Microbial Strategies for Cellulase and Xylanase Production through Solid-State Fermentation of Digestate from Biowaste



Abstract
:1. Introduction
2. Materials and Methods
2.1. Substrate Characterization
2.2. Inoculum Preparation
2.3. Solid-State Fermentation (SSF)
2.3.1. Experimental Set-Up
2.3.2. Monitoring Parameters
2.3.3. Experimental Methodology
2.4. Analytical Methods
2.4.1. Enzyme Extraction
2.4.2. Enzymatic Activity Determination
2.4.3. Routine Methods
2.4.4. Statistics
3. Results and Discussion
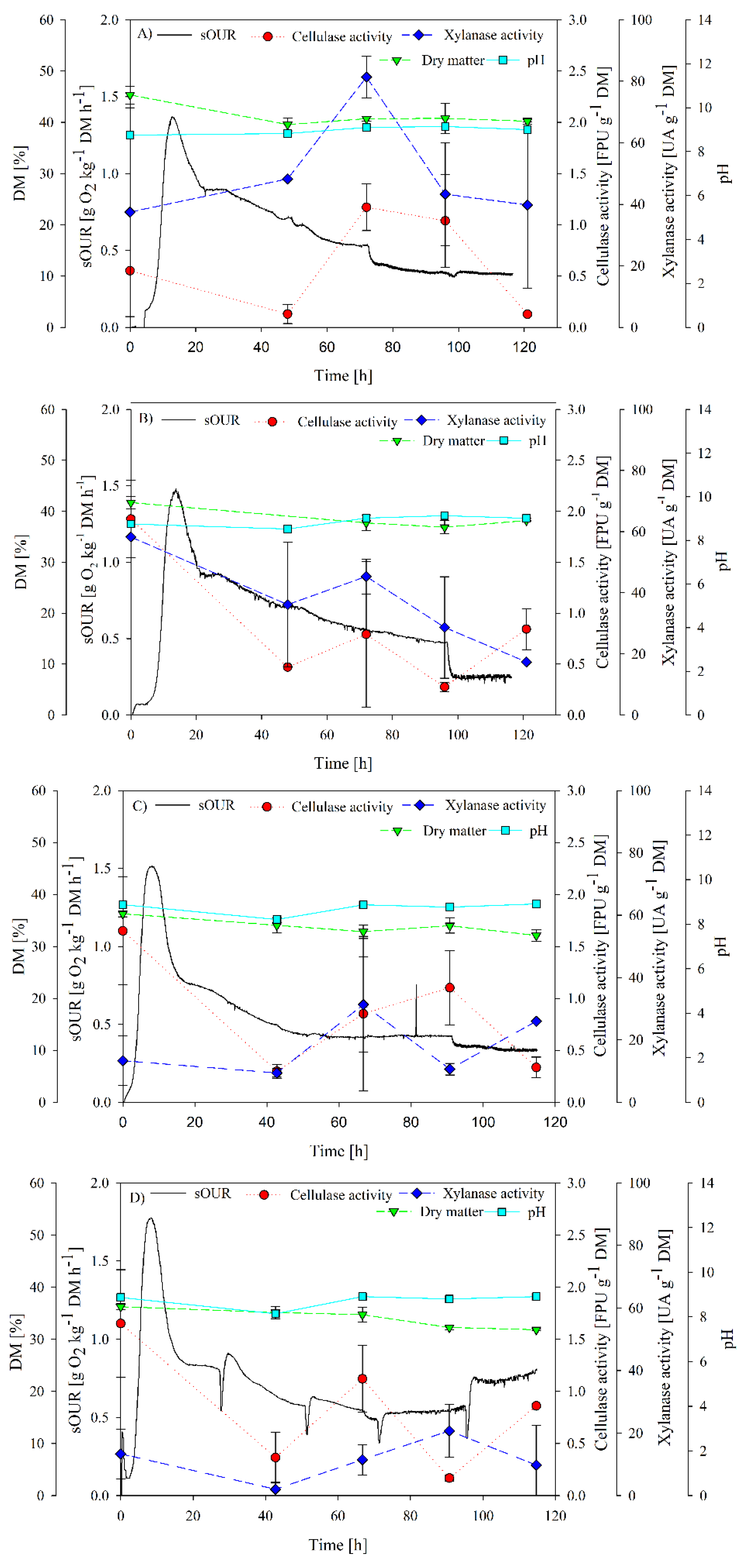
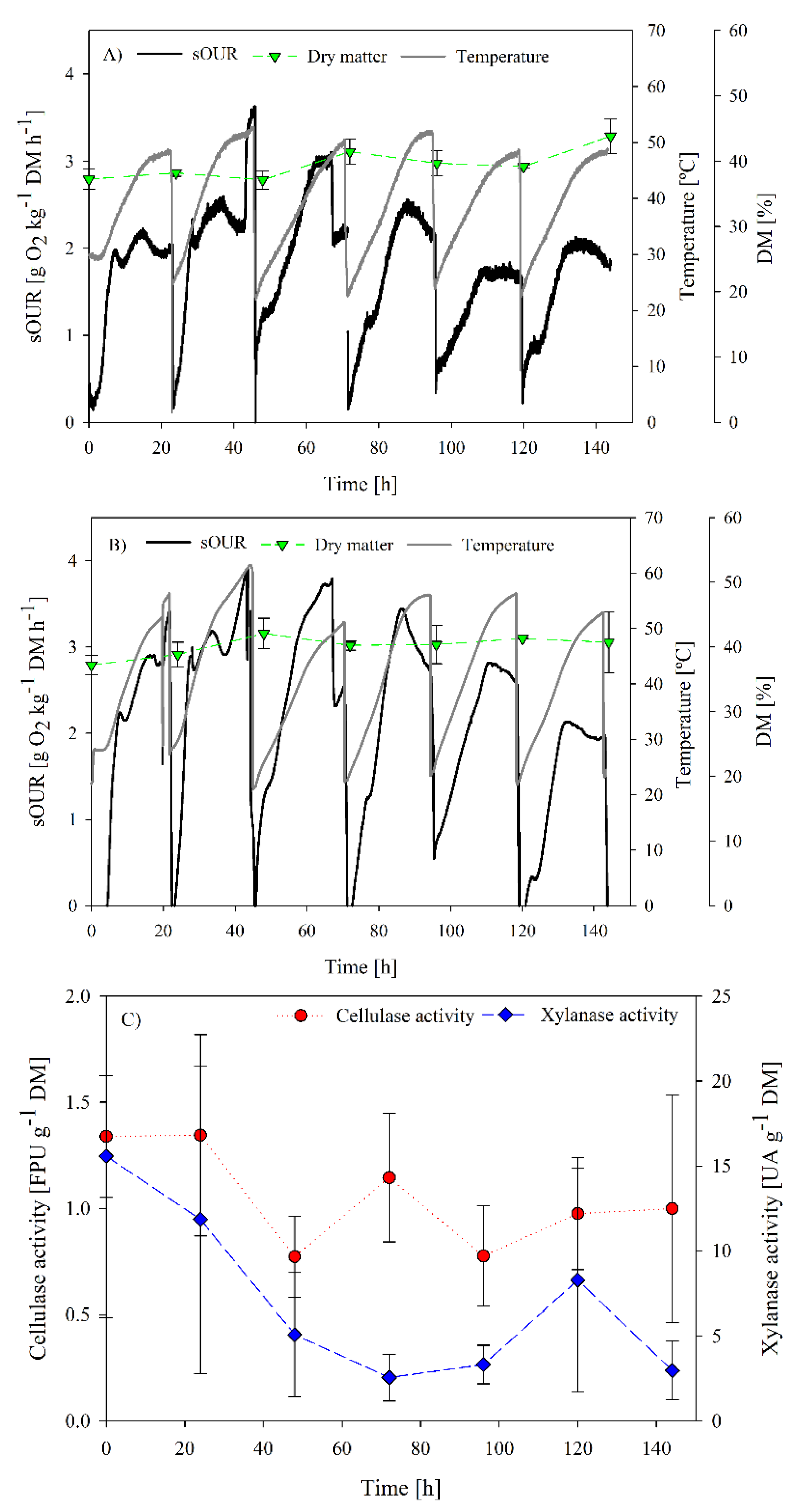
3.1. Inoculation Requirements Determination
