Genetic and Morphologic Variation in a Potential Mosquito Biocontrol Agent, Hydrochara Affinis (Coleoptera: Hydrophilidae)

 ,
,
Abstract
:1. Introduction
2. Materials and Methods
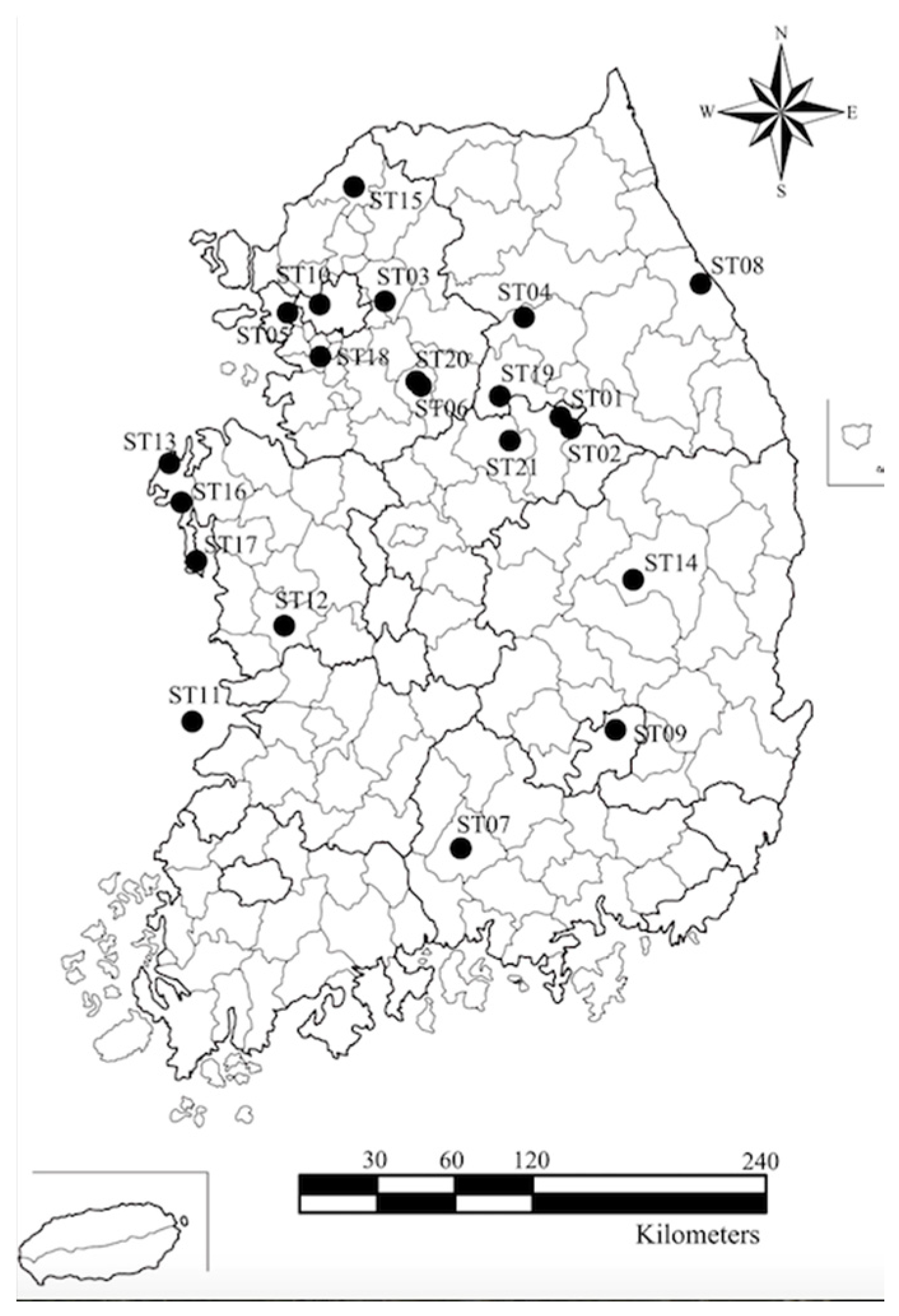
2.1. Sample Collection and COI Sequencing
2.2. Phylogenetic Reconstruction and Haplotype Network Analysis
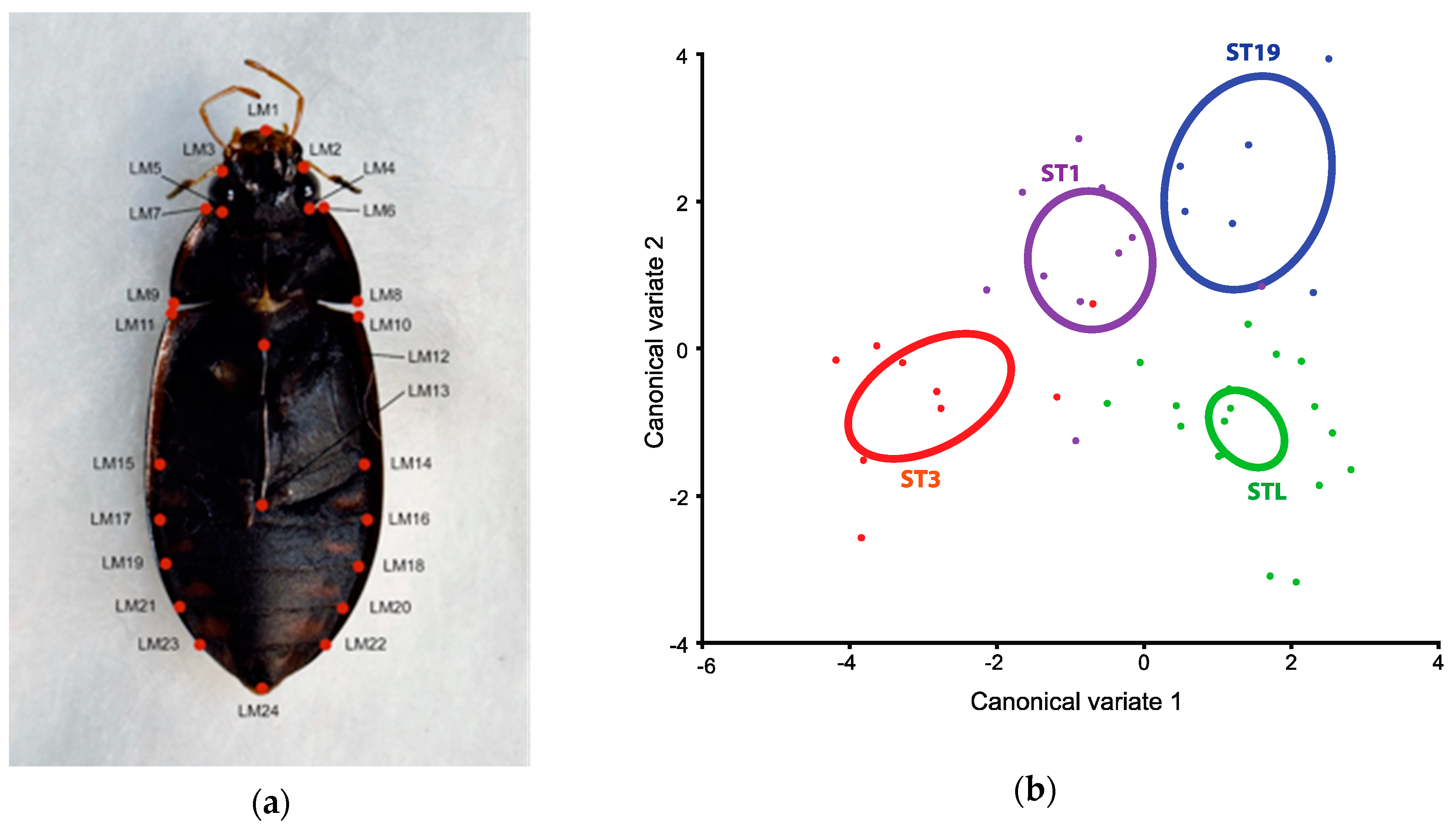
2.3. Geometric Morphometric Analyses
3. Results
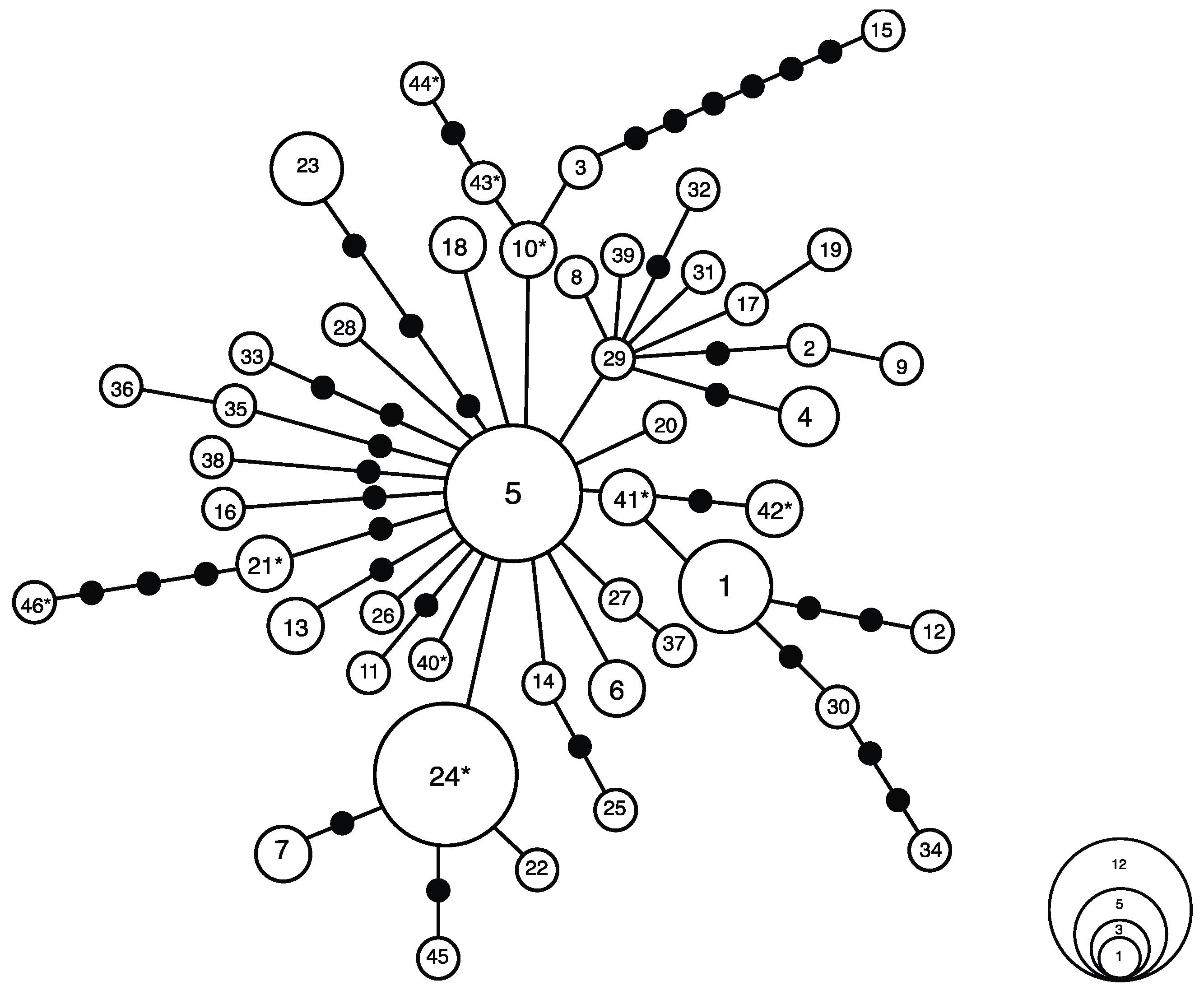
3.1. Haplotype Analysis
3.2. Geometric Morphometric Analysis
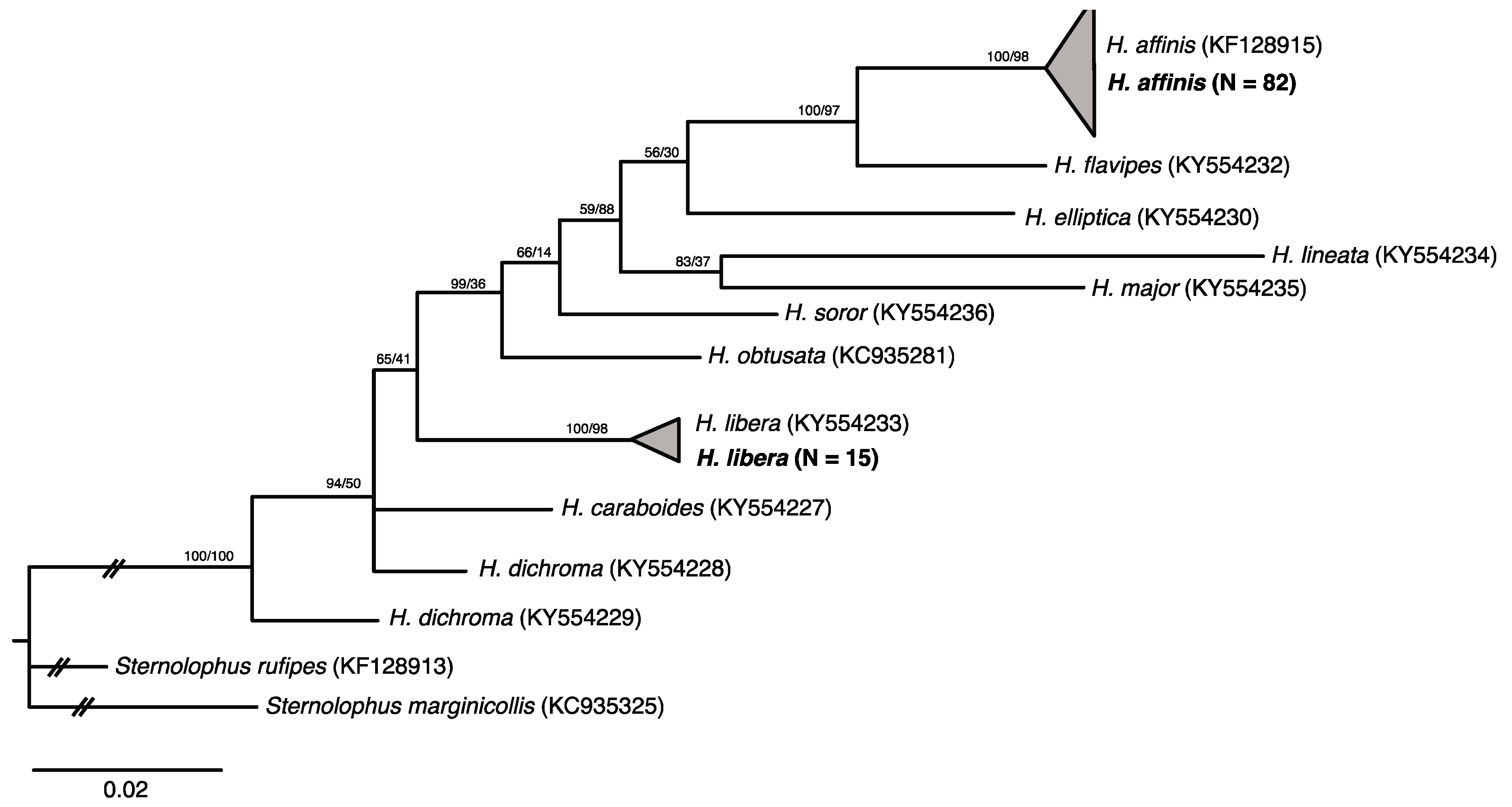
3.3. Phylogenetic Status of H. affinis and H. libera in the Genus Hydrochara
4. Discussion
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Hansen, M. The hydrophiloid beetles. Phylogeny, classification and a revision of the genera (Coleoptera, Hydrophiloidea). Biol. Skr. 1991, 40, 1–367. [Google Scholar]
- Kumar, R.; Hwang, J.S. Larvicidal efficiency of aquatic predators: A perspective for mosquito biocontrol. Zool. Stud. 2006, 45, 447. [Google Scholar]
- Baek, H.M.; Kim, D.G.; Baek, M.J.; Lee, C.Y.; Kang, H.J.; Kim, M.C.; Yoo, J.S.; Bae, Y.J. Predation efficiency and preference of the Hydrophilid Water Beetle Hydrochara affinis (Coleoptera: Hydrophilidae) larvae on two mosquitos Culex pipiens molestus and Ochlerotatus togoi under laboratory conditions. Korean J. Environ. Biol. 2014, 32, 112–117. [Google Scholar] [CrossRef]
- Shaalan, E.A.S.; Canyon, D.V. Aquatic insect predators and mosquito control. Trop. Biomed. 2009, 26, 223–261. [Google Scholar] [PubMed]
- Unruh, T.R.; Woolley, J.B. Handbook of Biological Control; Bellows, T.S., Fisher, T.W., Eds.; Academic Press: San Diego, CA, USA, 1999; pp. 57–85. [Google Scholar]
- Roderick, G.K.; Navajas, M. Genes in new environments, genetics and evolution in biological control. Nat. Rev. Genet. 2003, 4, 889–899. [Google Scholar] [CrossRef] [PubMed]
- Rauth, S.J.; Hinz, H.L.; Gerber, E.; Hufbauer, R.A. The benefits of pre-release population genetics: A case study using Ceutorhynchus scrobicollis, a candidate agent of garlic mustard. Alliaria Petiolata Biol. Control. 2011, 56, 67–75. [Google Scholar] [CrossRef]
