
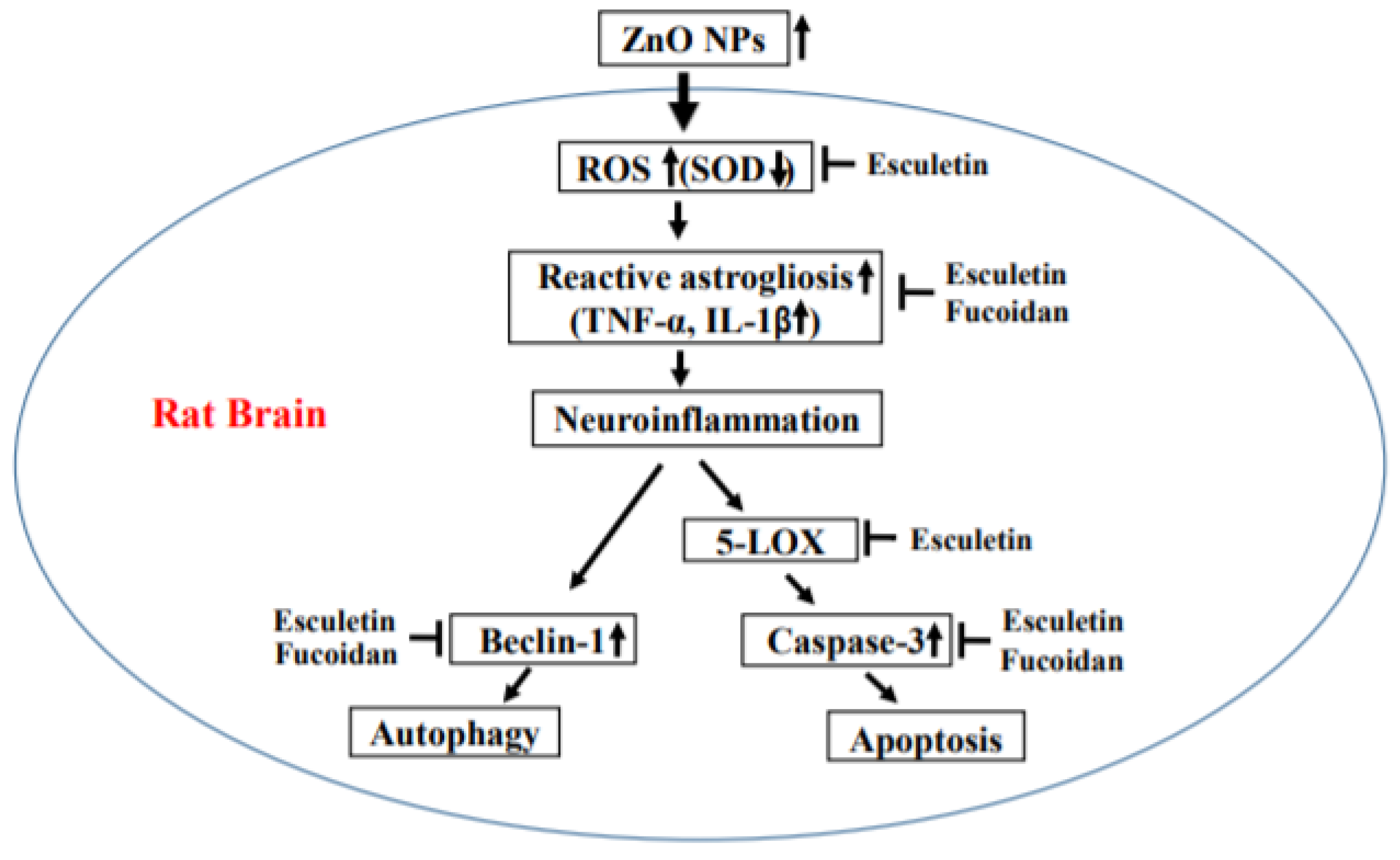
Esculetin and Fucoidan Attenuate Autophagy and Apoptosis Induced by Zinc Oxide Nanoparticles through Modulating Reactive Astrocyte and Proinflammatory Cytokines in the Rat Brain

 ,
,  , , and
, , and 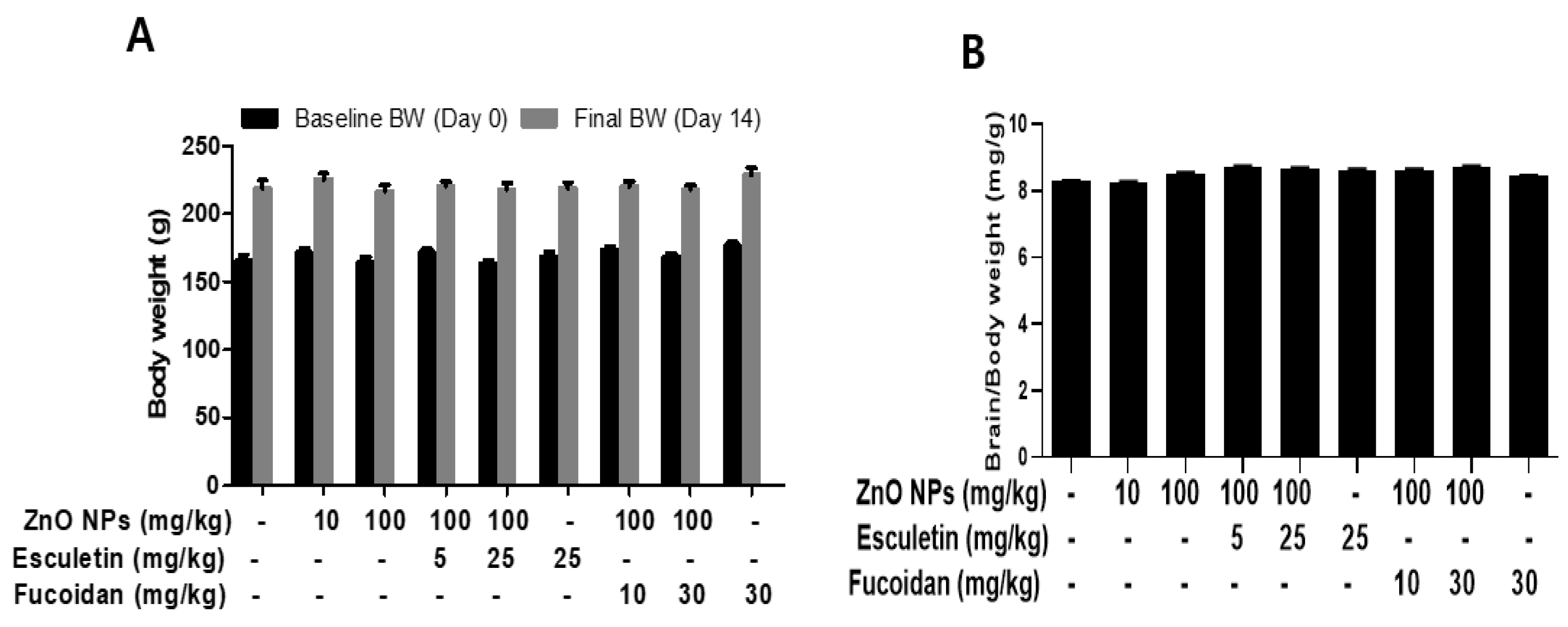{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Material and Methods
2.1. Animals and Test Chemicals
2.2. Production and Characterization of ZnO NPs
2.3. Assessment of Zinc Content
2.4. ELISA Assay
2.5. Protein Assay
2.6. Transmission Electron Microscopy
2.7. Tissue Preparation and Histological Examination
2.8. Immunohistochemistry
2.9. Statistical Analysis
3. Results and Discussion
3.1. Changes in the Body and Organ Weights and Brain Zinc Concentration by ZnO NP Administration after Pre-Treatment of Esculetin or Fucoidan
3.2. Changes in Serum Proinflammatory Cytokines, IL-1β and TNF-α, by ZnO NP Administration after Pre-Treatment of Esculetin or Fucoidan
3.3. Changes in Serum CAT, SOD, 3-NT, and OHdG after ZnO NP Administration after Pre-Treatment of Esculetin or Fucoidan
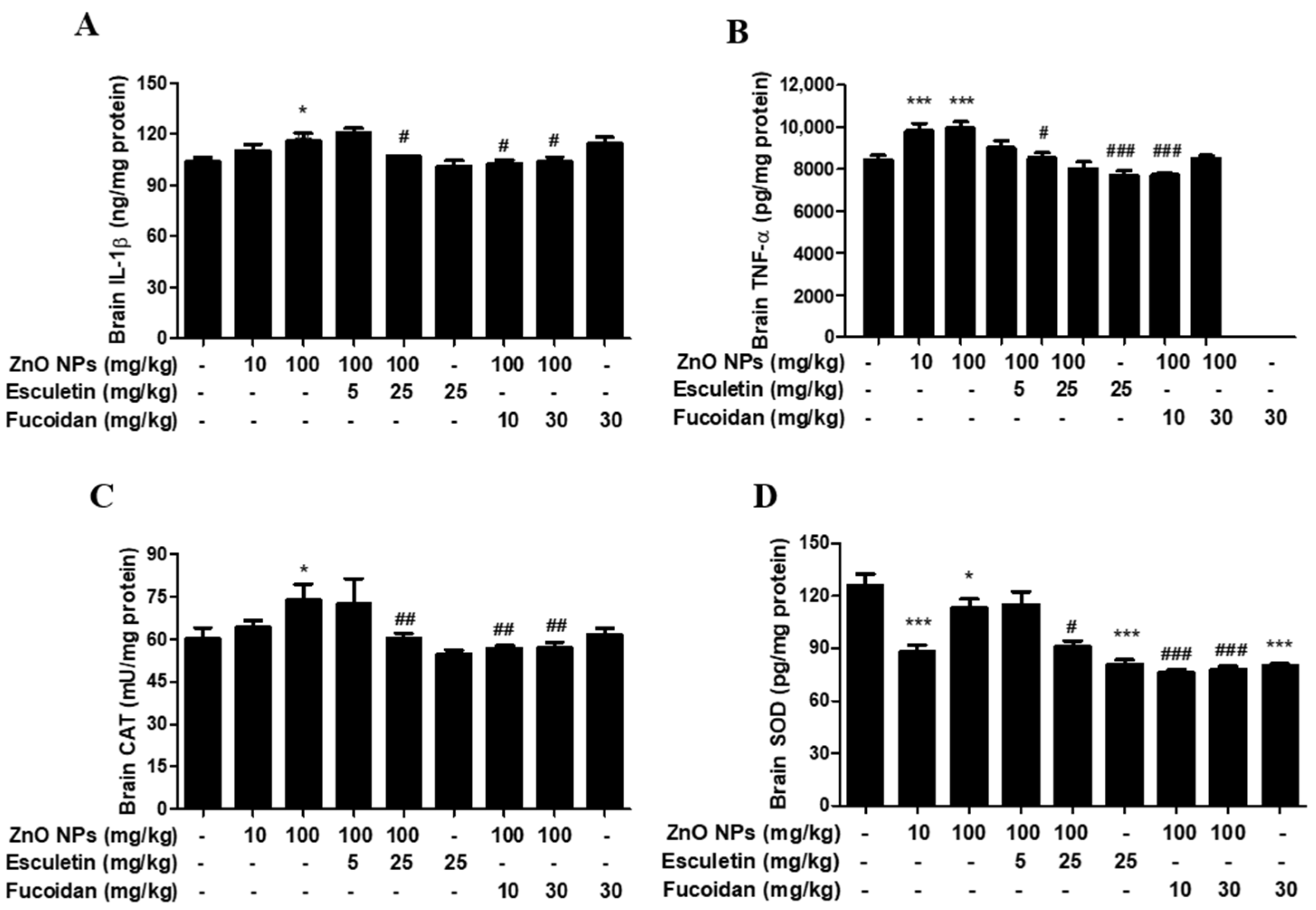
3.4. Changes in Brain Proinflammatory Cytokines and Antioxidant Enzymes by ZnO NP Administration of Pre-Treatment of Esculetin or Fucoidan
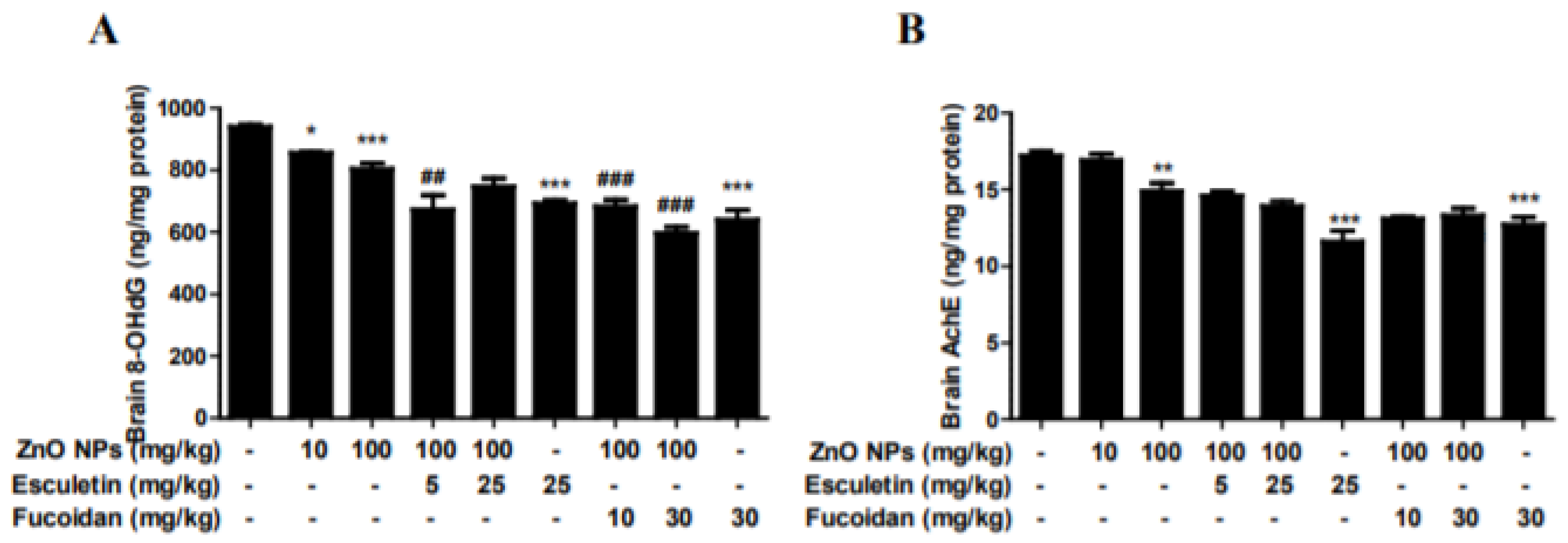
3.5. Changes in Brain 8-OHdG and AChE by ZnO NP Administration of Pre-Treatment of Esculetin or Fucoidan
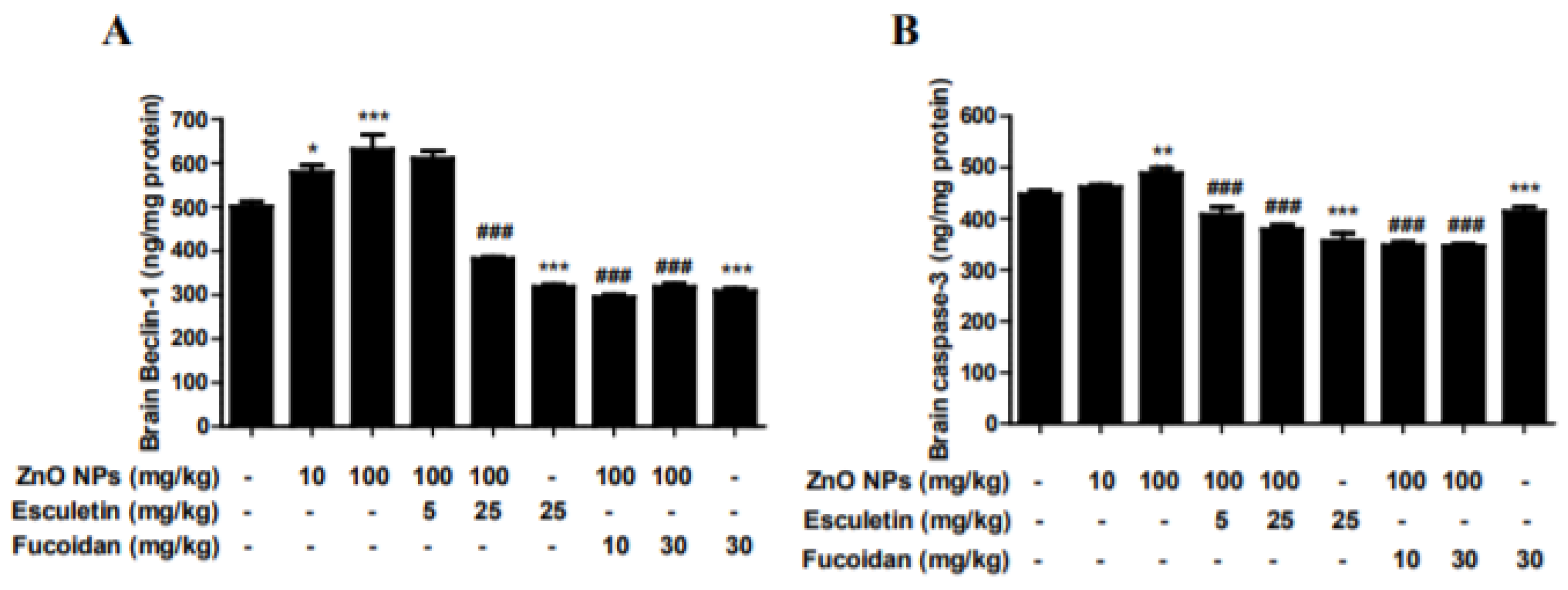
3.6. Changes in Brain Autophagy and Apoptosis by ZnO NP Administration after Pre-Treatment of Esculetin or Fucoidan
3.7. Changes in Brain TEM Findings by ZnO NP Treatment
3.8. Histological Examination of the Hippocampus with H&E Staining by ZnO NPadministration after Pre-Treatment of Esculetin or Fucoidan
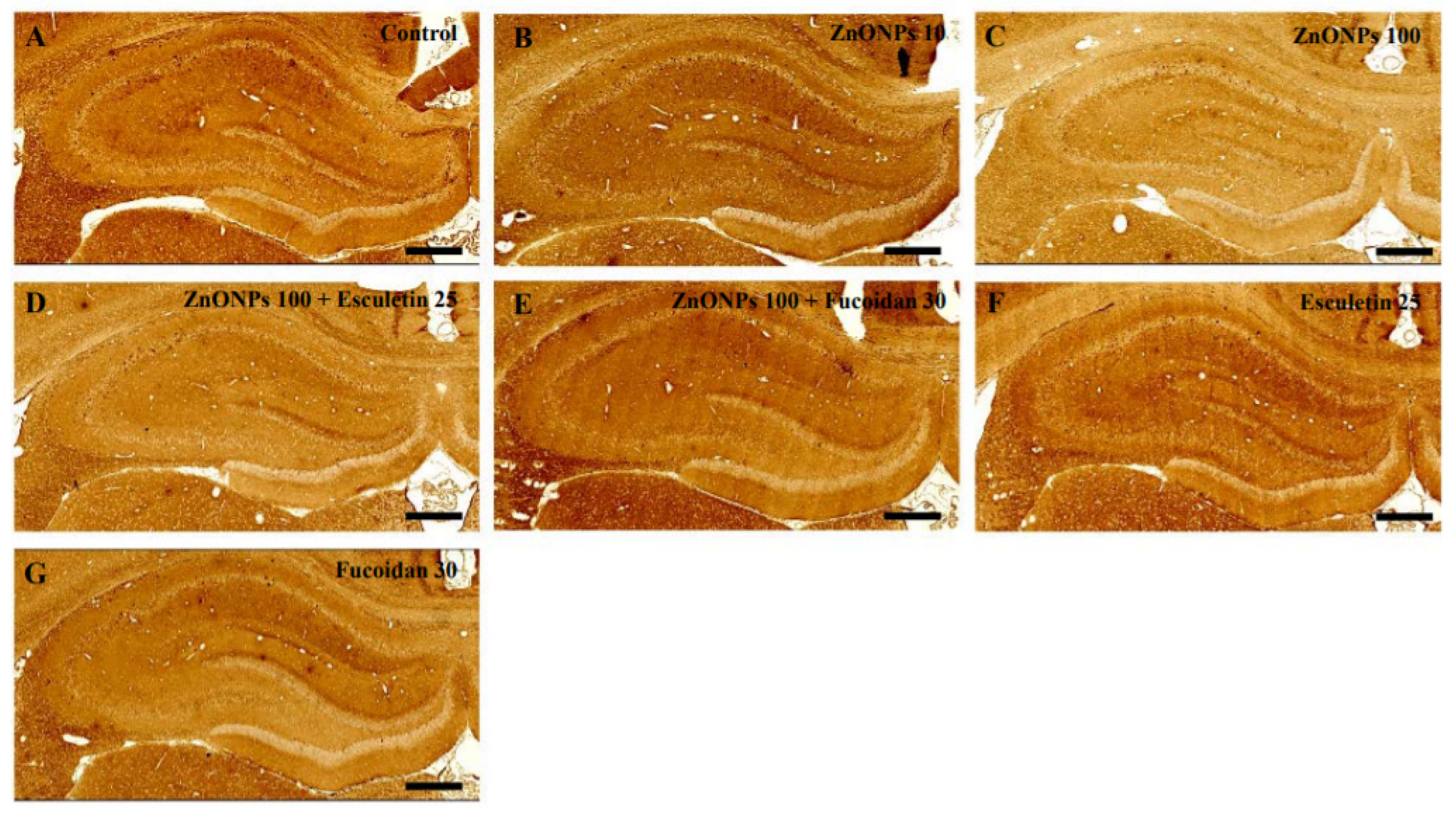
3.9. Histological Examination of the Hippocampus with Anti-NSE Antibody Staining by ZnO NP Administration after Pre-Treatment of Esculetin or Fucoidan
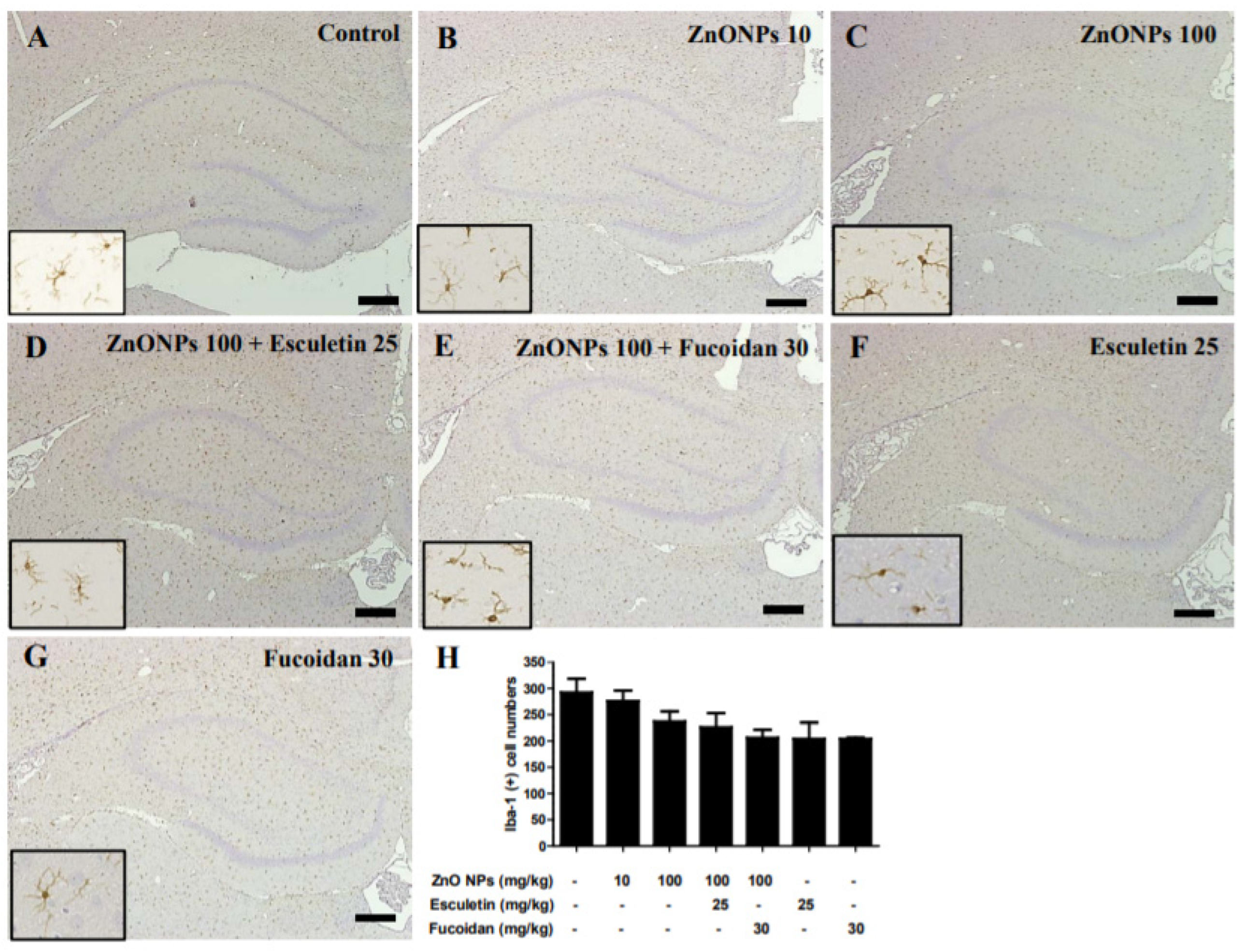
3.10. Histological Examination of the Hippocampus with Anti-Iba-1 Antibody by ZnO NP Administration after Pre-Treatment of Esculetin or Fucoidan
