Linking Organic Metabolites as Produced by Purpureocillium Lilacinum 6029 Cultured on Karanja Deoiled Cake Medium for the Sustainable Management of Root-Knot Nematodes

 , ,
, ,  and
and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Microorganism, Karanja Cake, and Culture Filtrate Preparation
2.2. Nematode Cultures
2.3. Extraction of Culture Filtrate Using Different Solvents
2.4. In Vitro Bioassays of Solvent Fractions
2.4.1. Effect on Juvenile Mortality
2.4.2. Effect on Egg Mass Hatching Inhibition
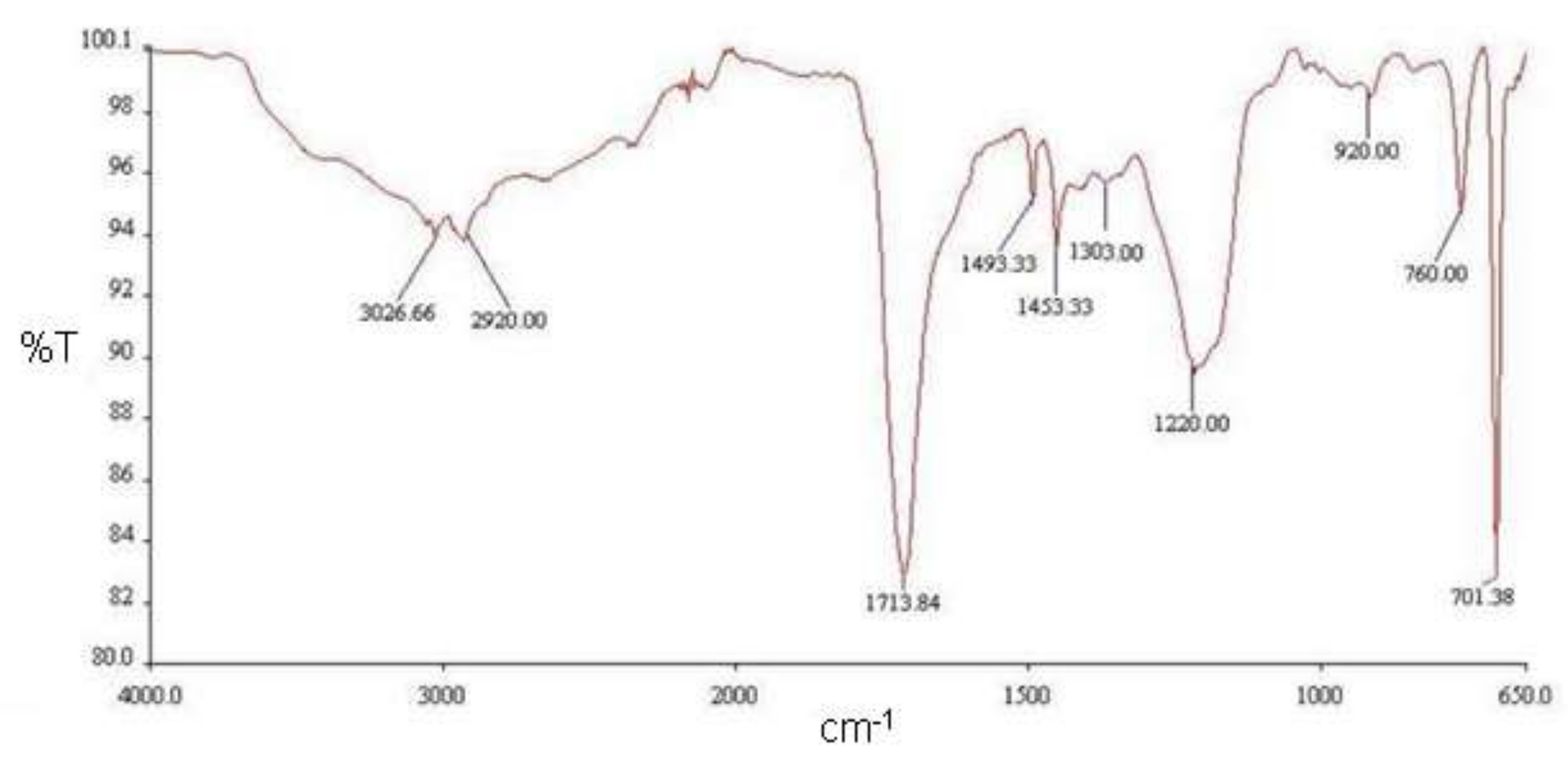
2.5. Fourier Transform Infrared (FT-IR) Spectroscopy Analysis
2.6. Identification of Nematocidal Volatile Compounds
2.7. Statistical Analysis
3. Results
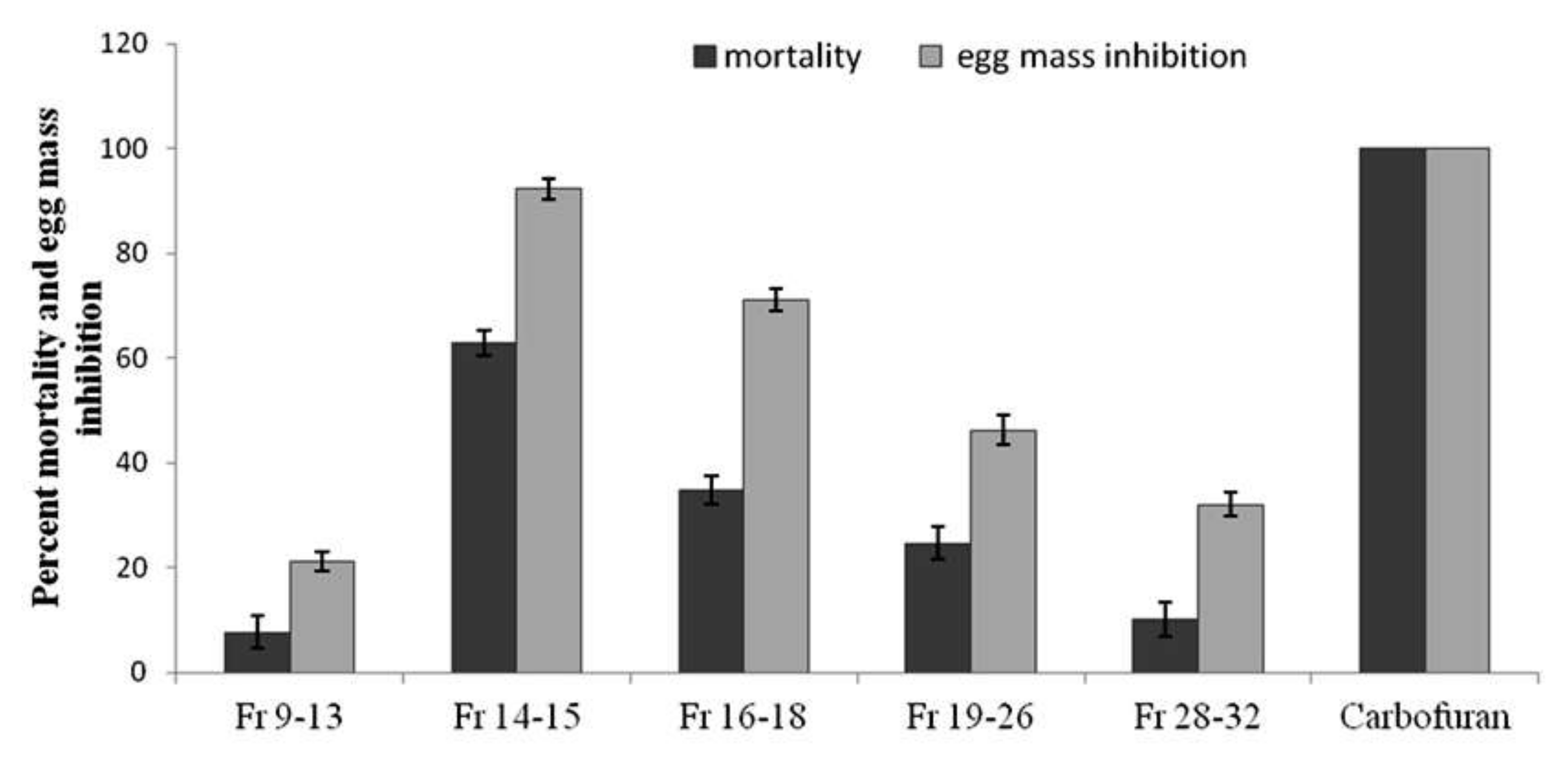
3.1. In Vitro Bioassays of Solvent Fractions of P. lilacinum Filtrate
3.2. Identification of Nematocidal Volatiles
4. Discussion
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Sato, K.; Kadota, Y.; Shirasu, K. Plant immune responses to plant parasitic nematodes. Front. Plant Sci. 2019, 10, 1165. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ibrahim, H.M.M.; Ahmad, E.M.; Martínez-Medina, A.; Aly, M.A.M. Effective approaches to study the plant-root knot nematode interaction. Plant Physiol. Biochem. 2019, 141, 332–342. [Google Scholar] [CrossRef] [PubMed]
- Mhatre, P.H.; Karthik, C.; Kadirvelu, K.; Divya, K.L.; Venkatasalam, E.P.; Srinivasan, S.; Ramkumar, G.; Saranya, C.; Shanmuganathan, R. Plant growth promoting rhizobacteria (PGPR): A potential alternative tool for nematodes bio-control. Biocatal. Agric. Biotechnol. 2019, 17, 119–128. [Google Scholar] [CrossRef]
- Abd-Elgawad, M.M.M.; Askary, T.H. Factors affecting success of biological agents used in controlling the plant-parasitic nematodes. Egypt. J. Biol. Pest Control 2020, 30, 17. [Google Scholar] [CrossRef]
- Poveda, J.; Abril-Urias, P.; Escobar, C. Biological control of plant-parasitic nematodes by filamentous fungi inducers of resistance: Trichoderma, mycorrhizal and endophytic fungi. Front. Microbiol. 2020, 11, 992. [Google Scholar] [CrossRef]
