
Subacute Exposure to an Environmentally Relevant Dose of Di-(2-ethylhexyl) Phthalate during Gestation Alters the Cecal Microbiome, but Not Pregnancy Outcomes in Mice


 and
and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Chemicals
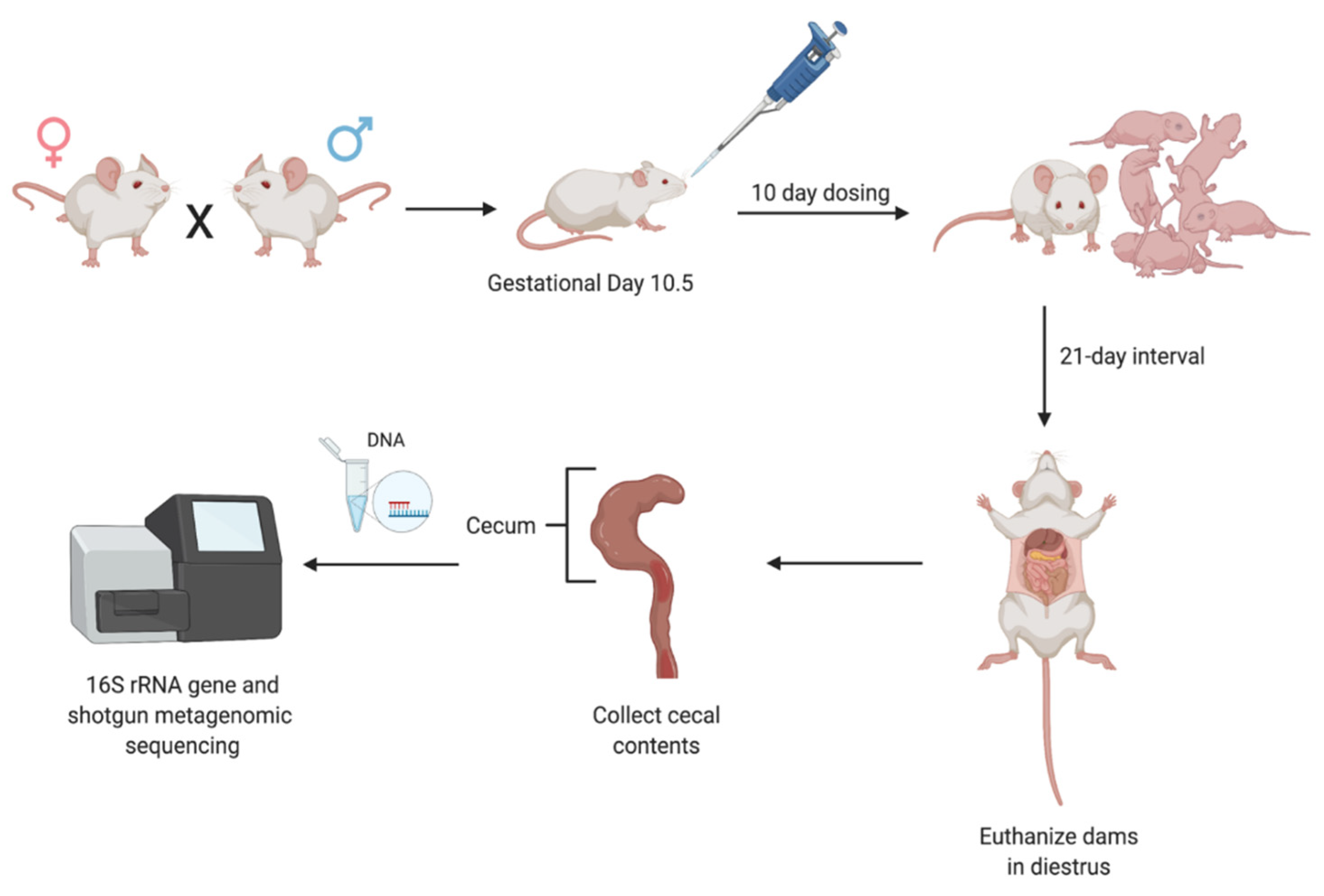