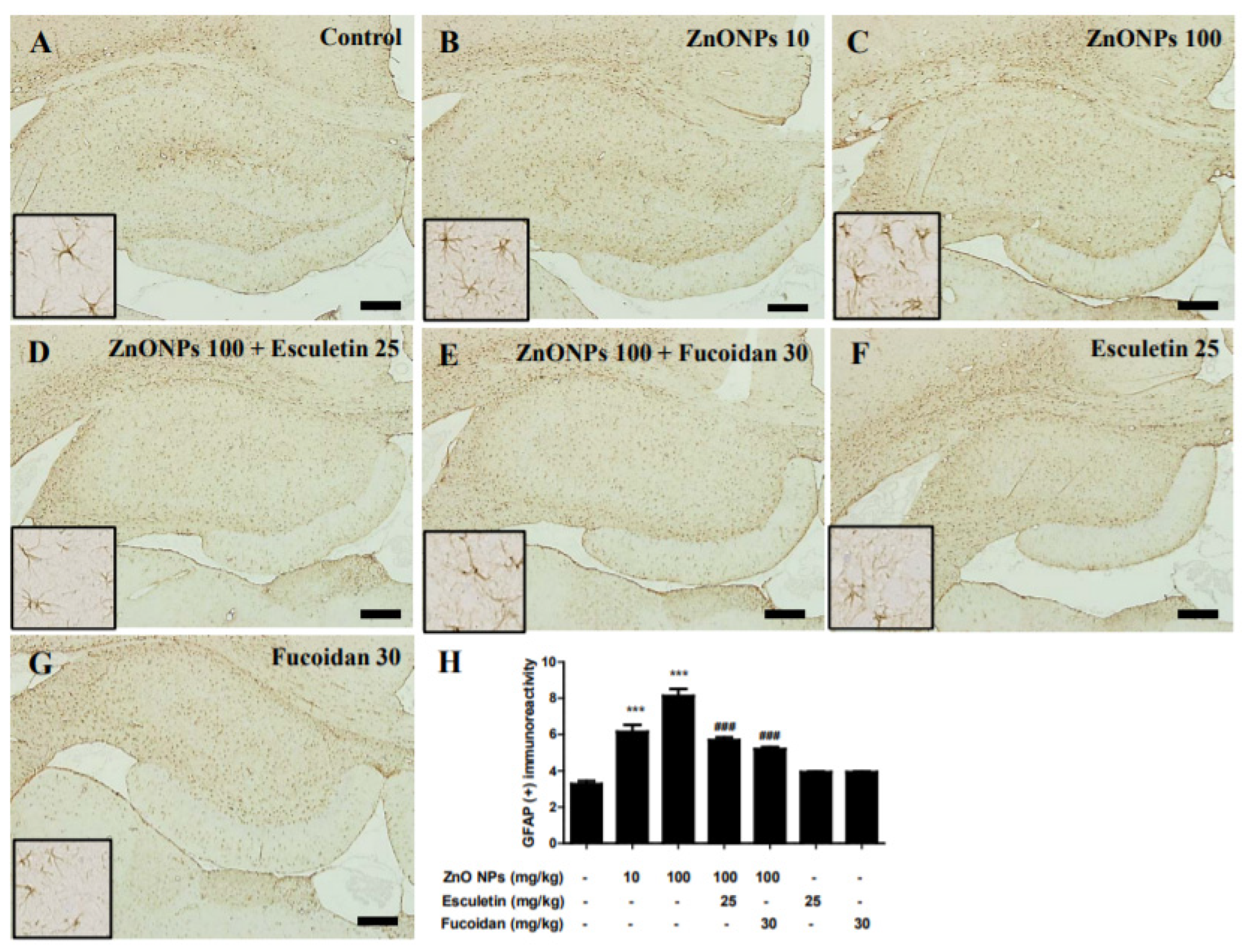
3.11. Immunohistological Examination of the Hippocampus with Anti-GFAP Antibody by ZnO NP Administration after Pre-Treatment of Esculetin or Fucoidan
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Nel, A.; Xia, T.; Mädler, L.; Li, N. Toxic potential of materials at the nanolevel. Science 2006, 311, 622–627. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Najahi-Missaoui, W.; Arnold, R.D.; Cummings, B.S. Safe nanoparticles: Are we there yet? Int. J. Mol. Sci. 2021, 22, 385. [Google Scholar] [CrossRef] [PubMed]
- Gupta, R.; Xie, H. Nanoparticles in daily life: Applications, toxicity and regulations. J. Environ. Pathol. Toxicol. Oncol. 2018, 37, 209–230. [Google Scholar] [CrossRef] [PubMed]
- Guo, Z.; Zhang, P.; Chakraborty, S.; Chetwynd, A.J.; Monikh, F.A.; Stark, C.; Ali-Boucetta, H.; Wilson, S.; Lynch, I.; Valsami-Jones, E. Biotransformation modulates the penetration of metallic nanomaterials across an artificial blood-brain barrier model. Proc. Natl. Acad. Sci. USA 2021, 118, e2105245118. [Google Scholar] [CrossRef]
- Grabrucker, A.M.; Ruozi, B.; Bellettti, D.; Pederzoli, F.; Forni, F.; Vandelli, M.A.; Tosi, G. Nanoparticle transport across the blood brain barrier. Tissue Barriers 2016, 4, e1153568. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Boyes, W.K.; Thriel, C.V. Neurotoxicology of nanomateri als. Chem. Res. Toxicol. 2020, 33, 1121–1144. [Google Scholar] [CrossRef] [PubMed]
- Yarjanli, Z.; Ghaedi, K.; Esmaeili, A.; Rahgozar, S.; Zarrabi, A. Iron oxide nanoparticles may damage to the neural tissue through iron accumulation, oxidative stress, and protein aggregation. BMC Neurosci. 2017, 18, 51. [Google Scholar] [CrossRef] [Green Version]
- Liu, G.; Men, P.; Kudo, W.; Perry, G.; Smith, M.A. Nanoparticle-chelator conjugates as inhibitors of amyloid-β aggregation and neurotoxicity: A Novel therapeutic approach for Alzheimer’s disease. Neurosci. Lett. 2009, 455, 187–190. [Google Scholar] [CrossRef] [Green Version]
