Actinomycete Strains Isolated from Saline Soils: Plant-Growth-Promoting Traits and Inoculation Effects on Solanum lycopersicum

 ,
,  ,
,  and
and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Actinomycete Strains and Culture Conditions
2.2. Plant-Growth-Promoting Traits
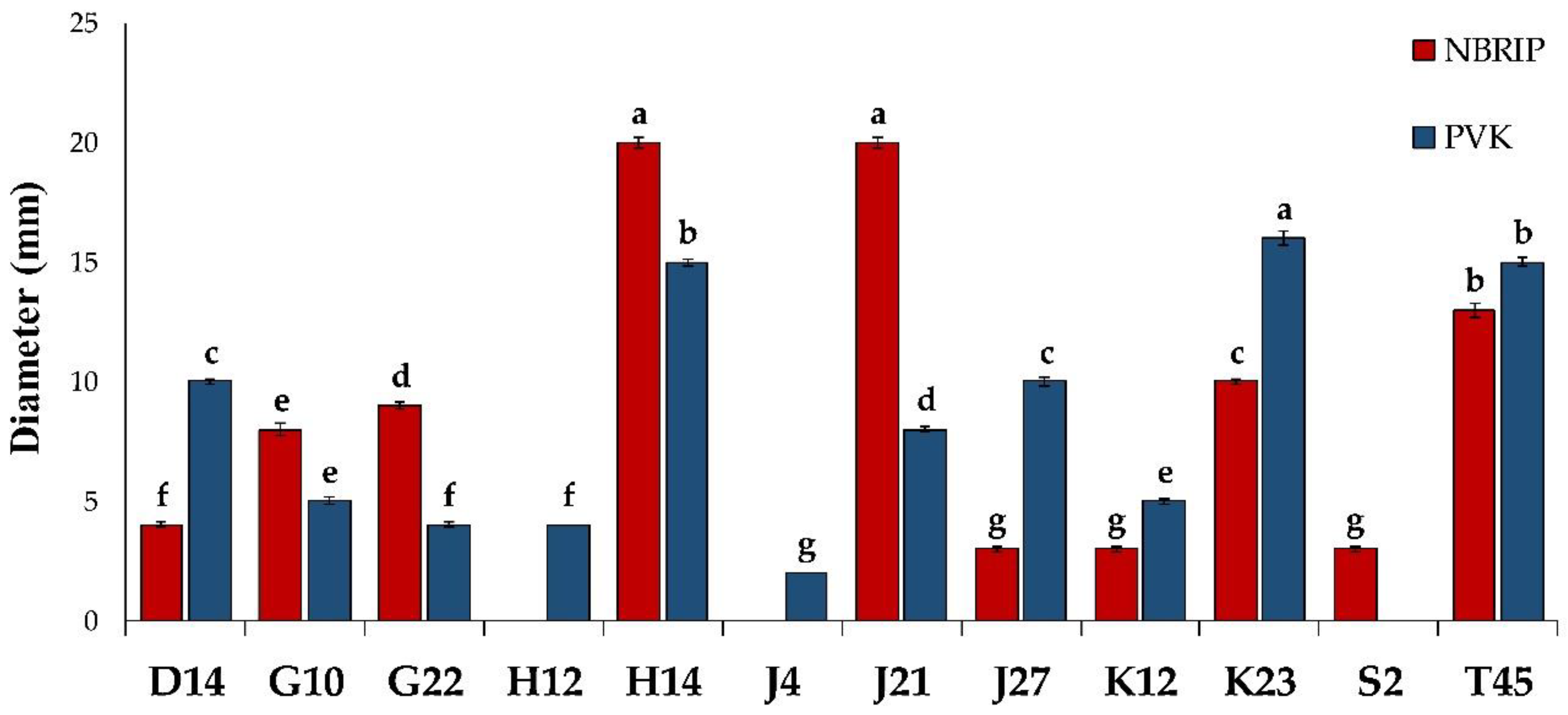
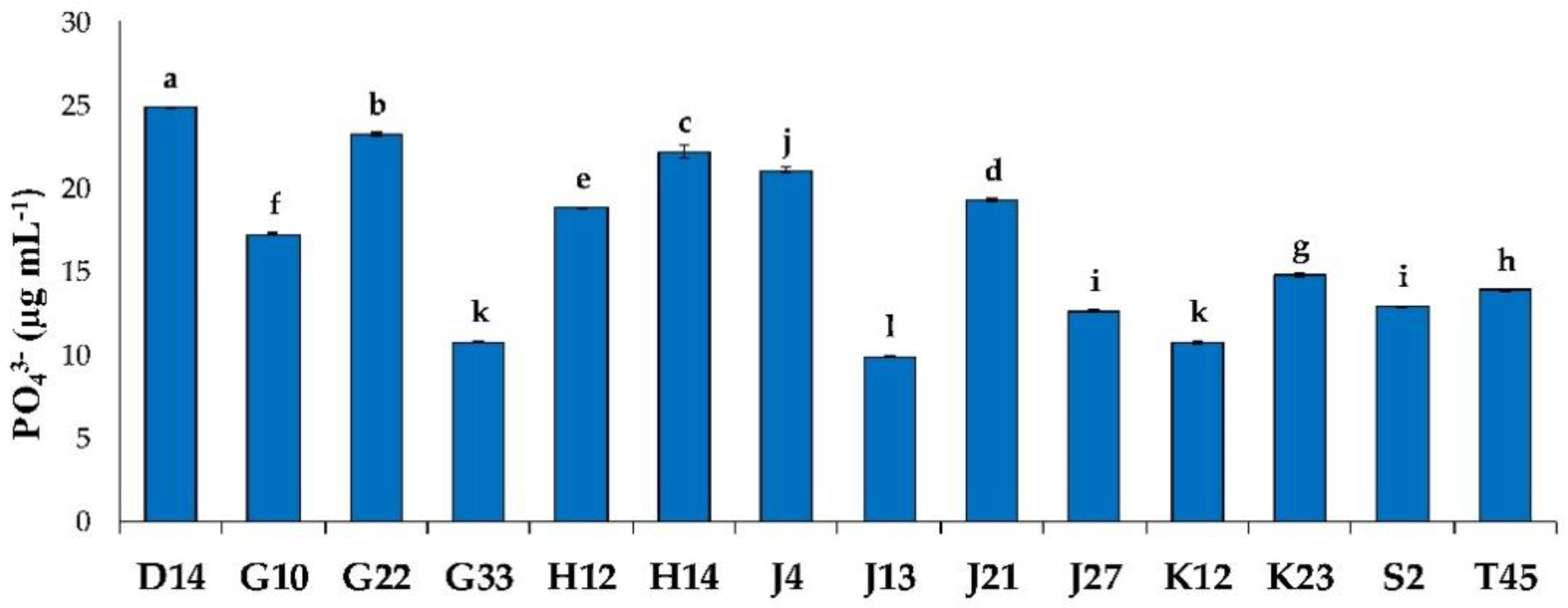
2.2.1. Phosphate Solubilization
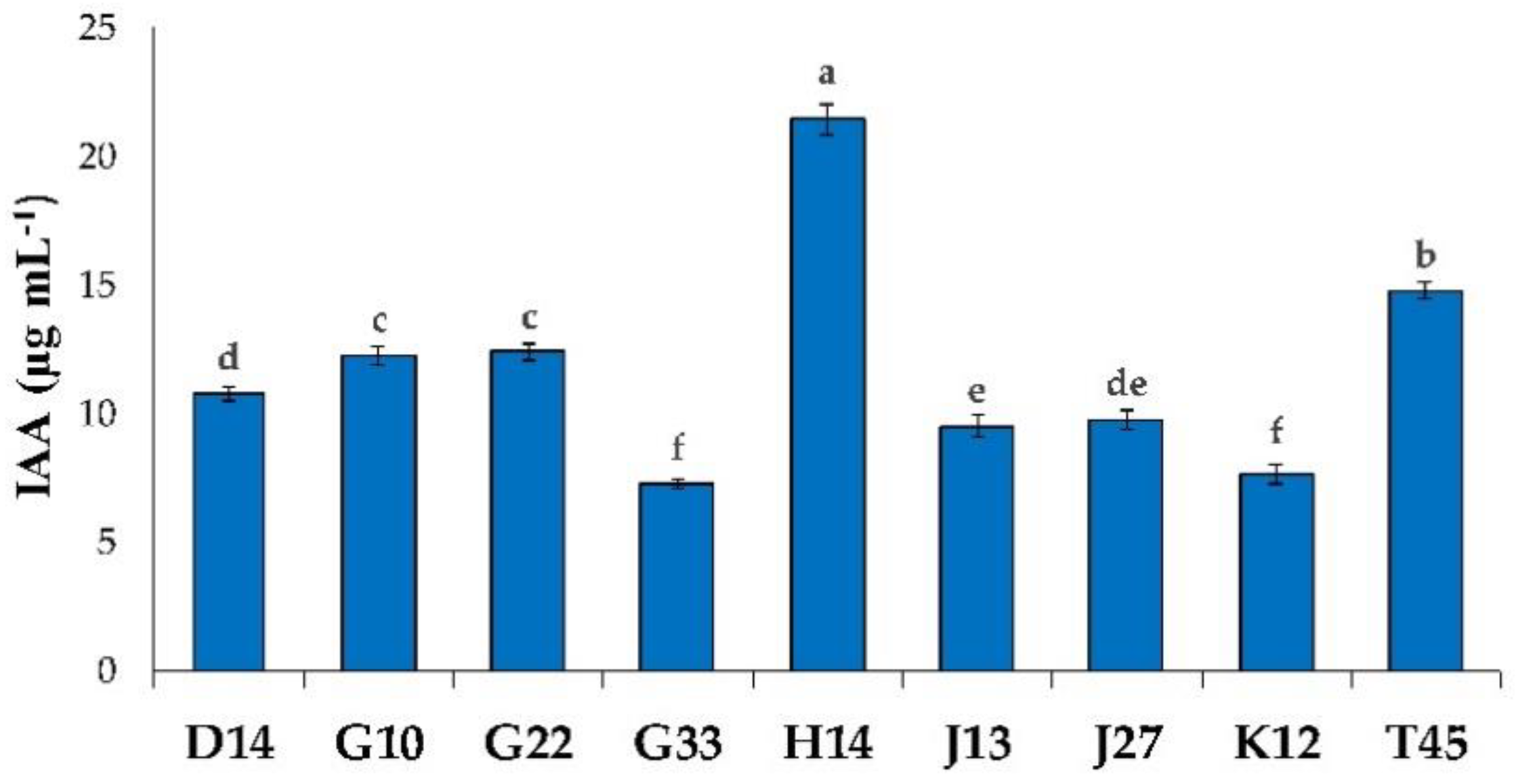
2.2.2. Production of Indole-3-Acetic Acid
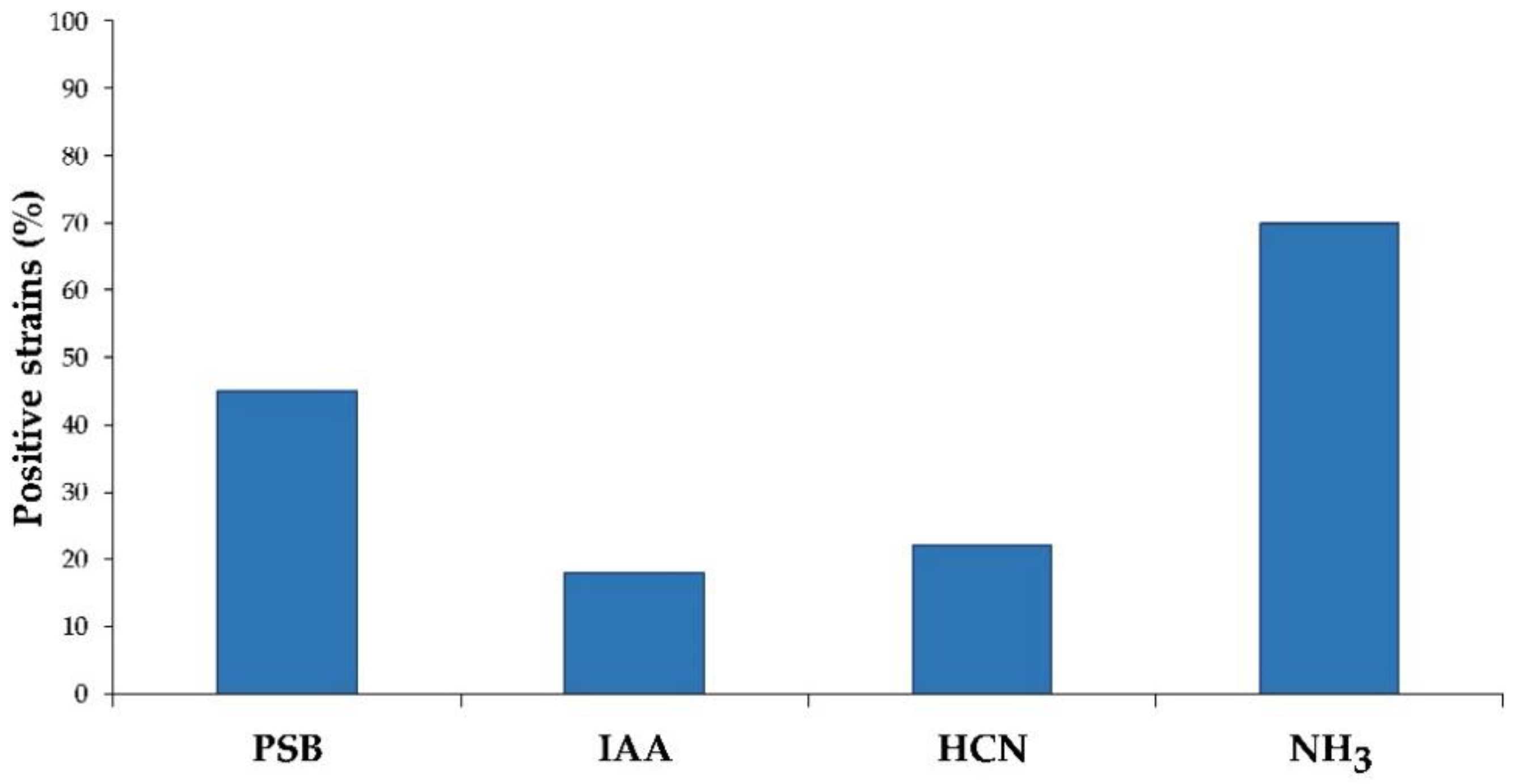
2.2.3. Production of Hydrocyanic Acid and Ammonia and Enzymatic Activities
- Amylase: Strains were streaked on the Glycerol Bouillon Agar (GBA) medium (20 g L−1 soluble starch, 20 mL L−1 glycerol, 10 g L−1 peptone, 5 g L−1 meat extract, 3 g L−1 CaCO3, 15 g L−1 agar—pH 7.0 ± 0.2). The cultures were incubated at 30 °C for 7 days, and amylase production was detected after immersion of the dishes with a Lugol solution. A positive outcome was underlined by the presence of a clear discoloration zone around the colonies [34].
- Catalase: An isolated colony from ISP2 agar culture was placed on a slide and a few drops of hydrogen peroxide were deposited on the colony. A positive catalase reaction results from the appearance of bubbles after the release of O2 [37].
