
Saved by the Shell: Molecular Analysis Detects the Cryptic Sea Hare, Aplysia concava G. B. Sowerby I, 1833 (Mollusca: Heterobranchia: Aplysiidae), from Oceania, with a Redescription


Abstract
:1. Introduction
2. Experimental Section
2.1. Source of Material
2.2. Molecular Study
2.3. Morphological Study
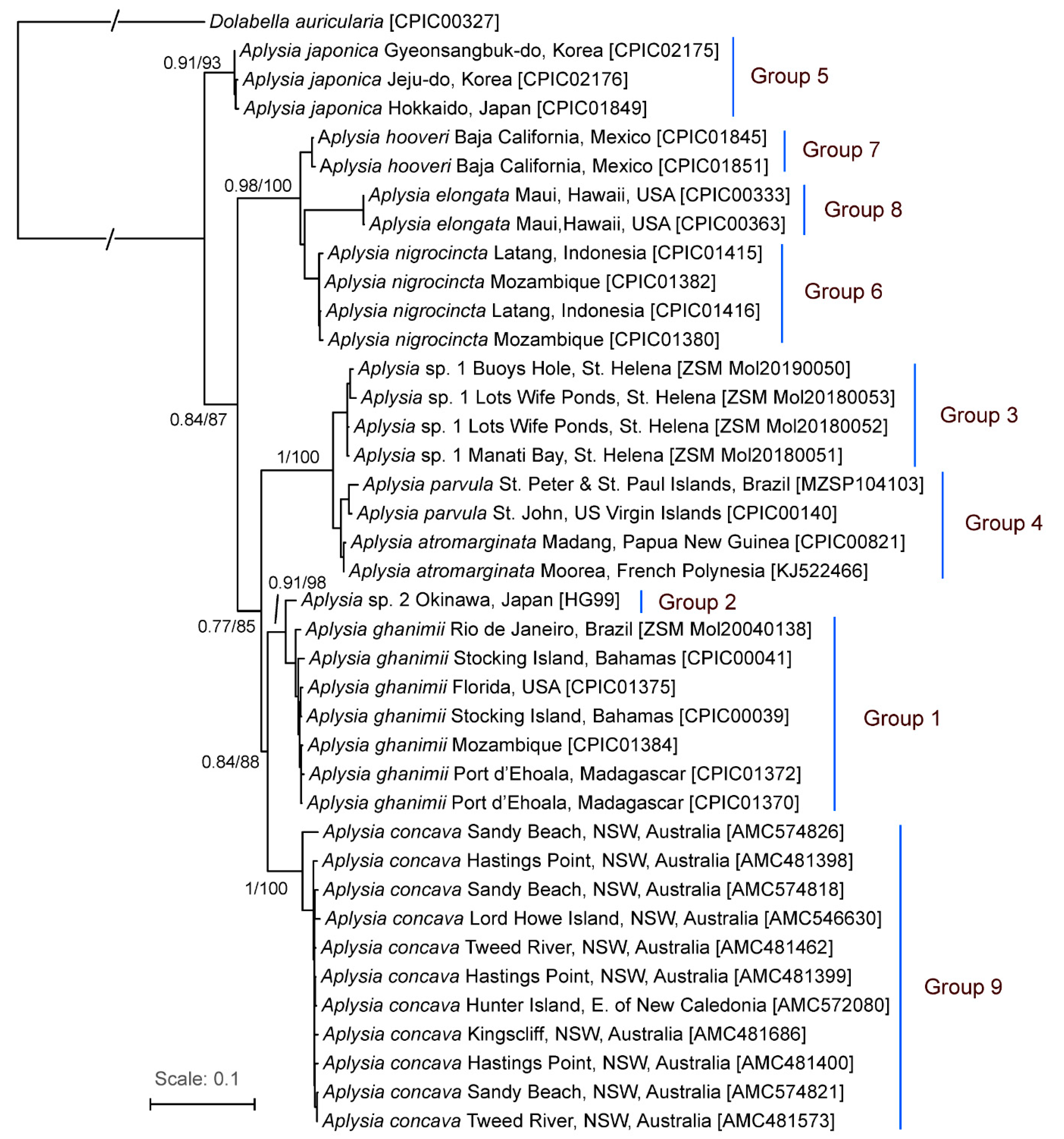
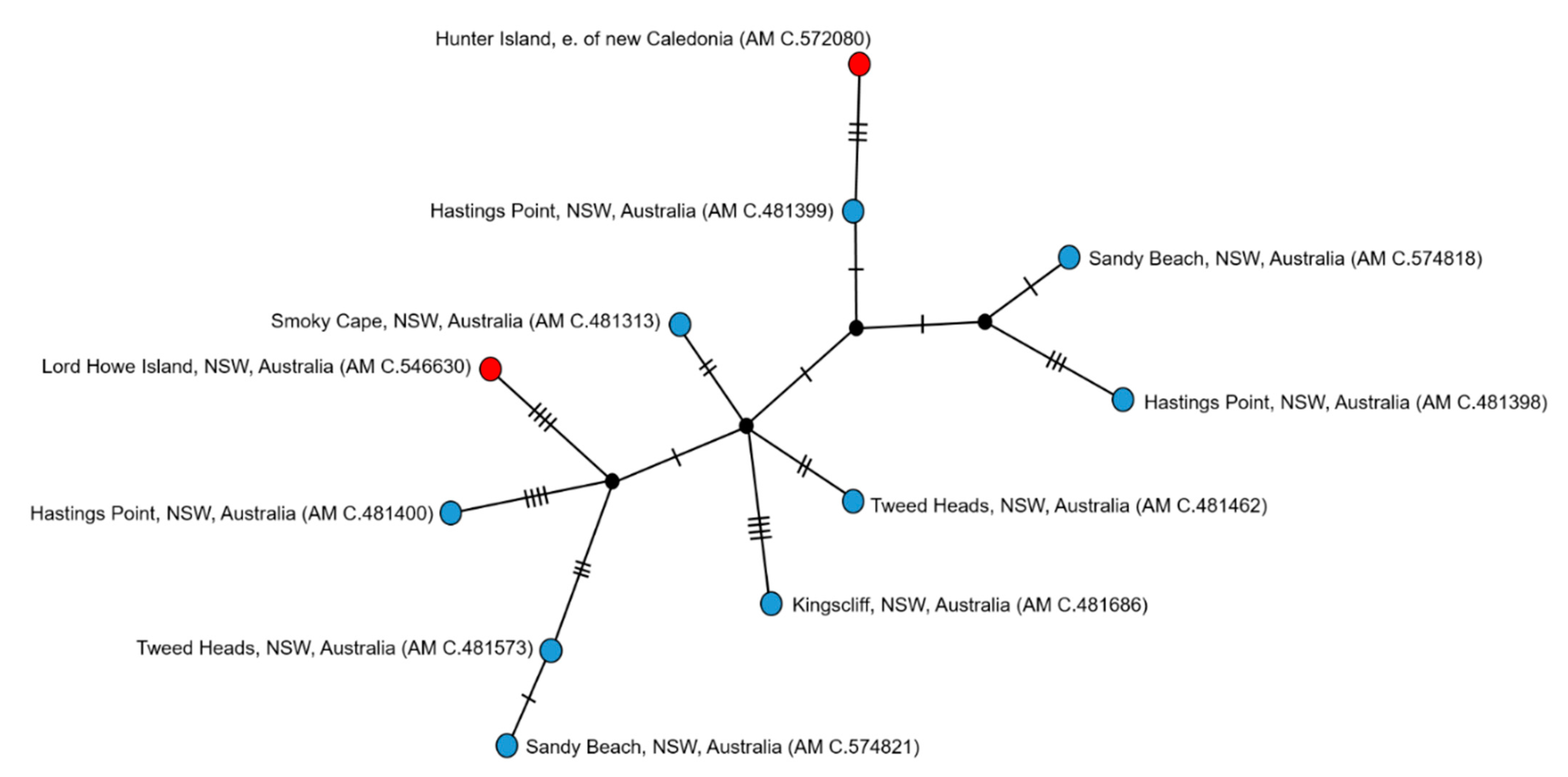
3. Results
3.1. Species Delimitation

3.2. Material Examined
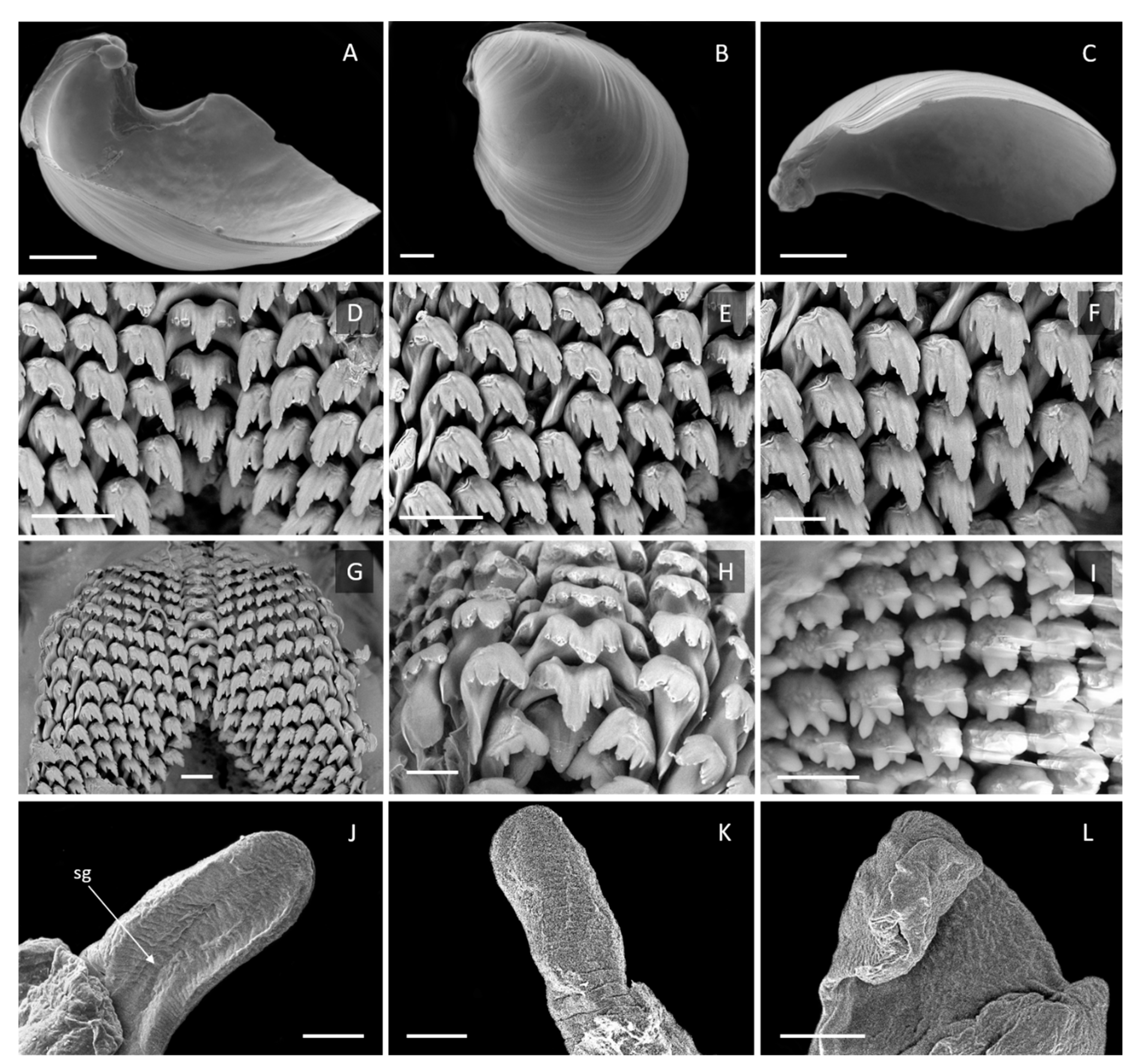
3.3. Description
3.3.1. External Morphology
3.3.2. Colour Variation
3.3.3. Internal Morphology
4. Discussion
4.1. Diagnosis
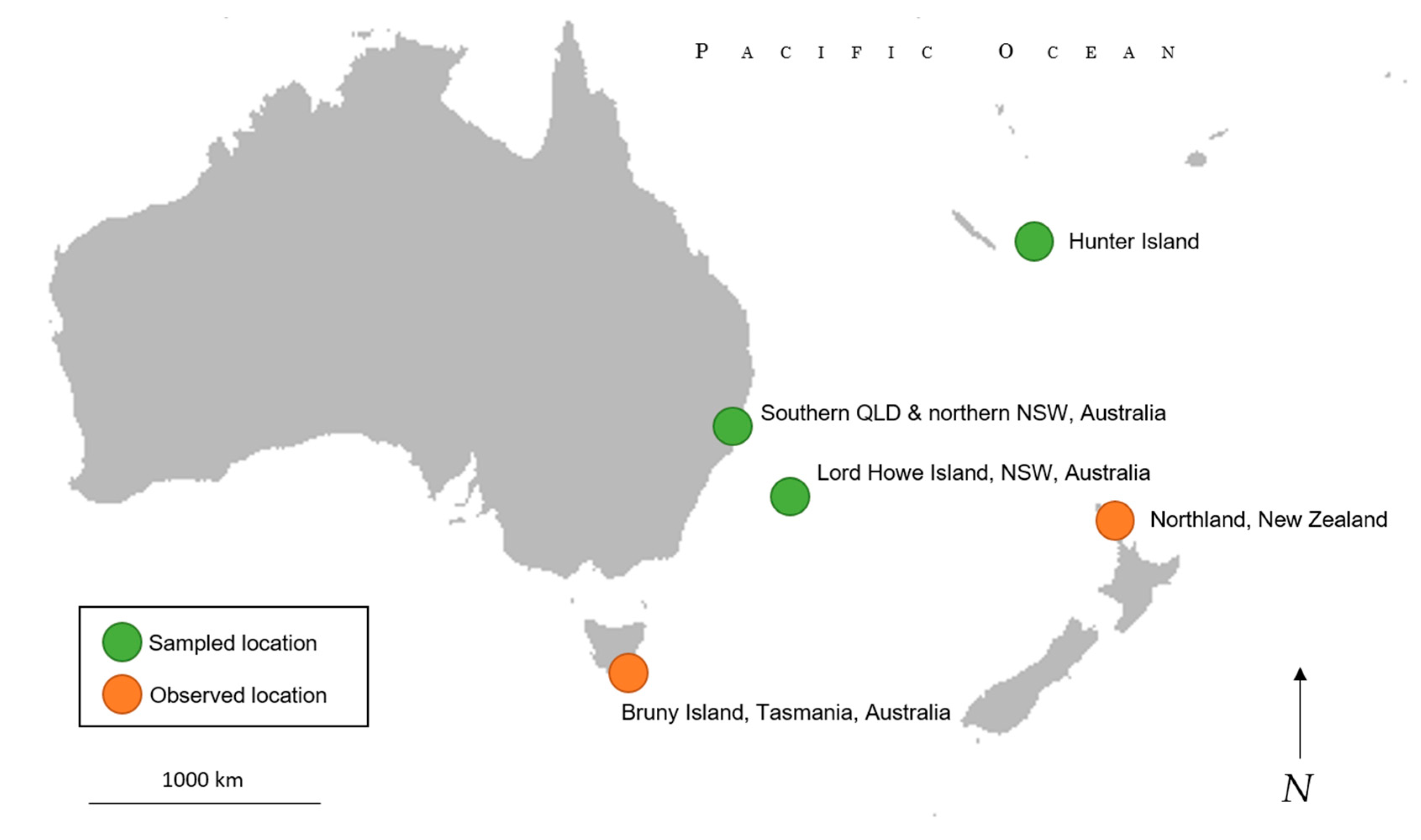
4.2. Range
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Valdés, Á.; Breslau, E.; Padula, V.; Schrödl, M.; Camacho, Y.; Malaquias, M.A.E.; Alexander, J.; Bottomley, M.; Vital, X.G.; Hooker, Y.; et al. Molecular and morphological systematics of Dolabrifera Gray, 1847 (Mollusca: Gastropoda: Heterobranchia: Aplysiomorpha). Zool. J. Linn. Soc. 2017, 1, 31–35. [Google Scholar] [CrossRef] [Green Version]
- Krug, P.J.; Berriman, J.S.; Valdés, Á. Phylogenetic systematics of the shelled sea slug genus Oxynoe Rafinesque, 1814 (Heterobranchia: Sacoglossa), with integrative descriptions of seven new species. Invertebr. Syst. 2018, 32, 950–1003. [Google Scholar] [CrossRef]
- Golestani, H.; Crocetta, F.; Padula, V.; Camacho-García, Y.; Langeneck, J.; Poursanidis, D.; Pola, M.; Yokeş, M.B.; Cervera, J.L.; Jung, D.-W.; et al. The little Aplysia coming of age: From one species to a complex of species complexes in Aplysia parvula (Mollusca: Gastropoda: Heterobranchia). Zool. J. Linn. Soc. 2019, 187, 279–330. [Google Scholar] [CrossRef]
