A Phenotypic and Genotypic Evaluation of Developmental Toxicity of Polyhexamethylene Guanidine Phosphate Using Zebrafish Embryo/Larvae

 , and
, and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Test Chemicals and Solution
2.2. Zebrafish Embryos
2.3. Embryonic Toxicity
2.4. Embryo/Larva Imaging
2.5. Heart Rate Count
2.6. RNA Sequencing
2.7. GO Analysis
2.8. Gene Expression Measurement
2.9. Statistical Analysis
3. Results
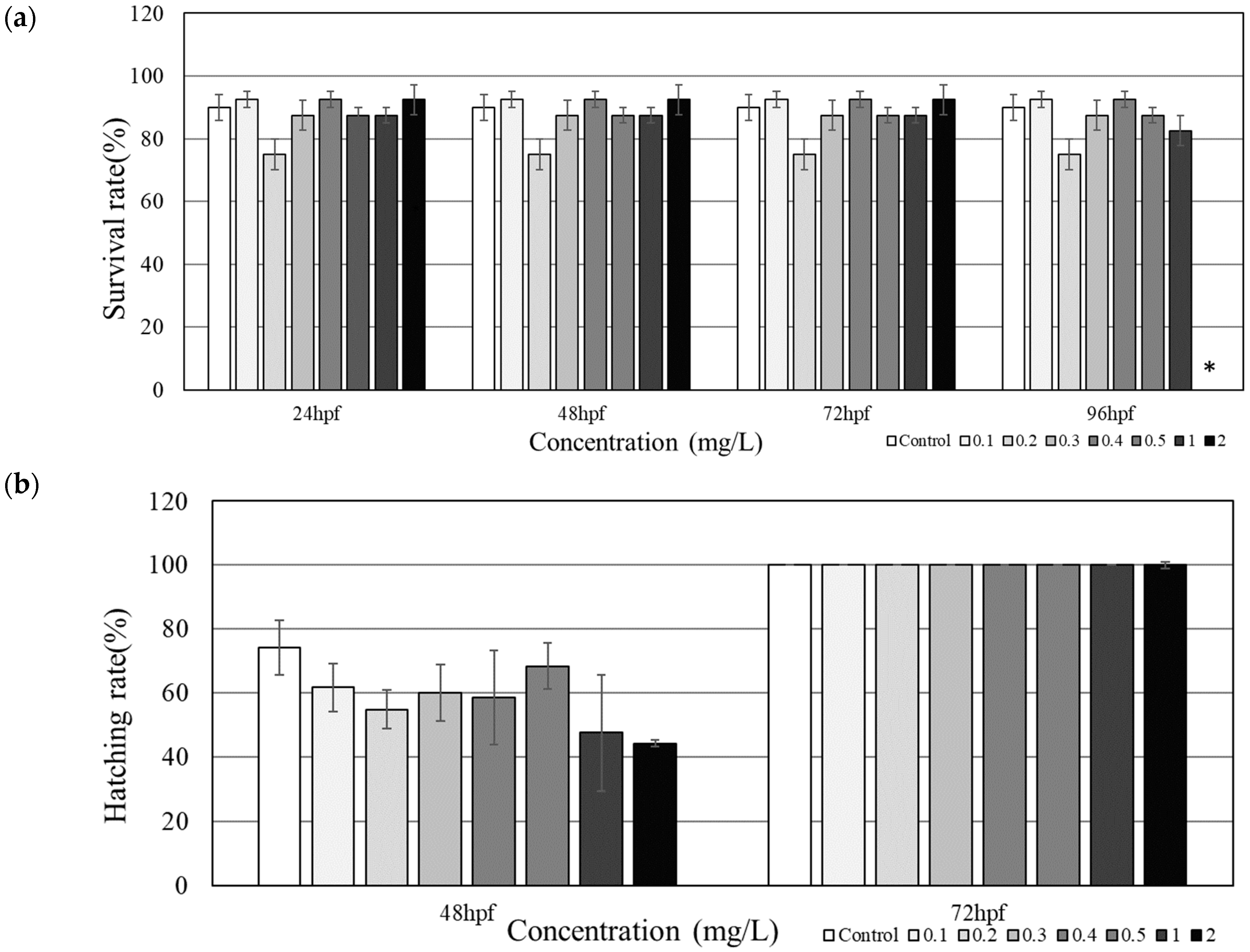
3.1. Mortality and Developmental Toxicity
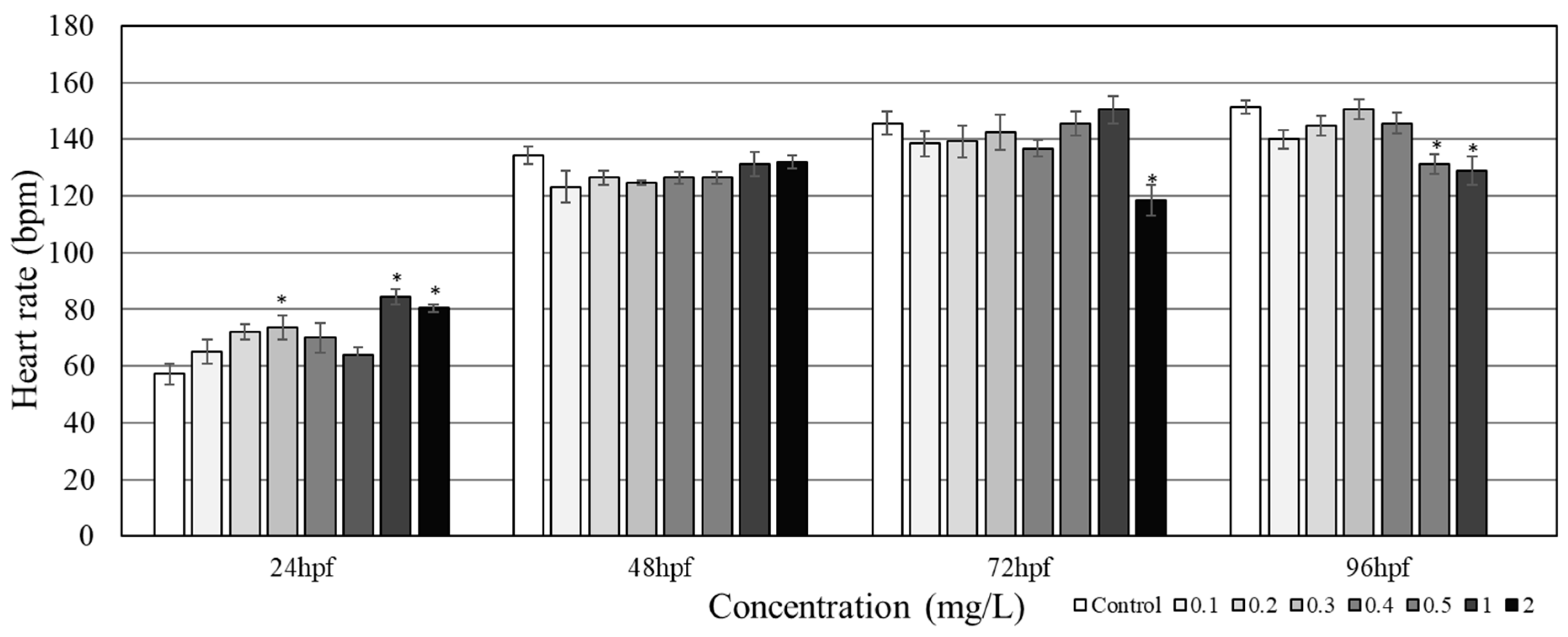
3.2. Cardiotoxicity
3.3. RNA Sequencing Analysis
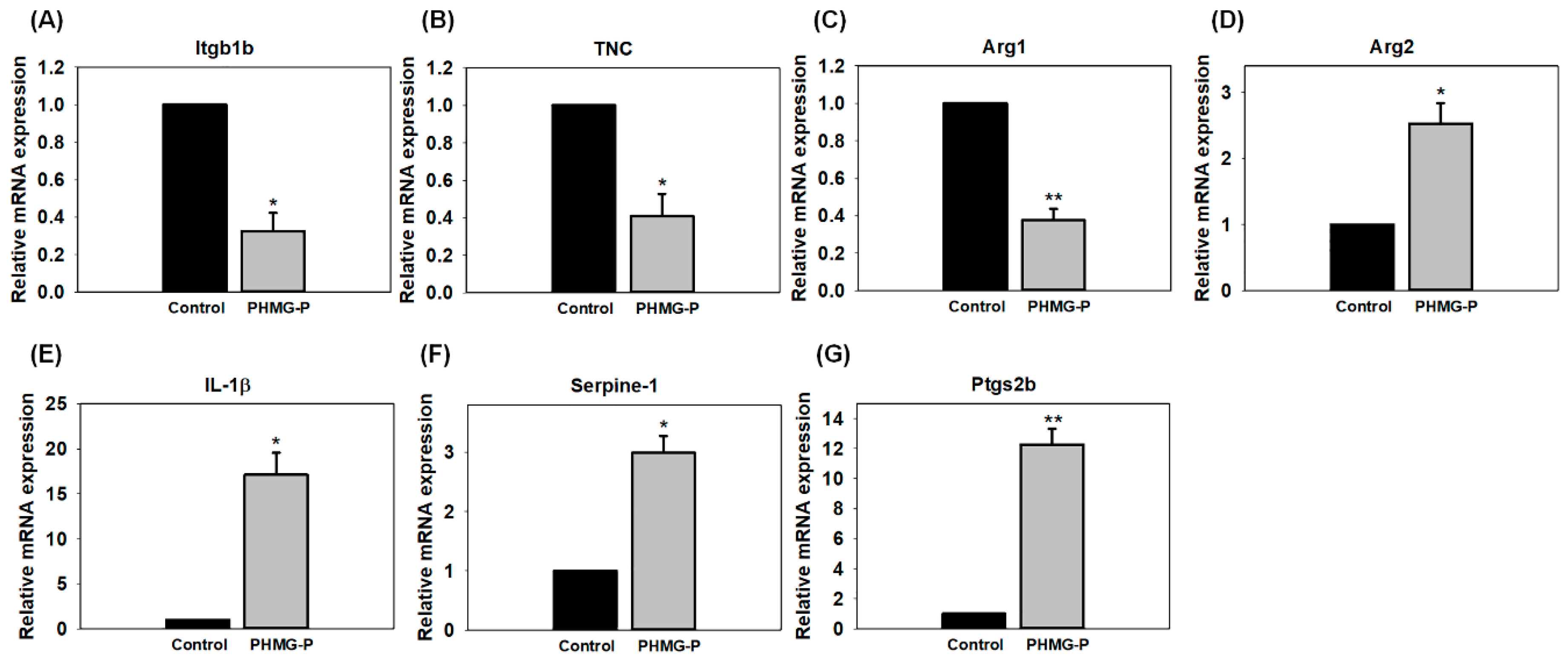
3.4. Quantitative PCR Analysis
4. Discussion
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Zhang, Y.; Jiang, J.; Chen, Y. Synthesis and antimicrobial activity of polymeric guanidine and biguanidine salts. Polymer 1999, 40, 6189–6198. [Google Scholar] [CrossRef]
- Ohta, S.; Misawa, Y.; Miyamoto, H.; Makino, M.; Nagai, K.; Shiraishi, T.; Nakagawa, Y.; Yamato, S.; Tachikawa, E.; Zenda, H. A Comparative Study of Characteristics of Current-Type and Conventional-Type Cationic Bactericides. Biol. Pharm. Bull. 2001, 24, 1093–1096. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vitt, A.; Sofrata, A.; Slizen, V.; Sugars, R.V.; Gustafsson, A.; Gudkova, E.I.; Kazeko, L.A.; Ramberg, P.; Buhlin, K. Antimicrobial activity of polyhexamethylene guanidine phosphate in comparison to chlorhexidine using the quantitative suspension method. Ann. Clin. Microbiol. Antimicrob. 2015, 14, 36. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Park, E.J.; Park, S.J.; Kim, S.; Lee, K.; Chang, J. Lung fibroblasts may play an important role in clearing apoptotic bodies of bronchial epithelial cells generated by exposure to PHMG-P-containing solution. Toxicol. Lett. 2018, 286, 108–119. [Google Scholar] [CrossRef]
- Lee, J.H.; Kim, Y.H.; Kwon, J.H. Fatal misuse of humidifier disinfectants in Korea: importance of screening risk assessment and implications for management of chemicals in consumer products. Environ. Sci. Technol. 2012, 46, 2498–2500. [Google Scholar] [CrossRef]
- NICNAS. National Industrial Chemicals Notification and Assessment Scheme (NICNAS)—Full Public Report Polyhexamethyleneguanidine Phosphate; National Occupational Health and Safety Commission: Sydney, Australia, 2001. [Google Scholar]
- Ostapenko, Y.N.; Brusin, K.M.; Zobnin, Y.V.; Shchupak, A.Y.; Vishnevetskiy, M.K.; Sentsov, V.G.; Novikova, O.V.; Alekseenko, S.A.; Lebed’ko, O.A.; Puchkov, Y.B. Acute cholestatic liver injury caused by polyhexamethyleneguanidine hydrochloride admixed to ethyl alcohol. Clin. Toxicol. (Phila) 2011, 49, 471–477. [Google Scholar] [CrossRef]