2.2. Experimental Animals
2.3. Experimental Design
2.4. Cecal Collection
2.5. DNA Isolation and Quantification
2.6. Processing of Microbial Genomic DNA for 16S rRNA Gene and Metagenomic Sequencing
2.7. Bioinformatic Analyses of 16S rRNA Gene Data
2.8. Bioinformatic Analyses of Metagenomic Data
2.9. Pregnancy Outcomes
2.10. Statistical Analysis
3. Results
3.1. Cecal Microbiota
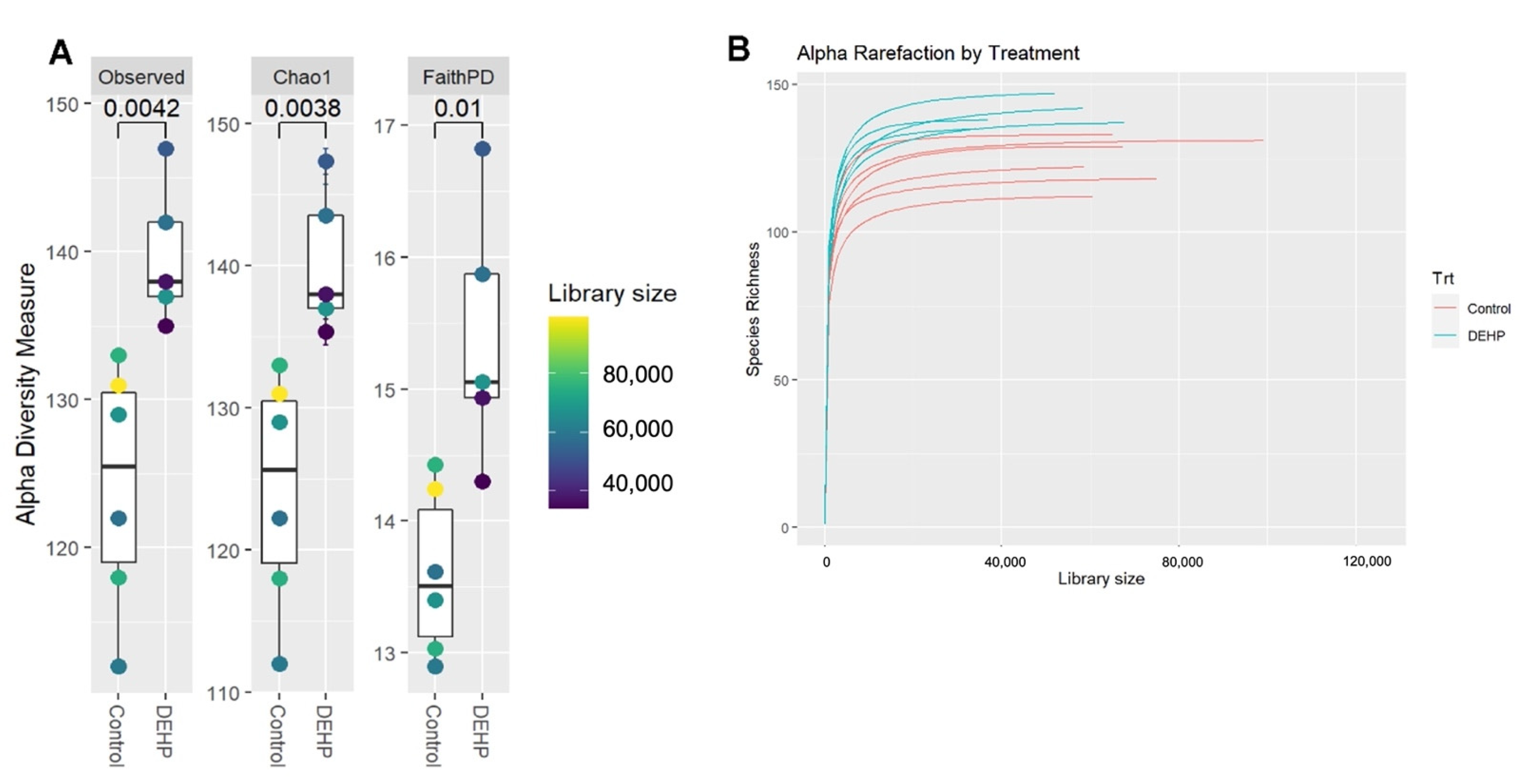
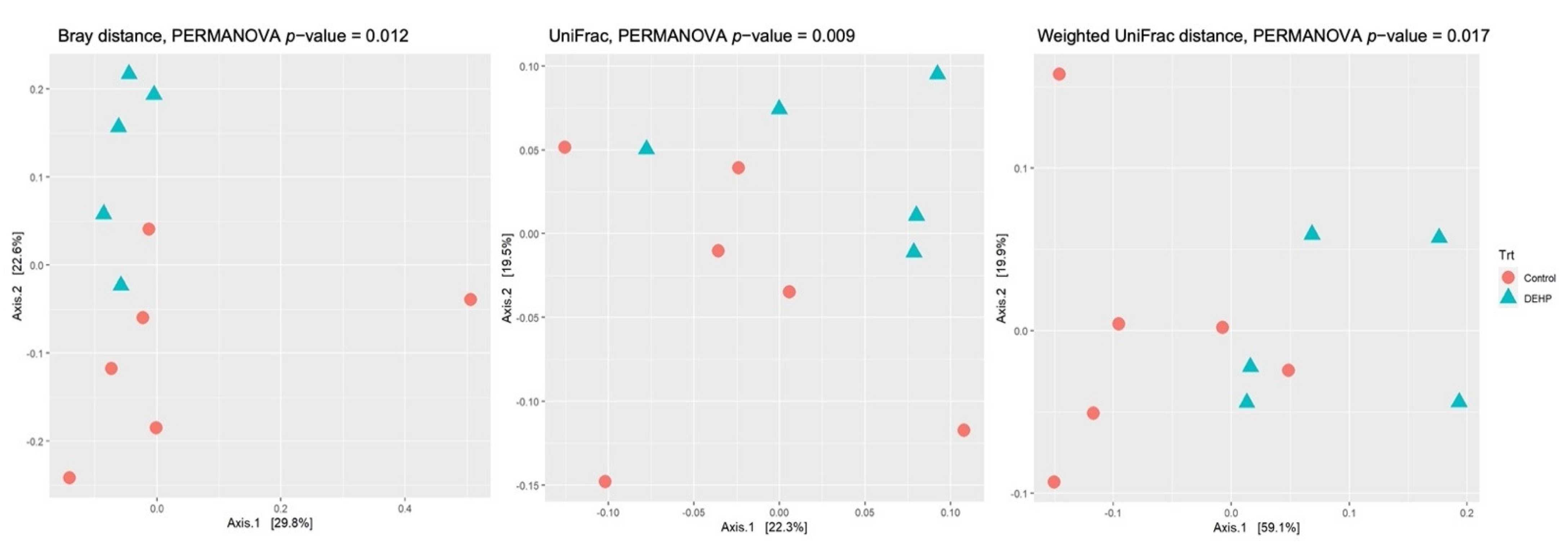
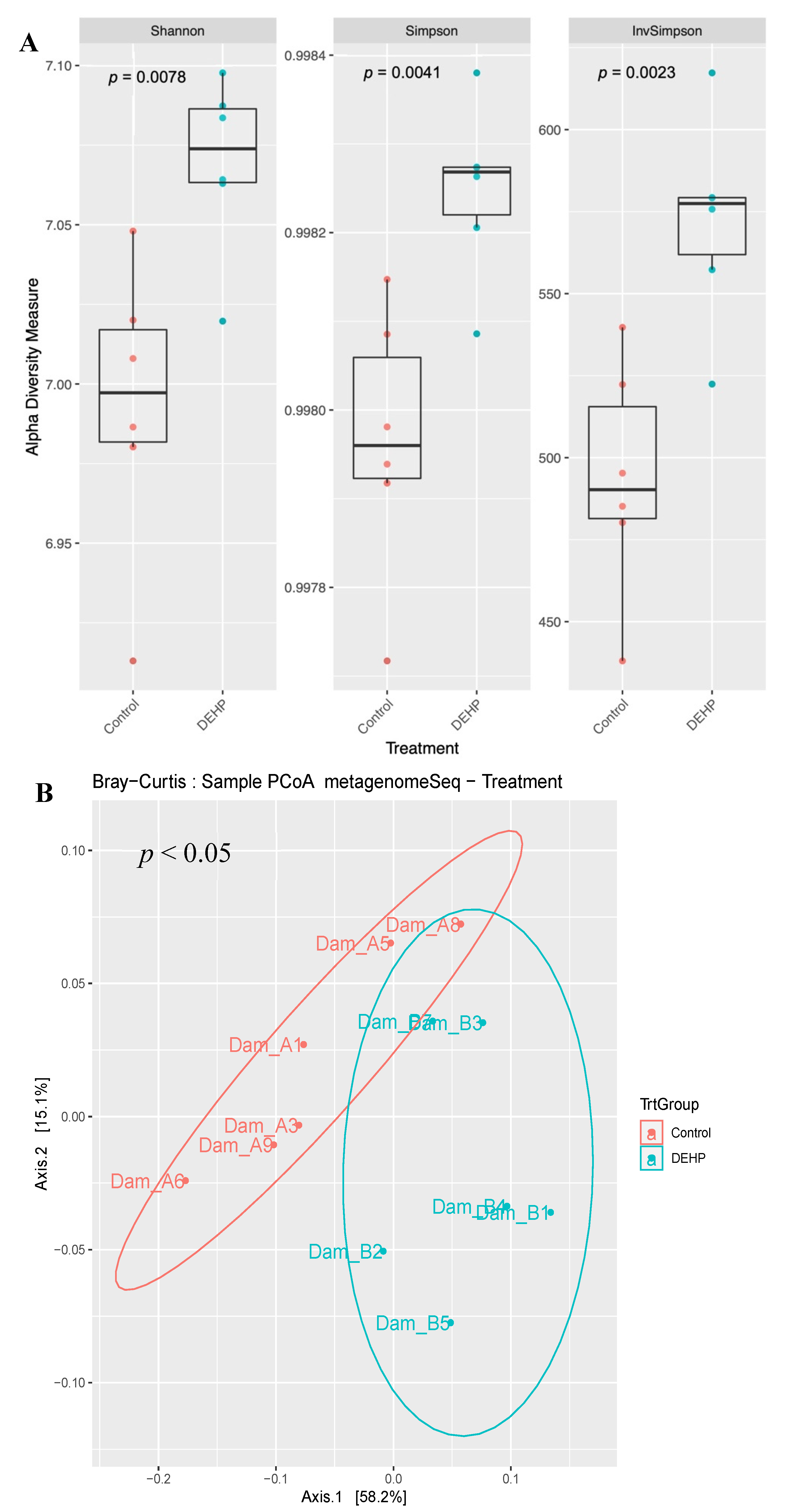
3.1.1. Alpha and Beta Diversity