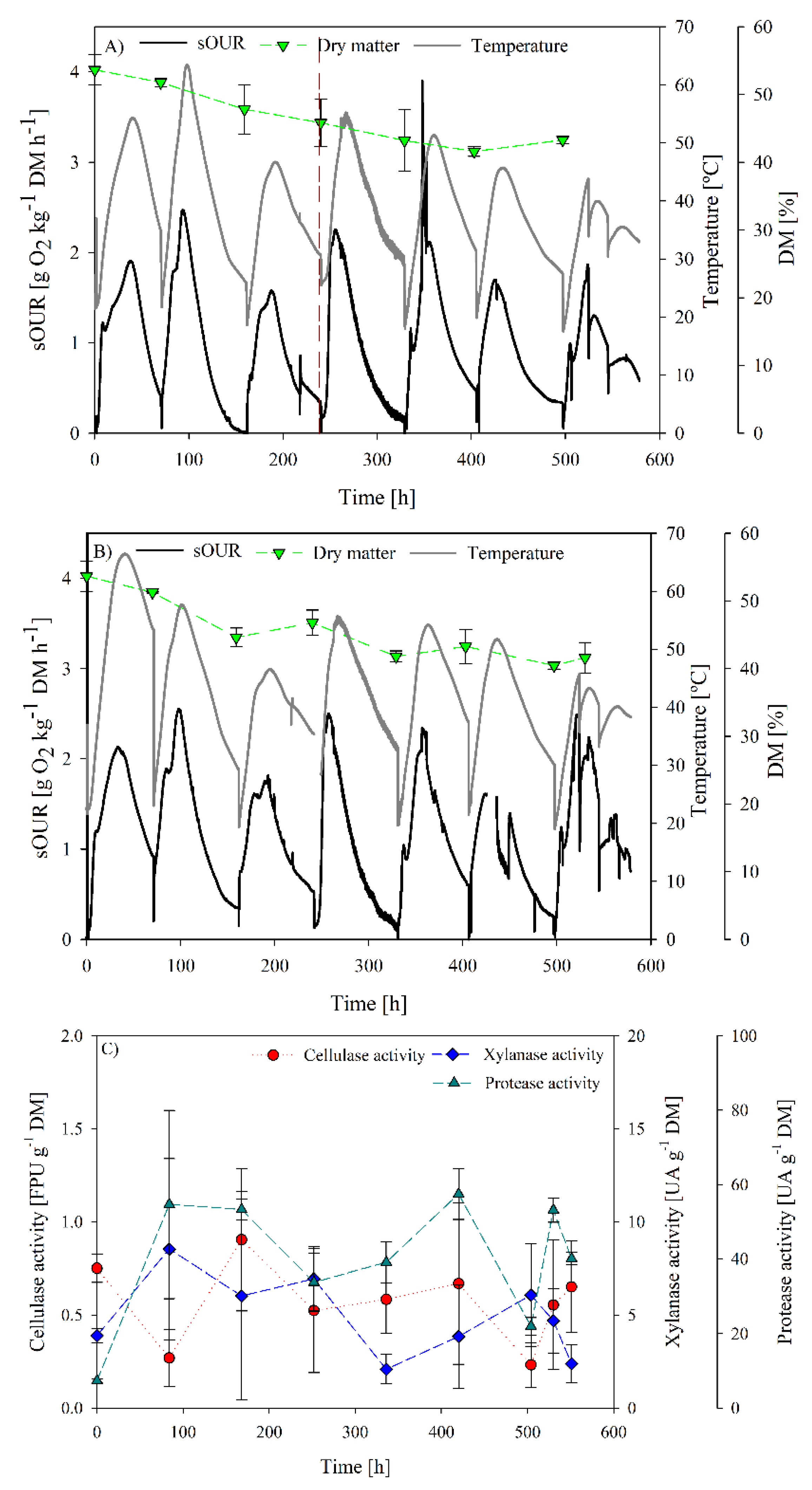
3.2. SBR as Strategy to Obtain a Specialized Inoculum for Cellulase and Xylanase Production
4. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Biodegradable Waste-Environment-European Commission. Available online: http://ec.europa.eu/environment/waste/compost/index.htm (accessed on 26 September 2017).