- Taylor, S.J.; Downie, D.A.; Paterson, I.D. Genetic diversity of introduced populations of the water hyacinth biological control agent Eccritotarsus catarinensis (Hemiptera: Miridae). Biol. Cont. 2011, 58, 330–336. [Google Scholar] [CrossRef]
- Mackauer, M. Genetic problems in the production of biological control agents. Annu. Rev. Entomol. 1976, 21, 369–385. [Google Scholar] [CrossRef]
- Kuriwada, T.; Kumano, N.; Shiromoto, K.; Haraguchi, D. Effect of mass rearing on life history traits and inbreeding depression in the sweetpotato weevil (Coleoptera: Brentidae). J. Econ. Entomol. 2010, 103, 1144–1148. [Google Scholar] [CrossRef]
- Sørensen, J.G.; Addison, M.F.; Terblanche, J.S. Mass-rearing of insects for pest management: Challenges; synergies and advances from evolutionary physiology. Crop. Prot. 2012, 38, 87–94. [Google Scholar] [CrossRef]
- Daly, H.V. Insect morphometrics. Annu. Rev. Entomol. 1985, 30, 415–438. [Google Scholar] [CrossRef]
- Grant, P.R. Speciation and the adaptive radiation of Darwin’s Finches: The complex diversity of Darwin’s finches may provide a key to the mystery of how intraspecific variation is transformed into interspecific variation. Am. Sci. 1981, 69, 653–663. [Google Scholar]
- Grant, P.R.; Price, T.D. Population variation in continuously varying traits as an ecological genetics problem. Am. Zool. 1981, 21, 795–811. [Google Scholar] [CrossRef]
- Monteiro, L.R.; Nogueira, M.R. Adaptive radiations, ecological specialization, and the evolutionary integration of complex morphological structures. Evolution 2010, 64, 724–744. [Google Scholar] [CrossRef] [PubMed]
- Dujardin, J.P. Modern morphometrics of medically important insects. In Genetics and Evolution of Infectious Disease; Elsevier: New York, NY, USA, 2011; pp. 473–501. [Google Scholar]
- Monteiro, L.R.; Diniz-Filho, J.A.F.; dos Reis, S.F.; Araújo, E.D. Geometric estimates of heritability in biological shape. Evolution 2002, 56, 563–572. [Google Scholar] [CrossRef]
- Claude, J.; Paradis, E.; Tong, H.; Auffray, J.C. A geometric morphometric assessment of the effects of environment and cladogenesis on the evolution of the turtle shell. Biol. J. Linn. Soc. 2003, 79, 485–501. [Google Scholar] [CrossRef] [Green Version]
- Sharpe, D.M.; Räsänen, K.; Berner, D.; Hendry, A.P. Genetic and environmental contributions to the morphology of lake and stream stickleback: Implications for gene flow and reproductive isolation. Evol. Ecol. Res. 2008, 10, 849–866. [Google Scholar]