- Nimbs, M.J.; Willan, R.C.; Smith, S.D. Is Port Stephens, eastern Australia, a global hotspot for biodiversity of Aplysiidae (Gastropoda: Heterobranchia)? Molluscan Res. 2017, 37, 45–65. [Google Scholar] [CrossRef]
- Von Clessin, S. Aplysiidae. In Conchylien-Cabinet; Verlag von Bauer & Raspe: Nurnburg, Germany, 1899; p. 22. [Google Scholar]
- Pilsbry, H.A. Philinidae, Gastropteridae, Aglajidae, Aplysiidae, Oxynoeidae, Runcinidae, Umbraculidae, Pleurobranchidae. Man. Conchol. 1896, 16, 262–274. [Google Scholar]
- Eales, N.B. Revision of the world species of Aplysia (Gastropoda, Opisthobranchia). In Bulletin of the British Musem, Zoology; British Museum (Natural History): London, UK, 1960; Volume 5, pp. 267–404. [Google Scholar]
- Tenison-Woods, J.E. Census; with brief descriptions of the marine shells of Tasmania and the adjacent islands. In Papers & Proceedings and Report of the Royal Society of Tasmania; 1877; pp. 26–57. Available online: https://eprints.utas.edu.au/15412/1/tenison-wood-census-marine-shells-1877.pdf (accessed on 9 April 2021).
- Geller, J.; Meyer, C.; Parker, M.; Hawk, H. Redesign of PCR primers for mitochondrial cytochrome c oxidase subunit I for marine invertebrates and application in all-taxa biotic surveys. Mol. Ecol. Resour. 2013, 13, 851–861. [Google Scholar] [CrossRef] [PubMed]
- Palumbi, S.R. Nucleic Acids II: The Polymerase Chain Reaction. In Molecular Systematics; Hillis, D.M., Moritz, C., Mable, B.K., Eds.; Sinauer Associates: Sunderland, MA, USA, 1996; pp. 205–247. [Google Scholar]
- Colgan, D.J.; McLauchlan, A.; Wilson, G.D.F.; Livingston, S.P.; Edgecombe, G.D.; Macaranas, J.; Cassis, G.; Gray, M.R. Histone H3 and U2 snRNA DNA sequences and arthropod molecular evolution. Aust. J. Zool. 1998, 46, 419–437. [Google Scholar] [CrossRef]