- Tambone, F.; Scaglia, B.; D’Imporzano, G.; Schievano, A.; Orzi, V.; Salati, S.; Adani, F. Assessing amendment and fertilizing properties of digestates from anaerobic digestion through a comparative study with digested sludge and compost. Chemosphere 2010, 81, 577–583. [Google Scholar] [CrossRef] [PubMed]
- Maynaud, G.; Druilhe, C.; Daumoin, M.; Jimenez, J.; Patureau, D.; Torrijos, M.; Pourcher, A.M.; Wéry, N. Characterisation of the biodegradability of post-treated digestates via the chemical accessibility and complexity of organic matter. Bioresour. Technol. 2017, 231, 65–74. [Google Scholar] [CrossRef] [PubMed]
- Costa, A.; Ely, C.; Pennington, M.; Rock, S.; Staniec, C.; Turgeon, J. Anaerobic Digestion and Its Applications; U.S. Environmental Protection Agency: Washington, DC, USA, 2015; p. 15.
- Teglia, C.; Tremier, A.; Martel, J.L. Characterization of solid digestates: Part 1, review of existing indicators to assess solid digestates agricultural use. Waste Biomass Valorization 2011, 2, 43–58. [Google Scholar] [CrossRef]
- Singhania, R.R.; Sukumaran, R.K.; Patel, A.K.; Larroche, C.; Pandey, A. Advancement and comparative profiles in the production technologies using solid-state and submerged fermentation for microbial cellulases. Enzyme Microb. Technol. 2010, 46, 541–549. [Google Scholar] [CrossRef]
- Lizardi-Jiménez, M.A.; Hernández-Martínez, R. Solid-state fermentation (SSF): Diversity of applications to valorize waste and biomass. 3 Biotech 2017, 7. [Google Scholar] [CrossRef] [PubMed]
- Yazid, N.A.; Barrena, R.; Komilis, D.; Sánchez, A. Solid-State Fermentation as a Novel Paradigm for Organic Waste Valorization: A Review. Sustainability 2017, 9, 224. [Google Scholar] [CrossRef]
- Wilson, D.B. Cellulases and biofuels. Curr. Opin. Biotechnol. 2009, 20, 295–299. [Google Scholar] [CrossRef] [PubMed]
- Srivastava, N.; Srivastava, M.; Mishra, P.K.; Gupta, V.K.; Molina, G.; Rodriguez-Couto, S.; Manikanta, A.; Ramteke, P.W. Applications of fungal cellulases in biofuel production: Advances and limitations. Renew. Sustain. Energy Rev. 2017, 2379–2386. [Google Scholar] [CrossRef]
- Hart, T.D.; De Leij, F.A.A.M.; Kinsey, G.; Kelley, J.; Lynch, J.M. Strategies for the isolation of cellulolytic fungi for composting of wheat straw. World J. Microbiol. Biotechnol. 2002, 18, 471–480. [Google Scholar] [CrossRef]
- Abraham, J.; Gea, T.; Sánchez, A. Potential of the SSF of soy fibers residues by native microbial populations for bench-scale alkaline protease production. Biochem. Eng. J. 2013, 74, 15–19. [Google Scholar] [CrossRef]
- Cerda, A.; Gea, T.; Vargas-García, M.C.; Sánchez, A. Towards a competitive solid-state fermentation: Cellulases production from coffee husk by sequential batch operation and role of microbial diversity. Sci. Total Environ. 2017, 589, 56–65. [Google Scholar] [CrossRef] [PubMed]
- Brijwani, K.; Oberoi, H.S.; Vadlani, P.V. Production of a cellulolytic enzyme system in mixed-culture solid-state fermentation of soybean hulls supplemented with wheat bran. Process Biochem. 2010, 45, 120–128. [Google Scholar] [CrossRef]
