
Formaldehyde in Hospitals Induces Oxidative Stress: The Role of GSTT1 and GSTM1 Polymorphisms

 , and
, and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Population
2.2. Questionnaire, Personal Air-FA Exposure Assessment, and Biological Measurements
2.2.1. Personal Air-FA Exposure
2.2.2. OS Measurement
2.2.3. Blood Sample Collection and Inflammatory Biomarkers
2.2.4. DNA Extraction and Genotyping
2.3. Statistical Analysis
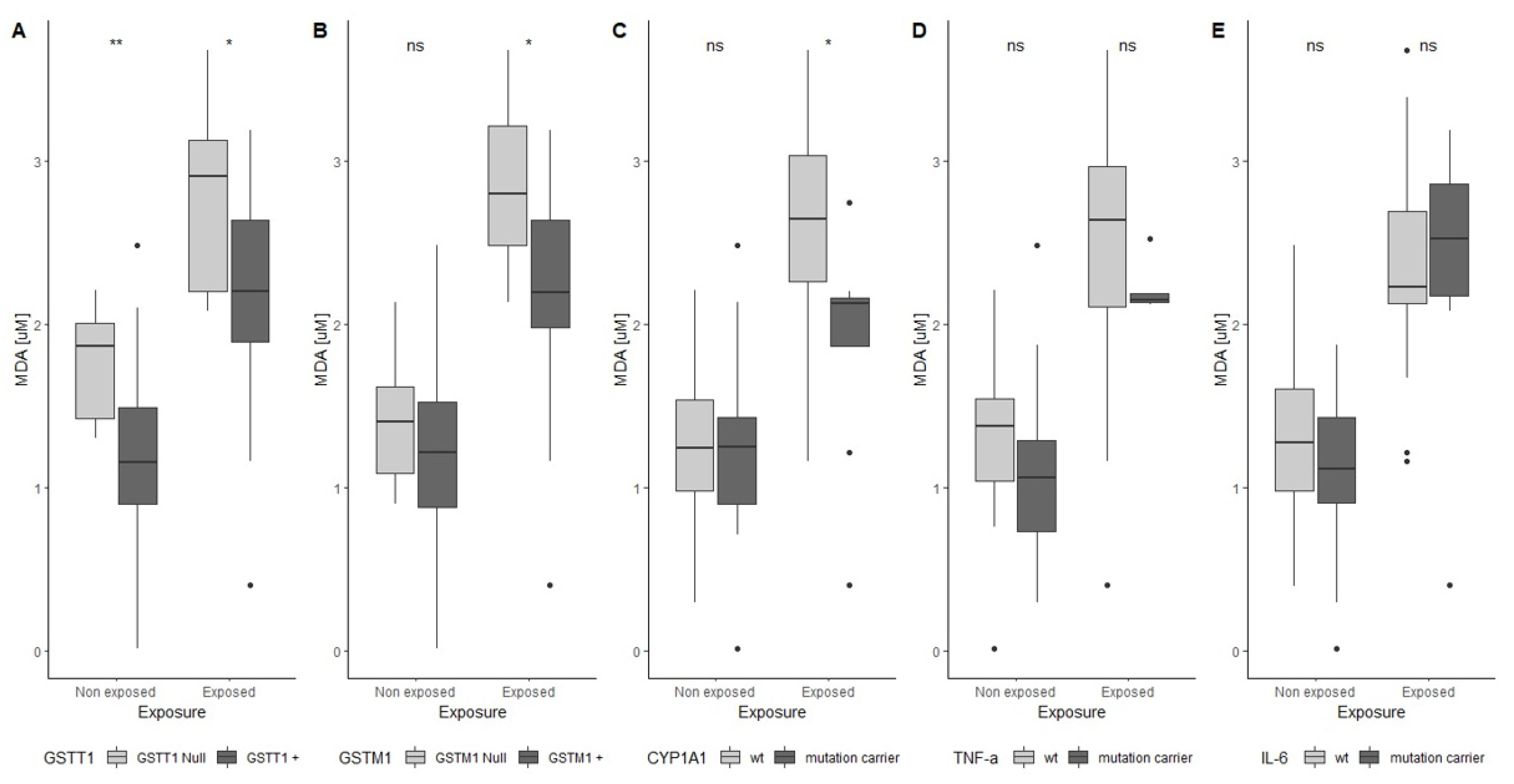
3. Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Santovito, A.; Cervella, P.; Chiarizio, M.; Meschiati, G.; Delsoglio, M.; Manitta, E.; Picco, G.; Delpero, M. Relationships between cytokine (IL-6 and TGF-β1) gene polymorphisms and chromosomal damage in hospital workers. J. Immunotoxicol. 2016, 13, 314–323. [Google Scholar] [CrossRef]
- da Silva, M.A.; Giuntini, P.B.; Meneguin, S.H. A percepção dos profissionais da saúde frente à exposição a riscos químicos. Rev. Bras. Enferm. 1997, 50, 591–598. [Google Scholar] [CrossRef] [Green Version]
- Santovito, A.; Schilirò, T.; Castellano, S.; Cervella, P.; Bigatti, M.P.; Gilli, G.; Bono, R.; DelPero, M. Combined analysis of chromosomal aberrations and glutathione S-transferase M1 and T1 polymorphisms in pathologists occupationally exposed to formaldehyde. Arch. Toxicol. 2011, 85, 1295–1302. [Google Scholar] [CrossRef]
- Santovito, A.; Cervella, P.; Delpero, M. Increased frequency of chromosomal aberrations and sister chromatid exchanges in peripheral lymphocytes of radiology technicians chronically exposed to low levels of ionizing radiations. Environ. Toxicol. Pharmacol. 2014, 37, 396–403. [Google Scholar] [CrossRef] [PubMed]
- Ioannou, S.; Andrianou, X.D.; Charisiadis, P.; Makris, K.C. Biomarkers of end of shift exposure to disinfection byproducts in nurses. J. Environ. Sci. 2017, 58, 217–223. [Google Scholar] [CrossRef] [PubMed]
- Santovito, A.; Delsoglio, M.; Manitta, E.; Picco, G.; Meschiati, G.; Chiarizio, M.; Gendusa, C.; Cervella, P. Association of GSTT1 null, XPD 751 CC and XPC 939 CC genotypes with increased levels of genomic damage among hospital pathologists. Biomarkers 2017, 22, 557–565. [Google Scholar] [CrossRef] [PubMed]
- Bellisario, V.; Mengozzi, G.; Grignani, E.; Bugiani, M.; Sapino, A.; Bussolati, G.; Bono, R. Towards a formalin-free hospital. Levels of 15-F2t-isoprostane and malondialdehyde to monitor exposure to formaldehyde in nurses from operating theatres. Toxicol. Res. (Camb.) 2016, 5, 1122–1129. [Google Scholar] [CrossRef] [Green Version]
- Ghelli, F.; Cocchi, E.; Buglisi, M.; Squillacioti, G.; Bellisario, V.; Bono, R.; Santovito, A. The role of phase I, phase II, and DNA-repair gene polymorphisms in the damage induced by formaldehyde in pathologists. Sci. Rep. 2021, 11, 10507. [Google Scholar] [CrossRef]