- Kearse, M.; Moir, R.; Wilson, A.; Stones-Havas, S.; Cheung, M.; Sturrock, S.; Buxton, S.; Cooper, A.; Markowitz, S.; Duran, C.; et al. Geneious Basic: An integrated and extendable desktop software platform for the organization and analysis of sequence data. Bioinformatics 2012, 28, 1647–1649. [Google Scholar] [CrossRef]
- Katoh, K.; Standley, D.M. MAFFT multiple sequence alignment software version 7: Improvements in performance and usability. Mol. Biol. Evol. 2013, 30, 772–780. [Google Scholar] [CrossRef] [Green Version]
- Johnson, M.; Zaretskaya, I.; Raytselis, Y.; Merezhuk, Y.; McGinnis, S.; Madden, T.L. NCBI BLAST: A better web interface. Nucleic Acids Res. 2008, 36, W5–W9. [Google Scholar] [CrossRef]
- Dereeper, A.; Guignon, V.; Blanc, G.; Audic, S.; Buffet, S.; Chevenet, F.; Dufayard, J.-F.; Guindon, S.; Lefort, V.; Lescot, M.; et al. Phylogeny.fr: Robust phylogenetic analysis for the non-specialist. Nucleic Acids Res. 2008, 36, W465–W469. [Google Scholar] [CrossRef]
- Castresana, J. Selection of Conserved Blocks from Multiple Alignments for Their Use in Phylogenetic Analysis. Mol. Biol. Evol. 2000, 17, 540–552. [Google Scholar] [CrossRef] [Green Version]
- Minh, B.Q.; Nguyen, M.A.T.; Von Haeseler, A. Ultrafast Approximation for Phylogenetic Bootstrap. Mol. Biol. Evol. 2013, 30, 1188–1195. [Google Scholar] [CrossRef]
- Trifinopoulos, J.; Nguyen, L.-T.; Von Haeseler, A.; Minh, B.Q. W-IQ-TREE: A fast online phylogenetic tool for maximum likelihood analysis. Nucleic Acids Res. 2016, 44, W232–W235. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chernomor, O.; Von Haeseler, A.; Minh, B.Q. Terrace Aware Data Structure for Phylogenomic Inference from Supermatrices. Syst. Biol. 2016, 65, 997–1008. