Water-Pipe Smoking Exposure Deregulates a Set of Genes Associated with Human Head and Neck Cancer Development and Prognosis

 ,
,  ,
,  ,
,  ,
,  and
and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Smoking Machine Protocol and WPS Preparation
2.2. Cell Lines
2.3. RNA Extraction and Reverse Transcriptase Real-Time PCR
2.4. NanoString
2.5. Gene Profile and In Silica Analyses
2.6. Network and Pathway Interaction
2.7. Statistical Analysis
3. Results
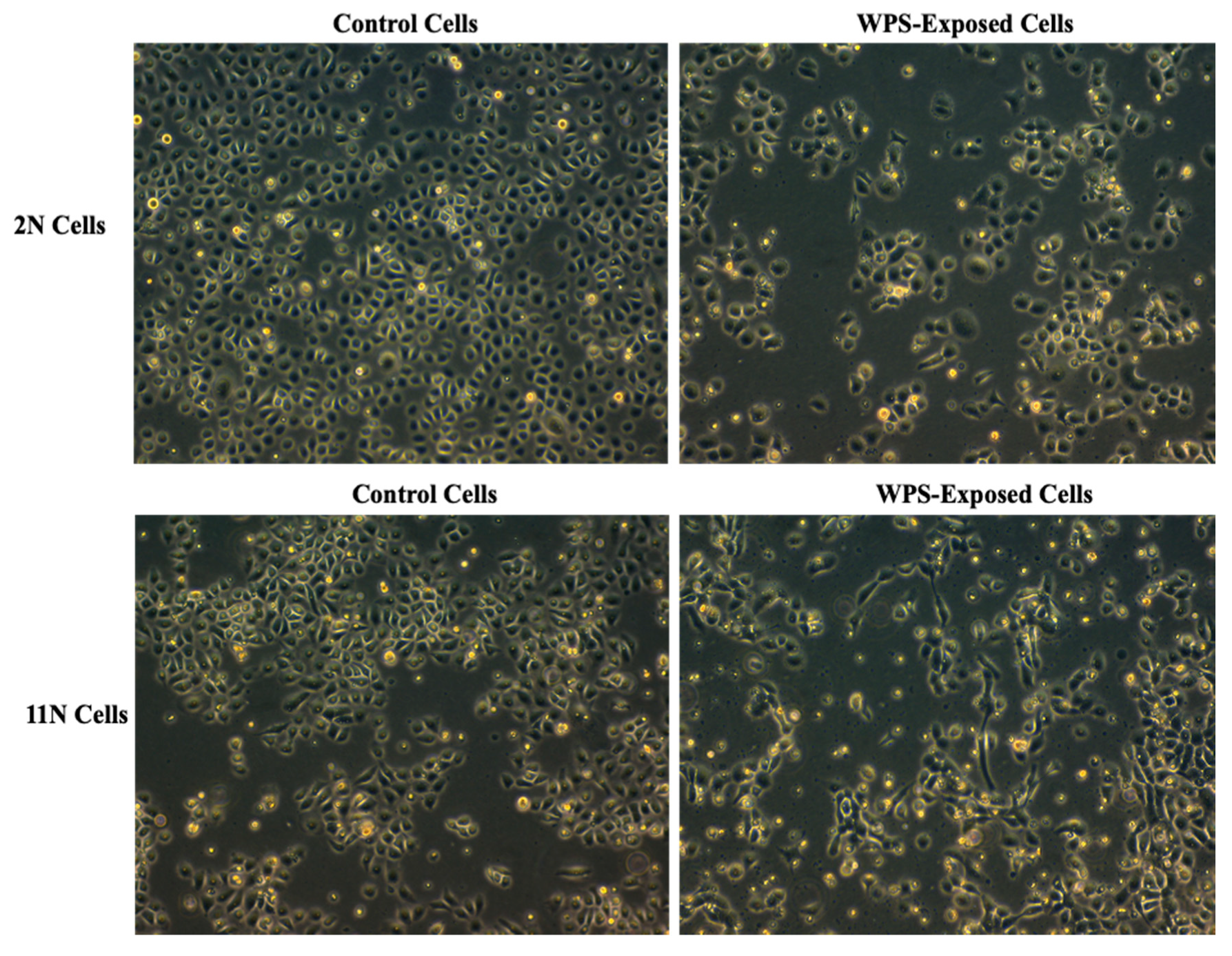
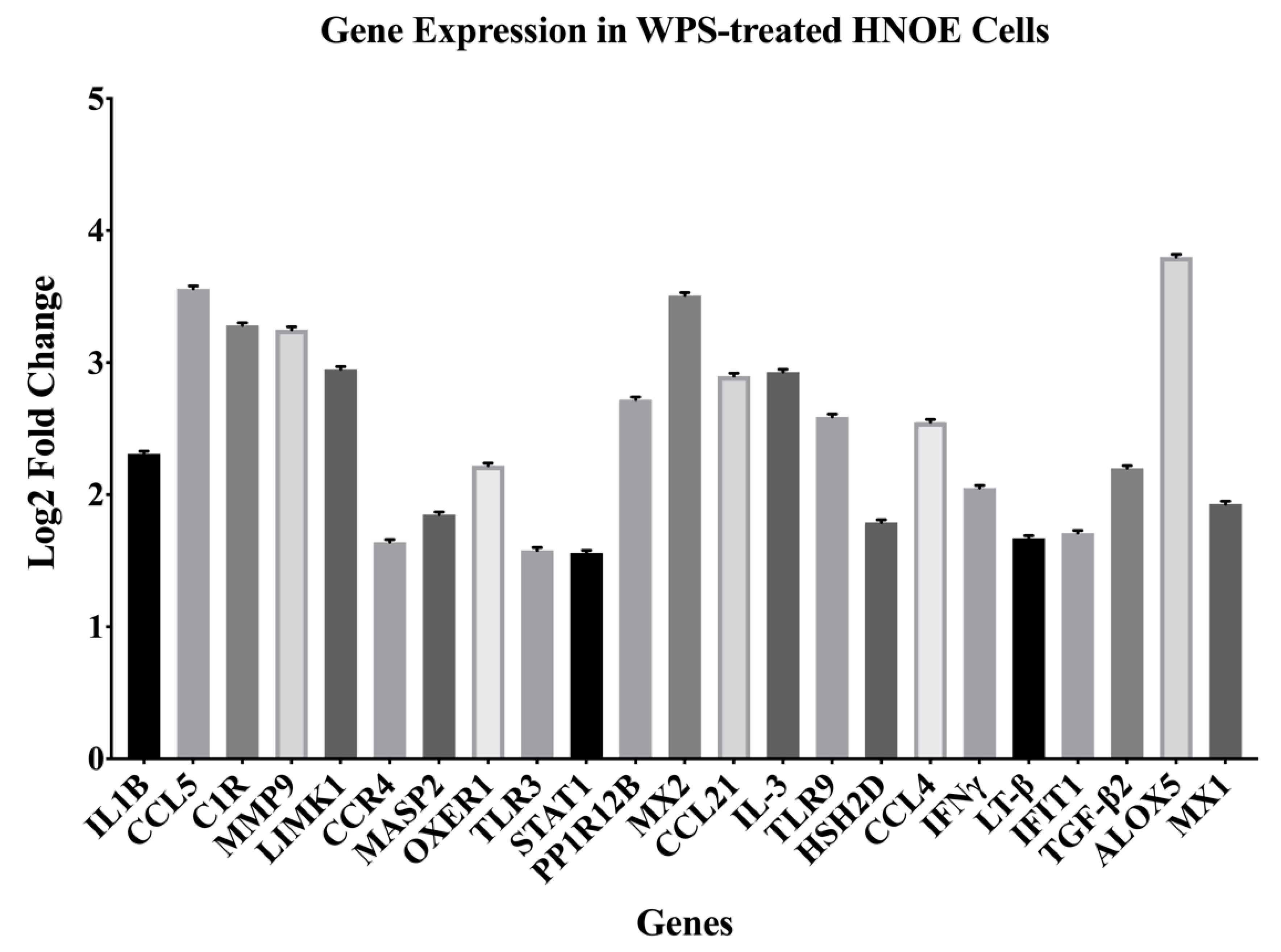
3.1. Identification of Differentially Expressed Genes (DEGs) Deregulated by WPS in HNOE Cells
3.2. Deregulated Genes Are Upregulated in HN Cancer Samples Compared with Normal Tissue
3.3. Deregulated Genes Are Upregulated in Smoking HN Cancer Patients Compared to Non-Smoker Patients
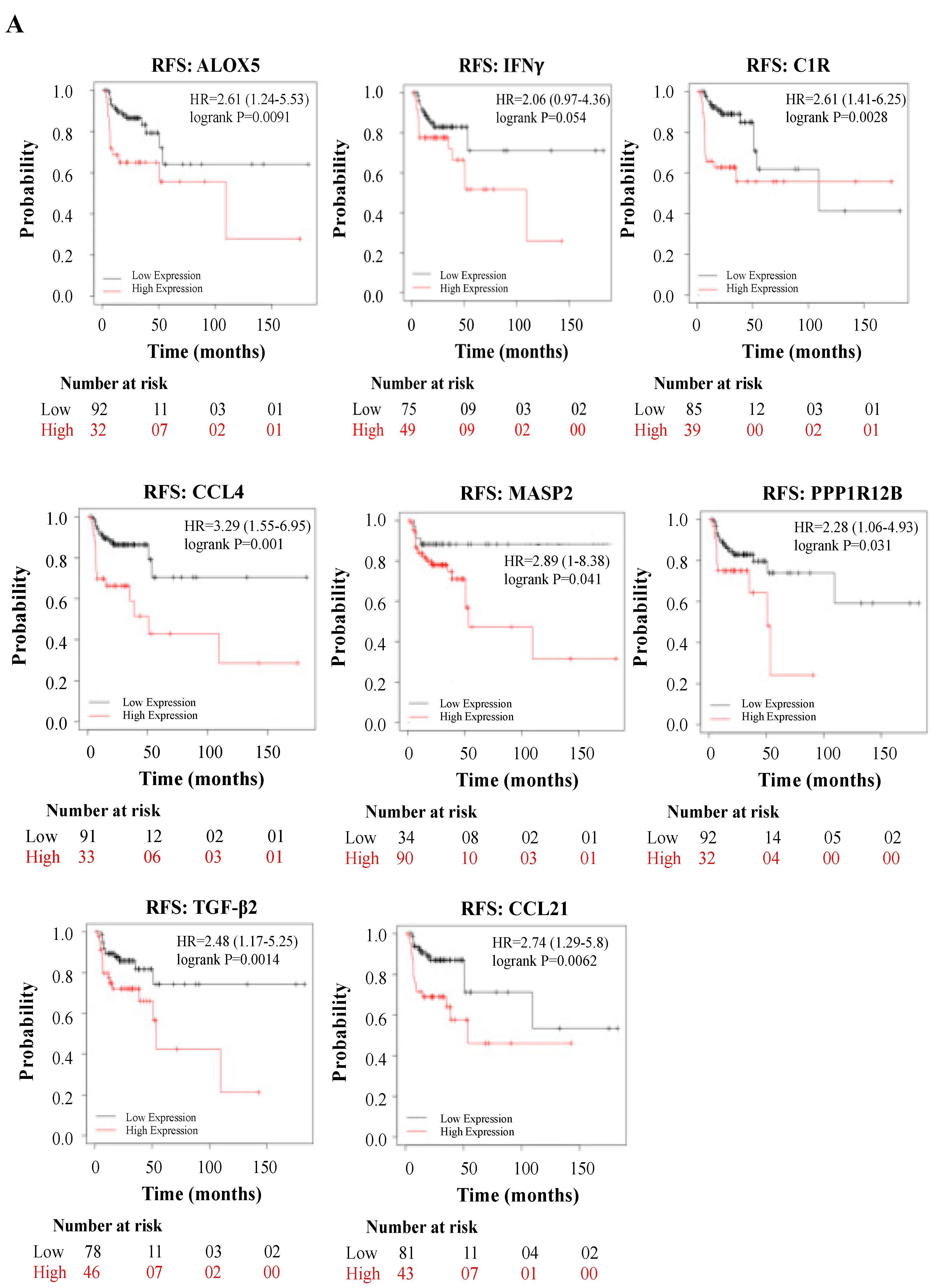
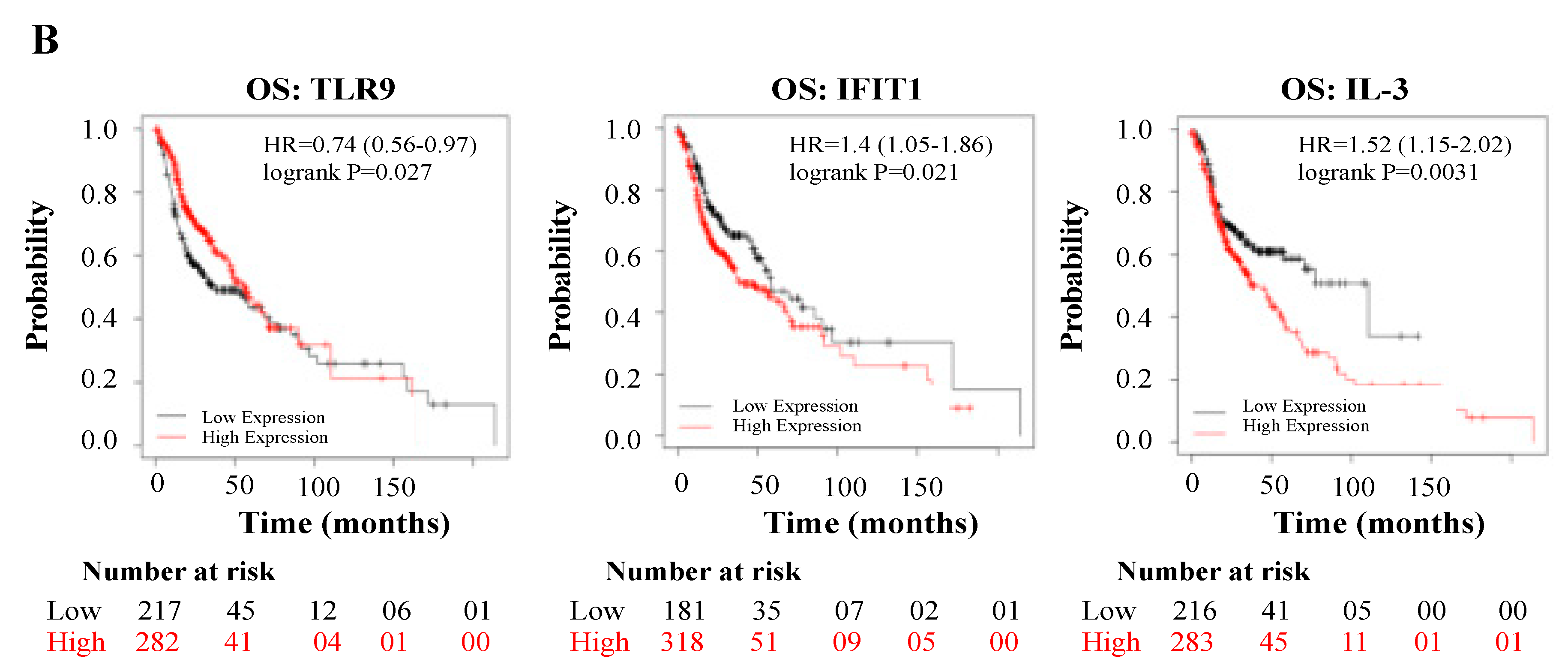
3.4. Deregulated Genes by WPS Have a Direct Impact on HN Cancer Patient’s Prognosis
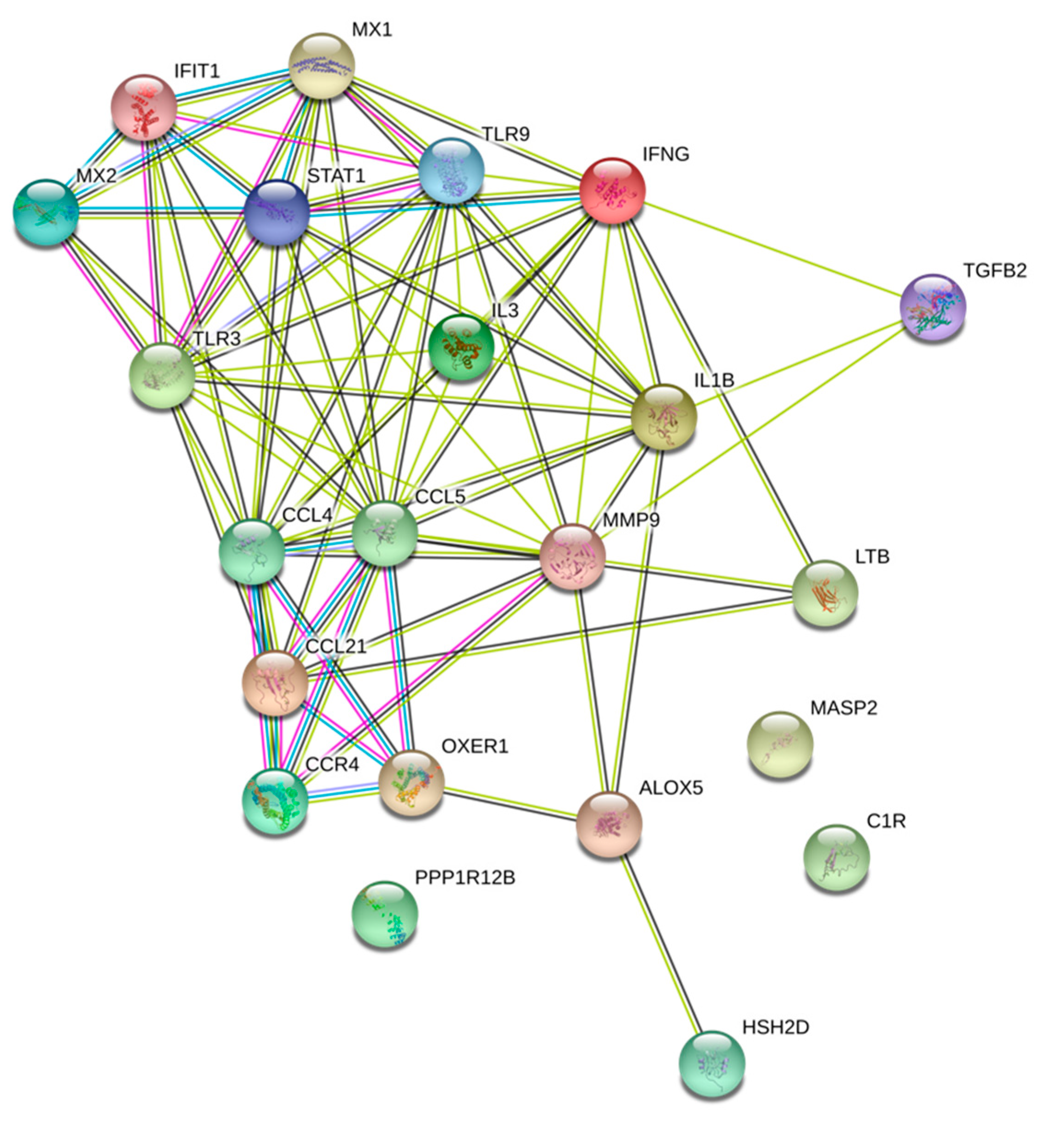
3.5. WPS Deregulated Genes Are Mainly Involved in Immune Response and Cytokine/Chemokine Mediated Pathways
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- WHO. WHO Global Report on Trends in Prevalence of Tobacco Smoking 2000–2025, 2nd ed.; WHO: Geneva, Switzerland, 2018. [Google Scholar]
- Onor, I.O.; Stirling, D.L.; Williams, S.R.; Bediako, D.; Borghol, A.; Harris, M.B.; Darensburg, T.B.; Clay, S.D.; Okpechi, S.C.; Sarpong, D.F. Clinical Effects of Cigarette Smoking: Epidemiologic Impact and Review of Pharmacotherapy Options. Int. J. Environ. Res. Public Health 2017, 14, 1147. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Maziak, W.; Taleb, Z.B.; Bahelah, R.; Islam, F.; Jaber, R.; Auf, R.; Salloum, R.G. The global epidemiology of waterpipe smoking. Tob. Control. 2015, 24 (Suppl. S1), i3–i12. [Google Scholar] [CrossRef]
- Jawad, M.; McEwen, A.; McNeill, A.; Shahab, L. To what extent should waterpipe tobacco smoking become a public health priority? Addiction 2013, 108, 1873–1884. [Google Scholar] [CrossRef] [PubMed]
- Glantz, S.A.; Bareham, D.W. E-Cigarettes: Use, Effects on Smoking, Risks, and Policy Implications. Annu. Rev. Public Health 2018, 39, 215–235. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wolfram, R.M.; Chehne, F.; Oguogho, A.; Sinzinger, H. Narghile (water pipe) smoking influences platelet function and (iso-)eicosanoids. Life Sci. 2003, 74, 47–53. [Google Scholar] [CrossRef] [PubMed]
- Neergaard, J.; Singh, P.; Job, J.; Montgomery, S. Waterpipe smoking and nicotine exposure: A review of the current evidence. Nicotine Tob. Res. 2007, 9, 987–994. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Maziak, W.; Nakkash, R.; Bahelah, R.; Husseini, A.; Fanous, N.; Eissenberg, T. Tobacco in the Arab world: Old and new epidemics amidst policy paralysis. Health Policy Plan. 2014, 29, 784–794. [Google Scholar] [CrossRef] [Green Version]
- Akl, E.A.; Gunukula, S.K.; Aleem, S.; Obeid, R.; Jaoude, P.A.; Honeine, R.; Irani, J. The prevalence of waterpipe tobacco smoking among the general and specific populations: A systematic review. BMC Public Health 2011, 11, 244. [Google Scholar] [CrossRef] [Green Version]
- Eissenberg, T.; Shihadeh, A. Waterpipe tobacco and cigarette smoking: Direct comparison of toxicant exposure. Am. J. Prev. Med. 2009, 37, 518–523. [Google Scholar] [CrossRef] [Green Version]
- Cobb, C.O.; Shihadeh, A.; Weaver, M.F.; Eissenberg, T. Waterpipe tobacco smoking and cigarette smoking: A direct comparison of toxicant exposure and subjective effects. Nicotine Tob. Res. 2011, 13, 78–87. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Maziak, W.; Ward, K.D.; Eissenberg, T. Factors related to frequency of narghile (waterpipe) use: The first insights on tobacco dependence in narghile users. Drug Alcohol Depend. 2004, 76, 101–106. [Google Scholar] [CrossRef] [PubMed]
- Rastam, S.; Eissenberg, T.; Ibrahim, I.; Ward, K.D.; Khalil, R.; Maziak, W. Comparative analysis of waterpipe and cigarette suppression of abstinence and craving symptoms. Addict. Behav. 2011, 36, 555–559. [Google Scholar] [CrossRef] [Green Version]
- Shihadeh, A.; Saleh, R. Polycyclic aromatic hydrocarbons, carbon monoxide, "tar", and nicotine in the mainstream smoke aerosol of the narghile water pipe. Food Chem. Toxicol 2005, 43, 655–661. [Google Scholar] [CrossRef] [PubMed]