- Cellulase: Strains were streaked on the ISP2 medium supplemented with 1% of cellulose. After incubation for 7 days at 30 °C, dishes were immersed in a Congo red solution (1% w/v) for 20 min and then washed with a NaCl solution (1N). A positive reaction is indicated by the development of a clear discoloration zone around the streak [38].
- Chitinase: Strains were streaked on nutrient agar supplemented with 1% chitin. After 7 days of incubation at 30 °C, the appearance of clear areas around the colonies was considered as a sign of chitinase production.
- Gelatinase: Strains were inoculated in tubes containing nutritive gelatin medium. After an incubation for 21 days at 30 °C, tubes were placed in the refrigerator for 1 h. If the gelatin becomes solid, this means that it was not attacked; if it remains liquid, this means that an extracellular enzyme has hydrolyzed it [39].
- Lecithinase, lipase, and lipoproteinase: Strains were streaked on agar medium containing egg yolk and prepared following the method described by Dellaras [39]. After plate incubation at 30 °C for 7 days, the presence of the enzymes was assessed as follows: (i) Lecithinase: Appearance of an opaque halo of white-yellowish pearl, with a clear edge under the streak or at the borders; (ii) Lipase: Appearance of an oily and shiny pearly white halo; (iii) Lipoproteinase: Appearance of a clear halo around the streak [40].
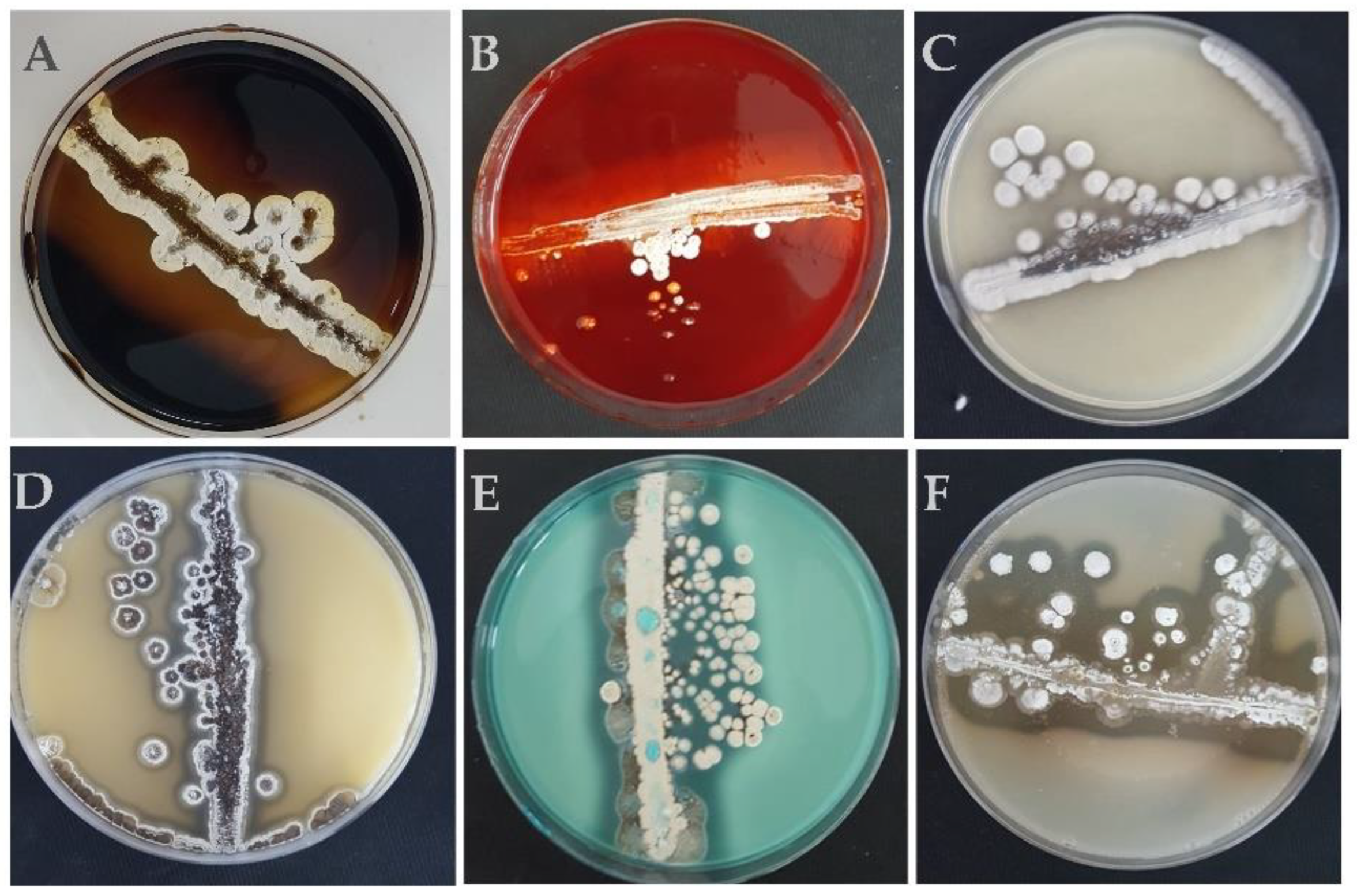
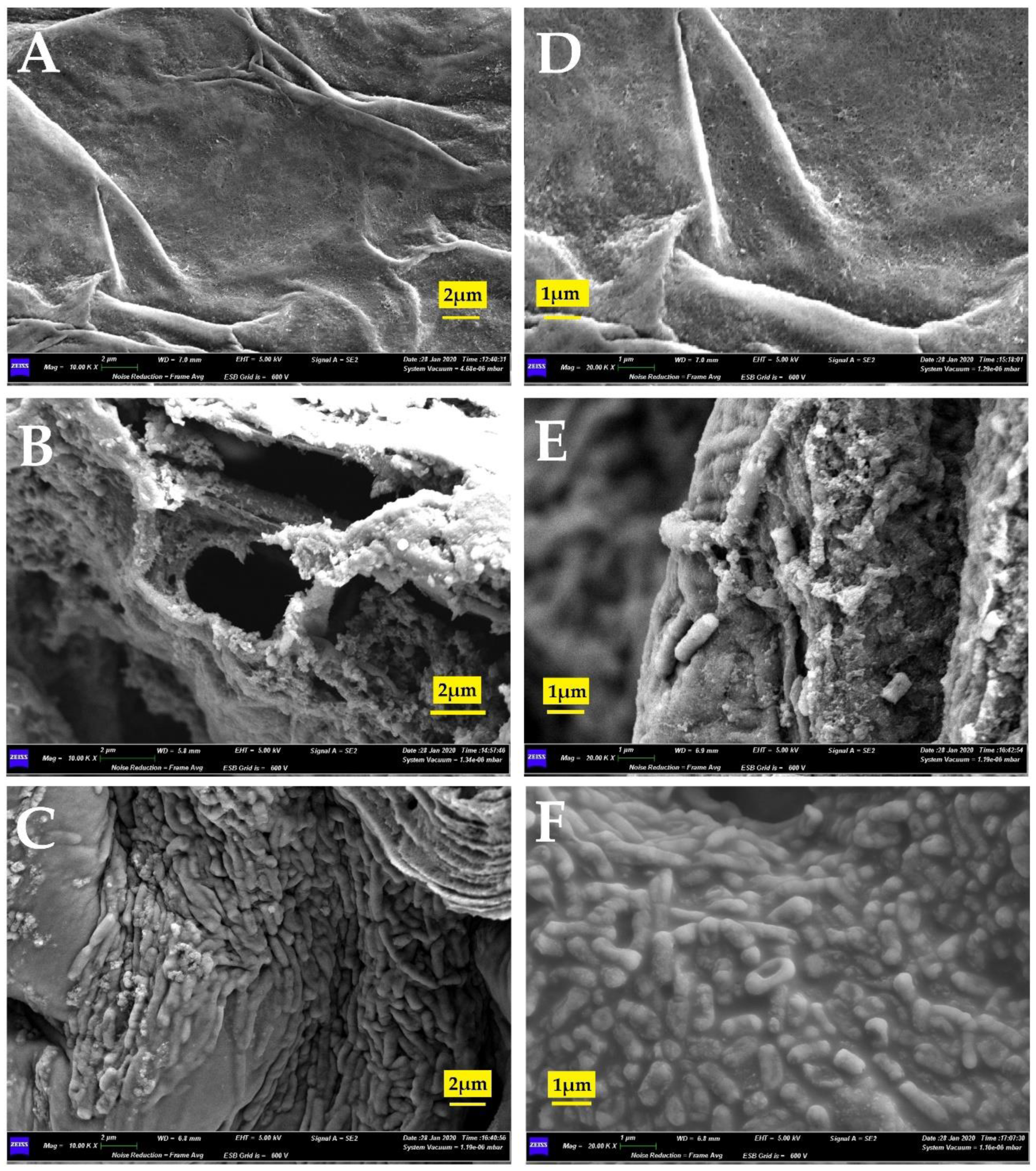
2.3. In Vitro Association Study by Scanning Electron Microscopy
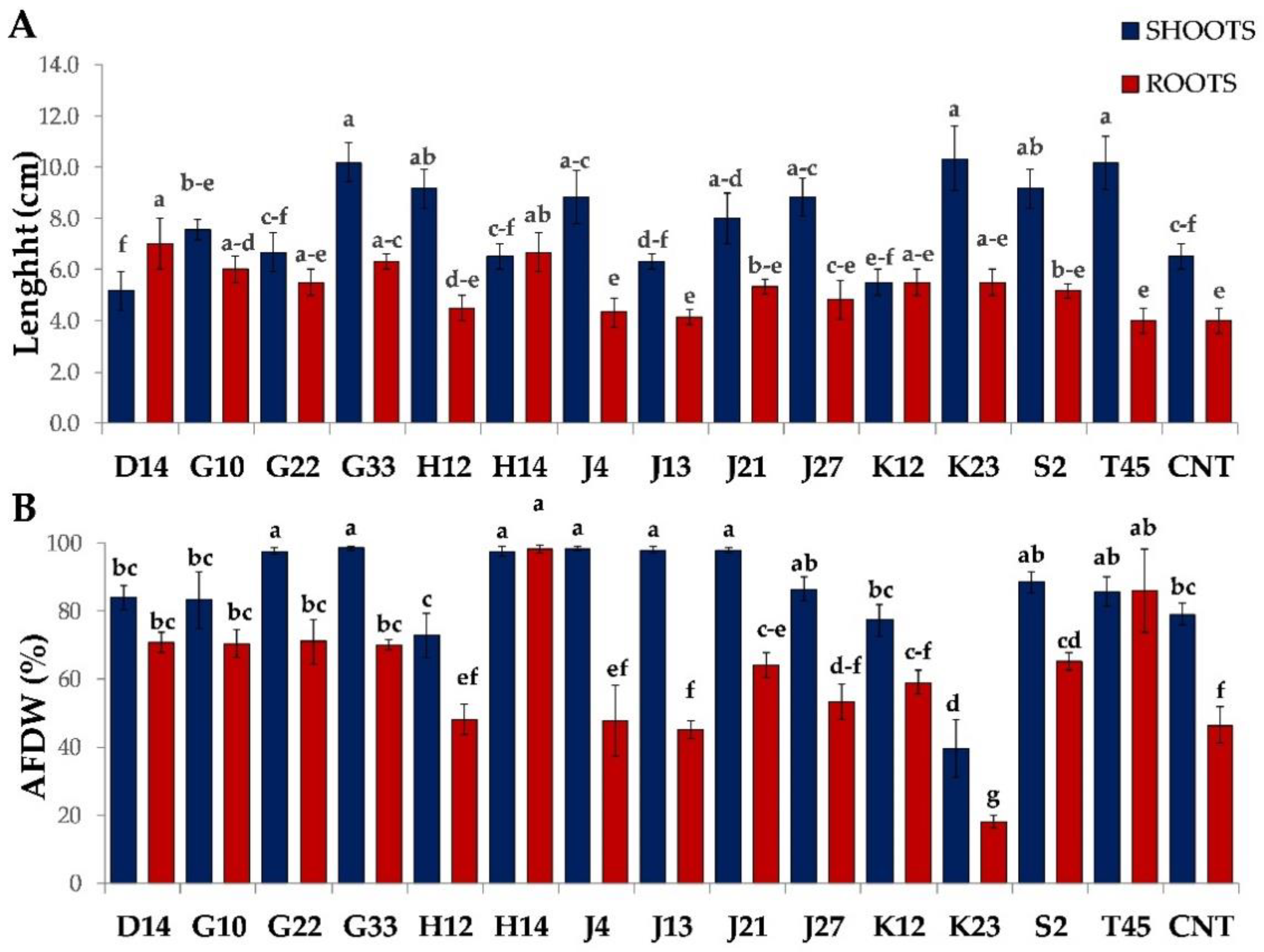
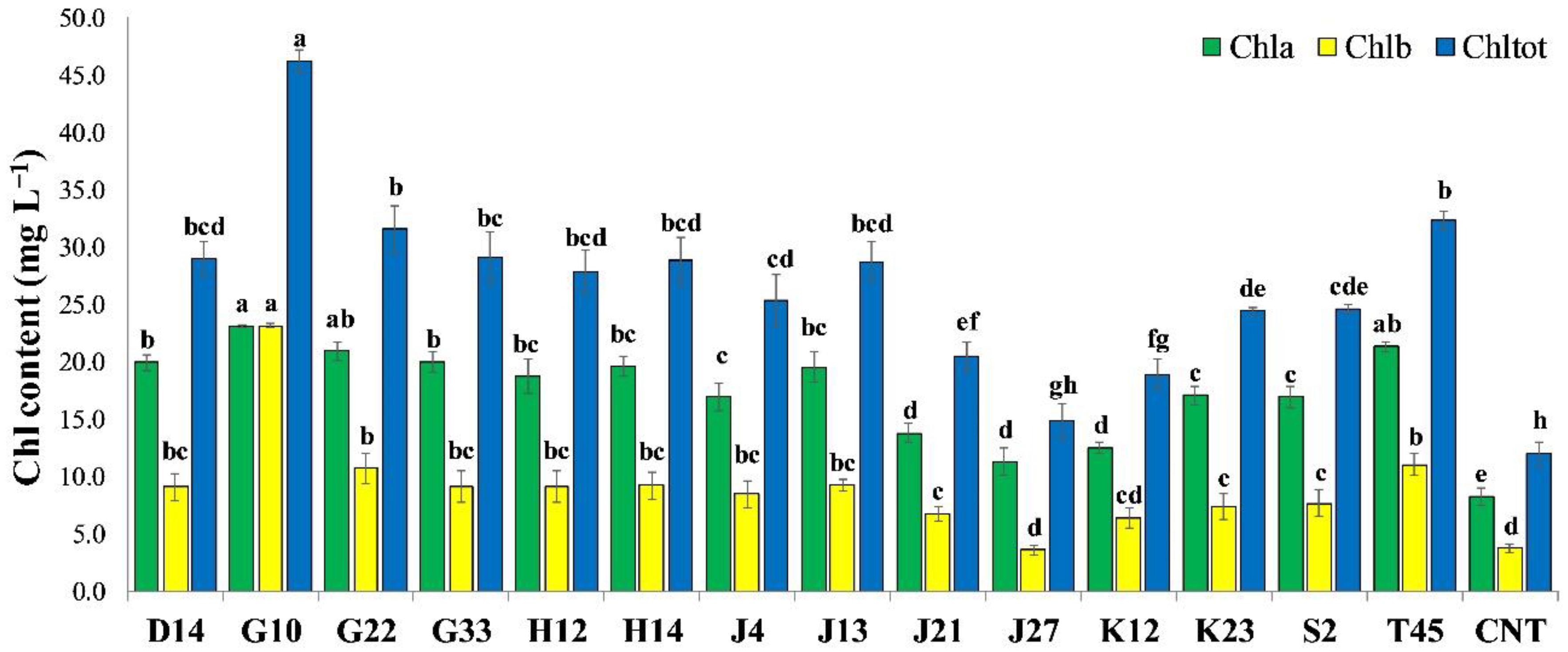
2.4. Greenhouse Experiment on Solanum Lycopersicum
2.5. Statistical Analysis
3. Results and Discussion
3.1. Plant-Growth-Promoting Traits
- D14, Nocardiopsis dassonvillei subsp. dassonvillei (Gene Bank accession MG597514).