- Rumchev, K.B.; Spickett, J.T.; Bulsara, M.K.; Phillips, M.R.; Stick, S.M. Domestic exposure to formaldehyde significantly increases the risk of asthma in young children. Eur. Respir. J. 2002, 20, 403–408. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, X.; Zhang, Y.P.; Chen, D.; Chen, W.G.; Wang, R. Eye irritation caused by formaldehyde as an indoor air pollution—A controlled human exposure experiment. Biomed. Environ. Sci. 2001, 14, 229–236. [Google Scholar] [PubMed]
- Hauptmann, M.; Lubin, J.H.; Stewart, P.A.; Hayes, R.B.; Blair, A. Mortality from Solid Cancers among Workers in Formaldehyde Industries. Am. J. Epidemiol. 2004, 159, 1117–1130. [Google Scholar] [CrossRef] [Green Version]
- Beane Freeman, L.E.; Blair, A.; Lubin, J.H.; Stewart, P.A.; Hayes, R.B.; Hoover, R.N.; Hauptmann, M. Mortality from solid tumors among workers in formaldehyde industries: An update of the NCI cohort. Am. J. Ind. Med. 2013, 56, 1015–1026. [Google Scholar] [CrossRef]
- Allegra, A.; Spatari, G.; Mattioli, S.; Curti, S.; Innao, V.; Ettari, R.; Allegra, A.G.; Giorgianni, C.; Gangemi, S.; Musolino, C. Formaldehyde exposure and acute myeloid leukemia: A review of the literature. Medicina 2019, 55, 638. [Google Scholar] [CrossRef] [Green Version]
- Nielsen, G.D.; Larsen, S.T.; Wolkoff, P. Re-evaluation of the WHO (2010) formaldehyde indoor air quality guideline for cancer risk assessment. Arch. Toxicol. 2017, 91, 35–61. [Google Scholar] [CrossRef] [Green Version]
- IARC Working Group on the Evaluation of Carcinogenic Risks to Humans; World Health Organization; International Agency for Research on Cancer. Formaldehyde, 2-Butoxyethanol and 1-tert-Butoxypropan-2-ol; International Agency for Research on Cancer: Lyon, France, 2006; ISBN 9283212886. [Google Scholar]
- Shaham, J.; Gurvich, R.; Kaufman, Z. Sister chromatid exchange in pathology staff occupationally exposed to formaldehyde. Mutat. Res. Genet. Toxicol. Environ. Mutagen. 2002, 514, 115–123. [Google Scholar] [CrossRef]
- Motta, O.; Charlier, B.; De Caro, F.; Coglianese, A.; De Rosa, F.; Moccia, G.; Pironti, C.; Capunzo, M.; Borrelli, A.; Filippelli, A.; et al. Environmental and biological monitoring of formaldehyde inside a hospital setting: A combined approach to manage chemical risk in workplaces. J. Public Health Res. 2021, 10, 2012. [Google Scholar] [CrossRef]
- Zhang, L.; Beane Freeman, L.E.; Nakamura, J.; Hecht, S.S.; Vandenberg, J.J.; Smith, M.T.; Sonawane, B.R. Formaldehyde and leukemia: Epidemiology, potential mechanisms, and implications for risk assessment. Environ. Mol. Mutagen. 2010, 51, 181–191. [Google Scholar] [CrossRef] [Green Version]
- Chiarella, P.; Tranfo, G.; Pigini, D.; Carbonari, D. Is it possible to use biomonitoring for the quantitative assessment of formaldehyde occupational exposure? Biomark. Med. 2016, 10, 1287–1303. [Google Scholar] [CrossRef] [PubMed]