- Sarwar, S.; Ali, A.; Pal, M.; Chakrabarti, P. Zinc oxide nanoparticles provide anti-cholera activity by disrupting the interaction of cholera toxin with the human GM1 receptor. J. Biol. Chem. 2017, 292, 18303–18311. [Google Scholar] [CrossRef] [Green Version]
- Asai, T. Nanoparticle-mediated delivery of anticancer agents to tumor angiogenic vessels. Biol. Pharm. Bull. 2012, 35, 1855–1861. [Google Scholar] [CrossRef] [Green Version]
- Wang, Y.; Yuan, L.; Yao, C.; Ding, L.; Li, C.; Fang, J.; Sui, K.; Liu, Y.; Wu, M. A combined toxicity study of zinc oxide nanoparticles and vitamin C in food additives. Nanoscale 2014, 6, 15333–15342. [Google Scholar] [CrossRef] [PubMed]
- Schneider, S.L.; Lim, H.W. A review of inorganic UV filters zinc oxide and titanium dioxide. Photodermatol. Photoimmunol. Photomed. 2019, 35, 442–446. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tang, K.S. The current and future perspectives of zinc oxide nanoparticles in the treatment of diabetes mellitus. Life Sci. 2019, 239, 117011. [Google Scholar] [CrossRef] [PubMed]
- Shiraishi, K.; Nakazawa, S.; Ito, H. Zinc enhances kainate neurotoxicity in the rat brain. Neurol. Res. 1993, 15, 113–116. [Google Scholar] [CrossRef] [PubMed]
- Galasso, S.L.; Dyck, R.H. The role of zinc in cerebral ischemia. Mol. Med. 2007, 13, 380–387. [Google Scholar] [CrossRef]
- Zhao, Y.; Pan, R.; Li, S.; Luo, Y.; Yan, F.; Yin, J.; Qi, Z.; Yan, Y.; Xi, X.; Liu, K.J. Chelating intracellularly accumulated zinc decreased ischemic brain injury through reducing neuronal apoptotic death. Stroke 2014, 45, 1139–1147. [Google Scholar] [CrossRef] [Green Version]
- Cuajungco, M.P.; Lees, G.J. Zinc and Alzheimer’s disease: Is there a direct link? Brain Res. Brain Res. Rev. 1997, 23, 219–236. [Google Scholar] [CrossRef]
- Narayanan, S.E.; Rehuman, N.A.; Harilal, S.; Vincent, A.; Rajamma, R.G.; Behl, T.; Uddin, M.S.; Ashraf, G.M. Molecular mechanism of zinc neurotoxicity in Alzheimer’s disease. Environ. Sci. Pollut. Res. Int. 2020, 27, 43542–43552. [Google Scholar] [CrossRef]