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kalyaanamoorthy, S.; Minh, B.Q.; Wong, T.K.F.; Von Haeseler, A.; Jermiin, L.S. ModelFinder: Fast model selection for accurate phylogenetic estimates. Nat. Methods 2017, 14, 587–589. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Medina, M.; Walsh, P.J. Molecular Systematics of the Order Anaspidea Based on Mitochondrial DNA Sequence (12S, 16S, and COI). Mol. Phylogenet. Evol. 2000, 15, 41–58. [Google Scholar] [CrossRef] [Green Version]
- Kumar, S.; Stecher, G.; Tamura, K. MEGA7: Molecular Evolutionary Genetics Analysis Version 7.0 for Bigger Datasets. Mol. Biol. Evol. 2016, 33, 1870–1874. [Google Scholar] [CrossRef] [Green Version]
- Tamura, K.; Nei, M. Estimation of the number of nucleotide substitutions in the control region of mitochondrial DNA in humans and chimpanzees. Mol. Biol. Evol. 1993, 10, 512–526. [Google Scholar] [CrossRef]
- Puillandre, N.; Lambert, A.; Brouillet, S.; Achaz, G. ABGD, Automatic Barcode Gap Discovery for primary species delimitation. Mol. Ecol. 2011, 21, 1864–1877. [Google Scholar] [CrossRef]
- Clement, M.J.; Snell, Q.; Walker, P.; Posada, D.; Crandall, K.A. TCS: Estimating gene genealogies. In Proceedings of the 16th Parallel and Distributed Processing Symposium; IEEE Computer Society: Los Alamitos, CA, USA, 2002; Volume 2, p. 184. [Google Scholar]
- Leigh, J.W.; Bryant, D. Popart: Full-feature software for haplotype network construction. Methods Ecol. Evol. 2015, 6, 1110–1116. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Voucher | Locality | Date | Collected by | Length (mm) | GenBank Accession | ||