- Kim, J.Y.; Kim, H.H.; Cho, K.H. Acute cardiovascular toxicity of sterilizers, PHMG, and PGH: Severe inflammation in human cells and heart failure in zebrafish. Cardiovasc. Toxicol. 2013, 13, 148–160. [Google Scholar] [CrossRef]
- Song, J.A.; Park, H.J.; Yang, M.J.; Jung, K.J.; Yang, H.S.; Song, C.W.; Lee, K. Polyhexamethyleneguanidine phosphate induces severe lung inflammation, fibrosis, and thymic atrophy. Food Chem. Toxicol. 2014, 69, 267–275. [Google Scholar] [CrossRef]
- Kim, H.R.; Lee, K.; Park, C.W.; Song, J.A.; Shin, D.Y.; Park, Y.J.; Chung, K.H. Polyhexamethylene guanidine phosphate aerosol particles induce pulmonary inflammatory and fibrotic responses. Arch. Toxicol. 2016, 90, 617–632. [Google Scholar] [CrossRef]
- Kim, M.S.; Jeong, S.W.; Choi, S.J.; Han, J.Y.; Kim, S.H.; Yoon, S.; Oh, J.H.; Lee, K. Analysis of genomic responses in a rat lung model treated with a humidifier sterilizer containing polyhexamethyleneguanidine phosphate. Toxicol. Lett. 2017, 268, 36–43. [Google Scholar] [CrossRef]
- Kim, H.; Ji, K. Exposure to humidifier disinfectants induces developmental effects and disrupts thyroid endocrine systems in zebrafish larvae. Ecotoxicol. Environ. Saf. 2019, 184, 109663. [Google Scholar] [CrossRef] [PubMed]
- Veldman, M.B.; Lin, S. Zebrafish as a Developmental Model Organism for Pediatric Research. Pediatr. Res. 2008, 64, 470–476. [Google Scholar] [CrossRef] [Green Version]
- Foster, W.R.; Chen, S.J.; He, A.; Truong, A.; Bhaskaran, V.; Nelson, D.M.; Dambach, D.M.; Lehman-McKeeman, L.D.; Car, B.D. A retrospective analysis of toxicogenomics in the safety assessment of drug candidates. Toxicol. Pathol. 2007, 35, 621–635. [Google Scholar] [CrossRef] [PubMed]
- Jung, H.N.; Zerin, T.; Podder, B.; Song, H.Y.; Kim, Y.S. Cytotoxicity and gene expression profiling of polyhexamethylene guanidine hydrochloride in human alveolar A549 cells. Toxicol. In Vitro 2014, 28, 684–692. [Google Scholar] [CrossRef] [PubMed]
- Nüsslein-Volhard, C.; Dahm, R. Zebrafish: A Practical Approach; Oxford University Press: New York, NY, USA, 2002. [Google Scholar]
- Oh, H.; Kim, C.Y.; Ryu, B.; Kim, U.; Kim, J.; Lee, J.M.; Lee, B.H.; Moon, J.; Jung, C.R.; Park, J.H. Respiratory Toxicity of Polyhexamethylene Guanidine Phosphate Exposure in Zebrafish. Zebrafish 2018, 15, 460–472. [Google Scholar] [CrossRef] [PubMed]
- Langheinrich, U.; Vacun, G.; Wagner, T. Zebrafish embryos express an orthologue of HERG and are sensitive toward a range of QT-prolonging drugs inducing severe arrhythmia. Toxicol. Appl. Pharmacol. 2003, 193, 370–382. [Google Scholar] [CrossRef]