- G10, Streptomyces iakyrus (MG597593).
- G22, Streptomyces xantholiticus (MG597582).
- G33, Streptomyces xantholiticus (MG597585).
- H12, Streptomyces albidoflavus (MG597552).
- H14, Nocardiopsis aegyptica (MG597543).
- J4, Streptomyces thinghirensis (MG597590).
- J13, Streptomyces anulatus (MG597579).
- J21, Nocardiopsis alba (MG597576).
- J27, Streptomyces ambofaciens (MG597599).
- K12, Streptomyces xantholiticus (MG597545).
- K23, Streptomyces thinghirensis (MG597560).
- S2, Nocardiopsis aegyptica (MG597572).
- T45, Nocardiopsis dassonvillei subsp. dassonvillei (MG597502).
3.1.1. Phosphate Solubilization
3.1.2. Production of Indole-3-Acetic Acid
3.1.3. Production of Hydrocyanic Acid, Ammonia, and Enzymatic Activities
3.2. In Vitro Association Study by Scanning Electron Microscopy (SEM)
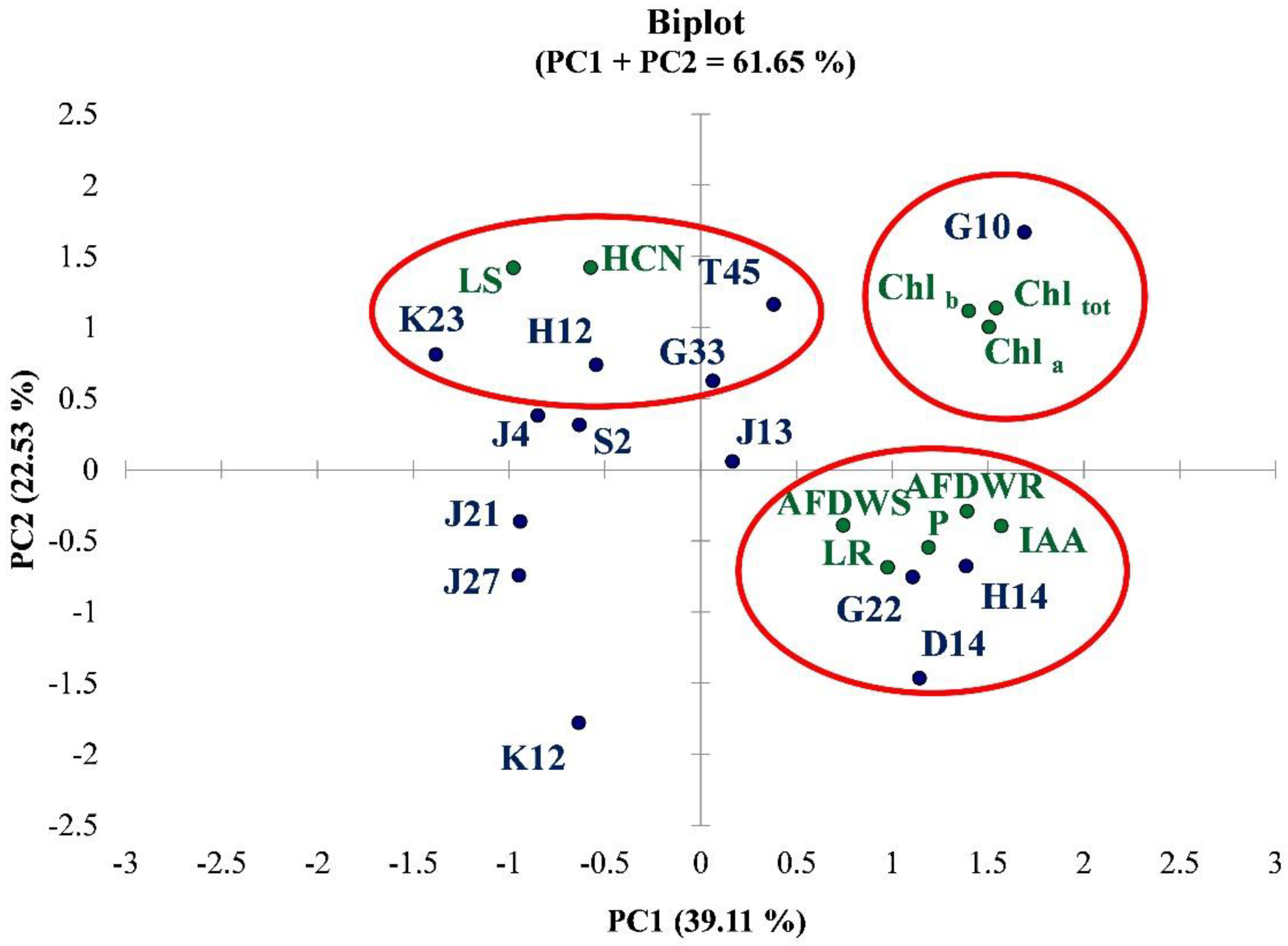
3.3. Greenhouse Experiment on Solanum Lycopersicum
4. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Nath, S.; Matozzo, V.; Bhandari, D.; Faggio, C. Growth and liver histology of Channa punctatus exposed to a common biofertilizer. Nat. Prod. Res. 2019, 33, 1591–1598. [Google Scholar] [CrossRef] [PubMed]
- Savci, S. An agricultural pollutant: Chemical fertilizer. Int. J. Environ. Sci. Dev. 2012, 3, 73. [Google Scholar] [CrossRef] [Green Version]
- Campos, E.V.; Proença, P.L.; Oliveira, J.L.; Bakshi, M.; Abhilash, P.C.; Fraceto, L.F. Use of botanical insecticides for sustainable agriculture: Future perspectives. Ecol. Indic. 2019, 105, 483–495. [Google Scholar] [CrossRef] [Green Version]
- Mia, M.B.; Shamsuddin, Z.H.; Mahmood, M. Use of plant growth promoting bacteria in banana: A new insight for sustainable banana production. Int. J. Agric. Biol. 2010, 12, 459–467. [Google Scholar]
- Gamez, R.; Cardinale, M.; Montes, M.; Ramirez, S.; Schnell, S.; Rodriguez, F. Screening, plant growth promotion and root colonization pattern of two rhizobacteria (Pseudomonas fluorescens Ps006 and Bacillus amyloliquefaciens Bs006) on banana cv. Williams (Musa acuminata Colla). Microbiol. Res. 2019, 220, 12–20. [Google Scholar] [CrossRef] [PubMed]
- Wu, S.C.; Cao, Z.H.; Li, Z.G.; Cheung, K.C.; Wong, M.H. Effects of biofertilizer containing N-fixer, P and K solubilizers and AM fungi on maize growth: A greenhouse trial. Geoderma 2005, 125, 155–166. [Google Scholar] [CrossRef]
- Gupta, G.; Parihar, S.S.; Ahirwar, N.K.; Snehi, S.K.; Singh, V. Plant growth promoting rhizobacteria (PGPR): Current and future prospects for development of sustainable agriculture. J. Microb. Biochem. Technol. 2015, 7, 096–102. [Google Scholar]
- Shilev, S. Soil rhizobacteria regulating the uptake of nutrients and undesirable elements by plants. In Plant Microbe Symbiosis: Fundamentals and Advances; Springer: New Delhi, India, 2013; pp. 147–167. [Google Scholar]
- Bhattacharyya, P.N.; Jha, D.K. Plant growth-promoting rhizobacteria (PGPR): Emergence in agriculture. World J. Microbiol. Biotechnol. 2012, 28, 1327–1350. [Google Scholar] [CrossRef] [PubMed]
- Glick, B.R.; Bashan, Y. Genetic manipulation of plant growth-promoting bacteria to enhance biocontrol of phytopathogens. Biotechnol. Adv. 1997, 15, 353–378. [Google Scholar] [CrossRef] [Green Version]
- Kyaw, E.P.; Soe, M.M.; San San Yu, Z.K.L.; Lynn, T.M. Study on plant growth promoting activities of Azotobacter isolates for sustainable agriculture in Myanmar. J. Biotech. Biores. 2019, 1, JBB.000524. [Google Scholar]
- Ivushkin, K.; Bartholomeus, H.; Bregt, A.K.; Pulatov, A.; Franceschini, M.H.D.; Kramer, H.; van Loo, E.N.; Jaramillo Roman, V.; Finkers, R. UAV based soil salinity assessment of cropland. Geoderma 2019, 338, 502–512. [Google Scholar] [CrossRef]
- Zanetti, F.; Zegada-Lizarazu, W.; Lambertini, C.; Monti, A. Salinity effects on germination, seedlings and full-grown plants of upland and lowland switchgrass cultivars. Biomass Bioenergy 2019, 120, 273–280. [Google Scholar] [CrossRef]
- Egamberdieva, D.; Wirth, S.; Bellingrath-Kimura, S.D.; Mishra, J.; Arora, N.K. Salt-Tolerant Plant Growth Promoting Rhizobacteria for Enhancing Crop Productivity of Saline Soils. Front. Microbiol. 2019, 10, 2791. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pace, L.; Pellegrini, M.; Palmieri, S.; Rocchi, R.; Lippa, L.; Del Gallo, M. Plant growth-promoting rhizobacteria for In Vitro and Ex Vitro performance enhancement of Apennines’ genepì (Artemisia umbelliformis subsp. eriantha), an endangered phytotherapeutic plant. Vitro Cell. Dev. Biol. Plant 2020, 56, 134–142. [Google Scholar] [CrossRef]
- Olanrewaju, O.S.; Babalola, O.O. Streptomyces: Implications and interactions in plant growth promotion. Appl. Microbiol. Biotechnol. 2019, 103, 1179–1188. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Toumatia, O.; Yekkour, A.; Goudjal, Y.; Riba, A.; Coppel, Y.; Mathieu, F.; Sabaou, N.; Zitouni, A. Antifungal properties of an actinomycin D-producing strain, Streptomyces sp. IA1, isolated from a Saharan soil. J. Basic Microbiol. 2015, 55, 221–228. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Olanrewaju, O.S.; Glick, B.R.; Babalola, O.O. Mechanisms of action of plant growth promoting bacteria. World J. Microbiol. Biotechnol. 2017, 33, 197. [Google Scholar] [CrossRef] [Green Version]
