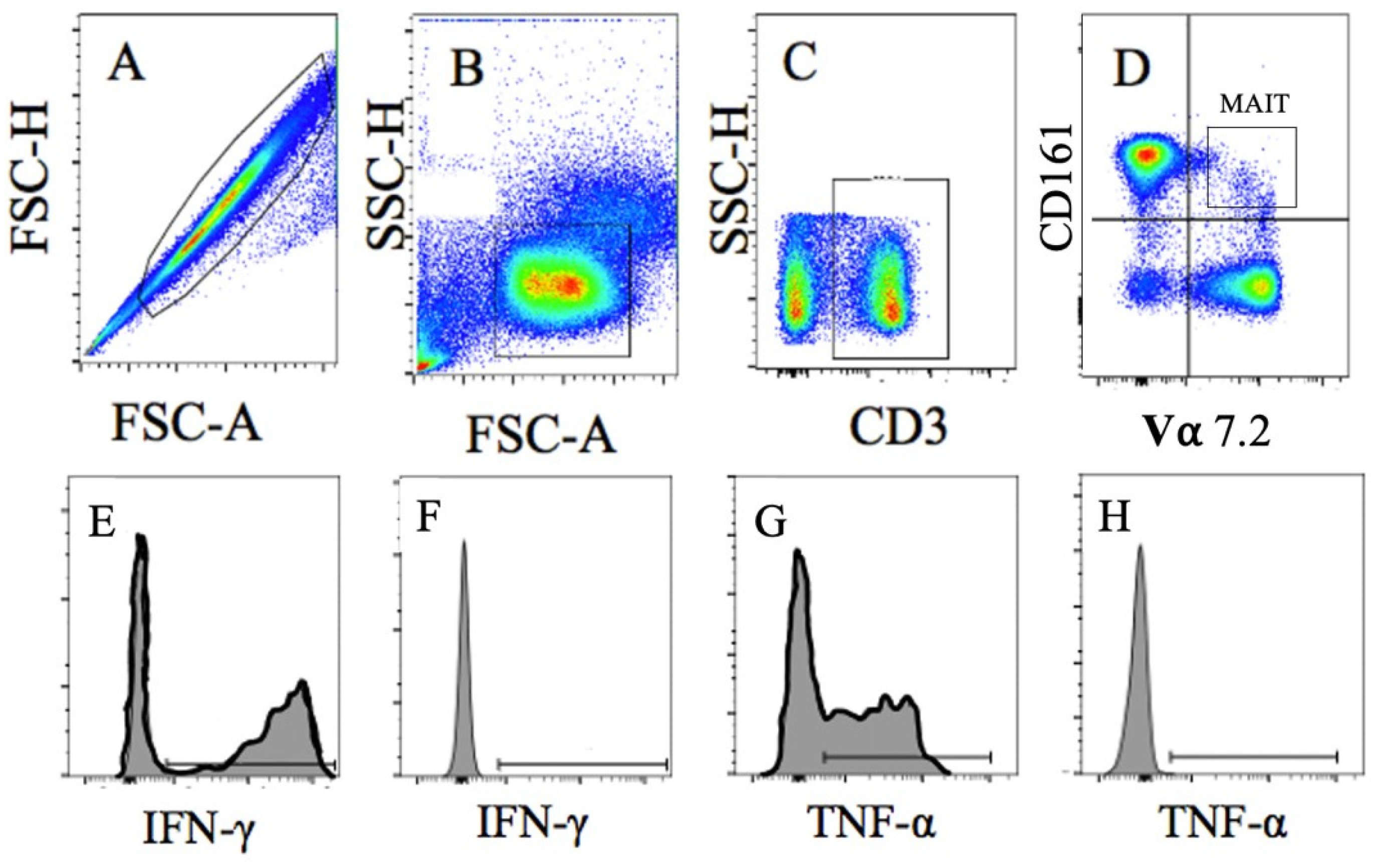
2.7. Blood Analysis
2.8. Statistical Analysis
3. Results
3.1. Participants
3.2. Baseline Comparison-Pre-Training
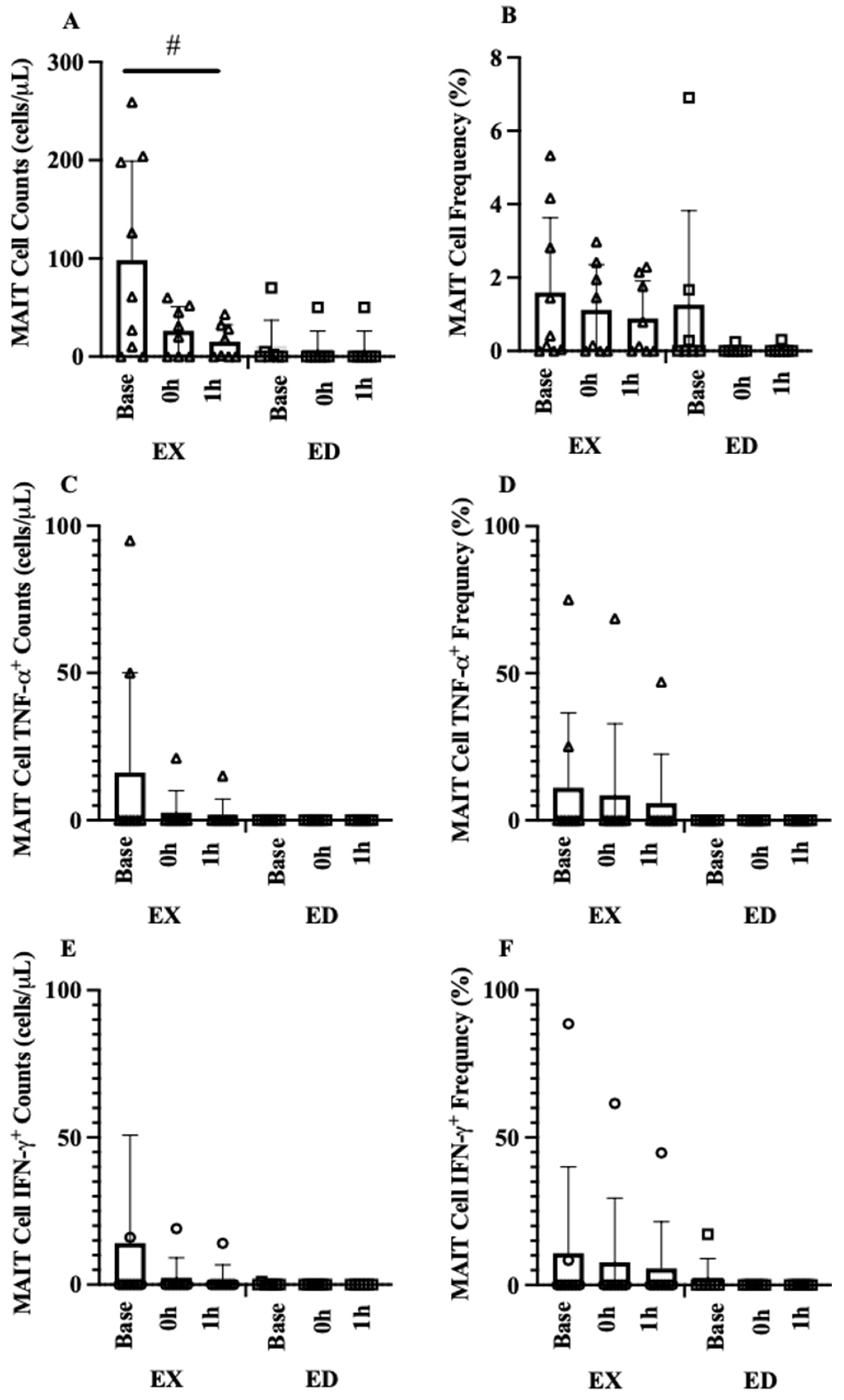
3.3. Acute Exercise
3.3.1. Acute Hematological Response
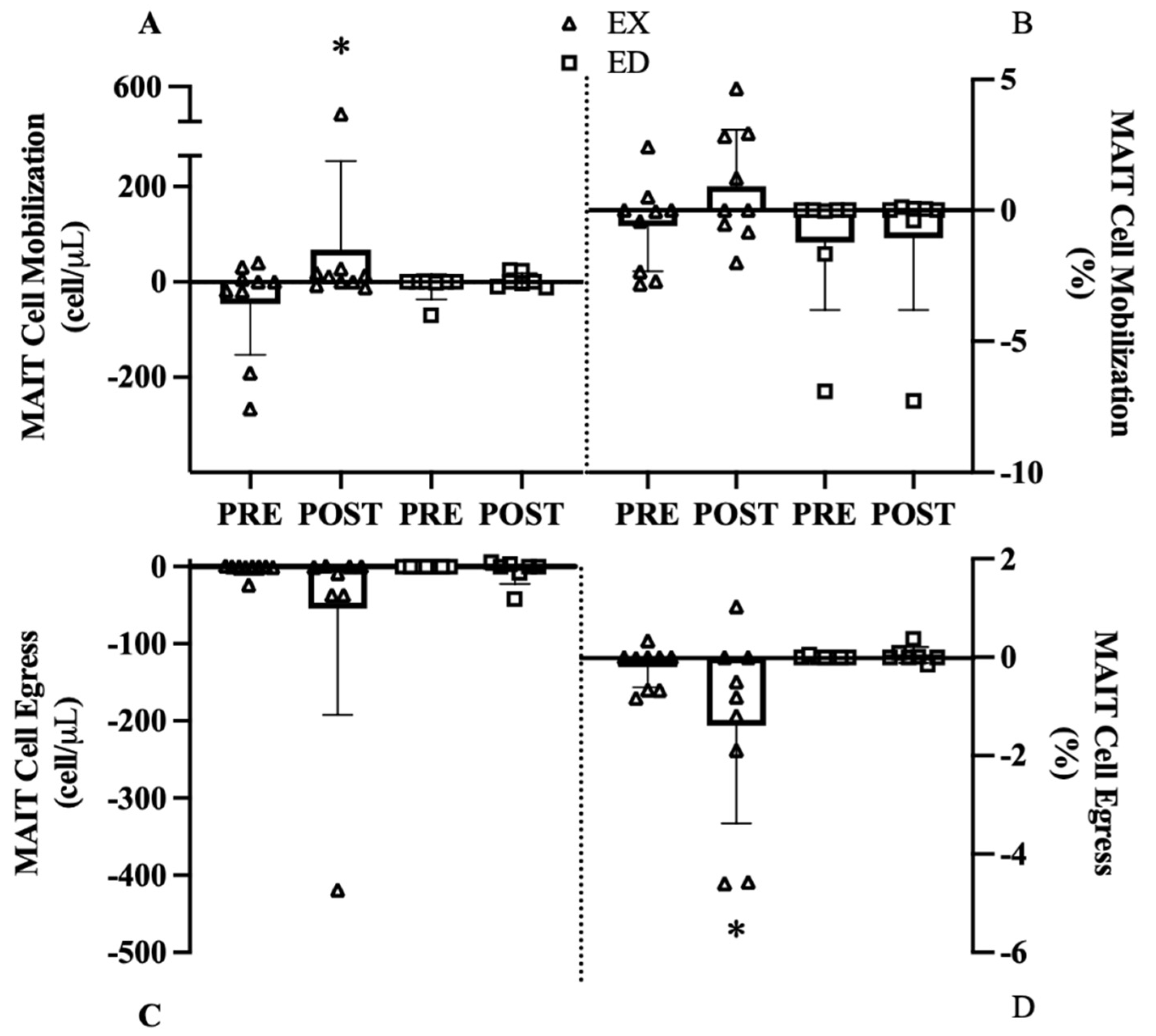
3.3.2. Acute MAIT-Cell Counts and Frequencies
3.4. Exercise Training
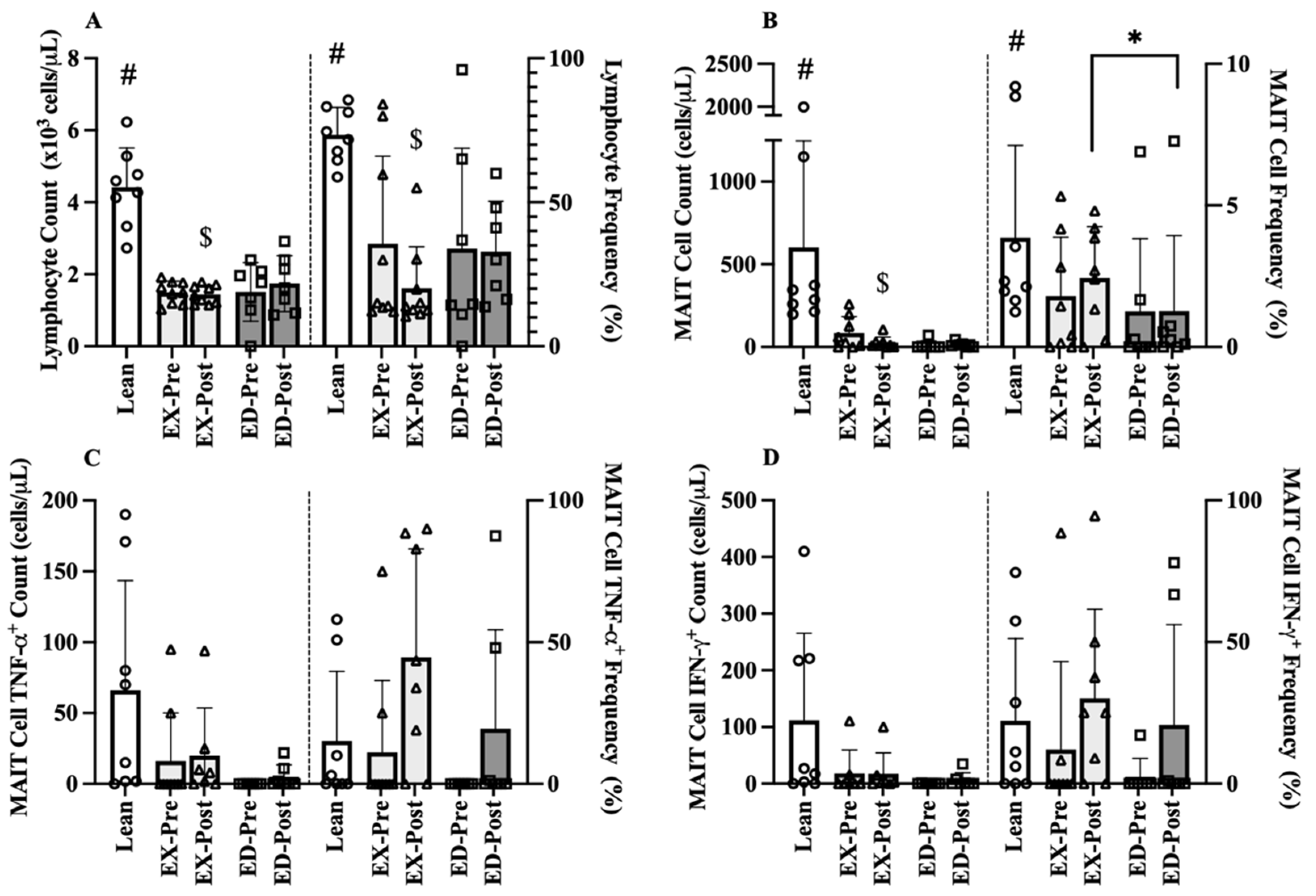
3.4.1. Baseline Comparison-Post-Training
3.4.2. MAIT-Cell Counts and Frequencies
3.4.3. MAIT-Cell Cytokine Counts and Frequencies
4. Discussion
4.1. Limitations and Strengths
4.2. Acute Hemotological Response
4.3. Circulating Baseline MAIT-Cells
4.4. Acute Exercise Effect
4.5. Exercise Training Effect
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- World Health Organization. Obesity and Overweight; World Health Organization: Geneva, Switzerland, 2020; pp. 1–2. [Google Scholar]
- Van Greevenbroek, M.M.; Schalkwijk, C.G.; Stehouwer, C.D. Obesity-associated low-grade inflammation in type 2 diabetes mellitus: Causes and consequences. Neth. J. Med. 2013, 71, 174–187. [Google Scholar]
- Brien, A.O.; Kedia-Mehta, N.; Tobin, L.; Veerapen, N.; Besra, G.S.; Shea, D.O.; Hogan, A.E. Targeting mitochondrial dysfunction in MAIT cells limits IL-17 production in obesity. Cell. Mol. Immunol. 2020, 17, 1193–1195. [Google Scholar] [CrossRef]