- Dugheri, S.; Bonari, A.; Pompilio, I.; Colpo, M.; Mucci, N.; Arcangeli, G. An Integrated Air Monitoring Approach for Assessment of Formaldehyde in the Workplace. Saf. Health Work 2018, 9, 479–485. [Google Scholar] [CrossRef] [PubMed]
- Dugheri, S.; Massi, D.; Mucci, N.; Berti, N.; Cappelli, G.; Arcangeli, G. Formalin safety in anatomic pathology workflow and integrated air monitoring systems for the formaldehyde occupational exposure assessment. Int. J. Occup. Med. Environ. Health 2020, 34, 1–20. [Google Scholar] [CrossRef]
- Fustinoni, S.; Campo, L.; Spinazzè, A.; Cribiù, F.M.; Chiappa, L.; Sapino, A.; Mercadante, R.; Olgiati, L.; Boniardi, L.; Cavallo, D.M.; et al. Exposure and Management of the Health Risk for the Use of Formaldehyde and Xylene in a Large Pathology Laboratory. Ann. Work Expo. Health 2021, 1–14. [Google Scholar] [CrossRef]
- D’Ettorre, G.; Criscuolo, M.; Mazzotta, M. Managing Formaldehyde indoor pollution in anatomy pathology departments. Work 2017, 56, 397–402. [Google Scholar] [CrossRef]
- Jie Seow, W.; Zhang, L.; Vermeulen, R.; Tang, X.; Hu, W.; Bassig, B.A.; Ji, Z.; Shiels, M.S.; Kemp, T.J.; Shen, M.; et al. Circulating immune/inflammation markers in Chinese workers occupationally exposed to formaldehyde. Carcinogenesis 2015, 36, 852–857. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lima, L.F.; Murta, G.L.; Bandeira, A.C.B.; Nardeli, C.R.; Lima, W.G.; Bezerra, F.S. Short-term exposure to formaldehyde promotes oxidative damage and inflammation in the trachea and diaphragm muscle of adult rats. Ann. Anat. 2015, 202, 45–51. [Google Scholar] [CrossRef] [PubMed]
- Li, L.; Hua, L.; He, Y.; Bao, Y. Differential effects of formaldehyde exposure on airway inflammation and bronchial hyperresponsiveness in BALB/c and C57BL/6 mice. PLoS ONE 2017, 12, e0179231. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Czerska, M.; Mikołajewska, K.; Zieliński, M.; Gromadzińska, J.; Wąsowicz, W. Today’s oxidative stress markers. Med. Pr. 2015, 66, 393–405. [Google Scholar] [CrossRef]
- Li, Y.R.; Trush, M. Defining ROS in Biology and Medicine. React. Oxyg. Species 2016, 1, 9. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Khoubnasabjafari, M.; Ansarin, K.; Jouyban, A. Reliability of malondialdehyde as a biomarker of oxidative stress in psychological disorders. BioImpacts 2015, 5, 123–127. [Google Scholar] [CrossRef]
- Niki, E. Biomarkers of lipid peroxidation in clinical material. Biochim. Biophys. Acta Gen. Subj. 2014, 1840, 809–817. [Google Scholar] [CrossRef]
- Milne, G.L.; Dai, Q.; Roberts, L.J. The isoprostanes—25 years later. Biochim. Biophys. Acta 2015, 1851, 433–445. [Google Scholar] [CrossRef] [Green Version]