- Shihadeh, A.; Schubert, J.; Klaiany, J.; El Sabban, M.; Luch, A.; Saliba, N.A. Toxicant content, physical properties and biological activity of waterpipe tobacco smoke and its tobacco-free alternatives. Tob. Control. 2015, 24 (Suppl. S1), i22–i30. [Google Scholar] [CrossRef] [Green Version]
- Shihadeh, A.; Salman, R.; Jaroudi, E.; Saliba, N.; Sepetdjian, E.; Blank, M.D.; Cobb, C.O.; Eissenberg, T. Does switching to a tobacco-free waterpipe product reduce toxicant intake? A crossover study comparing CO, NO, PAH, volatile aldehydes, "tar" and nicotine yields. Food Chem. Toxicol. 2012, 50, 1494–1498. [Google Scholar] [CrossRef] [Green Version]
- Joseph, S.; Pascale, S.; Georges, K.; Mirna, W. Cigarette and waterpipe smoking decrease respiratory quality of life in adults: Results from a national cross-sectional study. Pulm. Med. 2012, 2012, 868294. [Google Scholar] [CrossRef] [PubMed]
- Radwan, G.; Hecht, S.S.; Carmella, S.G.; Loffredo, C.A. Tobacco-specific nitrosamine exposures in smokers and nonsmokers exposed to cigarette or waterpipe tobacco smoke. Nicotine Tob. Res. 2013, 15, 130–138. [Google Scholar] [CrossRef]
- Ali, M.; Jawad, M. Health Effects of Waterpipe Tobacco Use: Getting the Public Health Message Just Right. Tob. Use Insights 2017, 10, 1179173X17696055. [Google Scholar] [CrossRef] [Green Version]
- Layoun, N.; Saleh, N.; Barbour, B.; Awada, S.; Rachidi, S.; Al-Hajje, A.; Bawab, W.; Waked, M.; Salameh, P. Waterpipe effects on pulmonary function and cardiovascular indices: A comparison to cigarette smoking in real life situation. Inhal. Toxicol. 2014, 26, 620–627. [Google Scholar] [CrossRef] [PubMed]
- Javed, F.; Al-Kheraif, A.A.; Rahman, I.; Millan-Luongo, L.T.; Feng, C.; Yunker, M.; Malmstrom, H.; Romanos, G.E. Comparison of Clinical and Radiographic Periodontal Status Between Habitual Water-Pipe Smokers and Cigarette Smokers. J. Periodontol. 2016, 87, 142–147. [Google Scholar] [CrossRef]
- Ashour, A.A.; Haik, M.Y.; Sadek, K.W.; Yalcin, H.C.; Bitharas, J.; Aboulkassim, T.; Batist, G.; Yasmeen, A.; Al Moustafa, A.E. Substantial Toxic Effect of Water-Pipe Smoking on the Early Stage of Embryonic Development. Nicotine Tob. Res. 2018, 20, 502–507. [Google Scholar] [CrossRef] [PubMed]
- Montazeri, Z.; Nyiraneza, C.; El-Katerji, H.; Little, J. Waterpipe smoking and cancer: Systematic review and meta-analysis. Tob. Control. 2017, 26, 92–97. [Google Scholar] [CrossRef]
- Rastam, S.; Li, F.M.; Fouad, F.M.; Al Kamal, H.M.; Akil, N.; Al Moustafa, A.E. Water pipe smoking and human oral cancers. Med. Hypotheses 2010, 74, 457–459. [Google Scholar] [CrossRef] [PubMed]
- Fouad, H.; Awa, F.E.; Naga, R.A.; Emam, A.H.; Labib, S.; Palipudi, K.M.; Andes, L.J.; Asma, S.; Talley, B. Prevalence of tobacco use among adults in Egypt, 2009. Glob. Health Promot. 2016, 23, 38–47. [Google Scholar] [CrossRef] [Green Version]
- Waziry, R.; Jawad, M.; Ballout, R.A.; Al Akel, M.; Akl, E.A. The effects of waterpipe tobacco smoking on health outcomes: An updated systematic review and meta-analysis. Int. J. Epidemiol 2017, 46, 32–43. [Google Scholar] [CrossRef] [Green Version]
- Sadek, K.W.; Haik, M.Y.; Ashour, A.A.; Baloch, T.; Aboulkassim, T.; Yasmeen, A.; Vranic, S.; Zeidan, A.; Al Moustafa, A.-E. Water-pipe smoking promotes epithelial-mesenchymal transition and invasion of human breast cancer cells via ERK1/ERK2 pathways. Cancer Cell Int. 2018, 18, 180. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Al Moustafa, A.-E.; Foulkes, W.D.; Benlimame, N.; Wong, A.; Yen, L.; Bergeron, J.; Batist, G.; Alpert, L.; Alaoui-Jamali, M.A. E6/E7 proteins of HPV type 16 and ErbB-2 cooperate to induce neoplastic transformation of primary normal oral epithelial cells. Oncogene 2004, 23, 350–358. [Google Scholar] [CrossRef] [Green Version]
- Sun, X.; Deng, Q.; Liang, Z.; Liu, Z.; Geng, H.; Zhao, L.; Zhou, Q.; Liu, J.; Ma, J.; Wang, D.; et al. Cigarette smoke extract induces epithelial-mesenchymal transition of human bladder cancer T24 cells through activation of ERK1/2 pathway. Biomed. Pharmacother. 2017, 86, 457–465. [Google Scholar] [CrossRef]
- Yu, D.; Geng, H.; Liu, Z.; Zhao, L.; Liang, Z.; Zhang, Z.; Xie, D.; Wang, Y.; Zhang, T.; Min, J.; et al. Cigarette smoke induced urocystic epithelial mesenchymal transition via MAPK pathways. Oncotarget 2017, 8, 8791–8800. [Google Scholar] [CrossRef] [Green Version]
- Chuang, J.-Y.; Yang, W.-H.; Chen, H.-T.; Huang, C.-Y.; Tan, T.-W.; Lin, Y.-T.; Hsu, C.-J.; Fong, Y.-C.; Tang, C.-H. CCL5/CCR5 axis promotes the motility of human oral cancer cells. J. Cell. Phys. 2009, 220, 418–426. [Google Scholar] [CrossRef]