|---|---|---|---|---|---|---|---|
| COI | 16S | H3 | |||||
| AMS C.481313 | Smoky Cape, NSW, Australia | 30 February 2014 | J.H. Waterhouse, A.C. Miller, F. Noss, D. Holmes | 9 | - | - | - |
| AMS C.481398 | Hastings Point, NSW, Australia | 27 February 2014 | J.H. Waterhouse, A.C. Miller, F. Noss, D. Holmes | 7 | MT107263 | MT108945 | MW854246 |
| AMS C.481399 | Hastings Point, NSW, Australia | 27 February 2014 | J.H. Waterhouse, A.C. Miller, F. Noss, D. Holmes | 19 | MT107259 | MT108948 | MW854247 |
| AMS C.481400 | Hastings Point, NSW, Australia | 27 February 2014 | J.H. Waterhouse, A.C. Miller, F. Noss, D. Holmes | 16 | MT107267 | MT108950 | MW854248 |
| AMS C.481462 | Tweed Heads, NSW, Australia | 28 February 2014 | J.H. Waterhouse, A.C. Miller, F. Noss, D. Holmes | 7 | MT107262 | MT108947 | MW854249 |
| AMS C.481573 | Tweed Heads, NSW, Australia | 28 February 2014 | J.H. Waterhouse, A.C. Miller, F. Noss, D. Holmes | 12 | MT107265 | MT108951 | MW854250 |
| AMS C.481686 | Kingscliff, NSW, Australia | 03 March 2014 | J.H. Waterhouse, A.C. Miller, F. Noss, D. Holmes | 14 | MT107264 | MT108944 | MW854251 |
| AMS C.546630 | Lord Howe Island, NSW, Australia | 04 April 2017 | K. Layton, A. Reid | 9 | MT107268 | MT108943 | MW854252 |
| AMS C.572080 | Hunter Island, e. of New Caledonia | 04 August 2017 | S. Hannam, A. Reid, E.K. Kupriyanova, L. Vogel, I. Middleton | 16 | MT107261 | MT108949 | MW854253 |
| AMS C.574818 | Sandy Beach, NSW, Australia | 04 March 2016 | S. D. A. Smith | 7 | MT107260 | MT108946 | MW854254 |
| AMS C.574821 | Sandy Beach, NSW, Australia | 06 April 2016 | M. Nimbs | 8 | MT107266 | MT108953 | MW854255 |
| AMS C.574820 | Newcastle, NSW, Australia | 14 December 2016 | S. D. A. Smith | 14 | - | - | - |