- Raffini, F.; Fruciano, C.; Meyer, A. Morphological and genetic correlates in the left–right asymmetric scale-eating cichlid fish of Lake Tanganyika. Biol. J. Linn. Soc. 2018, 124, 67–84. [Google Scholar] [CrossRef]
- Foster, D.J.; Podos, J.; Hendry, A.P. A geometric morphometric appraisal of beak shape in Darwin’s finches. J. Evol. Biol. 2017, 21, 263–275. [Google Scholar] [CrossRef]
- Franssen, N.R. Anthropogenic habitat alteration induces rapid morphological divergence in a native stream fish. Evol. Appl. 2011, 4, 791–804. [Google Scholar] [CrossRef]
- Cordell, S.; Goldstein, G.; Mueller-Dombois, D.; Webb, D.; Vitousek, P.M. Physiological and morphological variation in Metrosideros polymorpha, a dominant Hawaiian tree species, along an altitudinal gradient: The role of phenotypic plasticity. Oecologia 1998, 113, 188–196. [Google Scholar] [CrossRef] [PubMed]
- Ree, H.I.; Lee, W.J. Laboratory studies on predation efficacy of some Odonata nymphs and Coleoptera larvae against mosquito larvae. Korean J. Appl. Entomol. 1983, 13, 31–38. [Google Scholar]
- Han, M.S.; Bang, H.S.; Kim, M.H.; Kang, K.K.; Jung, M.P.; Lee, D.B. Distribution characteristics of water scavenger beetles (Hydrophilidae) in Korean paddy field. Korean J. Environ. Agric. 2010, 29, 427–433. [Google Scholar] [CrossRef] [Green Version]
- Choi, S.K.; Kim, M.H.; Choe, L.J.; Eo, J.; Bang, H.S. Prediction of the flight times of Hydrochara affinis and Sternolophus rufipes in paddy fields based on RCP 8.5 Scenario. Korean J. Agric. For. Meteorol. 2016, 18, 16–29. [Google Scholar] [CrossRef]
- ESRI. ArcGIS Desktop Release 10.5.1; Environmental Systems Research Institute: Redlands, CA, USA, 2017. [Google Scholar]
- Simon, C.; Frati, F.; Beckenbach, A.; Crespi, B.; Liu, H.; Flook, P. Evolution, weighting, and phylogenetic utility of mitochondrial gene sequences and a compilation of conserved polymerase chain reaction primers. Ann. Entomol. Soc. Am. 1994, 87, 651–701. [Google Scholar] [CrossRef]
- Thompson, J.D.; Higgins, D.G.; Gibson, T.J. CLUSTAL W: Improving the sensitivity of progressive multiple sequence alignment through sequence weighting, position-specific gap penalties and weight matrix choice. Nucleic Acids Res. 1994, 22, 4673–4680. [Google Scholar] [CrossRef] [Green Version]