- Tan, B.L.; Norhaizan, M.E.; Liew, W.P.P.; Rahman, H.S. Antioxidant and oxidative stress: A mutual interplay in age-related diseases. Front. Pharmacol. 2018, 9, 1162. [Google Scholar] [CrossRef] [Green Version]
- Oeltzschner, G.; Butz, M.; Wickrath, F.; Wittsack, H.J.; Schnitzler, A. Covert hepatic encephalopathy: Elevated total glutathione and absence of brain water content changes. Metab. Brain Dis. 2016, 31, 517–527. [Google Scholar] [CrossRef] [PubMed]
- Haffner, M.J.; Oakes, P.; Demerdash, A.; Yammine, K.C.; Watanabe, K.; Loukas, M.; Tubbs, R.S. Formaldehyde exposure and its effects during pregnancy: Recommendations for laboratory attendance based on available data. Clin. Anat. 2015, 28, 972–979. [Google Scholar] [CrossRef] [PubMed]
- Zmorzyński, S.; Świderska-Kołacz, G.; Koczkodaj, D.; Filip, A.A. Significance of Polymorphisms and Expression of Enzyme-Encoding Genes Related to Glutathione in Hematopoietic Cancers and Solid Tumors. BioMed Res. Int. 2015, 2015, 853573. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pisoschi, A.M.; Pop, A. The role of antioxidants in the chemistry of oxidative stress: A review. Eur. J. Med. Chem. 2015, 97, 55–74. [Google Scholar] [CrossRef]
- Moreno-Macias, H.; Romieu, I. Effects of antioxidant supplements and nutrients on patients with asthma and allergies. J. Allergy Clin. Immunol. 2014, 133, 1237–1244. [Google Scholar] [CrossRef] [PubMed]
- Jia, X.; Jia, Q.; Zhang, Z.; Gao, W.; Zhang, X.; Niu, Y.; Meng, T.; Feng, B.; Duan, H.; Ye, M.; et al. Effects of Formaldehyde on Lymphocyte Subsets and Cytokines in the Peripheral Blood of Exposed Workers. PLoS ONE 2014, 9, e104069. [Google Scholar] [CrossRef] [Green Version]
- Aydın, S.; Canpınar, H.; Ündeğer, Ü.; Güç, D.; Çolakoğlu, M.; Kars, A.; Başaran, N. Assessment of immunotoxicity and genotoxicity in workers exposed to low concentrations of formaldehyde. Arch. Toxicol. 2013, 87, 145–153. [Google Scholar] [CrossRef]
- Kastner, P.E.; Casset, A.; Pons, F. Formaldehyde interferes with airway epithelium integrity and functions in a dose- and time-dependent manner. Toxicol. Lett. 2011, 200, 109–116. [Google Scholar] [CrossRef]
- Balharry, D.; Sexton, K.; BéruBé, K.A. An in vitro approach to assess the toxicity of inhaled tobacco smoke components: Nicotine, cadmium, formaldehyde and urethane. Toxicology 2008, 244, 66–76. [Google Scholar] [CrossRef]
- Wang, F.; Li, C.; Liu, W.; Jin, Y. Effect of exposure to volatile organic compounds (VOCs) on airway inflammatory response in mice. J. Toxicol. Sci. 2012, 37, 739–748. [Google Scholar] [CrossRef] [Green Version]
- Christiani, D.C.; Mehta, A.J.; Yu, C.L. Genetic susceptibility to occupational exposures. Occup. Environ. Med. 2008, 65, 430–436. [Google Scholar] [CrossRef] [Green Version]
- Parveen, F.; Faridi, R.M.; Das, V.; Tripathi, G.; Agrawal, S. Genetic association of phase I and phase II detoxification genes with recurrent miscarriages among North Indian women. Mol. Hum. Reprod. 2010, 16, 207–214. [Google Scholar] [CrossRef] [Green Version]