- Wiklund, P. The role of physical activity and exercise in obesity and weight management: Time for critical appraisal. J. Sport Heal. Sci. 2016, 5, 151–154. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- You, T.; Arsenis, N.C.; Disanzo, B.L.; LaMonte, M.J. Effects of Exercise Training on Chronic Inflammation in Obesity. Sports Med. 2013, 43, 243–256. [Google Scholar] [CrossRef] [PubMed]
- Andersen, C.J.; Murphy, K.E.; Fernandez, M.L. Impact of Obesity and Metabolic Syndrome on Immunity. Adv. Nutr. 2016, 7, 66–75. [Google Scholar] [CrossRef] [Green Version]
- Hanson, E.D.; Bates, L.C.; Bartlett, D.B.; Campbell, J.P. Does exercise attenuate age- and disease-associated dysfunction in unconventional T cells? Shining a light on overlooked cells in exercise immunology. Eur. J. Appl. Physiol. 2021, 121, 1815–1834. [Google Scholar] [CrossRef] [PubMed]
- Touch, S.; Clément, K.; André, S. T Cell Populations and Functions Are Altered in Human Obesity and Type 2 Diabetes. Curr. Diab. Rep. 2017, 17, 81. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Costanzo, A.E.; Taylor, K.R.; Dutt, S.; Han, P.P.; Fujioka, K.; Jameson, J.M. Obesity impairs γδ T cell homeostasis and antiviral function in humans. PLoS ONE 2015, 10, e0120918. [Google Scholar] [CrossRef]
- Magalhaes, I.; Kiaf, B.; Lehuen, A. iNKT and MAIT cell alterations in diabetes. Front. Immunol. 2015, 6, 6–12. [Google Scholar] [CrossRef]
- Carolan, E.; Tobin, L.M.; Mangan, B.A.; Corrigan, M.; Gaoatswe, G.; Byrne, G.; Geoghegan, J.; Cody, D.; O’Connell, J.; Winter, D.C.; et al. Altered Distribution and Increased IL-17 Production by Mucosal-Associated Invariant T Cells in Adult and Childhood Obesity. J. Immunol. 2015, 194, 5775–5780. [Google Scholar] [CrossRef]