- Pan, E.; Zhang, X.A.; Huang, Z.; Krezel, A.; Zhao, M.; Tinberg, C.E.; Lippard, S.J.; Mcnamara, J.O. Vesicular zinc promotes presynaptic and inhibits postsynaptic long-term potentiation of mossy fiber-CA3 synapse. Neuron 2011, 71, 1116–1126. [Google Scholar] [CrossRef] [Green Version]
- Ceccom, J.; Hally, H.; Daumas, S.; Lassalle, J.M. A specific role for hippocampal mossy fiber’s zinc in rapid storage of emotional memories. Learn. Mem. 2014, 21, 287–297. [Google Scholar] [CrossRef] [Green Version]
- Attia, H.; Nounou, H.; Shalaby, M. Zinc oxide nanoparticles induced oxidative DNA damage, inflammation and apoptosis in rat’s brain after oral exposure. Toxics 2018, 6, 29. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Farokhcheh, M.; Hejazian, L.; Akbarnejad, Z.; Pourabdolhossein, F.; Hosseini, S.M.; Mehraei, T.M.; Soltanpour, N. Geraniol improved memory impairment and neurotoxicity induced by zinc oxide nanoparticles in male wistar rats through its antioxidant effect. Life Sci. 2021, 282, 119823. [Google Scholar] [CrossRef] [PubMed]
- Xiaoli, F.; Junrong, W.; Xuan, L.; Yanli, Z.; Limin, W.; Jia, L.; Longquan, S. Prenatal exposure to nanosized zinc oxide in rats: Neurotoxicity and postnatal impaired learning and memory ability. Nanomedicine 2017, 12, 777–795. [Google Scholar] [CrossRef] [PubMed]
- Tian, L.; Lin, B.; Wu, L.; Li, K.; Liu, H.; Yan, J.; Liu, X.; Xi, Z. Neurotoxicity induced by zinc oxide nanoparticles: Age-related differences and interaction. Sci. Rep. 2015, 5, 16117. [Google Scholar] [CrossRef] [Green Version]
- Han, D.; Tian, Y.; Zhang, T.; Ren, G.; Yang, Z. Nano-zinc oxide damages spatial cognition capability via over-enhanced long-term potentiation in hippocampus of Wistar rats. Int. J. Nanomedicine 2011, 6, 1453–1461. [Google Scholar]
- Amer, M.G.; Karam, R.A. Morphological and biochemical features of cerebellar cortex after exposure to zinc oxide nanoparticles: Possible protective role of curcumin. Anat. Rec. 2018, 301, 1454–1466. [Google Scholar] [CrossRef] [Green Version]
- Kim, J.H.; Jeong, M.S.; Kim, D.Y.; Her, S.; Wie, M.B. Zinc oxide nanoparticles induce lipoxygenase-mediated apoptosis and necrosis in human neuroblastoma SH-SY5Y cells. Neurochem. Int. 2015, 90, 204–214. [Google Scholar] [CrossRef]