- Goudjal, Y.; Toumatia, O.; Yekkour, A.; Sabaou, N.; Mathieu, F.; Zitouni, A. Biocontrol of Rhizoctonia solani damping-off and promotion of tomato plant growth by endophytic actinomycetes isolated from native plants of Algerian Sahara. Microbiol. Res. 2014, 169, 59–65. [Google Scholar] [CrossRef] [PubMed]
- Jog, R.; Nareshkumar, G.; Rajkumar, S. Enhancing soil health and plant growth promotion by actinomycetes. In Plant Growth Promoting Actinobacteria; Springer: Singapore, 2016; pp. 33–45. [Google Scholar]
- Wahyudi, A.T.; Priyanto, J.A.; Afrista, R.; Kurniati, D.; Astuti, R.I.; Akhdiya, A. Plant growth promoting activity of actinomycetes isolated from soybean rhizosphere. Online J. Biol. Sci. 2019, 19, 1–8. [Google Scholar] [CrossRef] [Green Version]
- Jog, R.; Pandya, M.; Nareshkumar, G.; Rajkumar, S. Mechanism of phosphate solubilization and antifungal activity of Streptomyces spp. isolated from wheat roots and rhizosphere and their application in improving plant growth. Microbiology 2014, 160, 778–788. [Google Scholar] [CrossRef] [Green Version]
- Boukaya, N.; Goudjal, Y.; Zamoum, M.; Chaabane Chaouch, F.; Sabaou, N.; Mathieu, F.; Zitouni, A. Biocontrol and plant-growth-promoting capacities of actinobacterial strains from the Algerian Sahara and characterisation of Streptosporangium becharense SG1 as a promising biocontrol agent. Biocontrol Sci. Technol. 2018, 28, 858–873. [Google Scholar] [CrossRef]
- Chen, W.; Xie, T.-T.; Zeng, H. Formation, antibiotic resistance, and control strategies of Staphylococcus epidermidis biofilm. In Bacterial Biofilms; IntechOpen: London, UK, 2019. [Google Scholar]
- Smati, M.; Kitouni, M. Diversity of actinobacteria in the marshes of Ezzemoul and Djendli in northeastern Algeria. Eur. J. Ecol. 2019, 5, 41–53. [Google Scholar] [CrossRef] [Green Version]
- Nautiyal, C.S. An efficient microbiological growth medium for screening phosphate solubilizing microorganisms. FEMS Microbiol. Lett. 1999, 170, 265–270. [Google Scholar] [CrossRef] [PubMed]
- Pikovskaya, R.I. Mobilization of phosphorus in soil in connection with the vital activity of some microbial species. Microbiology 1948, 17, 362–370. [Google Scholar]
- Hafsa, C.S.; Allaoua, S.; Mostefa, G.; Bilal, Y.; Fouzia, A. Solubilization of phosphate by the Bacillus under salt stress and in the presence of osmoprotectant compounds. Afr. J. Microbiol. Res. 2013, 7, 4562–4571. [Google Scholar] [CrossRef] [Green Version]
- Olsen, S.R.; Sommers, L.E.; Page, A.L. Phosphorus. In Methods of Soil Analisis, 2nd ed.; Page, A.L., Miller, R.H., Keeney, D.R., Eds.; American Society of Agronomy: Madison, WI, USA, 1982; pp. 403–430. [Google Scholar]
- Gordon, S.A.; Weber, R.P. Colorimetric estimation of indoleacetic acid. Plant Physiol. 1951, 26, 192. [Google Scholar] [CrossRef] [Green Version]
- Fatmawati, U.; Meryandini, A.; Nawangsih, A.A.; Wahyudi, A.T. Screening and characterization of actinomycetes isolated from soybean rhizosphere for promoting plant growth. Biodivers. J. Biol. Divers. 2019, 20, 2970–2977. [Google Scholar] [CrossRef]
- Donate-Correa, J.; León-Barrios, M.; Pérez-Galdona, R. Screening for plant growth-promoting rhizobacteria in Chamaecytisus proliferus (tagasaste), a forage tree-shrub legume endemic to the Canary Islands. Plant Soil 2005, 266, 261–272. [Google Scholar] [CrossRef]
- Cappuccino, J.G.; Sherman, N. Microbiology—A Laboratory Manual; The Benjamin/Cummings Publishing Co., Inc.: Menlo Park, CA, USA, 1992; pp. 188–247. [Google Scholar]
- Pranay, K.; Padmadeo, S.R.; Jha, V.; Prasad, B. Screening and identification of amylase producing strains of Bacillus. J. Appl. Biol. Biotech. 2019, 7, 57–62. [Google Scholar]
- Chaiharn, M.; Chunhaleuchanon, S.; Kozo, A.; Lumyong, S. Screening of rhizobacteria for their plant growth promoting activities. Curr. Appl. Sci. Technol. 2008, 8, 18–23. [Google Scholar]
- Raval, K.M.; Vaswani, P.S.; Majumder, D.R. Biotransformation of a single amino-acid L-tyrosine into a bioactive molecule L-DOPA. Int. J. Sci. Res. 2012, 2, 2250–3153. [Google Scholar]
- Nonomura, H. Key for classification and identification of 458 species of the Streptomycetes included in ISP. J. Ferment. Technol. 1974, 52, 78–92. [Google Scholar]
- Prasad, P.; Bedi, S.; Singh, T. In Vitro cellulose rich organic material degradation by cellulolytic Streptomyces albospinus (MTCC 8768). Malays. J. Microbiol. 2012, 8, 164–169. [Google Scholar] [CrossRef] [Green Version]
- Dellaras, C. Pratique en Microbiologie de Laboratoire? Recherche de Bactéries et de Levures-Moisissures; Lavoisier: Paris, France, 2014; pp. 65–120. [Google Scholar]
- Nitsch, B.; Kutzner, H.J. Egg-yolk agar as a diagnostic medium for Streptomycetes. Experientia 1969, 25, 220–221. [Google Scholar] [CrossRef]
- Hankin, L.; Zucker, M.; Sands, D.C. Improved solid medium for the detection and enumeration of pectolytic bacteria. Appl. Environ. Microbiol. 1971, 22, 205–209. [Google Scholar] [CrossRef] [Green Version]
- Pagnani, G.; Galieni, A.; Stagnari, F.; Pellegrini, M.; Del Gallo, M.; Pisante, M. Open field inoculation with PGPR as a strategy to manage fertilization of ancient Triticum genotypes. Biol. Fertil. Soils 2020, 56, 111–124. [Google Scholar] [CrossRef]