- Da Silva Campos, M.A. Bioprotection by arbuscular mycorrhizal fungi in plants infected with Meloidogyne nematodes: A sustainable alternative. Crop Prot. 2020, 135, 105203. [Google Scholar] [CrossRef]
- Sharapova, I.E. Prospects of using entomopathogenic fungus in development of a biopesticide product with nematicidal activity. Biocatal. Agric. Biotechnol. 2019, 19, 101098. [Google Scholar] [CrossRef]
- Degenkolb, T.; Vilcinskas, A. Metabolites from nematophagous fungi and nematicidal natural products from fungi as an alternative for biological control. Part I: Metabolites from nematophagous ascomycetes. Appl. Microbiol. Biotechnol. 2016, 100, 3799–3812. [Google Scholar] [CrossRef]
- Degenkolb, T.; Vilcinskas, A. Metabolites from nematophagous fungi and nematicidal natural products from fungi as an alternative for biological control. Part II: Metabolites from nematophagous basidiomycetes and non-nematophagous fungi. Appl. Microbiol. Biotechnol. 2016, 100, 3813–3824. [Google Scholar] [CrossRef] [Green Version]
- Laatsch, H. Antibase 2014 SciDex v. 1.2.495—The Natural Compounds Identifier; Wiley-VCH: Weinheim, Germany, 2014. [Google Scholar]
- Prasad, P.; Varshney, D.; Adholeya, A. Whole genome annotation and comparative genomic analyses of bio-control fungus Purpureocillium lilacinum. BMC Genom. 2015, 16, 1004. [Google Scholar] [CrossRef] [Green Version]
- Jatala, P.; Salas, R.; Kaltenback, R.; Bocangel, M. Multiple application and long term effect of Paecilomyces lilacinus in controlling Meloidogyne incognita under field conditions (abstr.). J. Nematol. Soc. Nematol. 1981, 13, 445. [Google Scholar]
- Sikora, R.; Kiewnick, S. Evaluation of Paecilomyces lilacinus strain 251 for the biological control of the northern root-knot nematode Meloidogyne hapla Chitwood. Nematology 2006, 8, 69–78. [Google Scholar]
- Sharma, S.; Verma, M.; Sharma, A. Utilization of non edible oil seed cakes as substrate for growth of Paecilomyceslilacinus and as biopesticide against termites. Waste Biomass Valoriz. 2013, 4, 325–330. [Google Scholar] [CrossRef]
- Sharma, A.; Sharma, S.; Mittal, A.; Naik, S.N. Statistical optimization of growth media for Paecilomyceslilacinus 6029 using non-edible oil cakes. Ann. Microbiol. 2014, 64, 515–520. [Google Scholar] [CrossRef]