- Bièche, I.; Lerebours, F.; Tozlu, S.; Espie, M.; Marty, M.; Lidereau, R. Molecular Profiling of Inflammatory Breast Cancer: Identification of a Poor-Prognosis Gene Expression Signature. Clin. Cancer Res. 2004, 10, 6789–6795. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Niwa, Y.; Akamatsu, H.; Niwa, H.; Sumi, H.; Ozaki, Y.; Abe, A. Correlation of Tissue and Plasma RANTES Levels with Disease Course in Patients with Breast or Cervical Cancer. Clin. Cancer Res. 2001, 7, 285–289. [Google Scholar] [PubMed]
- Yi, E.H.; Lee, C.S.; Lee, J.-K.; Lee, Y.J.; Shin, M.K.; Cho, C.-H.; Kang, K.W.; Lee, J.W.; Han, W.; Noh, D.-Y.; et al. STAT3-RANTES autocrine signaling is essential for tamoxifen resistance in human breast cancer cells. Mol. Cancer Res. 2013, 11, 31–42. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lv, D.; Zhang, Y.; Kim, H.-J.; Zhang, L.; Ma, X. CCL5 as a potential immunotherapeutic target in triple-negative breast cancer. Cell. Mol. Immunol. 2013, 10, 303–310. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Swamydas, M.; Ricci, K.; Rego, S.L.; Dréau, D. Mesenchymal stem cell-derived CCL-9 and CCL-5 promote mammary tumor cell invasion and the activation of matrix metalloproteinases. Cell Adhes. Migr. 2013, 7, 315–324. [Google Scholar] [CrossRef] [Green Version]
- Lotfi, A.; Mohammadi, G.; Tavassoli, A.; Mousaviagdas, M.; Chavoshi, H.; Saniee, L. Serum Levels of MMP9 and MMP2 in Patients with Oral Squamous Cell Carcinoma. Asian Pac. J. Cancer Prev. 2015, 16, 1327–1330. [Google Scholar] [CrossRef] [Green Version]
- Vilen, S.-T.; Salo, T.; Sorsa, T.; Nyberg, P. Fluctuating Roles of Matrix Metalloproteinase-9 in Oral Squamous Cell Carcinoma. Sci. World J. 2013, 2013, 920595. [Google Scholar] [CrossRef] [Green Version]
- Kummer, N.T.; Nowicki, T.S.; Azzi, J.P.; Reyes, I.; Iacob, C.; Xie, S.; Swati, I.; Darzynkiewicz, Z.; Gotlinger, K.H.; Suslina, N.; et al. Arachidonate 5 lipoxygenase expression in papillary thyroid carcinoma promotes invasion via MMP-9 induction. J. Cell. Biochem. 2012, 113, 1998–2008. [Google Scholar] [CrossRef] [Green Version]
- Tulah, A.S.; Parker, S.G.; Moffatt, M.F.; Wardlaw, A.J.; Connolly, M.J.; Sayers, I. The role of ALOX5AP, LTA4H and LTB4R polymorphisms in determining baseline lung function and COPD susceptibility in UK smokers. BMC Med. Genet. 2011, 12, 173. [Google Scholar] [CrossRef] [Green Version]
- Mougey, E.; Lang, J.E.; Allayee, H.; Teague, W.G.; Dozor, A.J.; Wise, R.A.; Lima, J.J. ALOX5 polymorphism associates with increased leukotriene production and reduced lung function and asthma control in children with poorly controlled asthma. Clin. Exp. Allergy 2013, 43, 512–520. [Google Scholar] [CrossRef] [Green Version]
- Kennedy-Feitosa, E.; Pinto, R.F.S.; Pires, K.M.P.; Monteiro, A.P.T.; Machado, M.N.; Santos, J.C.; Ribeiro, M.L.; Zin, W.A.; Canetti, C.A.; Romana-Souza, B.; et al. The influence of 5-lipoxygenase on cigarette smoke-induced emphysema in mice. Biochim. Biophys. Acta (BBA) Gen. Subj. 2014, 1840, 199–208. [Google Scholar] [CrossRef] [PubMed]
- Lien, M.-Y.; Lin, C.-W.; Tsai, H.-C.; Chen, Y.-T.; Tsai, M.-H.; Hua, C.-H.; Yang, S.-F.; Tang, C.-H. Impact of CCL4 gene polymorphisms and environmental factors on oral cancer development and clinical characteristics. Oncotarget 2017, 8, 31424–31434. [Google Scholar] [CrossRef] [PubMed]
- Kowshik, J.; Baba, A.B.; Giri, H.; Deepak Reddy, G.; Dixit, M.; Nagini, S. Astaxanthin inhibits JAK/STAT-3 signaling to abrogate cell proliferation, invasion and angiogenesis in a hamster model of oral cancer. PLoS ONE 2014, 9, e109114. [Google Scholar] [CrossRef]
- Li, L.; Liu, Y.-D.; Zhan, Y.-T.; Zhu, Y.-H.; Li, Y.; Xie, D.; Guan, X.-Y. High levels of CCL2 or CCL4 in the tumor microenvironment predict unfavorable survival in lung adenocarcinoma. Thorac. Cancer 2018, 9, 775–784. [Google Scholar] [CrossRef]
- Zhang, J.-F.; Li, Y.; Zhang, A.-Z.; He, Q.-Q.; Du, Y.-C.; Cao, W. Expression and pathological significance of CC chemokine receptor 7 and its ligands in the airway of asthmatic rats exposed to cigarette smoke. J. Thorac. Dis. 2018, 10, 5459–5467. [Google Scholar] [CrossRef]