- Apostolopoulos, V.; de Courten, M.P.J.; Stojanovska, L.; Blatch, G.L.; Tangalakis, K.; de Courten, B. The Complex Immunological and Inflammatory Network of Adipose Tissue in Obesity. Mol. Nutr. Food Res. 2016, 60, 43–57. [Google Scholar] [CrossRef] [PubMed]
- O’Brien, A.; Loftus, R.M.; Pisarska, M.M.; Tobin, L.M.; Bergin, R.; Wood, N.A.W.; Foley, C.; Mat, A.; Tinley, F.C.; Bannan, C. Obesity reduces mTORC1 activity in mucosal-associated invariant T cells, driving defective metabolic and functional responses. J. Immunol. 2019, 202, 3404–3411. [Google Scholar] [CrossRef]
- Magalhaes, I.; Pingris, K.; Poitou, C.; Bessoles, S.; Venteclef, N.; Kiaf, B.; Beaudoin, L.; Da Silva, J.; Allatif, O.; Rossjohn, J.; et al. Mucosal-associated invariant T cell alterations in obese and type 2 diabetic patients. J. Clin. Investig. 2015, 125, 1752–1762. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Simpson, R.J.; Lowder, T.W.; Spielmann, G.; Bigley, A.B.; LaVoy, E.C.; Kunz, H. Exercise and the aging immune system. Ageing Res. Rev. 2012, 11, 404–420. [Google Scholar] [CrossRef] [PubMed]
- Hanson, E.D.; Danson, E.; Nguyen-Robertson, C.V.; Fyfe, J.J.; Stepto, N.K.; Bartlett, D.B.; Sakkal, S. Maximal exercise increases mucosal associated invariant T cell frequency and number in healthy young men. Eur. J. Appl. Physiol. 2017, 117, 2159–2169. [Google Scholar] [CrossRef] [Green Version]