- Sharma, A.; Sharma, S.; Dalela, M. Nematicidal activity of Paecilomyces lilacinus 6029 cultured on Karanja cake medium. Microb. Pathog. 2014, 75, 16–20. [Google Scholar] [CrossRef]
- Sharma, A.; Sharma, S.; Yadav, S.; Naik, S.N. Role of Karanja deoiled cake based medium in production of protease and fatty acids by Paecilomyces lilacinus 6029. J. Biosci. Bioeng. 2014, 118, 270–271. [Google Scholar] [CrossRef]
- Sharma, A.; Sharma, S.; Mittal, A.; Naik, S.N. Evidence for the involvement of nematocidal toxins of Purpureocillium lilacinum 6029 cultured on Karanja deoiled cake liquid medium. World J. Microbiol. Biotechnol. 2016, 32, 82. [Google Scholar] [CrossRef]
- Huang, D.; Yu, C.; Shao, Z.; Cai, M.; Li, G.; Zheng, L.; Yu, Z.; Zhang, J. Identification and characterization of nematicidal volatile organic compounds from Deep-Sea Virgibacillus dokdonensis MCCC 1A00493. Molecules 2020, 25, 744. [Google Scholar] [CrossRef] [Green Version]
- Onifade, A.K.; Fatope, M.O.; Deadman, M.L.; Al-Kindy, S.M.Z. Nematicidal activity of Haplophyllumtuberculatum and Plectranthuscylindraceus oils against Meloidogyne javanica. Biochem. Syst. Ecol. 2008, 36, 679–683. [Google Scholar] [CrossRef]
- Kukula-Koch, W.; Grzybek, M.; Strachecka, A.; Jaworska, A.; Ludwiczuk, A. ATR-FTIR-based fingerprinting of some Cucurbitaceae extracts: A preliminary study. Acta Soc. Bot. Pol. 2018, 87, 92. [Google Scholar] [CrossRef]
- Nitao, J.K.; Meyer, S.L.F.; Chitwood, D.J. In-vitro assays of Meloidogyne incognita and Heterodera glycines for detection of nematode-antagonistic fungal compounds. J. Nematol. 1999, 31, 172. [Google Scholar] [PubMed]
- Rawani, A.; Ray, A.S.; Ghosh, A.; Sakar, M.; Chandra, G. Larvicidal activity of phytosteroid compounds from leaf extract of Solanumnigrum against Culexvishnui group and Anopheles subpictus. BMC Res. Notes 2017, 10, 135. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Siddiqui, I.A.; Qureshi, S.A.; Sultana, V.; Ehteshamul-Haque, S.; Ghaffar, A. Biological control of root rot-root knot disease complex of tomato. Plant Soil 2000, 227, 163–169. [Google Scholar] [CrossRef]