- Yang, X.-L.; Xie, S.; Jiang, Y.-Y.; Shi, C.; Cai, Z.-G.; Chen, S.-X. Association between CYP1A1 Ile462Val Polymorphism and Oral Squamous Cell Carcinoma Susceptibility: Evidence from 13 Investigations. J. Cancer 2015, 6, 302–309. [Google Scholar] [CrossRef] [Green Version]
- Hollman, A.L.; Tchounwou, P.B.; Huang, H.-C. The Association between Gene-Environment Interactions and Diseases Involving the Human GST Superfamily with SNP Variants. Int. J. Environ. Res. Public Health 2020, 13, 379. [Google Scholar] [CrossRef] [PubMed]
- Squillacioti, G.; Bellisario, V.; Grosso, A.; Ghelli, F.; Piccioni, P.; Grignani, E.; Corsico, A.; Bono, R. Formaldehyde, Oxidative Stress, and FeNO in Traffic Police Officers Working in Two Cities of Northern Italy. Int. J. Environ. Res. Public Health 2020, 17, 1655. [Google Scholar] [CrossRef] [Green Version]
- Ghelli, F.; Bellisario, V.; Squillacioti, G.; Grignani, E.; Garzaro, G.; Buglisi, M.; Bergamaschi, E.; Bono, R. Oxidative stress induction in woodworkers occupationally exposed to wood dust and formaldehyde. J. Occup. Med. Toxicol. 2021, 16, 4. [Google Scholar] [CrossRef]
- Bartels, H.; Cikes, M. Über chromogene der kreatininbestimmung nach Jaffé. Clin. Chim. Acta 1969, 26, 1–10. [Google Scholar] [CrossRef]
- De Moraes Martins, C.J.; Genelhu, V.; Pimentel, M.M.G.; Celoria, B.M.J.; Mangia, R.F.; Aveta, T.; Silvestri, C.; Di Marzo, V.; Francischetti, E.A. Circulating endocannabinoids and the polymorphism 385C>A in fatty acid amide hydrolase (FAAH) gene may identify the obesity phenotype related to cardiometabolic risk: A study conducted in a Brazilian population of complex interethnic admixture. PLoS ONE 2015, 10, e0142728. [Google Scholar] [CrossRef]
- Chen, S.; Xue, K.; Xu, L.; Ma, G.; Wu, J. Polymorphisms of the CYP1A1 and GSTM1 genes in relation to individual susceptibility to lung carcinoma in Chinese population. Mutat. Res. Mutat. Res. Genom. 2001, 458, 41–47. [Google Scholar] [CrossRef]
- Pemble, S.; Schroeder, K.R.; Spencer, S.R.; Meyer, D.J.; Hallier, E.; Bolt, H.M.; Ketterer, B.; Taylor, J.B. Human glutathione S-transferase Theta (GSTT1): cDNA cloning and the characterization of a genetic polymorphism. Biochem. J. 1994, 300, 271–276. [Google Scholar] [CrossRef] [Green Version]
- Zhong, S.; Wyllie, A.H.; Barnes, D.; Wolf, C.R.; Spurr, N.K. Relationship between the gstm1 genetic polymorphism and susceptibility to bladder, breast and colon cancer. Carcinogenesis 1993, 14, 1821–1824. [Google Scholar] [CrossRef]
- Perrey, C.; Turner, S.J.; Pravica, V.; Howell, W.M.; Hutchinson, I.V. ARMS-PCR methodologies to determine IL-10, TNF-α, TNF-β and TGF-β1 gene polymorphisms. Transpl. Immunol. 1999, 7, 127–128. [Google Scholar] [CrossRef]