| AMS C.574826 | Sandy Beach, NSW, Australia | 08 January 2019 | M. Nimbs | 7 | MT107269 | MT108942 | MW854256 |
| WAM S.29698 | Korff’s Islet, Coffs Harbour, NSW, Australia | 24 May 2019 | M. Nimbs | 12 | - | - | - |
| Species | Locality | Date | Voucher | GenBank Accession Number | ||
|---|---|---|---|---|---|---|
| COI | 16S | H3 | ||||
| Aplysia atromarginata | Moorea, French Polynesia | - | - | KJ522466 | - | - |
| Aplysia atromarginata | Madang, Papua New Guinea | 06 December 2012 | CPIC 00821 | MK422836 | MK422738 | MK422627 |
| Aplysia elongata | Maliko Bay, Maui, Hawaii, USA | 16 January 2011 | CPIC 00333 | MK422876 | MK422773 | MK422669 |
| Aplysia elongata | Maliko Bay, Maui, Hawaii, USA | 18 June 2011 | CPIC 00363 | MK422875 | MK422772 | MK422668 |
| Aplysia ghanimii | Florida, USA | 14 April 2013 | CPIC 01375 | MK422853 | MK422752 | MK422645 |
| Aplysia ghanimii | Mozambique | CPIC 01384 | MK422856 | MK422755 | MK422648 | |
| Aplysia ghanimii | Port d’Ehoala, Madagascar | 15 May 2010 | CPIC 01370 | MK422857 | MK422756 | MK422649 |
| Aplysia ghanimii | Port d’Ehoala, Madagascar | May 2010 | CPIC 01372 | MK422858 | MK422757 | MK422650 |
| Aplysia ghanimii | Rio de Janeiro, Brazil | 29 November 2002 | ZSM Mol 20040138 | MK422863 | MK422761 | MK422655 |
| Aplysia ghanimii | Stocking Island, Bahamas | 19 January 2007 | CPIC 00039 | MK422848 | MK422750 | MK422639 |
| Aplysia ghanimii | Stocking Island, Bahamas | 19 January 2007 | CPIC 00041 | MK422847 | MK422749 | MK422638 |
| Aplysia hooveri | Baja California, Mexico | 01 September 2015 | CPIC 01845 | MK422878 | MK422777 | MK422673 |
| Aplysia hooveri | Baja California, Mexico | 01 September 2015 | CPIC 01851 | MK422877 | MK422776 | MK422672 |