- Kuznar-Kaminska, B.; Mikuła-Pietrasik, J.; Sosińska, P.; Książek, K.; Batura-Gabryel, H. COPD promotes migration of A549 lung cancer cells: The role of chemokine CCL21. Int. J. Chron. Obstruct. Pulmon. Dis. 2016, 11, 1061–1066. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Campofiorito, C.M.M.; Mangone, F.R.R.; Pasini, F.S.; Maistro, S.; Brunialti, K.C.S.; Snitcovsky, I.M.L.; Lehn, C.N.; Walder, F.; Carvalho, M.B.; Brentani, M.; et al. CCR7/CCL21 receptor ligand system may play a role in the lymph node metastasis of oral squamous cell carcinoma. Clin. Cancer Res. 2006, 12, A62. [Google Scholar]
- Domingueti, C.B.; Janini, J.B.M.; Paranaíba, L.M.R.; Lozano-Burgos, C.; Olivero, P.; González-Arriagada, W.A. Prognostic value of immunoexpression of CCR4, CCR5, CCR7 and CXCR4 in squamous cell carcinoma of tongue and floor of the mouth. Med. Oral Patol. Oral Cir. Bucal 2019, 24, e354–e363. [Google Scholar] [CrossRef]
- González-Arriagada, W.A.; Lozano-Burgos, C.; Zúñiga-Moreta, R.; González-Díaz, P.; Coletta, R.D. Clinicopathological significance of chemokine receptor (CCR1, CCR3, CCR4, CCR5, CCR7 and CXCR4) expression in head and neck squamous cell carcinomas. J. Oral Pathol. Med. 2018, 47, 755–763. [Google Scholar] [CrossRef]
- Lee, J.H.; Cho, Y.-S.; Lee, J.Y.; Kook, M.C.; Park, J.-W.; Nam, B.-H.; Bae, J.-M. The chemokine receptor CCR4 is expressed and associated with a poor prognosis in patients with gastric cancer. Ann. Surg. 2009, 249, 933–941. [Google Scholar] [CrossRef]
- Olkhanud, P.B.; Baatar, D.; Bodogai, M.; Hakim, F.; Gress, R.; Anderson, R.L.; Deng, J.; Xu, M.; Briest, S.; Biragyn, A. Breast cancer lung metastasis requires expression of chemokine receptor CCR4 and regulatory T cells. Cancer Res. 2009, 69, 5996–6004. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, J.Y.; Ou, Z.L.; Yu, S.J.; Gu, X.L.; Yang, C.; Chen, A.X.; Di, G.H.; Shen, Z.Z.; Shao, Z.M. The chemokine receptor CCR4 promotes tumor growth and lung metastasis in breast cancer. Breast Cancer Res. Treat. 2012, 131, 837–848. [Google Scholar] [CrossRef] [PubMed]
- Tsujikawa, T.; Yaguchi, T.; Ohmura, G.; Ohta, S.; Kobayashi, A.; Kawamura, N.; Fujita, T.; Nakano, H.; Shimada, T.; Takahashi, T.; et al. Autocrine and paracrine loops between cancer cells and macrophages promote lymph node metastasis via CCR4/CCL22 in head and neck squamous cell carcinoma. Int. J. Cancer 2013, 132, 2755–2766. [Google Scholar] [CrossRef]
- Gupta, S.; Srivastava, M.; Ahmad, N.; Sakamoto, K.; Bostwick, D.G.; Mukhtar, H. Lipoxygenase-5 is overexpressed in prostate adenocarcinoma. Cancer 2001, 91, 737–743. [Google Scholar] [CrossRef]
- Tomar, S.; Graves, C.A.; Altomare, D.; Kowli, S.; Kassler, S.; Sutkowski, N.; Gillespie, M.B.; Creek, K.E.; Pirisi, L. Human papillomavirus status and gene expression profiles of oropharyngeal and oral cancers from European American and African American patients. Head Neck 2016, 38 (Suppl. 1), E694–E704. [Google Scholar] [CrossRef] [Green Version]
- Kim, S.M. Human papilloma virus in oral cancer. J. Korean Assoc. Oral Maxillofac. Surg. 2016, 42, 327–336. [Google Scholar] [CrossRef] [Green Version]
- Sabatini, M.E.; Chiocca, S. Human papillomavirus as a driver of head and neck cancers. Br. J. Cancer 2020, 122, 306–314. [Google Scholar] [CrossRef]
- Chuang, H.C.; Huang, C.C.; Chien, C.Y.; Chuang, J.H. Toll-like receptor 3-mediated tumor invasion in head and neck cancer. Oral Oncol. 2012, 48, 226–232. [Google Scholar] [CrossRef]
- Luo, Q.; Hu, S.; Yan, M.; Sun, Z.; Chen, W.; Chen, F. Activation of Toll-like receptor 3 induces apoptosis of oral squamous carcinoma cells in vitro and in vivo. Int. J. Biochem. Cell Biol. 2012, 44, 1266–1275. [Google Scholar] [CrossRef]
- Pries, R.; Hogrefe, L.; Xie, L.; Frenzel, H.; Brocks, C.; Ditz, C.; Wollenberg, B. Induction of c-Myc-dependent cell proliferation through toll-like receptor 3 in head and neck cancer. Int. J. Mol. Med. 2008, 21, 209–215. [Google Scholar] [CrossRef] [Green Version]
- Pries, R.; Wulff, S.; Wollenberg, B. Toll-like receptor modulation in head and neck cancer. Crit. Rev. Immunol. 2008, 28, 201–213. [Google Scholar] [CrossRef] [PubMed]
- Min, R.; Zun, Z.; Siyi, L.; Wenjun, Y.; Lizheng, W.; Chenping, Z. Increased expression of Toll-like receptor-9 has close relation with tumour cell proliferation in oral squamous cell carcinoma. Arch. Oral Biol. 2011, 56, 877–884. [Google Scholar] [CrossRef]
- Min, R.; Siyi, L.; Wenjun, Y.; Shengwen, L.; Ow, A.; Lizheng, W.; Chenping, Z. Toll-like receptor-9 agonists increase cyclin D1 expression partly through activation of activator protein-1 in human oral squamous cell carcinoma cells. Cancer Sci. 2012, 103, 1938–1945. [Google Scholar] [CrossRef]