- Hall, T.A. BioEdit: A user-friendly biological sequence alignment editor and analysis program for Windows 95/98/NT. Nucleic Acids Symp. Ser. 1999, 41, 95–98. [Google Scholar]
- Kumar, S.; Stecher, G.; Tamura, K. MEGA7: Molecular evolutionary genetics analysis version 7.0 for bigger datasets. Mol. Biol. Evol. 2016, 33, 1870–1874. [Google Scholar] [CrossRef] [Green Version]
- Ronquist, F.; Teslenko, M.; Van Der Mark, P.; Ayres, D.L.; Darling, A.; Höhna, S.; Larget, B.; Liu, L.; Suchard, M.A.; Huelsenbeck, J.P. MrBayes 3.2: Efficient Bayesian phylogenetic inference and model choice across a large model space. Syst. Biol. 2012, 61, 539–542. [Google Scholar] [CrossRef] [Green Version]
- Darriba, D.; Taboada, G.L.; Doallo, R.; Posada, D. jModelTest 2: More models, new heuristics and parallel computing. Nat. Methods 2012, 9, 772. [Google Scholar] [CrossRef] [Green Version]
- Guindon, S.; Dufayard, J.F.; Lefort, V.; Anisimova, M.; Hordijk, W.; Gascuel, O. New algorithms and methods to estimate maximum-likelihood phylogenies: Assessing the performance of PhyML 3.0. Syst. Biol. 2010, 59, 307–321. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Librado, P.; Rozas, J. DnaSP v5: A software for comprehensive analysis of DNA polymorphism data. Bioinformatics 2009, 25, 1451–1452. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Teacher, A.G.F.; Griffiths, D.J. HapStar: Automated haplotype network layout and visualization. Mol. Ecol. Resour. 2011, 11, 151–153. [Google Scholar] [CrossRef] [PubMed]
- Excoffier, L.; Laval, G.; Schneider, S. Arlequin (version 3.0): An integrated software package for population genetics data analysis. Evol. Bioinform. 2005, 1, 47–50. [Google Scholar] [CrossRef] [Green Version]
- Petit, R.J.; Mousadik, A.E.; Pons, O. Identifying populations for conservation on the basis of genetic markers. Biol. Conserv. 1998, 12, 844–855. [Google Scholar] [CrossRef]
- Rohlf, F.J. tpsUtil, Version 1.74; State University of New York at Stony Brook: New York, NY, USA, 2017. [Google Scholar]