- Zakharyan, R.; Petrek, M.; Arakelyan, A.; Mrazek, F.; Atshemyan, S.; Boyajyan, A. Interleukin-6 promoter polymorphism and plasma levels in patients with schizophrenia. Tissue Antigens 2012, 80, 136–142. [Google Scholar] [CrossRef] [PubMed]
- Titford, M.E.; Horenstein, M.G. Histomorphologic Assessment of Formalin Substitute Fixatives for Diagnostic Surgical Pathology. Arch. Pathol. Lab. Med. 2005, 129, 502–506. [Google Scholar] [CrossRef] [PubMed]
- Patil, S.; Rao, R.S.; Ganavi, B.S.; Majumdar, B. Natural sweeteners as fixatives in histopathology: A longitudinal study. J. Nat. Sci. Biol. Med. 2015, 6, 67–70. [Google Scholar] [CrossRef] [Green Version]
- Ye, X.; Ji, Z.; Wei, C.; McHale, C.M.; Ding, S.; Thomas, R.; Yang, X.; Zhang, L. Inhaled formaldehyde induces DNA-protein crosslinks and oxidative stress in bone marrow and other distant organs of exposed mice. Environ. Mol. Mutagen. 2013, 54, 705–718. [Google Scholar] [CrossRef]
- Li, X.; Zhang, F.; Zhou, H.; Hu, Y.; Guo, D.; Fang, X.; Chen, Y. Interplay of TNF-α, soluble TNF receptors and oxidative stress in coronary chronic total occlusion of the oldest patients with coronary heart disease. Cytokine 2020, 125, 154836. [Google Scholar] [CrossRef]
- Oztan, O.; Tutkun, L.; Turksoy, V.A.; Deniz, S.; Dip, A.; Iritas, S.B.; Eravci, D.B.; Alaguney, M.E. The relationship between impaired lung functions and cytokine levels in formaldehyde exposure. Arch. Environ. Occup. Health 2021, 76, 248–254. [Google Scholar] [CrossRef]
- Reuter, S.; Gupta, S.C.; Chaturvedi, M.M.; Aggarwal, B.B. Oxidative stress, inflammation, and cancer: How are they linked? Free Radic. Biol. Med. 2010, 49, 1603–1616. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Klaunig, J.E.; Wang, Z. Oxidative stress in carcinogenesis. Curr. Opin. Toxicol. 2018, 7, 116–121. [Google Scholar] [CrossRef]
- Bono, R.; Romanazzi, V.; Munnia, A.; Piro, S.; Allione, A.; Ricceri, F.; Guarrera, S.; Pignata, C.; Matullo, G.; Wang, P.; et al. Malondialdehyde−Deoxyguanosine Adduct Formation in Workers of Pathology Wards: The Role of Air Formaldehyde Exposure. Chem. Res. Toxicol. 2010, 23, 1342–1348. [Google Scholar] [CrossRef] [Green Version]
- Martinez-Moral, M.P.; Kannan, K. How stable is oxidative stress level? An observational study of intra- and inter-individual variability in urinary oxidative stress biomarkers of DNA, proteins, and lipids in healthy individuals. Environ. Int. 2019, 123, 382–389. [Google Scholar] [CrossRef] [PubMed]
- Basu, S. The enigma of in vivo oxidative stress assessment: Isoprostanes as an emerging target. Scand. J. Food Nutr. 2007, 51, 48–61. [Google Scholar] [CrossRef]