- Ruan, M.; Zhang, Z.; Li, S.; Yan, M.; Liu, S.; Yang, W.; Wang, L.; Zhang, C. Activation of Toll-like receptor-9 promotes cellular migration via up-regulating MMP-2 expression in oral squamous cell carcinoma. PLoS ONE 2014, 9, e92748. [Google Scholar] [CrossRef] [PubMed]
- Pisani, L.P.; Estadella, D.; Ribeiro, D.A. The Role of Toll Like Receptors (TLRs) in Oral Carcinogenesis. Anticancer Res. 2017, 37, 5389–5394. [Google Scholar]
- Riihilä, P.; Nissinen, L.; Farshchian, M.; Kivisaari, A.; Ala-Aho, R.; Kallajoki, M.; Grénman, R.; Meri, S.; Peltonen, S.; Peltonen, J.; et al. Complement factor I promotes progression of cutaneous squamous cell carcinoma. J. Invest. Dermatol. 2015, 135, 579–588. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Riihilä, P.; Viiklepp, K.; Nissinen, L.; Farshchian, M.; Kallajoki, M.; Kivisaari, A.; Meri, S.; Peltonen, J.; Peltonen, S.; Kähäri, V.M. Tumour-cell-derived complement components C1r and C1s promote growth of cutaneous squamous cell carcinoma. Br. J. Dermatol. 2020, 182, 658–670. [Google Scholar] [CrossRef] [PubMed]
- Kang, S.Y.; Toland, A.E. High risk cutaneous squamous cell carcinoma of the head and neck. World J. Otorhinolaryngol. Head Neck Surg. 2016, 2, 136–140. [Google Scholar] [CrossRef] [Green Version]
- Unterberger, C.; Hanson, S.; Klingenhoff, A.; Oesterle, D.; Frankenberger, M.; Endo, Y.; Matsushita, M.; Fujita, T.; Schwaeble, W.; Weiss, E.H.; et al. Stat3 is involved in control of MASP2 gene expression. Biochem. Biophys. Res. Commun. 2007, 364, 1022–1025. [Google Scholar] [CrossRef]
- Verma, A.; Matta, A.; Shukla, N.K.; Deo, S.V.; Gupta, S.D.; Ralhan, R. Clinical significance of mannose-binding lectin-associated serine protease-2 expression in esophageal squamous cell carcinoma. Int. J. Cancer 2006, 118, 2930–2935. [Google Scholar] [CrossRef]
- Ytting, H.; Jarle Christensen, I.; Thiel, S.; Jensenius, J.C.; Nielsen, H.J. Serum Mannan-Binding Lectin-Associated Serine Protease 2 Levels in Colorectal Cancer: Relation to Recurrence and Mortality. Clin. Cancer Res. 2005, 11, 1441–1446. [Google Scholar] [CrossRef] [Green Version]
- Swierzko, A.S.; Szala, A.; Sawicki, S.; Szemraj, J.; Sniadecki, M.; Sokolowska, A.; Kaluzynski, A.; Wydra, D.; Cedzynski, M. Mannose-Binding Lectin (MBL) and MBL-associated serine protease-2 (MASP-2) in women with malignant and benign ovarian tumours. Cancer Immunol. Immunother. 2014, 63, 1129–1140. [Google Scholar] [CrossRef] [Green Version]
- Huang, Y.-H.; Chang, C.-Y.; Kuo, Y.-Z.; Fang, W.-Y.; Kao, H.-Y.; Tsai, S.-T.; Wu, L.-W. Cancer-associated fibroblast-derived interleukin-1β activates protumor C-C motif chemokine ligand 22 signaling in head and neck cancer. Cancer Sci. 2019, 110, 2783–2793. [Google Scholar] [CrossRef] [Green Version]
- Jablonska, E.; Piotrowski, L.; Grabowska, Z. Serum Levels of IL-1b, IL-6, TNF-a, sTNF-RI and CRP in Patients with Oral Cavity Cancer. Pathol. Oncol. Res. POR 1997, 3, 126–129. [Google Scholar] [CrossRef]
- Lee, C.H.; Chang, J.S.; Syu, S.H.; Wong, T.S.; Chan, J.Y.; Tang, Y.C.; Yang, Z.P.; Yang, W.C.; Chen, C.T.; Lu, S.C.; et al. IL-1β promotes malignant transformation and tumor aggressiveness in oral cancer. J. Cell. Physiol. 2015, 230, 875–884. [Google Scholar] [CrossRef]
- Dentelli, P.; Rosso, A.; Calvi, C.; Ghiringhello, B.; Garbarino, G.; Camussi, G.; Pegoraro, L.; Brizzi, M.F. IL-3 affects endothelial cell-mediated smooth muscle cell recruitment by increasing TGFβ activity: Potential role in tumor vessel stabilization. Oncogene 2004, 23, 1681–1692. [Google Scholar] [CrossRef] [Green Version]
- Yamamoto, T.; Yoneda, K.; Ueta, E.; Osaki, T. Serum cytokines, interleukin-2 receptor, and soluble intercellular adhesion molecule-1 in oral disorders. Oral Surg. Oral Med. Oral Pathol. 1994, 78, 727–735. [Google Scholar] [CrossRef]
- Testa, U.; Riccioni, R.; Militi, S.; Coccia, E.; Stellacci, E.; Samoggia, P.; Latagliata, R.; Mariani, G.; Rossini, A.; Battistini, A.; et al. Elevated expression of IL-3Ralpha in acute myelogenous leukemia is associated with enhanced blast proliferation, increased cellularity, and poor prognosis. Blood 2002, 100, 2980–2988. [Google Scholar] [CrossRef]
- Almeida, V.L.; Santana, I.T.S.; Santos, J.N.A.; Fontes, G.S.; Lima, I.F.P.; Matos, A.L.P.; Matos, F.R.; Paranhos, L.R. Influence of interleukins on prognosis of patients with oral squamous cells carcinoma. J. Bras. Patol. Med. Lab. 2019, 55, 550–567. [Google Scholar] [CrossRef]