- Anders, S.; Pyl, P.; Huber, W. HTSeq—a Python framework to work with high-throughput sequencing data. Bioinformatics 2015, 31, 166–169. [Google Scholar] [CrossRef]
- Sun, J.; Nishiyama, T.; Shimizu, K.; Kadota, K. TCC: an R package for comparing tag count data with robust normalization strategies. BMC Bioinform. 2013, 14, 219. [Google Scholar] [CrossRef] [Green Version]
- van Aerle, R.; Lange, A.; Moorhouse, A.; Paszkiewicz, K.; Ball, K.; Johnston, B.D.; de-Bastos, E.; Booth, T.; Tyler, C.R.; Santos, E.M. Molecular mechanisms of toxicity of silver nanoparticles in zebrafish embryos. Environ. Sci. Technol. 2013, 47, 8005–8014. [Google Scholar] [CrossRef] [Green Version]
- Rao, X.; Huang, X.; Zhou, Z.; Lin, X. An improvement of the 2^(-delta delta CT) method for quantitative real-time polymerase chain reaction data analysis. Biostat. Bioinform. Biomath. 2013, 3, 71–85. [Google Scholar]
- De la Paz, J.F.; Beiza, N.; Paredes-Zuniga, S.; Hoare, M.S.; Allende, M.L. Triazole Fungicides Inhibit Zebrafish Hatching by Blocking the Secretory Function of Hatching Gland Cells. Int. J. Mol. Sci. 2017, 18. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Teixido, E.; Barenys, M.; Pique, E.; Llobet, J.M.; Gomez-Catalan, J. Cardiovascular Effects of PCB 126 (3,3’,4,4’,5-Pentachlorobiphenyl) in Zebrafish Embryos and Impact of Co-Exposure to Redox Modulating Chemicals. Int. J. Mol. Sci. 2019, 20. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fischer, S.; Kluver, N.; Burkhardt-Medicke, K.; Pietsch, M.; Schmidt, A.M.; Wellner, P.; Schirmer, K.; Luckenbach, T. Abcb4 acts as multixenobiotic transporter and active barrier against chemical uptake in zebrafish (Danio rerio) embryos. BMC Biol. 2013, 11, 69. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hallare, A.; Nagel, K.; Kohler, H.R.; Triebskorn, R. Comparative embryotoxicity and proteotoxicity of three carrier solvents to zebrafish (Danio rerio) embryos. Ecotoxicol. Environ. Saf. 2006, 63, 378–388. [Google Scholar] [CrossRef] [PubMed]
- Hennig, B.; Meerarani, P.; Slim, R.; Toborek, M.; Daugherty, A.; Silverstone, A.E.; Robertson, L.W. Proinflammatory properties of coplanar PCBs: In vitro and in vivo evidence. Toxicol. Appl. Pharmacol. 2002, 181, 174–183. [Google Scholar] [CrossRef]
- Kim, H.R.; Shin, D.Y.; Chung, K.H. The role of NF-kappaB signaling pathway in polyhexamethylene guanidine phosphate induced inflammatory response in mouse macrophage RAW264.7 cells. Toxicol. Lett. 2015, 233, 148–155. [Google Scholar] [CrossRef]
- Scholz, S. Zebrafish embryos as an alternative model for screening of drug-induced organ toxicity. Arch. Toxicol. 2013, 87, 767–769. [Google Scholar] [CrossRef] [Green Version]