- Sukumaran, R.K.; Singhania, R.R.; Mathew, G.M.; Pandey, A. Cellulase production using biomass feed stock and its application in lignocellulose saccharification for bio-ethanol production. Renew. Energy 2009, 34, 421–424. [Google Scholar] [CrossRef]
- Xia, L.; Cen, P. Cellulase production by solid-state fermentation on lignocellulosic waste from the xylose industry. Process Biochem. 1999, 34, 909–912. [Google Scholar] [CrossRef]
- Brijwani, K.; Vadlani, P.V.; Hohn, K.L.; Maier, D.E. Experimental and theoretical analysis of a novel deep-bed solid-state bioreactor for cellulolytic enzymes production. Biochem. Eng. J. 2011, 58–59, 110–123. [Google Scholar] [CrossRef]
- Dhillon, G.S.; Oberoi, H.S.; Kaur, S.; Bansal, S.; Brar, S.K. Value-addition of agricultural wastes for augmented cellulase and xylanase production through solid-state tray fermentation employing mixed-culture of fungi. Ind. Crops Prod. 2011, 34, 1160–1167. [Google Scholar] [CrossRef]
- Santi, G.; Muzzini, V.G.; Galli, E.; Proietti, S.; Moscatello, S.; Battistelli, A. Mycelial growth and enzymatic activities of white-rot fungi on anaerobic digestates from industrial biogas plants. Environ. Eng. Manag. J. 2015, 14, 1713–1719. [Google Scholar]
- European Parliament and Council Regulation (EC) No 1069/2009; Official Journal of the European Union: Brussels, Belgium, 2009; Volume 300, pp. 1–33.
- Ponsá, S.; Gea, T.; Sánchez, A. Different Indices to Express Biodegradability in Organic Solid Wastes. J. Environ. Quality 2010, 39, 706. [Google Scholar] [CrossRef] [PubMed]
- Gea, T.; Barrena, R.; Artola, A.; Sánchez, A. Optimal bulking agent particle size and usage for heat retention and disinfection in domestic wastewater sludge composting. Waste Manag. 2007, 27, 1108–1116. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- López-González, J.A.; del Vargas-García, M.C.; López, M.J.; Suárez-Estrella, F.; Jurado, M.; Moreno, J. Enzymatic characterization of microbial isolates from lignocellulose waste composting: Chronological evolution. J. Environ. Manag. 2014, 145, 137–146. [Google Scholar] [CrossRef] [PubMed]
- Eichorst, S.A.; Varanasi, P.; Stavila, V.; Zemla, M.; Auer, M.; Singh, S.; Simmons, B.A.; Singer, S.W. Community dynamics of cellulose-adapted thermophilic bacterial consortia. Environ. Microbiol. 2013, 15, 2573–2587. [Google Scholar] [CrossRef] [PubMed]
- Dhillon, G.S.; Kaur, S.; Brar, S.K.; Verma, M. Potential of apple pomace as a solid substrate for fungal cellulase and hemicellulase bioproduction through solid-state fermentation. Ind. Crops Prod. 2012, 38, 6–13. [Google Scholar] [CrossRef]
- Ghose, T.K. Measurement of cellulase activities. Pure Appl. Chem. 1987, 59. [Google Scholar] [CrossRef]
- Ang, S.K.; Shaza, E.M.; Adibah, Y.A.; Suraini, A.A.; Madihah, M.S. Production of cellulases and xylanase by Aspergillus fumigatus SK1 using untreated oil palm trunk through solid-state fermentation. Process Biochem. 2013, 48, 1293–1302. [Google Scholar] [CrossRef]
- Alef, K.; Nannipieri, P. Enzyme Activities; Academic Press: London, UK, 1995; ISBN 9780125138406. [Google Scholar]
- Test Methods US Composting Council. Available online: https://compostingcouncil.org/test-methods-parameters/ (accessed on 5 March 2018).