- Sun, L.; Diamond, M.E.; Ottaviano, A.J.; Joseph, M.J.; Ananthanarayan, V.; Munshi, H.G. Transforming growth factor-beta 1 promotes matrix metalloproteinase-9-mediated oral cancer invasion through snail expression. Mol. Cancer Res. 2008, 6, 10–20. [Google Scholar] [CrossRef] [Green Version]
- Smith, A.; Teknos, T.N.; Pan, Q. Epithelial to mesenchymal transition in head and neck squamous cell carcinoma. Oral Oncol. 2013, 49, 287–292. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cirillo, N.; Hassona, Y.; Celentano, A.; Lim, K.P.; Manchella, S.; Parkinson, E.K.; Prime, S.S. Cancer-associated fibroblasts regulate keratinocyte cell–cell adhesion via TGF-β-dependent pathways in genotype-specific oral cancer. Carcinogenesis 2016, 38, 76–85. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Davies, M.; Prime, S.S.; Eveson, J.W.; Price, N.; Ganapathy, A.; D’Mello, A.; Paterson, I.C. Transforming growth factor-β enhances invasion and metastasis in Ras-transfected human malignant epidermal keratinocytes. Int. J. Exp. Pathol. 2012, 93, 148–156. [Google Scholar] [CrossRef]
- Lu, Z.; Ding, L.; Ding, H.; Hao, F.; Pu, Y.; Wang, Y.; Chen, S.; Yang, Y.; Zhao, X.; Huang, X.; et al. Tumor cell-derived TGF-β at tumor center independently predicts recurrence and poor survival in oral squamous cell carcinoma. J. Oral Pathol. Med. 2019, 48, 696–704. [Google Scholar] [CrossRef]
- Tian, S.; Jiang, C.; Liu, X.; Xu, S.; Zhang, Z.; Chen, H.; Zhang, Y.; Liu, Y.; Ma, D. Hypermethylation of IFN-γ in oral cancer tissues. Clin. Oral Investig. 2017, 21, 2535–2542. [Google Scholar] [CrossRef]
- Katayama, A.; Ogino, T.; Bandoh, N.; Nonaka, S.; Harabuchi, Y. Expression of CXCR4 and Its Down-Regulation by IFN-γ in Head and Neck Squamous Cell Carcinoma. Clin. Cancer Res. 2005, 11, 2937–2946. [Google Scholar] [CrossRef] [Green Version]
- Wang, S.; Sun, M.; Gu, C.; Wang, X.; Chen, D.; Zhao, E.; Jiao, X.; Zheng, J. Expression of CD163, interleukin-10, and interferon-gamma in oral squamous cell carcinoma: Mutual relationships and prognostic implications. Eur. J. Oral Sci. 2014, 122, 202–209. [Google Scholar] [CrossRef]
- Li, K.; Guo, Q.; Zhang, X.; Dong, X.; Liu, W.; Zhang, A.; Li, Y.; Yan, J.; Jia, G.; Zheng, Z.; et al. Oral cancer-associated tertiary lymphoid structures: Gene expression profile and prognostic value. Clin. Exp. Immunol. 2020, 199, 172–181. [Google Scholar] [CrossRef] [PubMed]
- Das, R.; Coupar, J.; saleh, A.; Clavijo, P.E.; Chen, Z.; VanWaes, C. Abstract 358: LTB and LTBR mediates alternative Nf-kB activation through NIk and RELB/NF-kB2 to promote cell migration of HNSCC. Cancer Res. 2017, 77, 358. [Google Scholar] [CrossRef]
- Hsu, D.S.; Hwang, W.L.; Yuh, C.H.; Chu, C.H.; Ho, Y.H.; Chen, P.B.; Lin, H.S.; Lin, H.K.; Wu, S.P.; Lin, C.Y.; et al. Lymphotoxin-β Interacts with Methylated EGFR to Mediate Acquired Resistance to Cetuximab in Head and Neck Cancer. Clin. Cancer Res. 2017, 23, 4388–4401. [Google Scholar] [CrossRef] [Green Version]
- Buettner, R.; Mora, L.B.; Jove, R. Activated STAT signaling in human tumors provides novel molecular targets for therapeutic intervention. Clin. Cancer Res. 2002, 8, 945–954. [Google Scholar] [PubMed]
- Leibowitz, M.S.; Andrade Filho, P.A.; Ferrone, S.; Ferris, R.L. Deficiency of activated STAT1 in head and neck cancer cells mediates TAP1-dependent escape from cytotoxic T lymphocytes. Cancer Immunol. Immunother. CII 2011, 60, 525–535. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, Y.; Molavi, O.; Su, M.; Lai, R. The clinical and biological significance of STAT1 in esophageal squamous cell carcinoma. BMC Cancer 2014, 14, 791. [Google Scholar] [CrossRef] [Green Version]
- Pidugu, V.K.; Wu, M.M.; Yen, A.H.; Pidugu, H.B.; Chang, K.W.; Liu, C.J.; Lee, T.C. IFIT1 and IFIT3 promote oral squamous cell carcinoma metastasis and contribute to the anti-tumor effect of gefitinib via enhancing p-EGFR recycling. Oncogene 2019, 38, 3232–3247. [Google Scholar] [CrossRef]
- Pidugu, V.K.; wu, M.-M.; Pidugu, H.B.; Lee, T.-C. Abstract 2098: IFIT1 and IFIT3 modulate the drug response in human oral squamous cell carcinoma through interaction and activation of Hsp90. Cancer Res. 2019, 79, 2098. [Google Scholar] [CrossRef]
- Pidugu, V.K.; Yen, A.-H.; Chen, Y.-C.; Wu, M.-M.; Liu, C.-J.; Lee, T.-C. Abstract 3933: Characterization of oncogenic activity of interferon-induced protein with tetratricopeptide repeats 1 and 3 in human oral squamous cell carcinoma progression. Cancer Res. 2017, 77, 3933. [Google Scholar] [CrossRef]