- Wang, X.; Li, L.; Liu, D. Expression analysis of integrin beta1 isoforms during zebrafish embryonic development. Gene Expert Patterns 2014, 16, 86–92. [Google Scholar] [CrossRef]
- Yu, Y.M.; Cristofanilli, M.; Valiveti, A.; Ma, L.; Yoo, M.; Morellini, F.; Schachner, M. The extracellular matrix glycoprotein tenascin-C promotes locomotor recovery after spinal cord injury in adult zebrafish. Neuroscience 2011, 183, 238–250. [Google Scholar] [CrossRef]
- Golias, C.; Batistatou, A.; Bablekos, G.; Charalabopoulos, A.; Peschos, D.; Mitsopoulos, P.; Charalabopoulos, K. Physiology and pathophysiology of selectins, integrins, and IgSF cell adhesion molecules focusing on inflammation. A paradigm model on infectious endocarditis. Cell Commun. Adhes. 2011, 18, 19–32. [Google Scholar] [CrossRef] [Green Version]
- Gonzalez-Amaro, R.; Diaz-Gonzalez, F.; Sanchez-Madrid, F. Adhesion molecules in inflammatory diseases. Drugs 1998, 56, 977–988. [Google Scholar] [CrossRef] [PubMed]
- Mills, C.D.; Kincaid, K.; Alt, J.M.; Heilman, M.J.; Hill, A.M. M-1/M-2 Macrophages and the Th1/Th2 Paradigm. J. Immunol. 2000, 164, 6166–6173. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, G.; Sidney, M. Morris, J. Arginine metabolism: nitric oxide and beyond. Biochem. J. 1998, 336, 1–17. [Google Scholar] [CrossRef] [PubMed]
- Ming, X.F.; Rajapakse, A.G.; Yepuri, G.; Xiong, Y.; Carvas, J.M.; Ruffieux, J.; Scerri, I.; Wu, Z.; Popp, K.; Li, J.; et al. Arginase II Promotes Macrophage Inflammatory Responses Through Mitochondrial Reactive Oxygen Species, Contributing to Insulin Resistance and Atherogenesis. J. Am. Heart Assoc. 2012, 1, e000992. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Klasen, S.; Hammermann, R.; Fuhrmann, M.; Lindemann, D.; Beck, K.-F.; Pfeilschifter, J.; Racke, K. Glucocorticoids inhibit lipopolysaccharide-induced up-regulation of arginase in rat alveolar macrophages. Br. J. Pharmacol. 2001, 132, 1349–1357. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Que, L.G.; Kantrow, S.P.; Jenkinson, C.P.; Piantadosi, C.A.; Hua, Y.-C.T. Induction of arginase isoforms in the lung during hyperoxia. Am. J. Physiol. 1998, 275, 96–102. [Google Scholar] [CrossRef] [PubMed]
- Maarsingh, H.; Pera, T.; Meurs, H. Arginase and pulmonary diseases. Naunyn Schmiedeberg’s Arch. Pharmacol. 2008, 378, 171–184. [Google Scholar] [CrossRef] [Green Version]
- Yan, B.; Han, P.; Pan, L.; Lu, W.; Xiong, J.; Zhang, M.; Zhang, W.; Li, L.; Wen, Z. IL-1beta and reactive oxygen species differentially regulate neutrophil directional migration and Basal random motility in a zebrafish injury-induced inflammation model. J. Immunol. 2014, 192, 5998–6008. [Google Scholar] [CrossRef] [Green Version]