- Van Soest, P.J.; Robertson, J.B.; Lewis, B.A. Methods for Dietary Fiber, Neutral Detergent Fiber, and Nonstarch Polysaccharides In Relation To Animal Nutrition. J. Dairy Sci. 1991, 74, 3583–3597. [Google Scholar] [CrossRef]
- Barrena, R.; Gea, T.; Ponsá, S.; Ruggieri, L.; Artola, A.; Font, X.; Sánchez, A. Categorizing Raw Organic Material Biodegradability Via Respiration Activity Measurement: A Review. Compost Sci. Util. 2011, 19, 105–113. [Google Scholar] [CrossRef] [Green Version]
- Yang, F.; Hanna, M.; Sun, R. Value-added uses for crude glycerol—A byproduct of biodiesel production. Biotechnol. Biofuels 2012, 5, 13. [Google Scholar] [CrossRef] [PubMed]
- Flodman, H.R.; Noureddini, H. Effects of intermittent mechanical mixing on solid-state fermentation of wet corn distillers grain with Trichoderma reesei. Biochem. Eng. J. 2013, 81, 24–28. [Google Scholar] [CrossRef]
- Deschamps, F.; Giuliano, C.; Asther, M.; Huet, M.C.; Roussost, S. Cellulase production by Trichoderma harzianum in static and mixed solid-state fermentation reactors under mesophilic conditions. Biotechnol. Bioeng. 1985, 27, 1385–1388. [Google Scholar] [CrossRef] [PubMed]
- Mitchell, D.A.; Krieger, N.; Berovič, M. Solid-State Fermentation Bioreactors; Springer: Berlin, Germany, 2006; ISBN 978-3-540-31285-7. [Google Scholar]
- Lever, M.; Ho, G.; Cord-Ruwisch, R. Ethanol from lignocellulose using crude unprocessed cellulase from solid-state fermentation. Bioresour. Technol. 2010, 101, 7083–7087. [Google Scholar] [CrossRef] [PubMed]
- Gao, J.; Weng, H.; Zhu, D.; Yuan, M.; Guan, F.; Xi, Y. Production and characterization of cellulolytic enzymes from the thermoacidophilic fungal Aspergillus terreus M11 under solid-state cultivation of corn stover. Bioresour. Technol. 2008, 99, 7623–7629. [Google Scholar] [CrossRef] [PubMed]
- Liu, D.; Zhang, R.; Yang, X.; Wu, H.; Xu, D.; Tang, Z.; Shen, Q. Thermostable cellulase production of Aspergillus fumigatus Z5 under solid-state fermentation and its application in degradation of agricultural wastes. Int. Biodeterior. Biodegrad. 2011, 65, 717–725. [Google Scholar] [CrossRef]
- Chapla, D.; Divecha, J.; Madamwar, D.; Shah, A. Utilization of agro-industrial waste for xylanase production by Aspergillus foetidus MTCC 4898 under solid-state fermentation and its application in saccharification. Biochem. Eng. J. 2010, 49, 361–369. [Google Scholar] [CrossRef]
- Nava-Cruz, N.Y.; Contreras-Esquivel, J.C.; Aguilar-González, M.A.; Nuncio, A.; Rodríguez-Herrera, R.; Aguilar, C.N. Agave atrovirens fibers as substrate and support for solid-state fermentation for cellulase production by Trichoderma asperellum. 3 Biotech 2016, 6. [Google Scholar] [CrossRef] [PubMed]