- Hanson, E.D.; Danson, E.; Evans, W.S.; Wood, W.A.; Battaglini, C.L.; Sakkal, S. Exercise Increases Mucosal-associated Invariant T Cell Cytokine Expression but Not Activation or Homing Markers. Med. Sci. Sports Exerc. 2019, 51, 379–388. [Google Scholar] [CrossRef] [PubMed]
- Graff, R.; Kunz, H.; Agha, N.; Baker, F.; Laughlin, M.; Bigley, A.; Markofski, M.; LaVoy, E.; Katsanis, E.; Bond, R. β2-Adrenergic receptor signaling mediates the preferential mobilization of differentiated subsets of CD8+ T-cells, NK-cells and non-classical monocytes in response to acute exercise in humans. Brain. Behav. Immun. 2018, 74, 143–153. [Google Scholar] [CrossRef]
- Hanson, E.D.; Bates, L.C.; Harrell, E.P.; Bartlett, D.B.; Lee, J.T.; Wagoner, C.W.; Alzer, M.S.; Amatuli, D.J.; Jensen, B.C.; Deal, A.M. Exercise training partially rescues impaired mucosal associated invariant t-cell mobilization in breast cancer survivors compared to healthy older women. Exp. Gerontol. 2021, 152, 111454. [Google Scholar] [CrossRef]
- Cardenas, M.A.; Levitt, M.M.; Richie, B.; Lu, S.; Cook, C.; Haynes, J.; Orr, K.; Kreutzer, A.; Mitchell, J.B.; Phillips, M.D. Exercise-induced Th17 Lymphocyte Response and their Rela-tionship to Cardiovascular Disease Risk Factors in Obese, Post-menopausal Women. Int. J. Exerc. Sci. Conf. Proc. 2018, 2, 13. [Google Scholar]
- Cannon, J.G.; Orencole, S.F.; Fielding, R.A.; Meydani, M.; Meydani, S.N.; Fiatarone, M.A.; Blumberg, J.B.; Evans, W.J. Acute phase response in exercise: Interaction of age and vitamin E on neutrophils and muscle enzyme release. Am. J. Physiol. Integr. Comp. Physiol. 1990, 259, R1214–R1219. [Google Scholar] [CrossRef]