- Lopez-Castejon, G.; Brough, D. Understanding the mechanism of IL-1beta secretion. Cytokine Growth Factor Rev. 2011, 22, 189–195. [Google Scholar] [CrossRef]
- Ferrero-Miliani, L.; Nielsen, O.H.; Andersen, P.S.; Girardin, S.E. Chronic inflammation: importance of NOD2 and NALP3 in interleukin-1beta generation. Clin. Exp. Immunol. 2007, 147, 227–235. [Google Scholar] [CrossRef]
- Bonniaud, P.; Margetts, P.J.; Ask, K.; Flanders, K.; Gauldie, J.; Kolb, M. TGF- and Smad3 Signaling Link Inflammation to Chronic Fibrogenesis. J. Immunol. 2005, 175, 5390–5395. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kolb, M.; Margetts, P.J.; Anthony, D.C.; Pitossi, F.; Gauldie, J. Transient expression of IL-1beta induces acute lung injury and chronic repair leading to pulmonary fibrosis. J. Clin. Invest. 2001, 107, 1529–1536. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kaiko, G.E.; Chen, F.; Lai, C.W.; Chiang, I.L.; Perrigoue, J.; Stojmirovic, A.; Li, K.; Muegge, B.D.; Jain, U.; VanDussen, K.L.; et al. PAI-1 augments mucosal damage in colitis. Sci. Transl. Med. 2019, 11. [Google Scholar] [CrossRef] [PubMed]
- Simone, T.M.; Higgins, C.E.; Czekay, R.P.; Law, B.K.; Higgins, S.P.; Archambeault, J.; Kutz, S.M.; Higgins, P.J. SERPINE1: A Molecular Switch in the Proliferation-Migration Dichotomy in Wound-”Activated” Keratinocytes. Adv. Wound Care (New Rochelle) 2014, 3, 281–290. [Google Scholar] [CrossRef] [Green Version]
- Galindo-Villegas, J.; Garcia-Moreno, D.; de Oliveira, S.; Meseguer, J.; Mulero, V. Regulation of immunity and disease resistance by commensal microbes and chromatin modifications during zebrafish development. Proc. Natl. Acad. Sci. USA 2012, 109, E2605–E2614. [Google Scholar] [CrossRef] [Green Version]
- Park, D.U.; Ryu, S.H.; Lim, H.K.; Kim, S.K.; Choi, Y.Y.; Ahn, J.J.; Lee, E.; Hong, S.B.; Do, K.H.; Cho, J.L.; et al. Types of household humidifier disinfectant and associated risk of lung injury (HDLI) in South Korea. Sci. Total Environ. 2017, 596–597, 53–60. [Google Scholar] [CrossRef]
- Kim, S.; Park, S.; Jo, H.; Song, S.; Ham, S.; Yoon, C. Behavioral characteristics of polyhexamethyleneguanidine (PHMG) particles in aqueous solution and air when sprayed into an ultrasonic humidifier. Environ. Res. 2020, 182, 109078. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Ontology | Term | GO ID | p-Value (at 96 h) | ||
|---|---|---|---|---|---|
| I | II | III | |||
| BP | response to molecule of bacterial origin | GO:0002237 | 0.000006 | 0.000002 | 0.000388 |