| Aplysia japonica, | Gyeongsangbuk-do, Korea | 24 August 2011 | CPIC 02175 | MK422869 | MK422764 | MK422661 |
| Aplysia japonica | Hokkaido, Japan | 12 September 2014 | CPIC 01849 | MK422867 | MK422763 | MK422659 |
| Aplysia japonica | Jeju-do, Korea | 28 April 2013 | CPIC 02176 | MK422870 | MK422765 | MK422662 |
| Aplysia nigrocincta | Latang, Indonesia | 11 March 2014 | CPIC 01415 | MK422888 | MK422788 | MK422687 |
| Aplysia nigrocincta | Latang, Indonesia | 11 March 2014 | CPIC 01416 | MK422889 | MK422789 | MK422688 |
| Aplysia nigrocincta | Mozambique | CPIC 01380 | MK422885 | MK422785 | MK422684 | |
| Aplysia nigrocincta | Mozambique | CPIC 01382 | MK422886 | MK422786 | MK422685 | |
| Aplysia parvula | St. John, U.S. Virgin Is. | 15 April 2006 | CPIC 00140 | MK422834 | MK422735 | MK422625 |
| Aplysia parvula | St. Peter and St. Paul Is., Brazil | 03 May 2011 | MZSP 104103 | MK422835 | MK422737 | MK422626 |
| Aplysia sp. 1 | Buoys Hole, St. Helena | 02 October 2013 | ZSM Mol 20190050 | MK422838 | MK422740 | MK422629 |
| Aplysia sp. 1 | Lots Wife Ponds, St. Helena | 22 January 2014 | ZSM Mol 20180052 | MK422840 | MK422742 | MK422631 |
| Aplysia sp. 1 | Lots Wife Ponds, St. Helena | 22 January 2014 | ZSM Mol 20180053 | MK422841 | MK422743 | MK422632 |
| Aplysia sp. 1 | Manati Bay, St. Helena | 14 October 2013 | ZSM Mol 20180051 | MK422839 | MK422741 | MK422630 |
| Aplysia sp. 2 | Okinawa, Japan | 15 March 1987 | HG99 | MK422864 | MK422762 | MK422656 |
| Dolabella auricularia | Napili Bay, Maui, Hawaii, USA | 15 June 2011 | CPIC 00327 | MF669619 | MF669573 | MF669657 |