- Zhang, Y.; An, Y.; He, X.; Zhang, D.; He, W. Esculetin protects human corneal epithelial cells from oxidative stress through Nrf-2 signaling pathway. Exp. Eye Res. 2021, 202, 108360. [Google Scholar] [CrossRef]
- Tubaro, A.; Del Negro, P.; Rogazzi, E.; Zampiron, S.; Della Loggia, R. Anti-inflammatory and peripheral analgesic activity of esculetin in vivo. Pharmacol. Res. Commun. 1988, 20, 83–85. [Google Scholar] [CrossRef]
- Tien, Y.C.; Liao, J.C.; Chiu, C.S.; Huang, T.H.; Huang, C.Y.; Chang, W.T.; Peng, W.H. Esculetin ameliorates carbon tetrachloride-mediated hepatic apoptosis in rats. Int. J. Mol. Sci. 2011, 12, 4053–4067. [Google Scholar] [CrossRef] [Green Version]
- Subramaniam, S.R.; Ellis, E.M. Neuroprotective effects of umbelliferone and esculetin in a mouse model of Parkinson’s disease. J. Neurosci. Res. 2013, 91, 453–461. [Google Scholar] [CrossRef] [PubMed]
- Song, W.J.; Jeong, M.S.; Choi, D.M.; Kim, K.N.; Wie, M.B. Zinc oxide nanoparticles induce autophagy and apoptosis via oxidative injury and pro-inflammatory cytokines in primary astrocyte cultures. Nanomaterials 2019, 9, 1043. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, D.Y.; Kim, J.H.; Lee, J.C.; Won, M.H.; Yang, S.R.; Kim, H.C.; Wie, M.B. Zinc oxide nanoparticles exhibit both cyclooxygenase- and lipoxygenase-mediated apoptosis in human bone marrow-derived mesenchymal stem cells. Toxicol. Res. 2019, 35, 83–91. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, B.; Lu, F.; Wei, X.; Zhao, R. Fucoidan: Structure and bioactivity. Molecules 2008, 23, 1671–1695. [Google Scholar] [CrossRef] [Green Version]
- Xue, M.; Ge, Y.; Zhang, J.; Wang, Q.; Hou, L.; Liu, Y.; Sun, L.; Li, Q. Anticancer properties and mechanisms of fucoidan on mouse breast cancer in vitro and in vivo. PLoS ONE 2012, 7, e43483. [Google Scholar] [CrossRef]
- Lee, S.H.; Ko, C.I.; Ahn, G.; You, S.G.; Kim, J.S.; Heu, M.S.; Kim, J.; Jee, Y.; Jeon, Y.J. Molecular characteristics and anti-inflammatory activity of the fucoidan extracted from Ecklonia cava. Carbohydr. Polym. 2012, 89, 599–606. [Google Scholar] [CrossRef]