| BP | immune system process | GO:0002376 | <0.000001 | <0.000001 | <0.000001 |
| BP | antigen processing and presentation of peptide antigen via MHC class I | GO:0002474 | 0.000917 | 0.000005 | 0.000082 |
| BP | UDP-N-acetylglucosamine metabolic process | GO:0006047 | 0.000256 | 0.000352 | 0.000321 |
| BP | protein glycosylation | GO:0006486 | 0.000252 | 0.000006 | 0.000049 |
| BP | proteolysis | GO:0006508 | 0.000184 | 0.000172 | 0.000098 |
| BP | fatty acid biosynthetic process | GO:0006633 | <0.000001 | <0.000001 | 0.000011 |
| BP | unsaturated fatty acid biosynthetic process | GO:0006636 | 0.000002 | 0.000003 | 0.000035 |
| BP | response to stress | GO:0006950 | <0.000001 | <0.000001 | <0.000001 |
| BP | defense response | GO:0006952 | <0.000001 | <0.000001 | <0.000001 |
| BP | inflammatory response | GO:0006954 | <0.000001 | <0.000001 | <0.000001 |
| BP | immune response | GO:0006955 | <0.000001 | <0.000001 | <0.000001 |
| BP | nucleotide-sugar biosynthetic process | GO:0009226 | 0.000001 | 0.000001 | 0.000001 |
| BP | GDP-mannose biosynthetic process | GO:0009298 | 0.000350 | 0.000448 | 0.000418 |
| BP | response to biotic stimulus | GO:0009607 | <0.000001 | <0.000001 | <0.000001 |
| BP | response to wounding | GO:0009611 | <0.000001 | <0.000001 | <0.000001 |
| BP | response to virus | GO:0009615 | <0.000001 | <0.000001 | <0.000001 |
| BP | response to bacterium | GO:0009617 | <0.000001 | <0.000001 | <0.000001 |
| BP | cytokine-mediated signaling pathway | GO:0019221 | 0.000032 | 0.000068 | 0.000238 |
| BP | GDP-mannose metabolic process | GO:0019673 | 0.000018 | 0.000025 | 0.000023 |
| BP | neutrophil chemotaxis | GO:0030593 | 0.000323 | 0.000471 | 0.000049 |
| BP | leukocyte chemotaxis | GO:0030595 | 0.000010 | 0.000018 | 0.000002 |
| BP | response to lipopolysaccharide | GO:0032496 | 0.000022 | 0.000006 | 0.000216 |
| BP | response to cytokine | GO:0034097 | 0.000006 | 0.000015 | 0.000011 |
| BP | positive regulation of MAPK cascade | GO:0043410 | 0.000206 | 0.000363 | 0.000002 |
| BP | innate immune response | GO:0045087 | 0.000051 | 0.000008 | <0.000001 |
| BP | leukocyte migration | GO:0050900 | <0.000001 | 0.000002 | <0.000001 |
| BP | cell chemotaxis | GO:0060326 | 0.000016 | 0.000029 | 0.000003 |
| CC | cell-cell junction | GO:0005911 | 0.000003 | 0.000011 | 0.000345 |
| CC | tight junction | GO:0005923 | <0.000001 | <0.000001 | 0.000026 |
| CC | NADPH oxidase complex | GO:0043020 | 0.000133 | 0.000172 | 0.00016 |
| CC | apical junction complex | GO:0043296 | 0.000001 | <0.000001 | 0.000042 |
| MF | alpha-1,6-mannosyltransferase activity | GO:0000009 | 0.000383 | 0.000125 | 0.000105 |