- Datta, S.K.; Kumar, V.; Pathak, R.; Tripathi, A.K.; Ahmed, R.S.; Kalra, O.P.; Banerjee, B.D. Association of glutathione S -transferase M1 and T1 gene polymorphism with oxidative stress in diabetic and nondiabetic chronic kidney disease. Ren. Fail. 2010, 32, 1189–1195. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hong, Y.C.; Lee, K.H.; Yi, C.H.; Ha, E.H.; Christiani, D.C. Genetic susceptibility of term pregnant women to oxidative damage. Toxicol. Lett. 2002, 129, 255–262. [Google Scholar] [CrossRef]
- Vibhuti, A.; Arif, E.; Mishra, A.; Deepak, D.; Singh, B.; Rahman, I.; Mohammad, G.; Pasha, M.A.Q. CYP1A1, CYP1A2 and CYBA gene polymorphisms associated with oxidative stress in COPD. Clin. Chim. Acta 2010, 411, 474–480. [Google Scholar] [CrossRef] [PubMed]
- Allocati, N.; Masulli, M.; Di Ilio, C.; Federici, L. Glutathione transferases: Substrates, inihibitors and pro-drugs in cancer and neurodegenerative diseases. Oncogenesis 2018, 7, 1–15. [Google Scholar] [CrossRef]
- Ho, Y.-C.; Huang, F.-M.; Chang, Y.-C. Cytotoxicity of formaldehyde on human osteoblastic cells is related to intracellular glutathione levels. J. Biomed. Mater. Res. Part B Appl. Biomater. 2007, 83, 340–344. [Google Scholar] [CrossRef]
- Tulpule, K.; Hohnholt, M.C.; Dringen, R. Formaldehyde metabolism and formaldehyde-induced stimulation of lactate production and glutathione export in cultured neurons. J. Neurochem. 2013, 125, 260–272. [Google Scholar] [CrossRef]
- Kumar, V.; Yadav, C.S.; Datta, S.K.; Singh, S.; Ahmed, R.S.; Goel, S.; Gupta, S.; Mustafa, M.; Grover, R.K.; Banerjee, B.D. Association of GSTM1 and GSTT1 polymorphism with lipid peroxidation in benign prostate hyperplasia and prostate cancer: A pilot study. Dis. Mark. 2011, 30, 163–169. [Google Scholar] [CrossRef]
- Costa, S.; Carvalho, S.; Costa, C.; Coelho, P.; Silva, S.; Santos, L.S.; Gaspar, J.F.; Porto, B.; Laffon, B.; Teixeira, J.P. Increased levels of chromosomal aberrations and DNA damage in a group of workers exposed to formaldehyde. Mutagenesis 2015, 30, 463–473. [Google Scholar] [CrossRef] [PubMed] [Green Version]


{kind=link}
| Gene | Polymorphism NCBI | Main Function Protein | Sequence | T (°C) | Methodology | PCR Product Size (bp) | Reference |
|---|---|---|---|---|---|---|---|
| CYP1A1 (A > G) | rs1048943 | Phase-I metabolic enzyme | 5′-AAGACCTCCCAGCGGGCAAT-3′ 5′-AAGACCTCCCAGCGGGCAAC-3′ 5′-CTCTGGTTACAGGAAGCTAT-3′ | 60 | ARMS-PCR | 162 | [51] |
| GSTT1 (Presence/ Absence) | rs1601993659 | Phase-II metabolic enzyme | 5′-TTCCTTACTGGTCCTCACATCTC-3′ 5′-TCACCGGATCATGGCCAGCA-3′ | 63 | PCR | 480 | [52] |
| GSTM1 (Presence/ Absence) | rs1183423000 | Phase-II metabolic enzyme | 5-CTGGATTGTAGCAGATCATGC-3′ 5′-CTGCCCTACTTGATTGATGGG-3′ | 65 | PCR | 273 | [53] |
| TNF-α (−308, G > A) Antisense primer G-sense primer A-sense primer | rs1800629 | Pro-inflammatory | 5′-TCTCGGTTTCTTCTCCATCG-3′ 5′-ATAGGTTTTGAGGGGCATGG-3′ 5-AATAGGTTTTGAGGGGCATGA-3′ | 60 | ARMS-PCR | 184 | [54] |