- Albert, P.; Moureau, B.; Dommergues, J.L.; David, B. Differentiation at a microgeographical scale within two species of ground beetle, Carabus auronitens and C. nemoralis (Coleoptera, Carabidae): A geometrical morphometric approach. Zool. Scr. 2001, 30, 299–311. [Google Scholar] [CrossRef]
- Lemic, D.; Benítez, H.A.; Püschel, T.A.; Gašparić, H.V.; Šatvar, M.; Bažok, R. Ecological morphology of the sugar beet weevil Croatian populations: Evaluating the role of environmental conditions on body shape. Zool. Anz. 2016, 260, 25–32. [Google Scholar] [CrossRef]
- Rohlf, F.J. tpsDig, Version 2.30; Department of Ecology and Evolution, State University of New York at Stony Brook: New York, NY, USA, 2017. [Google Scholar]
- Klingenberg, C.P.; Barluenga, M.; Meyer, A. Shape analysis of symmetric structures: Quantifying variation among individuals and asymmetry. Evolution 2002, 56, 1909–1920. [Google Scholar] [CrossRef] [Green Version]
- Benítez, H.A.; Püschel, T.A.; Lemic, D.; Čačija, M.; Kozina, A.; Bažok, R. Ecomorphological variation of the wireworm cephalic capsule: Studying the interaction of environment and geometric shape. PLoS ONE 2014, 9, e102059. [Google Scholar] [CrossRef] [Green Version]
- Klingenberg, C.P. MorphoJ: An integrated software package for geometric morphometrics. Mol. Ecol. Resour. 2011, 11, 353–357. [Google Scholar] [CrossRef]
- Goodall, C. Procrustes methods in the statistical analysis of shape. J. R. Stat. Soc. Ser. B Stat. Methodol. 1991, 53, 285–339. [Google Scholar] [CrossRef]
- Klingenberg, C.P.; McIntyre, G.S. Geometric morphometrics of developmental instability: Analyzing patterns of fluctuating asymmetry with Procrustes methods. Evolution 1998, 52, 1363–1375. [Google Scholar] [CrossRef] [PubMed]
- Hajek, A.E.; Hurley, B.P.; Kenis, M.; Garnas, J.R.; Bush, S.J.; Wingfield, M.J.; Michael, J.; Van Lenteren, J.C.; Cock, M.J.W. Exotic biological control agents: A solution or contribution to arthropod invasions? Biol. Invasions 2016, 18, 953–969. [Google Scholar] [CrossRef] [Green Version]
- Benelli, G.; Jeffries, C.L.; Walker, T. Biological control of mosquito vectors: Past, present, and future. Insects 2016, 7, 4–52. [Google Scholar] [CrossRef] [PubMed]
- Kats, L.B.; Ferrer, R.P. Alien predators and amphibian declines: Review of two decades of science and the transition to conservation. Divers. Distrib. 2003, 9, 99–110. [Google Scholar] [CrossRef]