- Chi, L.-M.; Lee, C.-W.; Chang, K.-P.; Hao, S.-P.; Lee, H.-M.; Liang, Y.; Hsueh, C.; Yu, C.-J.; Lee, I.N.; Chang, Y.-J.; et al. Enhanced interferon signaling pathway in oral cancer revealed by quantitative proteome analysis of microdissected specimens using 16O/18O labeling and integrated two-dimensional LC-ESI-MALDI tandem MS. Mol. Cell Proteom. 2009, 8, 1453–1474. [Google Scholar] [CrossRef] [Green Version]
- Calmon, M.F.; Rodrigues, R.V.; Kaneto, C.M.; Moura, R.P.; Silva, S.D.; Mota, L.D.C.; Pinheiro, D.G.; Torres, C.; de Carvalho, A.F.; Cury, P.M.; et al. Epigenetic Silencing of CRABP2 and MX1 in Head and Neck Tumors. Neoplasia 2009, 11, 1329–1339. [Google Scholar] [CrossRef] [PubMed]
- Greene, T.; Powell, P.; Nzerem, C.; Shapiro, M.; Shapiro, V. Cloning and Characterization of ALX, an Adaptor Downstream of CD28. J. Biol. Chem. 2003, 278, 45128–45134. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tsukishiro, T.; Donnenberg, A.D.; Whiteside, T.L. Rapid turnover of the CD8+CD28- T-cell subset of effector cells in the circulation of patients with head and neck cancer. Cancer Immunol. Immunother. 2003, 52, 599–607. [Google Scholar] [CrossRef]
- Grassie, M.E.; Moffat, L.D.; Walsh, M.P.; MacDonald, J.A. The myosin phosphatase targeting protein (MYPT) family: A regulated mechanism for achieving substrate specificity of the catalytic subunit of protein phosphatase type 1δ. Arch. Biochem. Biophys. 2011, 510, 147–159. [Google Scholar] [CrossRef]
- Galván, S.; Felipe-Abrio, B.; Verdugo-Sivianes, E.; Jiménez-García, M.-P.; Suarez-Martinez, E.; Marco, P.; Otero Albiol, D.; Peinado-Serrano, J.; Navas, L.; Carnero, A. PO-106 Downregulation of mypt increases tumorigenesis and resistance to platin drugs in ovarian cancer. ESMO Open. 2018, 3, A62. [Google Scholar] [CrossRef] [Green Version]
- Liang, Y.; Zhuo, Y.; Lin, Z.; Jiang, F.; Dai, Q.; Lu, J.; Dong, W.; Zhu, X.; Han, Z.; Zhong, W. Decreased Expression of MYPT1 Contributes to Tumor Angiogenesis and Poor Patient Prognosis in Human Prostate Cancer. Curr Mol. Med. 2018, 18, 100–108. [Google Scholar] [CrossRef] [PubMed]
- Andersen, Z.J.; Jørgensen, J.T.; Grøn, R.; Brauner, E.V.; Lynge, E. Active smoking and risk of breast cancer in a Danish nurse cohort study. BMC Cancer 2017, 17, 556. [Google Scholar] [CrossRef]
- White, A.J.; D’Aloisio, A.A.; Nichols, H.B.; DeRoo, L.A.; Sandler, D.P. Breast cancer and exposure to tobacco smoke during potential windows of susceptibility. Cancer Causes Control. 2017, 28, 667–675. [Google Scholar] [CrossRef]
- Strumylaite, L.; Kregzdyte, R.; Poskiene, L.; Bogusevicius, A.; Pranys, D.; Norkute, R. Association between lifetime exposure to passive smoking and risk of breast cancer subtypes defined by hormone receptor status among non-smoking Caucasian women. PLoS ONE 2017, 12, e0171198. [Google Scholar] [CrossRef] [Green Version]
- Mele, A.; Mehta, P.; Slanetz, P.J.; Brook, A.; Recht, A.; Sharma, R. Breast-Conserving Surgery Alone for Ductal Carcinoma In Situ: Factors Associated with Increased Risk of Local Recurrence. Ann. Surg. Oncol. 2017, 24, 1221–1226. [Google Scholar] [CrossRef]
- Pillai, S.; Trevino, J.; Rawal, B.; Singh, S.; Kovacs, M.; Li, X.; Schell, M.; Haura, E.; Bepler, G.; Chellappan, S. β-arrestin-1 mediates nicotine-induced metastasis through E2F1 target genes that modulate epithelial-mesenchymal transition. Cancer Res. 2015, 75, 1009–1020. [Google Scholar] [CrossRef] [Green Version]
- Liu, Y.; Luo, F.; Xu, Y.; Wang, B.; Zhao, Y.; Xu, W.; Shi, L.; Lu, X.; Liu, Q. Epithelial-mesenchymal transition and cancer stem cells, mediated by a long non-coding RNA, HOTAIR, are involved in cell malignant transformation induced by cigarette smoke extract. Toxicol. Appl. Pharmacol. 2015, 282, 9–19. [Google Scholar] [CrossRef]
- Dinicola, S.; Masiello, M.G.; Proietti, S.; Coluccia, P.; Fabrizi, G.; Catizone, A.; Ricci, G.; de Toma, G.; Bizzarri, M.; Cucina, A. Nicotine increases colon cancer cell migration and invasion through epithelial to mesenchymal transition (EMT): COX-2 involvement. J. Cell. Physiol. 2018, 233, 4935–4948. [Google Scholar] [CrossRef]
- Chen, P.C.; Lee, W.Y.; Ling, H.H.; Cheng, C.H.; Chen, K.C.; Lin, C.W. Activation of fibroblasts by nicotine promotes the epithelial-mesenchymal transition and motility of breast cancer cells. J. Cell. Physiol. 2018, 233, 4972–4980. [Google Scholar] [CrossRef] [PubMed]