- Pagnani, G.; Pellegrini, M.; Galieni, A.; D’Egidio, S.; Matteucci, F.; Ricci, A.; Stagnari, F.; Sergi, M.; Sterzo, C.L.; Pisante, M. Plant growth-promoting rhizobacteria (PGPR) in Cannabis sativa ‘Finola’ cultivation: An alternative fertilization strategy to improve plant growth and quality characteristics. Ind. Crops Prod. 2018, 123, 75–83. [Google Scholar] [CrossRef]
- Murashige, T.; Skoog, F. A revised medium for rapid growth and bio assays with tobacco tissue cultures. Physiol. Plant. 1962, 15, 473–497. [Google Scholar] [CrossRef]
- Silini, A.; Cherif-Silini, H.; Yahiaoui, B. Growing varieties durum wheat (Triticum durum) in response to the effect of osmolytes and inoculation by Azotobacter chroococcum under salt stress. Afr. J. Microbiol. Res. 2016, 10, 387–399. [Google Scholar]
- Arnon, D.I. Copper enzymes in isolated chloroplasts. Polyphenoloxidase in Beta vulgaris. Plant Physiol. 1949, 24, 1–15. [Google Scholar] [CrossRef] [Green Version]
- Franco-Correa, M.; Quintana, A.; Duque, C.; Suarez, C.; Rodríguez, M.X.; Barea, J.-M. Evaluation of actinomycete strains for key traits related with plant growth promotion and mycorrhiza helping activities. Appl. Soil Ecol. 2010, 45, 209–217. [Google Scholar] [CrossRef]
- Kaur, T.; Sharma, D.; Kaur, A.; Manhas, R.K. Antagonistic and plant growth promoting activities of endophytic and soil actinomycetes. Arch. Phytopathol. Plant Prot. 2013, 46, 1756–1768. [Google Scholar] [CrossRef]
- Gangwar, M.; Dogra, S.; Gupta, U.P.; Kharwar, R.N. Diversity and biopotential of endophytic actinomycetes from three medicinal plants in India. Afr. J. Microbiol. Res. 2014, 8, 184–191. [Google Scholar]
- Sebihi, F.Z.; Benguedouar, A.; Benhizia, Y.; Sanchez, J.; Gallego, E. Evaluation of multi-trait plant growth promoting Pseudomonas fluorescens isolated from Constantine Wheat rhizosphere soil (Algeria) and screening there antifungal activity against two species of Fusarium. Adv. Environ. Biol. 2016, 10, 102–116. [Google Scholar]
- Khan, M.S.; Zaidi, A.; Ahmad, E. Mechanism of phosphate solubilization and physiological functions of phosphate-solubilizing microorganisms. In Phosphate Solubilizing Microorganisms; Khan, M.S., Zaidi, A., Musarrat, J., Eds.; Springer: Cham, Switzerland, 2014; pp. 31–62. [Google Scholar]
- Faried, A.-S.M. Isolation and Characterization of Phosphate Solubilizing Actinomycetes from Rhizosphere Soil. Ph.D. Thesis, Fac of Agric, Assiut University, Assiut, Egypt, 2019. [Google Scholar]
- Ahmad, F.; Ahmad, I.; KHAN, M.S. Indole acetic acid production by the indigenous isolates of Azotobacter and fluorescent Pseudomonas in the presence and absence of tryptophan. Turk. J. Biol. 2005, 29, 29–34. [Google Scholar]
- Radwan, T.E.-S.E.-D.; Mohamed, Z.K.; Reis, V. Production of indole-3-acetic acid by different strains of Azospirillum and Herbaspirillum spp. Symbiosis 2002, 32, 39–54. [Google Scholar]
- Barazani, O.Z.; Friedman, J. Is IAA the major root growth factor secreted from plant-growth-mediating bacteria? J. Chem. Ecol. 1999, 25, 2397–2406. [Google Scholar] [CrossRef]
- Silini, A. Effets des Molécules Osmoprotectrices sur la Survie et L’activité de Azotobacter et sur la Croissance du blé dur en Milieu Salin. Ph.D. Thesis, Université Ferhat Abbas de Sétif, Sétif, Algeria, 2013. [Google Scholar]
- Glick, B.R. The enhancement of plant growth by free-living bacteria. Can. J. Microbiol. 1995, 41, 109–117. [Google Scholar] [CrossRef]
- Dastager, S.G.; Deepa, C.K.; Pandey, A. Potential plant growth-promoting activity of Serratia nematodiphila NII-0928 on black pepper (Piper nigrum L.). World J. Microbiol. Biotechnol. 2011, 27, 259–265. [Google Scholar] [CrossRef]
- Patten, C.L.; Glick, B.R. Role of Pseudomonas putida indoleacetic acid in development of the host plant root system. Appl Environ. Microbiol. 2002, 68, 3795–3801. [Google Scholar] [CrossRef] [Green Version]
- Mirza, M.S.; Ahmad, W.; Latif, F.; Haurat, J.; Bally, R.; Normand, P.; Malik, K.A. Isolation, partial characterization, and the effect of plant growth-promoting bacteria (PGPB) on micro-propagated sugarcane In Vitro. Plant Soil 2001, 237, 47–54. [Google Scholar] [CrossRef]
- Ashwini, M.; Doddamani, L.C.; Babar, S.R. Characterization and evaluation of actinomycete isolates for traits associated with plant growth promotion. J. Pharmacogn. Phytochem. 2018, 7, 1177–1180. [Google Scholar]
- Sreevidya, M.; Gopalakrishnan, S.; Kudapa, H.; Varshney, R.K. Exploring plant growth-promotion actinomycetes from vermicompost and rhizosphere soil for yield enhancement in chickpea. Braz. J. Microbiol. 2016, 47, 85–95. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ahmad, F.; Ahmad, I.; Khan, M.S. Screening of free-living rhizospheric bacteria for their multiple plant growth promoting activities. Microbiol. Res. 2008, 163, 173–181. [Google Scholar] [CrossRef] [PubMed]
- Kumar, A.; Kumar, A.; Devi, S.; Patil, S.; Payal, C.; Negi, S. Isolation, screening and characterization of bacteria from Rhizospheric soils for different plant growth promotion (PGP) activities: An In Vitro study. Recent Res. Sci. Technol. 2012, 4, 1–5. [Google Scholar]
- Bashan, Y.; De-Bashan, L.E. Bacteria/Plant growth-promotion. In Encyclopedia of Soils in the Environment; Hillel, D., Ed.; Elsevier: Oxford, UK, 2005; Volume 1, pp. 103–115. [Google Scholar]
- Joseph, B.; Ranjan Patra, R.; Lawrence, R. Characterization of plant growth promoting rhizobacteria associated with chickpea (Cicer arietinum L.). Int. J. Plant Prod. 2012, 1, 141–152. [Google Scholar]