- Cannon, J.G.; Fiatarone, M.A.; Fielding, R.A.; Evans, W.J. Aging and stress-induced changes in complement activation and neutrophil mobilization. J. Appl. Physiol. 1994, 76, 2616–2620. [Google Scholar] [CrossRef] [PubMed]
- Sellami, M.; Gasmi, M.; Denham, J.; Hayes, L.D.; Stratton, D.; Padulo, J.; Bragazzi, N. Effects of Acute and Chronic Exercise on Immunological Parameters in the Elderly Aged: Can Physical Activity Counteract the Effects of Aging? Front. Immunol. 2018, 9, 2187. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nieman, D.C.; Henson, D.A.; Nehlsen-Cannarella, S.L.; Ekkens, M.; Utter, A.C.; Butterworth, D.E.; Fagoaga, O.R. Influence of obesity on immune function. J. Am. Diet. Assoc. 1999, 99, 294–299. [Google Scholar] [CrossRef]
- Nieman, D.C.; Miller, A.R.; Henson, D.A.; Warren, B.J.; Gusewitch, G.; Johnson, R.L.; Davis, J.M.; Butterworth, D.E.; Herring, J.L.; Nehlsen-Cannarella, S.L. Effect of high- versus moderate-intensity exercise on lymphocyte subpopulations and proliferative response. Int. J. Sports Med. 1994, 15, 199–206. [Google Scholar] [CrossRef]
- Walsh, N.P.; Gleeson, M.; Shephard, R.J.; Jeffrey, M.G.; Woods, A.; Bishop, N.C.; Fleshner, M.; Green, C.; Pedersen, K.; Hoffman-goetz, L.; et al. Part one: Immune function and exercise. Exerc. Immunol. Rev. 2011, 17, 6–63. [Google Scholar]