| Aplysia sp. 2 | Aplysia sp. 1 | A. parvula | A. nigrocincta | A. japonica | A. hooveri | A. ghanimii | A. elongata | A. concava | A. atromarginata | |
|---|---|---|---|---|---|---|---|---|---|---|
| Aplysia sp. 2 | 0.03 | |||||||||
| Aplysia sp. 1 | 0.11 | 0.01 | ||||||||
| A. parvula | 0.12 | 0.06 | 0.02 | |||||||
| A. nigrocincta | 0.13 | 0.14 | 0.12 | 0.01 | ||||||
| A. japonica | 0.12 | 0.13 | 0.14 | 0.12 | 0.01 | |||||
| A. hooveri | 0.12 | 0.13 | 0.12 | 0.05 | 0.11 | 0.01 | ||||
| A. ghanimii | 0.04 | 0.09 | 0.11 | 0.11 | 0.11 | 0.11 | 0.01 | |||
| A. elongata | 0.13 | 0.11 | 0.11 | 0.12 | 0.12 | 0.08 | 0.12 | 0.00 | ||
| A. concava | 0.09 | 0.12 | 0.12 | 0.11 | 0.13 | 0.10 | 0.12 | 0.14 | 0.02 | |
| A. atromarginata | 0.11 | 0.04 | 0.03 | 0.12 | 0.13 | 0.10 | 0.10 | 0.10 | 0.12 | 0.01 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Nimbs, M.J.; Wilson, N.G. Saved by the Shell: Molecular Analysis Detects the Cryptic Sea Hare, Aplysia concava G. B. Sowerby I, 1833 (Mollusca: Heterobranchia: Aplysiidae), from Oceania, with a Redescription. Taxonomy 2021, 1, 48-59. https://0-doi-org.brum.beds.ac.uk/10.3390/taxonomy1020006
Nimbs MJ, Wilson NG. Saved by the Shell: Molecular Analysis Detects the Cryptic Sea Hare, Aplysia concava G. B. Sowerby I, 1833 (Mollusca: Heterobranchia: Aplysiidae), from Oceania, with a Redescription. Taxonomy. 2021; 1(2):48-59. https://0-doi-org.brum.beds.ac.uk/10.3390/taxonomy1020006
Chicago/Turabian StyleNimbs, Matt J., and Nerida G. Wilson. 2021. "Saved by the Shell: Molecular Analysis Detects the Cryptic Sea Hare, Aplysia concava G. B. Sowerby I, 1833 (Mollusca: Heterobranchia: Aplysiidae), from Oceania, with a Redescription" Taxonomy 1, no. 2: 48-59. https://0-doi-org.brum.beds.ac.uk/10.3390/taxonomy1020006