- Mota, M.S.; Gomes, C.B.; Souza Júnior, I.T.; Moura, A.B. Bacterial selection for biological control of plant disease: Criterion determination and validation. Braz. J. Microbiol. 2017, 48, 62–70. [Google Scholar] [CrossRef] [Green Version]
- Passari, A.K.; Mishra, V.K.; Gupta, V.K.; Yadav, M.K.; Saikia, R.; Singh, B.P. In Vitro and In Vivo plant growth promoting activities and DNA fingerprinting of antagonistic endophytic actinomycetes associates with medicinal plants. PLoS ONE 2015, 10, e0139468. [Google Scholar] [CrossRef] [Green Version]
- Weise, T.; Kai, M.; Piechulla, B. Bacterial ammonia causes significant plant growth inhibition. PLoS ONE 2013, 8, e63538. [Google Scholar] [CrossRef]
- Reetha, S.; Selvakumar, G.; Bhuvaneswari, G.; Thamizhiniyan, P.; Ravimycin, T. Screening of cellulase and pectinase by using Pseudomonas fluorescens and Bacillus subtilis. Int. Lett. Nat. Sci. 2014, 13, 75–80. [Google Scholar]
- Turan, M.; Nikerel, E.; Kaya, K.; Kitir, N.; Gunes, A.; Mokhtari, N.E.P.; Tüfenkçi, Ş.; Karaman, M.R.; Çimrin, K.M. Enzyme Dynamic in Plant Nutrition Uptake and Plant Nutrition. In Enzyme Inhibitors and Activators; Senturk, M., Ed.; Inctechopen: London, UK, 2017; pp. 225–242. [Google Scholar]
- Mushtaq, S.; Shafiq, M.; Ashraf, T.; Haider, M.S.; Ashfaq, M.; Ali, M. Characterization of plant growth promoting activities of bacterial endophytes and their antibacterial potential isolated from citrus. J. Anim. Plant Sci. 2019, 29, 978–991. [Google Scholar]
- Vranova, V.; Rejsek, K.; Formanek, P. Proteolytic activity in soil: A review. Appl. Soil Ecol. 2013, 70, 23–32. [Google Scholar] [CrossRef]
- Stach, N.; Kaszycki, P.; W\ladyka, B.; Dubin, G. Extracellular proteases of Staphylococcus spp. In Pet-to-Man Travelling Staphylococci; Savini, V., Ed.; Academic Press: Cambridge, MA, USA, 2018; pp. 135–145. [Google Scholar]
- Vardharajula, S.; SkZ, A.; Shiva Krishna Prasad Vurukonda, S.; Shrivastava, M. Plant growth promoting endophytes and their interaction with plants to alleviate abiotic stress. Curr. Biotechnol. 2017, 6, 252–263. [Google Scholar] [CrossRef]
- Islam, R.; Datta, B. Diversity of chitinases and their industrial potential. Int. J. Appl. Res. 2015, 1, 55–60. [Google Scholar]
- Pascoal, A.; Estevinho, L.M.; Martins, I.M.; Choupina, A.B. Novel sources and functions of microbial lipases and their role in the infection mechanisms. Physiol. Mol. Plant Pathol. 2018, 104, 119–126. [Google Scholar] [CrossRef]
- Dal Cortivo, C.; Barion, G.; Visioli, G.; Mattarozzi, M.; Mosca, G.; Vamerali, T. Increased root growth and nitrogen accumulation in common wheat following PGPR inoculation: Assessment of plant-microbe interactions by ESEM. Agric. Ecosyst. Environ. 2017, 247, 396–408. [Google Scholar] [CrossRef]
- Primo, E.D.; Ruiz, F.; Masciarelli, O.; Giordano, W. Biofilm formation and biosurfactant activity in plant-associated bacteria. In Bacterial Metabolites in Sustainable Agroecosystem; Maheshwari, D., Ed.; Springer: Cham, Switzerland, 2015; Volume 12, pp. 337–349. [Google Scholar]
- Patel, K.B.; Thakker, J.N. Growth promotion and biocontrol activity of Nocardiopsis dassonvillei strain YM12: An isolate from coastal agricultural land of Khambhat. Vegetos 2019, 32, 571–582. [Google Scholar] [CrossRef]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| K12 | K23 | H12 | H14 | S2 | D14 | T45 | J4 | J13 | J21 | J27 | G10 | G22 | G33 | |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| HCN | + | + | + | + | + | − | − | + | + | + | + | + | − | + |
| NH3 | + | + | + | + | + | + | + | + | + | + | + | + | + | + |
| Amylase | + | + | + | + | + | + | + | + | + | + | + | − | + | + |
| Caseinase | + | + | − | + | + | + | + | + | + | + | + | + | + | + |
| Catalase | + | + | + | + | + | + | + | + | + | + | + | + | + | + |
| Cellulase | − | + | − | + | − | − | + | + | + | + | + | + | + | − |
| Chitinase | − | − | − | − | − | − | − | − | − | − | − | + | − | − |
| Gelatinase | + | + | + | + | + | + | + | + | + | + | + | + | + | + |
| Lecithinase | − | + | − | − | + | − | − | + | + | + | − | + | − | + |
| Lipase | − | − | − | + | − | − | + | − | − | − | − | − | − | + |
| Lipoprotease | − | + | + | + | + | + | + | + | + | + | + | − | − | + |
| Pectinase | + | + | − | + | + | + | − | + | + | + | − | + | + | + |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Djebaili, R.; Pellegrini, M.; Smati, M.; Del Gallo, M.; Kitouni, M. Actinomycete Strains Isolated from Saline Soils: Plant-Growth-Promoting Traits and Inoculation Effects on Solanum lycopersicum. Sustainability 2020, 12, 4617. https://0-doi-org.brum.beds.ac.uk/10.3390/su12114617
Djebaili R, Pellegrini M, Smati M, Del Gallo M, Kitouni M. Actinomycete Strains Isolated from Saline Soils: Plant-Growth-Promoting Traits and Inoculation Effects on Solanum lycopersicum. Sustainability. 2020; 12(11):4617. https://0-doi-org.brum.beds.ac.uk/10.3390/su12114617
Chicago/Turabian StyleDjebaili, Rihab, Marika Pellegrini, Maria Smati, Maddalena Del Gallo, and Mahmoud Kitouni. 2020. "Actinomycete Strains Isolated from Saline Soils: Plant-Growth-Promoting Traits and Inoculation Effects on Solanum lycopersicum" Sustainability 12, no. 11: 4617. https://0-doi-org.brum.beds.ac.uk/10.3390/su12114617