- Krüger, K.; Lechtermann, A.; Fobker, M.; Völker, K.; Mooren, F.C. Exercise-induced redistribution of T lymphocytes is regulated by adrenergic mechanisms. Brain. Behav. Immun. 2008, 22, 324–338. [Google Scholar] [CrossRef]
- Peake, J.M.; Neubauer, O.; Walsh, N.P.; Simpson, R.J. Recovery of the immune system after exercise. J. Appl. Physiol. 2017, 122, 1077–1087. [Google Scholar] [CrossRef]
- Lee, O.-J.; Cho, Y.-N.; Kee, S.-J.; Kim, M.-J.; Jin, H.-M.; Lee, S.-J.; Park, K.-J.; Kim, T.-J.; Lee, S.-S.; Kwon, Y.-S.; et al. Circulating mucosal-associated invariant T cell levels and their cytokine levels in healthy adults. Exp. Gerontol. 2014, 49, 47–54. [Google Scholar] [CrossRef]
- Novak, J.; Dobrovolny, J.; Novakova, L.; Kozak, T. The Decrease in Number and Change in Phenotype of Mucosal-Associated Invariant T cells in the Elderly and Differences in Men and Women of Reproductive Age. Scand. J. Immunol. 2014, 80, 271–275. [Google Scholar] [CrossRef] [Green Version]
- Womack, J.; Tien, P.C.; Feldman, J.; Shin, J.H.; Fennie, K.; Anastos, K.; Cohen, M.H.; Bacon, M.C.; Minkoff, H. Obesity and immune cell counts in women. Metabolism 2007, 56, 998–1004. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- O’Rourke, R.W.; Kay, T.; Scholz, M.H.; Diggs, B.; Jobe, B.A.; Lewinsohn, D.M.; Bakke, A.C. Alterations in T-Cell Subset Frequency in Peripheral Blood in Obesity. Obes. Surg. 2005, 15, 1463–1468. [Google Scholar] [CrossRef] [PubMed]