| IL-6 (−174, G > C) Antisense primer G-sense primer C-sense primer | rs1800796 | Pro-inflammatory | 5′-TCGTGCATGACTTCAGCTTTA-3′ 5′-AATGTGACGTCCTTTAGCATG-3′ 5′-AATGTGACGTCCTTTAGCATC-3′ | 60 | ARMS-PCR | 190 | [55] |
| Characteristics | Formalin-Employers (n = 23) | Non-Employers (n = 45) | p-Value | ||
|---|---|---|---|---|---|
| n (%) | n (%) | ||||
| Sex | |||||
| Male | 5 (21.7) | 7 (15.6) | 0.522 | ||
| Female | 18 (78.3) | 38 (84.4) | |||
| Smoking | |||||
| Yes | 7 (30.4) | 12 (26.7) | 0.780 | ||
| No | 16 (69.6) | 33 (73.3) | |||
| Task | |||||
| Healthcare assistant | 4 (17.4) | 11 (24.4) | 0.133 | ||
| Nurse | 7 (30.4) | 20 (44.4) | |||
| Laboratory technician | 7 (30.4) | 12 (26.7) | |||
| Pathologist | 5 (21.7) | 2 (4.4) | |||
| Working years | |||||
| <5 | 3 (13.0) | 5 (11.1) | 0.973 | ||
| 5–10 | 5 (21.7) | 10 (22.2) | |||
| >10 | 15 (65.2) | 30 (66.7) | |||
| Median | IQR | Median | IQR | ||
| Age (years) | 44.00 | 14.00 | 49.00 | 13.00 | 0.369 |
| BMI (kg/m2) | 23.44 | 4.20 | 23.40 | 7.32 | 0.429 |
| Parameters | Formalin-Employers (n = 23) | Non-Employers (n = 45) | p-Value | ||
|---|---|---|---|---|---|
| Median | IQR | Median | IQR | ||
| FA (µg/m3) | 57.3 | 125.9 | 13.5 | 12.7 | <0.001 |
| 15-F2t-IsoP (ng/mg CREA) | 16.06 | 14.98 | 2.16 | 2.67 | <0.001 |
| MDA (μM) | 2.29 | 0.67 | 1.25 | 0.64 | <0.001 |
| TNF-α (pg/mL) | 15.35 | 16.51 | 11.59 | 7.31 | 0.023 |
| n (%) | n (%) | ||||
| GSTT1 | |||||
| GSTT+ | 15 (65.2) | 38 (84.4) | 0.120 | ||
| GSTT Null | 8 (34.8) | 7 (15.6) | |||
| GSTM1 | |||||
| GSTM+ | 16 (69.6) | 34 (75.6) | 0.772 | ||
| GSTM Null | 7 (30.4) | 11 (24.4) | |||
| CYP1A1 | |||||
| wt | 15 (65.2) | 32 (71.1) | 0.782 | ||
| mutation carrier | 8 (34.8) | 13 (28.9) | |||
| TNF-α | |||||
| wt | 18 (78.3) | 30 (66.7) | 0.405 | ||
| mutation carrier | 5 (21.7) | 15 (33.3) | |||
| IL-6 | |||||
| wt | 15 (65.2) | 32 (71.1) | 0.782 | ||
| mutation carrier | 8 (34.8) | 13 (28.9) | |||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ghelli, F.; Bellisario, V.; Squillacioti, G.; Panizzolo, M.; Santovito, A.; Bono, R. Formaldehyde in Hospitals Induces Oxidative Stress: The Role of GSTT1 and GSTM1 Polymorphisms. Toxics 2021, 9, 178. https://0-doi-org.brum.beds.ac.uk/10.3390/toxics9080178
Ghelli F, Bellisario V, Squillacioti G, Panizzolo M, Santovito A, Bono R. Formaldehyde in Hospitals Induces Oxidative Stress: The Role of GSTT1 and GSTM1 Polymorphisms. Toxics. 2021; 9(8):178. https://0-doi-org.brum.beds.ac.uk/10.3390/toxics9080178
Chicago/Turabian StyleGhelli, Federica, Valeria Bellisario, Giulia Squillacioti, Marco Panizzolo, Alfredo Santovito, and Roberto Bono. 2021. "Formaldehyde in Hospitals Induces Oxidative Stress: The Role of GSTT1 and GSTM1 Polymorphisms" Toxics 9, no. 8: 178. https://0-doi-org.brum.beds.ac.uk/10.3390/toxics9080178