- Wang, Y.; Wang, Q.; Han, X.; Ma, Y.; Zhang, Z.; Zhao, L.; Guan, F.; Ma, S. Fucoidan: A promising agent for brain injury and neurodegenerative disease intervention. Food Funct. 2021, 12, 3820–3830. [Google Scholar] [CrossRef]
- Kim, K.; Choi, Y.; Ahn, M.; Jung, K.; Shin, T. Olfactory dysfunction in autoimmune central nervous system neuroinflammation. Mol. Neurobiol. 2018, 55, 8499–8508. [Google Scholar] [CrossRef]
- Amara, S.; Ben-slama, I.; Mrad, I.; Rihane, N.; Jeljeli, M.; El-Mir, L.; Ben-Rhouma, K.; Rachidi, W.; Sève, M.; Abdelmelek, H.; et al. Acute exposure to zinc oxide nanoparticles does not affect the cognitive capacity and neurotransmitter levels in adult rats. Nanotoxicology 2014, 8, 208–215. [Google Scholar] [CrossRef]
- Liang, H.; Chen, A.; Lai, X.; Liu, J.; Wu, J.; Kang, Y.; Wang, X.; Shao, L. Neuroinflammation is induced by tongue-instilled ZnO nanoparticles via the Ca2+-dependent NF-ĸB and MAPK pathways. Part. Fibre Toxicol. 2018, 15, 39. [Google Scholar] [CrossRef] [Green Version]
- Qin, X.; Tang, Q.; Jiang, X.; Zhang, J.; Wang, B.; Liu, X.; Zhang, Y.; Zou, Z.; Chen, C. Zinc oxide nanoparticles induce ferroptotic neuronal cell death in vitro and in vivo. Int. J. Nanomedicine 2020, 15, 5299–5315. [Google Scholar] [CrossRef] [PubMed]
- Ansar, S.; Abudawood, M.; Hamed, S.S.; Aleem, M.M. Exposure to zinc oxide nanoparticles induces neurotoxicity and proinflammatory response: Amelioration by hesperidin. Biol. Trace Elem. Res. 2017, 175, 360–366. [Google Scholar] [CrossRef] [PubMed]
- Kuhn, D.M.; Sakowski, S.A.; Sadidi, M.; Geddes, T.J. Nitrotyrosine as a marker for peroxynitrite-induced neurotoxicity: The beginning or the end of the end of dopamine neurons. J. Neurochem. 2004, 89, 529–536. [Google Scholar] [CrossRef] [PubMed]
- Shekaftik, S.O.; Nasirzadeh, N. 8-hydroxy-2’-deoxyguanosine (8-OHDA) as a biomarker of oxidative DNA damage induced by occupational exposure to nanomaterials: A systemic review. Nanotoxicology 2021, 15, 850–864. [Google Scholar] [CrossRef]