- Tanaka, S.; Isoda, F.; Ishihara, Y.; Kimura, M.; Yamakawa, T. T lymphopaenia in relation to body mass index and TNF-α in human obesity: Adequate weight reduction can be corrective. Clin. Endocrinol. 2001, 54, 347–354. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Pre | Post | ||||||||
|---|---|---|---|---|---|---|---|---|---|
| OW (n = 16) | Lean (n = 8) | OW vs. Lean | OW (n = 16) | EX vs. ED | |||||
| EX | ED | Lean | MD | ES | EX | ED | MD | ES | |
| Age (y) | 61 (3) | 65 (4) | 64 (7) | 1 (4) | 0.20 | 61 (3) | 65 (4) | 4 (1) | 1.13 |
| BMI (kg/m2) | 33.1 (5) | 32.9 (2) | 21 (2) # | 12 (2) | 4.75 | 33.0 (5) | 33.5 (2) | 1 (3) | 0.13 |
| Height (cm) | 164 (4) | 161 (6) | 161 (5) | 2 (0) | 0.39 | 164 (4) | 162 (7) | 2 (3) | 0.35 |
| Weight (kg) | 89 (12) | 86 (13) | 56 (5) # | 32 (8) | 8.74 | 89 (12) | 88 (13) | 1 (1) | 0.08 |
| VO2 (mL/kg/min) | 22 (3) | 22 (4) | 30 (5) # | 8 (2) | 2.22 | 24 (4) | 22 (4) | 2 (0) | 0.50 |
| Outcome | Group | Base | 0 h | 1 h | Mob. MD | Mob. ES | Egress MD | Egress ES |
|---|---|---|---|---|---|---|---|---|
| Leukocytes (× 103/µL) | EX | 4.8 (1.7) | 7.3 (2.2) † | 6.7 (2.0) † | 2.5 (0.4) | 5.1 | −0.6 (0.1) | 2.0 |
| ED | 5.7 (0.9) | 6.3 (1.2) † | 6.7 (1.3) † | 0.6 (.3) | 0.4 (0.0) | |||
| Lymphocytes (× 103/µL) | EX | 1.5 (0.3) | 2.5 (0.6) † | 1.3 (0.3) | 1.0 (0.3) | 3.6 | −1.1 (0.3) | 5.4 |
| ED | 1.8 (0.5) | 1.9 (0.6) | 2.1 (0.6) | 0.2 (0.1) | 0.1 (0.0) | |||
| Neutrophils (× 103/µL) | EX | 2.7 (1.3) | 4.0 (1.7) † | 4.7 (1.8) † | 1.2 (0.3) | 3.1 | 0.7 (0.1) | 5.0 |