- Rupp, H.R. Adverse assessments of Gambusia affinis: An alternate view for mosquito control practitioners. J. Am. Mosq. Control. Assoc. 1996, 12, 155–159. [Google Scholar]
- Chambers, D.L. Quality control in mass rearing. Annu. Rev. Entomol. 1977, 22, 289–308. [Google Scholar] [CrossRef]
- Quaglietti, B.; Tamisier, L.; Groussier, G.; Fleisch, A.; Le Goff, I.; Ris, N.; Kreiter, P.; Fauvergue, X.; Malausa, T. No inbreeding depression in laboratory-reared individuals of the parasitoid wasp Allotropa burrelli. Ecol. Evol. 2017, 7, 964–973. [Google Scholar] [CrossRef]
- Beever, E.A.; O’Leary, J.; Mengelt, C.; West, J.M.; Julius, S.; Green, N.; Magness, D.; Petes, L.; Stein, B.; Nicotra, A.B.; et al. Improving conservation outcomes with a new paradigm for understanding species’ fundamental and realized adaptive capacity. Conser. Lett. 2016, 9, 131–137. [Google Scholar] [CrossRef] [Green Version]
- Orlofske, J.M.; Baird, D.J. A geometric morphometric approach to establish body-shape trait criteria for aquatic insects. Freshw. Sci. 2014, 33, 978–994. [Google Scholar] [CrossRef] [Green Version]
- Gotthard, K.; Nylin, S. Adaptive plasticity and plasticity as an adaptation: A selective review of plasticity in animal morphology and life history. Oikos 1995, 74, 3–17. [Google Scholar] [CrossRef] [Green Version]
- Smetana, A. Revision of the genus Hydrochara Berth. (Coleoptera, Hydrophilidae). Mem. Entomol. Soc. Can. 1980, 112, 1–100. [Google Scholar] [CrossRef]
- Short, A.E. Phylogeny, evolution and classification of the giant water scavenger beetles (Coleoptera, Hydrophilidae: Hydrophilini: Hydrophilina). Syst. Biodivers. 2010, 8, 17–37. [Google Scholar] [CrossRef] [Green Version]
- Darılmaz, M.C.; Kıyak, S.; Short, A.E. Discovery of the water scavenger beetle genus Brownephilus Mouchamps in Turkey (Coleoptera, Hydrophilidae, Hydrophilini. ZooKeys 2010, 53, 13. [Google Scholar] [CrossRef] [PubMed]
- Short, A.E.; Fikáček, M. World catalogue of the Hydrophiloidea (Coleoptera): Additions and corrections II (2006–2010). Acta Entomol. Mus. Natl. Pragae. 2011, 51, 83–122. [Google Scholar]
- Lee, S.H.; Lee, C.E.; Park, H.C. A taxonomy of the aquatic Hydrophilidae of South Korea (Coleoptera). Nat. Life 1988, 18, 79–91. [Google Scholar]