- Loman, A.A.; Ju, L.K. Optimization of Enzymatic Process Condition for Protein Enrichment, Sugar Recovery and Digestibility Improvement of Soy Flour. JAOCS J. Am. Oil Chem. Soc. 2016, 93, 1063–1073. [Google Scholar] [CrossRef]
- Ortiz, G.E.; Guitart, M.E.; Cavalitto, S.F.; Albertó, E.O.; Fernández-Lahore, M.; Blasco, M. Characterization, optimization, and scale-up of cellulases production by Trichoderma reesei cbs 836.91 in solid-state fermentation using agro-industrial products. Bioprocess Biosyst. Eng. 2015, 38, 2117–2128. [Google Scholar] [CrossRef] [PubMed]
- Cerda, A.; Mejías, L.; Gea, T.; Sánchez, A. Cellulase and xylanase production at pilot scale by solid-state fermentation from coffee husk using specialized consortia: The consistency of the process and the microbial communities involved. Bioresour. Technol. 2017, 243, 1059–1068. [Google Scholar] [CrossRef] [PubMed]
- Cheirsilp, B.; Kitcha, S. Solid-state fermentation by cellulolytic oleaginous fungi for direct conversion of lignocellulosic biomass into lipids: Fed-batch and repeated-batch fermentations. Ind. Crops Prod. 2015, 66, 73–80. [Google Scholar] [CrossRef]
- Gao, D.; Haarmeyer, C.; Balan, V.; Whitehead, T.A.; Dale, B.E.; Chundawat, S.P.S. Lignin triggers irreversible cellulase loss during pretreated lignocellulosic biomass saccharification. Biotechnol. Biofuels 2014, 7, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Marín, M.; Artola, A.; Sánchez, A. Optimization of Down-Stream for Cellulases Produced Under Solid-State Fermentation of Coffee Husk. Waste Biomass Valoriz. 2018. [Google Scholar] [CrossRef]
- Cerda, A.; El-Bakry, M.; Gea, T.; Sánchez, A. Long term enhanced solid-state fermentation: Inoculation strategies for amylase production from soy and bread wastes by Thermomyces sp. in a sequential batch operation. J. Environ. Chem. Eng. 2016, 4, 2394–2401. [Google Scholar] [CrossRef]
- El-Bakry, M.; Abraham, J.; Cerda, A.; Barrena, R.; Ponsá, S.; Gea, T.; Sánchez, A. From Wastes to High Value Added Products: Novel Aspects of SSF in the Production of Enzymes. Crit. Rev. Environ. Sci. Technol. 2015, 45, 1999–2042. [Google Scholar] [CrossRef]
- El-Bakry, M.; Gea, T.; Sánchez, A. Inoculation effect of thermophilic microorganisms on protease production through solid-state fermentation under non-sterile conditions at lab and bench scale (SSF). Bioprocess Biosyst. Eng. 2016, 39, 585–592. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Reference | Enzyme | Microorganism | Substrate | SSF Time | Temperature | pH | Fermentation Sample | Max. Enzymatic Activity |
|---|---|---|---|---|---|---|---|---|
| [15] | Cellulase | Trichoderma reesei RUT C30 | Wheat bran | 72 h | 30 °C | - | 5 g | 22.8 FPU/g DS |
| Betaglucosidase | Aspergillus niger MTCC 7956 | Wheat bran | 5 g | 21.39 U/g DS | ||||