- Strobel, G.A.; Dirkse, E.; Sears, J.; Markworth, C. Volatile antimicrobials from Muscodoralbus, a novel endophytic fungus. Microbiology 2001, 147, 2943–2950. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gapasin, R.M.; Vasquez, E.A.; Rendon, G.A. Bioassay-guided identification of the nematicidal secondary metabolites from Paecilomyces lilicanus for the control of root-knot nematode (Meloidogyne graminicola, Golden and Birchfield). Ann. Trop. Res. 2011, 33, 22–43. [Google Scholar] [CrossRef]
- Breinig, S.; Schiltz, E.; Fuchs, G. Genes involved in anaerobic metabolism of phenol in the bacterium Thauera aromatica. J. Bacteriol. 2000, 182, 5849–5863. [Google Scholar] [CrossRef] [Green Version]
- Niu, Q.; Huang, X.; Zhang, L.; Xu, J.; Yang, D.; Wei, K.; Niu, X.; An, Z.; Bennett, J.W.; Zou, C. A Trojan horse mechanism of bacterial pathogenesis against nematodes. Proc. Natl. Acad. Sci. USA 2010, 107, 16631–16636. [Google Scholar] [CrossRef] [Green Version]



{kind=link}
{kind=link}
| Concentration (mg/L) | Ethyl Acetate Extract | Hexane Extract | Aqueous Extract | |||
|---|---|---|---|---|---|---|
| Mortality (%) at 48 h | Egg Mass Hatching Inhibition (%) | Mortality (%) at 48 h | Egg Mass Hatching Inhibition (%) | Mortality (%) at 48 h | Egg Mass Hatching Inhibition (%) | |
| 312.5 | 16.2 ± 2.6 b | 15.9 ± 1.2 b | 0.0 ± 0.0 a | 0.0 ± 0.0 a | 7.3 ± 0.0 a | 11.5 ± 0.0 a |
| 625 | 43.1 ± 1.8 c | 26.5 ± 1.0 c | 0.0 ± 0.0 a | 17.2 ± 1.2 b | 12.6 ± 1.1 b | 23.2 ± 1.9 b |
| 1250 | 76.9 ± 1.8 d | 51.7 ± 1.7 d | 10.8 ±1.8 b | 20.2 ± 1.8 c | 32.5 ± 1.9 c | 41.4 ±1.2 c |
| 2500 | 91.9 ± 2.8 e | 82.3 ± 2.2 e | 17.3 ± 1.6 c | 27.5 ± 1.9 d | 61.2 ± 2.6 d | 68.7 ± 1.8 d |
| 5000 | 100.0 ± 0.0 f | 100.0 ± 0.0 f | 23.9 ±1.2 d | 31.1 ± 1.6 e | 77.3 ± 1.2 e | 80.5 ± 1.3 e |
| 10,000 | 100.0 ± 0.0 f | 100.0 ± 0.0 f | 29.7 ± 1.9 e | 38.2 ± 2.8 f | 89.2 ± 2.1 f | 91.3 ± 0.9 f |
| DW+Tween blank | 0.0 ± 0.0 a | 0.0 ± 0.0 a | 0.0 ± 0.0 a | 0.0 ± 0.0 a | 0.0 ± 0.0 a | 0.0 ± 0.0 a |
| Ethyl acetate blank | 12.1 ± 2.3 b | 11.7 ± 1.1 b | Nd | Nd | 9.9 ± 0.1 a | 13.4 ± 0.2 a |