- Lee, S.H. Some additions to the Hydrophilidae of Korea (Coleoptera). Nat. Life 1990, 20, 1–7. [Google Scholar]
- Lee, S.H.; Cho, Y.B.; Lee, C.E. Some addition to the Hydrophilidae of Korea II (Coleoptera). Nat. Life 1993, 23, 145–149. [Google Scholar]
- Jung, S.W.; Lee, D.H.; Ham, S.A.; Huh, J.M.; Hwang, J.M.; Bae, Y.J. Revised checklist of the Korean aquatic insects. Bull. Entomol. Res. 2011, 27, 37–52. [Google Scholar]
- National Institute of Biological Resources. National Species list of Korea. III. Insects (Hexapoda); Designzip: Incheon, Korea, 2019; pp. 224–225. [Google Scholar]
- Toussaint, E.F.; Bloom, D.; Short, A.E. Cretaceous West Gondwana vicariance shaped giant water scavenger beetle biogeography. J. Biogeogr. 2017, 44, 1952–1965. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Site Nr | Localities | N | NH | h | hr | MPD (SD) | π (SD) | Tajima’s D | Fu’s Fs |
|---|---|---|---|---|---|---|---|---|---|
| ST1 | 37°10′16.7″ N, 128°12′12.9″ E | 10 | 9 | 0.978 (0.054) | 3.778 | 4.067 (2.214) | 0.005 (0.003) | −0.816 | −4.318 |
| ST2 | 37°08′25.7″ N, 128°13′54.4″ E | 8 | 8 | 1.000 (0.063) | 4.000 | 5.286 (2.855) | 0.007 (0.004) | −1.447 | −3.752 |
| ST3 | 37°35′01.1″ N, 127°14′15.7″ E | 6 | 6 | 1.000 (0.096) | 3.333 | 2.400 (1.510) | 0.003 (0.002) | −0.496 | −3.851 |
| ST4 | 37°33′04.1″ N, 127°57′10.3″ E | 2 | 1 | 0.000 (0.000) | - | - | - | 0 | 0.000 |
| ST5 | 37°30′11.0″ N, 126°45′57″ E | 1 | 1 | 1.000 (0.000) | - | - | - | 0 | 0.000 |
| ST6 | 37°14′46.0″ N, 127°28′07.4″ E | 2 | 2 | 1.000 (0.500) | - | 4.000 (3.162) | 0.005 (0.006) | 0 | 0.000 |
| ST7 | 35°20′43.0″ N, 127°50′20.0″ E | 2 | 2 | 1.000 (0.500) | - | 4.000 (3.162) | 0.005 (0.006) | 0 | 0.000 |
| ST8 | 37°46′01″ N, 128°53′12.0″ E | 1 | 1 | 1.000 (0.000) | - | - | - | 0 | 1.386 |
| ST9 | 35°54′54.2″ N, 128°36′01.7″ E | 3 | 2 | 1.000 (0.272) | - | 2.000 (1.512) | 0.003 (0.002) | 0 | 1.386 |
| ST10 | 37°34′02.0″ N, 126°53′14.8″ E | 1 | 1 | 1.000 (0.000) | - | - | - | 0 | 0.000 |
| ST11 | 35°48′48.2″ N, 126°24′26.4″ E | 1 | 1 | 1.000 (0.000) | - | - | - | 0 | 0.000 |
| ST12 | 36°14′55.1″ N, 126°51′33.5″ E | 1 | 1 | 1.000 (0.000) | - | - | - | 0 | 0.000 |