- Yasuhara, T.; Hara, K.; Sethi, K.D.; Morgan, J.C.; Borlongan, C.V. Increased 8-OHdG levels in the urine, serum, and substantia nigra of hemiparkinsonian rats. Brain Res. 2007, 1133, 49–52. [Google Scholar] [CrossRef]
- Chuang, H.C.; Juan, H.T.; Chang, C.N.; Yan, Y.H.; Yuan, T.H.; Wang, J.S.; Chen, H.C.; Hwang, Y.H.; Lee, C.H.; Cheng, T.J. Cardiopulmonary toxicity of pulmonary exposure to occupationally relevant zinc oxide nanoparticles. Nanotoxicology 2014, 8, 593–604. [Google Scholar] [CrossRef]
- Li, Y.S.; Ootsuyama, Y.; Kawasaki, Y.; Morimoto, Y.; Higashi, T.; Kawai, K. Oxidative DNA damage in the rat lung induced by intratracheal instillation and inhalation of nanoparticles. J. Clin. Biochem. Nutr. 2018, 62, 238–241. [Google Scholar] [CrossRef] [Green Version]
- Kumar, A.; Behl, T.; Jamwal, S.; Kaur, I.; Sood, A.; Kumar, P. Exploring the molecular approach of COX and LOX in Alzheimer’s and Parkinson’s disorder. Mol. Biol. Rep. 2020, 47, 9895–9912. [Google Scholar] [CrossRef]
- Saxena, M.; Dubey, R. Target enzyme in Alzheimer’s disease. Curr. Top. Med. Chem. 2019, 19, 264–275. [Google Scholar] [CrossRef]
- Giacconi, R.; Giuli, C.; Casoli, T.; Balietti, M.; Costarelli, L.; Provinciali, M.; Basso, A.; Piacenza, F.; Postacchini, D.; Galeazzi, R.; et al. Acetylcholinesterase inhibitors in Alzheimer’s disease influence zinc and copper homeostasis. J. Trace Elem. Med. Biol. 2019, 55, 58–63. [Google Scholar] [CrossRef]
- Zhang, Z.; Miah, M.; Culbreth, M.; Aschner, M. Autophagy in neurodegenerative diseases and metal neurotoxicity. Neurochem. Res. 2016, 41, 409–422. [Google Scholar] [CrossRef] [PubMed]
- Lu, J.; Wu, M.; Yue, Z. Autophagy and Parkinson’s disease. Adv. Exp. Med. Biol. 2020, 1207, 21–51. [Google Scholar] [PubMed]
- Gao, Y.; Li, C.; Yin, J.; Shen, J.; Wang, H.; Wu, Y.; Jin, H. Fucoidan, a sulfated polysaccharide from brown algae, improves cognitive impairment induced by infusion of Aβ peptide in rats. Environ. Toxicol. Pharmacol. 2012, 33, 304–311. [Google Scholar] [CrossRef] [PubMed]