| [37] | Endoglucanase | Aspergillus terreus M11 | Wheat straw | 96 h | 45 °C | - | 5 g (plus mineral solution) | 417 U/g DM |
| Cellulase | 166 U/g DM | |||||||
| Beta-glucanase | 87 U/g DM | |||||||
| [33] | Cellulase | Trichoderma reesei (ATCC 26921) Aspergillus oryzae (ATCC 12892) | Soybean hulls and wheat bran | 96 h | 30 °C | 5 | 100 g | 10.78 FPU/ g DS |
| Betaglucosidase | 10.71 U/g DS | |||||||
| Endoglucanase | 100.67 U/g DS | |||||||
| Xylanases | 504.98 U/g DS | |||||||
| [16] | Cellulase | Trichoderma reesei ZU-02 | Corn cob residue and wheat bran + mineral solution | 5 days | 30 °C | 4.5 (initial) | Trays (thickness about 30 cm) | 40–158 FPU/g (depending on fermentation time) |
| [38] | Cellulase | Aspergillus fumigatus Z5 | Cornmeal | 4 days | 50 °C | 9 | Substrate (plus mineral medium) | 98.1 FPU/g dw |
| Endoglucanase | 341.3 U/g dw | |||||||
| [39] | Xylanase | Aspergillus foetidus MTCC 4898 | Agricultural waste + wastewater from anaerobic digestion | 72–96 h | 30 °C | - | 5 g (plus liquid medium) | 100–40,000 U/g (depending on the agricultural residue) |
| [40] | Endoglucanase | Trichoderma asperellum | Agave atrovirens fibres | 210–310 h | 29 °C | - | - | 12,860 U/g |
| Exoglucanase | 3144.4 U/g | |||||||
| Betaglucosidase | 384.4 U/g | |||||||
| [19] | Endoglucanase | Pleurotus ostreatus | Corn silage digestate | 15–20 days | - | - | 15 g | 2.3 U/mg prot |
| Cellobiohydrolase | 0.7 U/mg prot | |||||||
| Xylanase | 3 U/mg prot | |||||||
| [17] | Cellulase | Trichoderma reesei Aspergillus oryzae | Soybean hull | 70 h | 30 | 5 | 10.24 kg | 5.4 FPU/g |
| Endoglucanse | 58.6 U/g | |||||||
| Betaglucosidase | 18.4 U/g | |||||||
| Xylanase | 242 U/g | |||||||
| [18] | Cellulase | Trichoderma reesei Aspergillus oryzae | Rice straw and wheat bran | 120 h | 30 | - | 500 g | 35.8 FPU/g |
| Endoglucanse | 132.3 U/g | |||||||
| Betaglucosidase | 33.7 U/g | |||||||
| Xylanase | 3106 U/g |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mejias, L.; Cerda, A.; Barrena, R.; Gea, T.; Sánchez, A. Microbial Strategies for Cellulase and Xylanase Production through Solid-State Fermentation of Digestate from Biowaste. Sustainability 2018, 10, 2433. https://0-doi-org.brum.beds.ac.uk/10.3390/su10072433
Mejias L, Cerda A, Barrena R, Gea T, Sánchez A. Microbial Strategies for Cellulase and Xylanase Production through Solid-State Fermentation of Digestate from Biowaste. Sustainability. 2018; 10(7):2433. https://0-doi-org.brum.beds.ac.uk/10.3390/su10072433
Chicago/Turabian StyleMejias, Laura, Alejandra Cerda, Raquel Barrena, Teresa Gea, and Antoni Sánchez. 2018. "Microbial Strategies for Cellulase and Xylanase Production through Solid-State Fermentation of Digestate from Biowaste" Sustainability 10, no. 7: 2433. https://0-doi-org.brum.beds.ac.uk/10.3390/su10072433