| ST13 | 36°50′30.0″ N, 126°11′48.9″ E | 1 | 1 | 1.000 (0.000) | - | - | - | 0 | −0.693 |
| ST14 | 36°29′19.0″ N, 128°40′24.7″ E | 1 | 1 | 1.000 (0.000) | - | - | - | 0 | 0.000 |
| ST15 | 38°01′57.4″ N, 127°04′33.4″ E | 2 | 2 | 1.000 (0.500) | - | 3.000 (2.449) | 0.004 (0.004) | 0 | 0.000 |
| ST16 | 36°40′02.5″ N, 126°18′35.3″ E | 2 | 2 | 1.000 (0.500) | - | 4.000 (3.162) | 0.005 (0.006) | 0 | 1.386 |
| ST17 | 36°26′10.5″ N, 126°23′47.0″ E | 1 | 1 | 1.000 (0.000) | - | - | - | 0 | 0.000 |
| ST18 | 37°21′13.3″ N, 126°55′06.4″ E | 2 | 2 | 1.000 (0.500) | - | 10.000 (7.416) | 0.013 (0.134) | 0 | 2.303 |
| ST19 | 37°21′50.8″ N, 127°54′21.7″ E | 9 | 7 | 0.833 (0.127) | 2.770 | 2.889 (1.672) | 0.004 (0.002) | −1.000 | −1.252 |
| ST20 | 37°14′46.2″ N, 127°28′07.1″ E | 5 | 4 | 0.900 (0.161) | 3.000 | 1.600 (1.128) | 0.002 (0.002) | −1.094 | −1.405 |
| ST21 | 37°06′04.8″ N, 127°56′24.2″ E | 2 | 2 | 1.000 (0.500) | - | 4.000 (3.162) | 0.005 (0.006) | 0 | 1.386 |
| STL | Laboratory rearing | 19 | 12 | 0.901 (0.059) | 3.193 | 3.064 (1.667) | 0.004 (0.002) | 0.378 | −5.439 |
| Total | 82 | 46 | 0.957 (0.013) | - | 3.566 (1.831) | 0.005 (0.003) | −2.068 | −26.129 |
| Sites | ST1 | ST3 | ST27 | STL |
|---|---|---|---|---|
| ST1 ST3 ST27 STL | - 0.0147 0.0136 0.0076 | 0.0021 - 0.0202 ** 0.0162 * | 0.0578 0.0841 - 0.0129 | 0.0811 ** 0.1215 ** 0.0655 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kang, J.H.; Lim, C.; Park, S.H.; Kim, W.G.; Sareein, N.; Bae, Y.J. Genetic and Morphologic Variation in a Potential Mosquito Biocontrol Agent, Hydrochara Affinis (Coleoptera: Hydrophilidae). Sustainability 2020, 12, 5481. https://0-doi-org.brum.beds.ac.uk/10.3390/su12135481
Kang JH, Lim C, Park SH, Kim WG, Sareein N, Bae YJ. Genetic and Morphologic Variation in a Potential Mosquito Biocontrol Agent, Hydrochara Affinis (Coleoptera: Hydrophilidae). Sustainability. 2020; 12(13):5481. https://0-doi-org.brum.beds.ac.uk/10.3390/su12135481
Chicago/Turabian StyleKang, Ji Hyoun, Changseob Lim, Sung Hwan Park, Wang Gyu Kim, Nattawut Sareein, and Yeon Jae Bae. 2020. "Genetic and Morphologic Variation in a Potential Mosquito Biocontrol Agent, Hydrochara Affinis (Coleoptera: Hydrophilidae)" Sustainability 12, no. 13: 5481. https://0-doi-org.brum.beds.ac.uk/10.3390/su12135481