- Inoue-Choi, M.; Hartge, P.; Liao, L.M.; Caporaso, N.; Freedman, N.D. Association between long-term low-intensity cigarette smoking and incidence of smoking-related cancer in the national institutes of health-AARP cohort. Int. J. Cancer 2018, 142, 271–280. [Google Scholar] [CrossRef] [PubMed]
- Lee, P.N.; Thornton, A.J.; Hamling, J.S. Epidemiological evidence on environmental tobacco smoke and cancers other than lung or breast. Regul. Toxicol. Pharmacol. 2016, 80, 134–163. [Google Scholar] [CrossRef] [Green Version]
- Liu, M.; Zhou, C.; Zheng, J. Cigarette smoking impairs the response of EGFR-TKIs therapy in lung adenocarcinoma patients by promoting EGFR signaling and epithelial-mesenchymal transition. Am. J. Transl. Res. 2015, 7, 2026–2035. [Google Scholar]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Forward (5′-3′) | Reverse (5′-3′) |
|---|---|---|
| CCL5 | GGTGCCAGCAAGATAACCCT | GCTTGCCTGACTTCCTCCTT |
| MX1 | AGGTTCCAGTAGGGCATGTG | TTGGAAAGAAGGTGCTTGCT |
| CCL21 | CTGGACAAGACACCATCCCC | TGTACTGGGGAGCCGTATCA |
| IFNγ | CTCATGTAAGCCCCCAGAAA | GCCCAGTTCCTGCAGAGTAG |
| ALOX5 | ACTTCGCCGACTTTGAGAAA | CAAGGGTGACCACAGTGATG |
| MMP9 | GTCTTGTGGAGGCTTTGAGC | CAGGGATCTCCCCTCCTTAG |
| CCL4 | GCTAAATCCAGTGGGTGGAA | GCTTGCTTCTTTTGGTTTGG |
| IL-3 | GTAGAGACGGGGTTTCACCA | GGCACAGGCCTAGAAGTGAG |
| TLR9 | CAGCAGCTCTGCAGTACGTC | AAGGCCAGGTAATTGTCACG |
| IL-1B | GGCTGCTGACTTTGAAGGAC | CATGGGAAGAAACTGGGAGA |
| LIMK1 | TCTGCAAGTGTTCGCCATAG | AGGGAGGCTCTGAAGGAAAG |
| C1R | GTTTTGGCAGGTGGCTCTTG | AGGCACAGTGGTTTCCCAAA |
| MASP2 | CCCTGGAGATTGATTCCTCA | AAACCCACTGGTCAGTTTCG |
| OXER1 | GAAACCCACCTAGGCCTCTC | TTGGAAGGGACAAACTGGAG |
| TLR3 | AGCCTTCAACGACTGATGCT | TTTCCAGAGCCGTGCTAAGT |
| STAT1 | GCAGAGACATGCCTTTGTCA | GCCACTCAGCTATTGCTTCC |
| PPP1R12B | CCAAGTTGATTCAAGCAGCA | GTTCAAGTCCAGGGCAACAT |
| MX2 | AGGTTCCAGTAGGGCATGTG | TTGGAAAGAAGGTGCTTGCT |
| HSH2D | CCACGCATGTAGGGAAGTTT | AGGGTCAGGGCTGTGTTATG |
| CCR4 | GTACTCCAACCTGGGCAAAA | CAGACTGGGTGACAGAGCAA |
| LT-β | AGGAGCCACTTCTCTGGTGA | AAAAGACCACAGGCACAACC |
| IFIT1 | CTGTGGTAGGCTCTGCTTCC | CCACCACACCCAGCTAAGTT |
| TGF-β2 | GGCAAATAGCCTGGTGTTGT | GCTGAGTTGGCATTCTGACA |
| Molecular and Cellular Functions | Genes Involved |
|---|---|
| Cellular Processes (Cell Cycle, Proliferation, Migration, Invasion, Apoptosis, and Angiogenesis) | CCR4, IL-1B, IL-3, LIMK1, MMP9 |
| Signal Transduction | CCL4, CCL21, HSH2D, IFNγ, IFIT1, MASP2, MX1, MX2, OXER1, PPP1R12B, STAT1, TGF-β2, TLR9 |
| Inflammatory Response | ALOX5, C1R, CCL4, IFNγ, LT-β |
| Molecular Function (GO) | |||
| Go-Term | Description | Count in Gene Set | False Discovery Rate |
| GO:0005126 | Cytokine receptor binding | 10 of 272 | 6.46 × 10−11 |
| GO:0005125 | Cytokine activity | 8 of 216 | 6.87 × 10−9 |
| GO:0048020 | CCR chemokine receptor binding | 4 of 41 | 5.61 × 10−6 |
| GO:0031730 | CCR5 chemokine receptor binding | 3 of 7 | 5.61 × 10−6 |
| GO:0005149 | Interlukin-1 receptor binding | 3 of 18 | 3.85 × 10−5 |
| Biological Process | |||
| Go-Term | Description | Count in Gene Set | False Discovery Rate |
| GO:0006955 | Immune response | 18 of 1560 | 1.54 × 10−13 |
| GO:0002376 | Immune system process | 20 of 2370 | 1.54 × 10−13 |
| GO:0019221 | Cytokine-mediated signaling pathway | 14 of 655 | 3.65 × 10−13 |
| GO:0071345 | Cellular response to cytokine stimulus | 15 of 953 | 1.22 × 10−12 |
| GO:0006952 | Defense response | 16 of 1234 | 1.22 × 10−12 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
López-Ozuna, V.M.; Gupta, I.; Kiow, R.L.C.; Matanes, E.; Kheraldine, H.; Yasmeen, A.; Khalil, A.; Vranic, S.; Al Moustafa, A.-E.; Farsi, H.F.A. Water-Pipe Smoking Exposure Deregulates a Set of Genes Associated with Human Head and Neck Cancer Development and Prognosis. Toxics 2020, 8, 73. https://0-doi-org.brum.beds.ac.uk/10.3390/toxics8030073
López-Ozuna VM, Gupta I, Kiow RLC, Matanes E, Kheraldine H, Yasmeen A, Khalil A, Vranic S, Al Moustafa A-E, Farsi HFA. Water-Pipe Smoking Exposure Deregulates a Set of Genes Associated with Human Head and Neck Cancer Development and Prognosis. Toxics. 2020; 8(3):73. https://0-doi-org.brum.beds.ac.uk/10.3390/toxics8030073
Chicago/Turabian StyleLópez-Ozuna, Vanessa M., Ishita Gupta, Ryan Liu Chen Kiow, Emad Matanes, Hadeel Kheraldine, Amber Yasmeen, Ashraf Khalil, Semir Vranic, Ala-Eddin Al Moustafa, and Halema F Al Farsi. 2020. "Water-Pipe Smoking Exposure Deregulates a Set of Genes Associated with Human Head and Neck Cancer Development and Prognosis" Toxics 8, no. 3: 73. https://0-doi-org.brum.beds.ac.uk/10.3390/toxics8030073