| Hexane blank | Nd | Nd | 2.1 ± 0.0 a | 12.3 ± 0.2 a | 11.2 ± 0.2 a | 15.3 ± 0.1 a |
| Chemical control ** | 100.0 ± 0.0 f | 100.0 ± 0.0 f | 100.0 ± 0.0 f | 100.0 ± 0.0 g | 100.0 ± 0.0 g | 100.0 ± 0.0 g |
| Toxicity Factor | Ethyl Acetate Extract | Aqueous Extract | ||
|---|---|---|---|---|
| Second Stage Juveniles | Egg Mass Hatching | Second Stage Juveniles | Egg Mass Hatching | |
| LC50 (mg/L) | 781.48 | 1004.58 | 2058.12 | 1556.24 |
| 95% Confidence intervals | (556.72–1040.83) | (725.94–1360.21) | (1785.83–2377.57) | (1333.46–1811.07) |
| LC90 (mg/L) | 2490.56 | 3198.06 | 9575.24 | 8385.22 |
| 95% Confidence intervals | (1748.16–4599.24) | (2192.86–6184.47) | (7571.27–12877.9) | (6545.68–11495.39) |
| Chi-squared (χ2) | 10.929 | 12.136 | 2.528 | 1.318 |
| Major Compounds | Retention Time (min) | Area (%) |
|---|---|---|
| 2-ethyl butyric acid | 8.21 | 0.75 |
| Phenyl ethyl alcohol | 8.75 | 1.55 |
| Benzoic Acid | 9.81 | 1.69 |
| Benzene acetic acid | 12.06 | 6.05 |
| 3,5-Di-t-butyl-phenol | 19.46 | 7.66 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sharma, A.; Gupta, A.; Dalela, M.; Sharma, S.; Sayyed, R.Z.; Enshasy, H.A.E.; Elsayed, E.A. Linking Organic Metabolites as Produced by Purpureocillium Lilacinum 6029 Cultured on Karanja Deoiled Cake Medium for the Sustainable Management of Root-Knot Nematodes. Sustainability 2020, 12, 8276. https://0-doi-org.brum.beds.ac.uk/10.3390/su12198276
Sharma A, Gupta A, Dalela M, Sharma S, Sayyed RZ, Enshasy HAE, Elsayed EA. Linking Organic Metabolites as Produced by Purpureocillium Lilacinum 6029 Cultured on Karanja Deoiled Cake Medium for the Sustainable Management of Root-Knot Nematodes. Sustainability. 2020; 12(19):8276. https://0-doi-org.brum.beds.ac.uk/10.3390/su12198276
Chicago/Turabian StyleSharma, Abhishek, Aditi Gupta, Manu Dalela, Satyawati Sharma, R. Z. Sayyed, Hesham Ali El Enshasy, and Elsayed Ahmed Elsayed. 2020. "Linking Organic Metabolites as Produced by Purpureocillium Lilacinum 6029 Cultured on Karanja Deoiled Cake Medium for the Sustainable Management of Root-Knot Nematodes" Sustainability 12, no. 19: 8276. https://0-doi-org.brum.beds.ac.uk/10.3390/su12198276