| MF | alpha-1,2-mannosyltransferase activity | GO:0000026 | 0.000438 | 0.000146 | 0.000123 |
| MF | alpha-1,3-mannosyltransferase activity | GO:0000033 | 0.000383 | 0.000125 | 0.000105 |
| MF | alpha-1,3-galactosyltransferase activity | GO:0001962 | 0.000383 | 0.000125 | 0.000105 |
| MF | NAD+ ADP-ribosyltransferase activity | GO:0003950 | <0.000001 | 0.000354 | 0.000034 |
| MF | UDP-glucose:glycoprotein glucosyltransferase activity | GO:0003980 | 0.000501 | 0.000170 | 0.000143 |
| MF | endopeptidase activity | GO:0004175 | <0.000001 | <0.000001 | <0.000001 |
| MF | cysteine-type endopeptidase activity | GO:0004197 | <0.000001 | <0.000001 | <0.000001 |
| MF | glycolipid mannosyltransferase activity | GO:0004376 | 0.000498 | 0.000169 | 0.000142 |
| MF | oligosaccharyl transferase activity | GO:0004576 | 0.000693 | 0.000247 | 0.000208 |
| MF | dolichyl-phosphate-glucose-glycolipid alpha-glucosyltransferase activity | GO:0004583 | 0.000437 | 0.000146 | 0.000122 |
| MF | phospholipase inhibitor activity | GO:0004859 | 0.000001 | 0.000002 | 0.000001 |
| MF | peptidase activity | GO:0008233 | 0.000010 | 0.000030 | 0.000003 |
| MF | cysteine-type peptidase activity | GO:0008234 | 0.000038 | 0.000103 | <0.000001 |
| MF | acetylglucosaminyltransferase activity | GO:0008375 | 0.000624 | 0.000070 | 0.000056 |
| MF | galactosyltransferase activity | GO:0008378 | 0.000194 | 0.000002 | 0.00001 |
| MF | fucosyltransferase activity | GO:0008417 | 0.000003 | 0.000001 | 0.000006 |
| MF | O antigen polymerase activity | GO:0008755 | 0.000383 | 0.000125 | 0.000105 |
| MF | lipopolysaccharide-1,6-galactosyltransferase activity | GO:0008921 | 0.000383 | 0.000125 | 0.000105 |
| MF | transferase activity, transferring glycosyl groups | GO:0016757 | 0.000001 | <0.000001 | 0.000001 |
| MF | transferase activity, transferring hexosyl groups | GO:0016758 | 0.000320 | 0.000001 | 0.000027 |
| MF | cellulose synthase activity | GO:0016759 | 0.000383 | 0.000125 | 0.000105 |
| MF | 9-phenanthrol UDP-glucuronosyltransferase activity | GO:0018715 | 0.000383 | 0.000125 | 0.000105 |
| MF | 1-phenanthrol glycosyltransferase activity | GO:0018716 | 0.000383 | 0.000125 | 0.000105 |
| MF | 9-phenanthrol glycosyltransferase activity | GO:0018717 | 0.000383 | 0.000125 | 0.000105 |
| MF | 1,2-dihydroxy-phenanthrene glycosyltransferase activity | GO:0018718 | 0.000383 | 0.000125 | 0.000105 |
| MF | phenanthrol glycosyltransferase activity | GO:0019112 | 0.000383 | 0.000125 | 0.000105 |
| MF | beta-1,4-mannosyltransferase activity | GO:0019187 | 0.000383 | 0.000125 | 0.000105 |
| MF | alpha-1,2-galactosyltransferase activity | GO:0031278 | 0.000383 | 0.000125 | 0.000105 |