- Huiliang, Z.; Mengzhe, Y.; Xiaochuan, W.; Hui, W.; Min, D.; Mengqi, W.; Jianzhi, W.; Zhongshan, C.; Caixia, P.; Rong, L. Zinc induces reactive astrogliosis through ERK-dependent activation of Stat3 and promotes synaptic degenration. J. Neurochem. 2021, 159, 1016–1027. [Google Scholar] [CrossRef]
- Freeman, M.R. Specification and morphogenesis of astrocytes. Science 2010, 330, 774–778. [Google Scholar] [CrossRef] [Green Version]
- Segawa, S.; Tatsumi, N.; Ohishi, A.; Nishida, K.; Nagasawa, K. Characterization of zinc uptake by mouse primary cultured astrocyte and microglia. Metallomics 2015, 7, 1067–1077. [Google Scholar] [CrossRef]
- Benveniste, E.N. Cytokine actions in the central nervous system. Cytokine Growth Factor Rev. 1998, 9, 259–275. [Google Scholar] [CrossRef]
- Gao, Z.; Zhu, Q.; Zhang, Y.; Zhao, Y.; Cai, L.; Shields, C.B.; Cai, J. Reciprocal modulation between microglia and astrocyte in reactive gliosis following the CNS injury. Mol. Neurobiol. 2013, 48, 690–701. [Google Scholar] [CrossRef] [Green Version]













Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Song, W.-J.; Kim, J.; Shin, T.; Jeong, M.-S.; Kim, K.-N.; Yun, J.-H.; Wie, M.-B. Esculetin and Fucoidan Attenuate Autophagy and Apoptosis Induced by Zinc Oxide Nanoparticles through Modulating Reactive Astrocyte and Proinflammatory Cytokines in the Rat Brain. Toxics 2022, 10, 194. https://0-doi-org.brum.beds.ac.uk/10.3390/toxics10040194
Song W-J, Kim J, Shin T, Jeong M-S, Kim K-N, Yun J-H, Wie M-B. Esculetin and Fucoidan Attenuate Autophagy and Apoptosis Induced by Zinc Oxide Nanoparticles through Modulating Reactive Astrocyte and Proinflammatory Cytokines in the Rat Brain. Toxics. 2022; 10(4):194. https://0-doi-org.brum.beds.ac.uk/10.3390/toxics10040194
Chicago/Turabian StyleSong, Woo-Ju, Jeongtae Kim, Taekyun Shin, Myeong-Seon Jeong, Kil-Nam Kim, Jang-Hyuk Yun, and Myung-Bok Wie. 2022. "Esculetin and Fucoidan Attenuate Autophagy and Apoptosis Induced by Zinc Oxide Nanoparticles through Modulating Reactive Astrocyte and Proinflammatory Cytokines in the Rat Brain" Toxics 10, no. 4: 194. https://0-doi-org.brum.beds.ac.uk/10.3390/toxics10040194