| ED | 3.2 (0.5) | 3.6 (0.6) † | 3.9 (0.7) † | 0.4 (0.2) | 0.2 (0.1) | |||
| Hemoglobin (g/dL) | EX | 12.9 (1.5) | 13.7 (1.5) | 13.0 (1.3) | 0.8 (0.2) | 1.9 | −0.7 (0.2) | 5.7 |
| ED | 12.9 (1.3) | 13.3 (1.3) | 13.5 (1.3) | 0.5 (0.1) | 0.2 (0.1) | |||
| Hematocrit (%) | EX | 38.1 (2.5) | 40.6 (3.3) † | 38.2 (2.2) | 2.2 (0.8) | 1.2 | −2.0 (1.1) | 2.9 |
| ED | 39.5 (3.0) | 39.2 (3.3) | 39.5 (1.9) | 1.4 (0.5) | 0.3 (1.0) | |||
| PV Shift (Δ%) | EX | - | −9.8 (4.3) † | −0.5 (4.6) | - | - | 9.3 (0.4) | 24.3 |
| ED | - | −5.6 (4.8) † | −7.3 (4.3) † | - | −1.7 (0.5) |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bates, L.C.; Hanson, E.D.; Levitt, M.M.; Richie, B.; Erickson, E.; Bartlett, D.B.; Phillips, M.D. Mucosal-Associated Invariant T Cell Response to Acute Exercise and Exercise Training in Older Obese Women. Sports 2021, 9, 133. https://0-doi-org.brum.beds.ac.uk/10.3390/sports9100133
Bates LC, Hanson ED, Levitt MM, Richie B, Erickson E, Bartlett DB, Phillips MD. Mucosal-Associated Invariant T Cell Response to Acute Exercise and Exercise Training in Older Obese Women. Sports. 2021; 9(10):133. https://0-doi-org.brum.beds.ac.uk/10.3390/sports9100133
Chicago/Turabian StyleBates, Lauren C., Erik D. Hanson, Michael M. Levitt, Bryan Richie, Elise Erickson, David B. Bartlett, and Melody D. Phillips. 2021. "Mucosal-Associated Invariant T Cell Response to Acute Exercise and Exercise Training in Older Obese Women" Sports 9, no. 10: 133. https://0-doi-org.brum.beds.ac.uk/10.3390/sports9100133