| MF | dolichyl pyrophosphate Man7GlcNAc2 alpha-1,3-glucosyltransferase activity | GO:0033556 | 0.000383 | 0.000125 | 0.000105 |
| MF | UDP-glucosyltransferase activity | GO:0035251 | 0.000810 | 0.000294 | 0.000248 |
| MF | lipopolysaccharide-1,5-galactosyltransferase activity | GO:0035496 | 0.000383 | 0.000125 | 0.000105 |
| MF | dolichyl pyrophosphate Man9GlcNAc2 alpha-1,3-glucosyltransferase activity | GO:0042281 | 0.000497 | 0.000169 | 0.000142 |
| MF | dolichyl pyrophosphate Glc1Man9GlcNAc2 alpha-1,3-glucosyltransferase activity | GO:0042283 | 0.000383 | 0.000125 | 0.000105 |
| MF | inositol phosphoceramide synthase activity | GO:0045140 | 0.000383 | 0.000125 | 0.000105 |
| MF | alpha-(1->3)-fucosyltransferase activity | GO:0046920 | 0.000059 | 0.000022 | 0.000099 |
| MF | alpha-(1->6)-fucosyltransferase activity | GO:0046921 | 0.000438 | 0.000146 | 0.000123 |
| MF | indole-3-butyrate beta-glucosyltransferase activity | GO:0052638 | 0.000383 | 0.000125 | 0.000105 |
| MF | salicylic acid glucosyltransferase (ester-forming) activity | GO:0052639 | 0.000383 | 0.000125 | 0.000105 |
| MF | salicylic acid glucosyltransferase (glucoside-forming) activity | GO:0052640 | 0.000383 | 0.000125 | 0.000105 |
| MF | benzoic acid glucosyltransferase activity | GO:0052641 | 0.000383 | 0.000125 | 0.000105 |
| MF | chondroitin hydrolase activity | GO:0052757 | 0.000383 | 0.000125 | 0.000105 |
| MF | dolichyl-pyrophosphate Man7GlcNAc2 alpha-1,6-mannosyltransferase activity | GO:0052824 | 0.000383 | 0.000125 | 0.000105 |
| MF | cytokinin 9-beta-glucosyltransferase activity | GO:0080062 | 0.000383 | 0.000125 | 0.000105 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Song, J.; Eghan, K.; Lee, S.; Park, J.-S.; Yoon, S.; Pimtong, W.; Kim, W.-K. A Phenotypic and Genotypic Evaluation of Developmental Toxicity of Polyhexamethylene Guanidine Phosphate Using Zebrafish Embryo/Larvae. Toxics 2020, 8, 33. https://0-doi-org.brum.beds.ac.uk/10.3390/toxics8020033
Song J, Eghan K, Lee S, Park J-S, Yoon S, Pimtong W, Kim W-K. A Phenotypic and Genotypic Evaluation of Developmental Toxicity of Polyhexamethylene Guanidine Phosphate Using Zebrafish Embryo/Larvae. Toxics. 2020; 8(2):33. https://0-doi-org.brum.beds.ac.uk/10.3390/toxics8020033
Chicago/Turabian StyleSong, Jeongah, Kojo Eghan, Sangwoo Lee, Jong-Su Park, Seokjoo Yoon, Wittaya Pimtong, and Woo-Keun Kim. 2020. "A Phenotypic and Genotypic Evaluation of Developmental Toxicity of Polyhexamethylene Guanidine Phosphate Using Zebrafish Embryo/Larvae" Toxics 8, no. 2: 33. https://0-doi-org.brum.beds.ac.uk/10.3390/toxics8020033