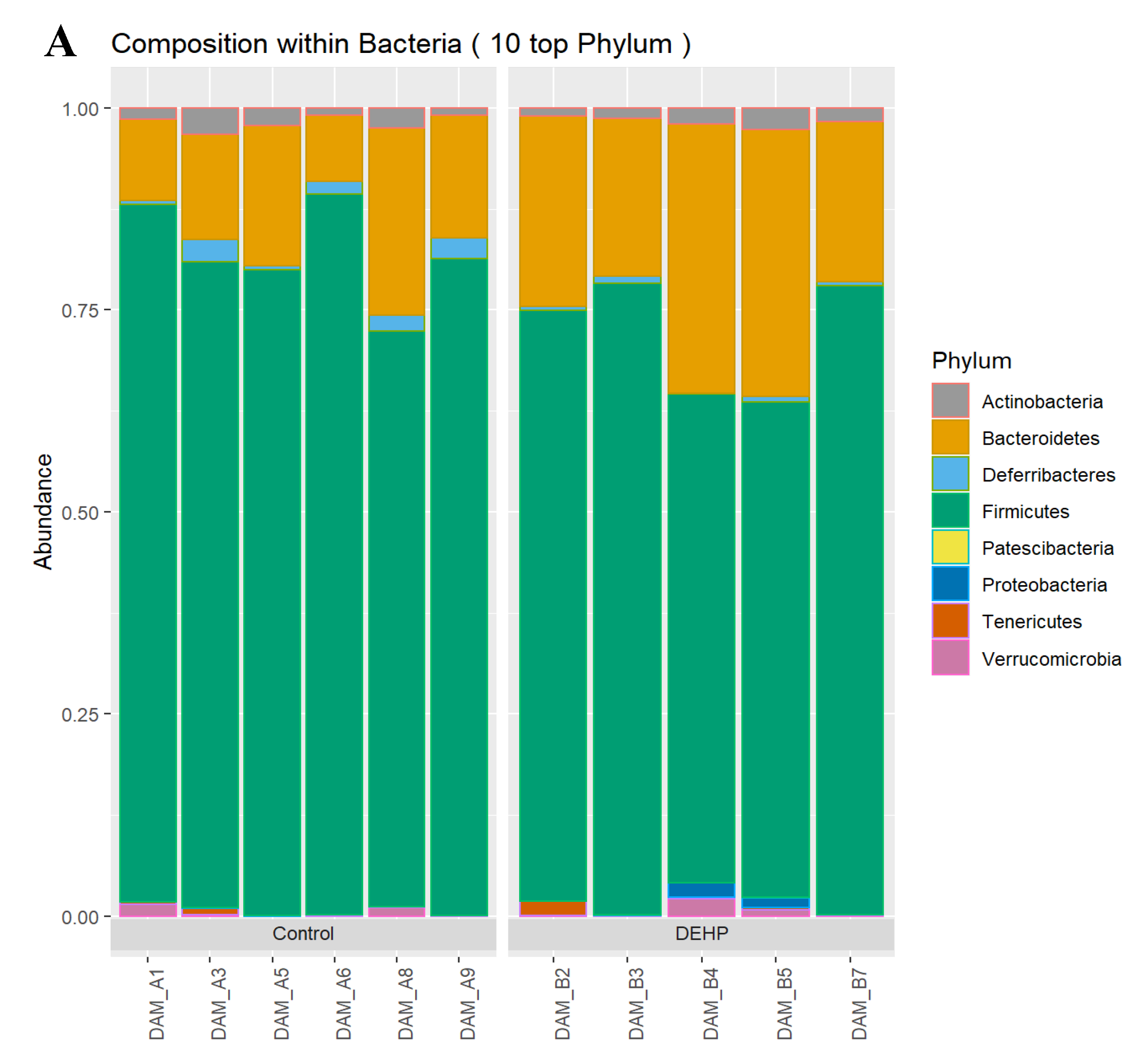
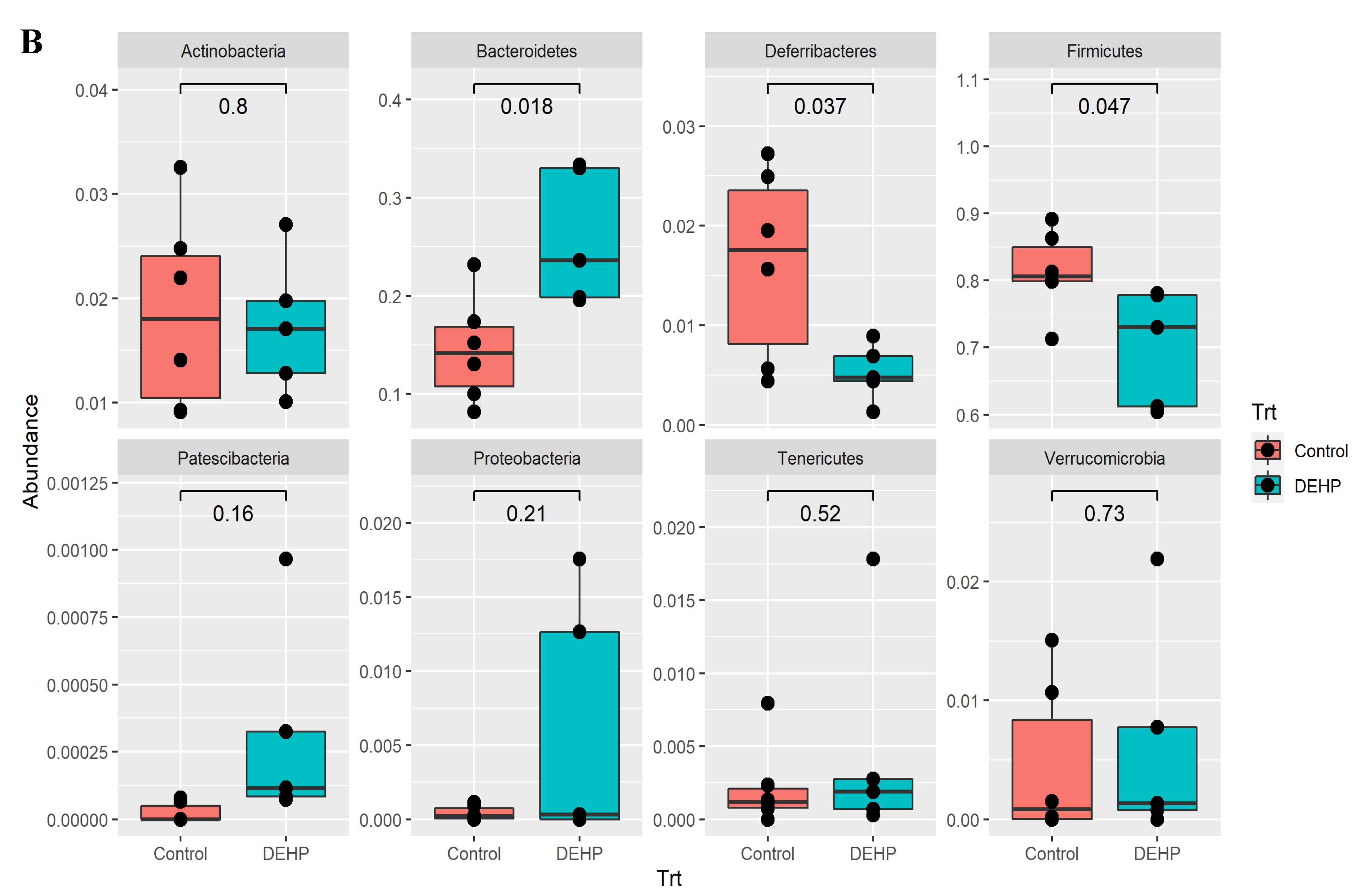
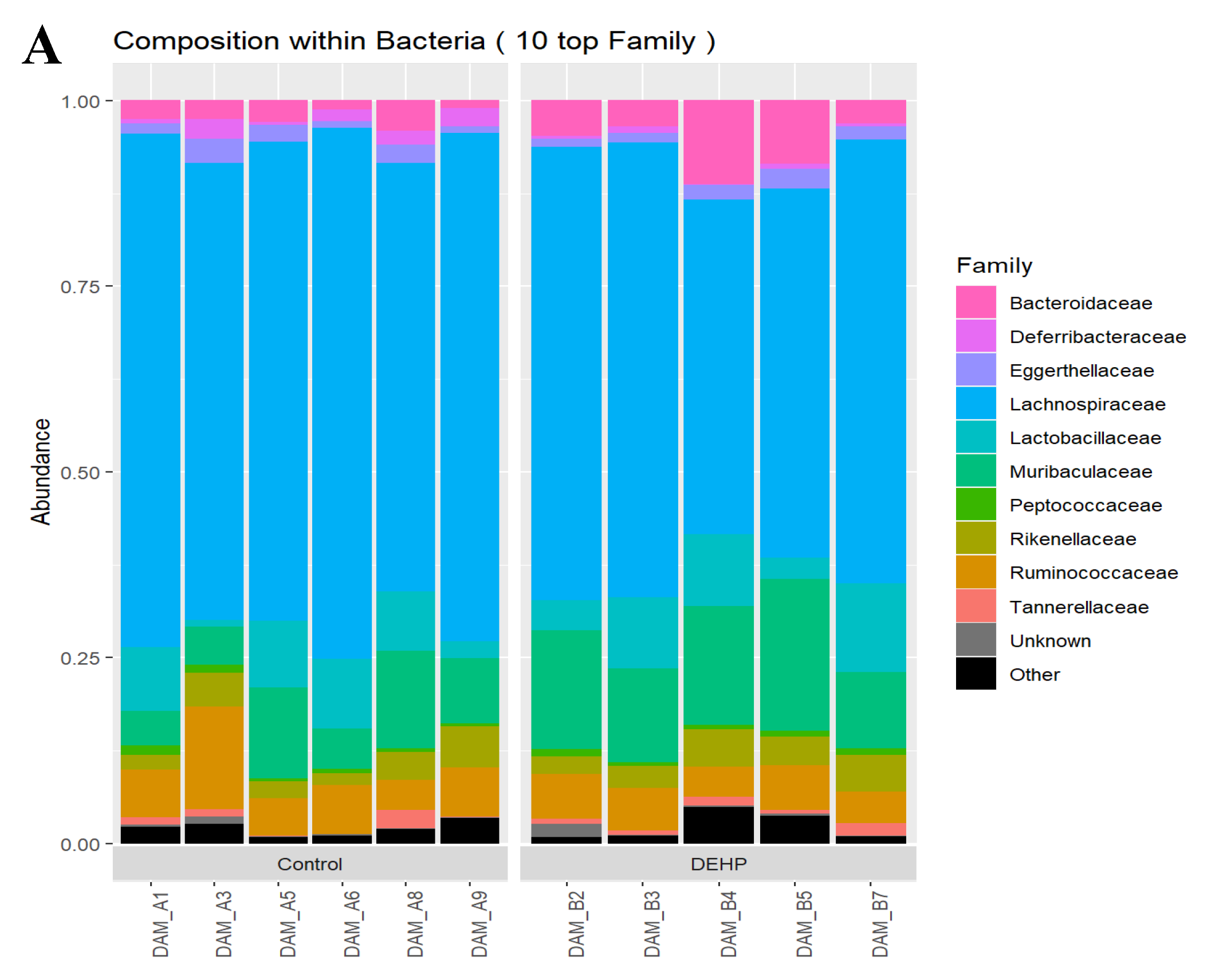
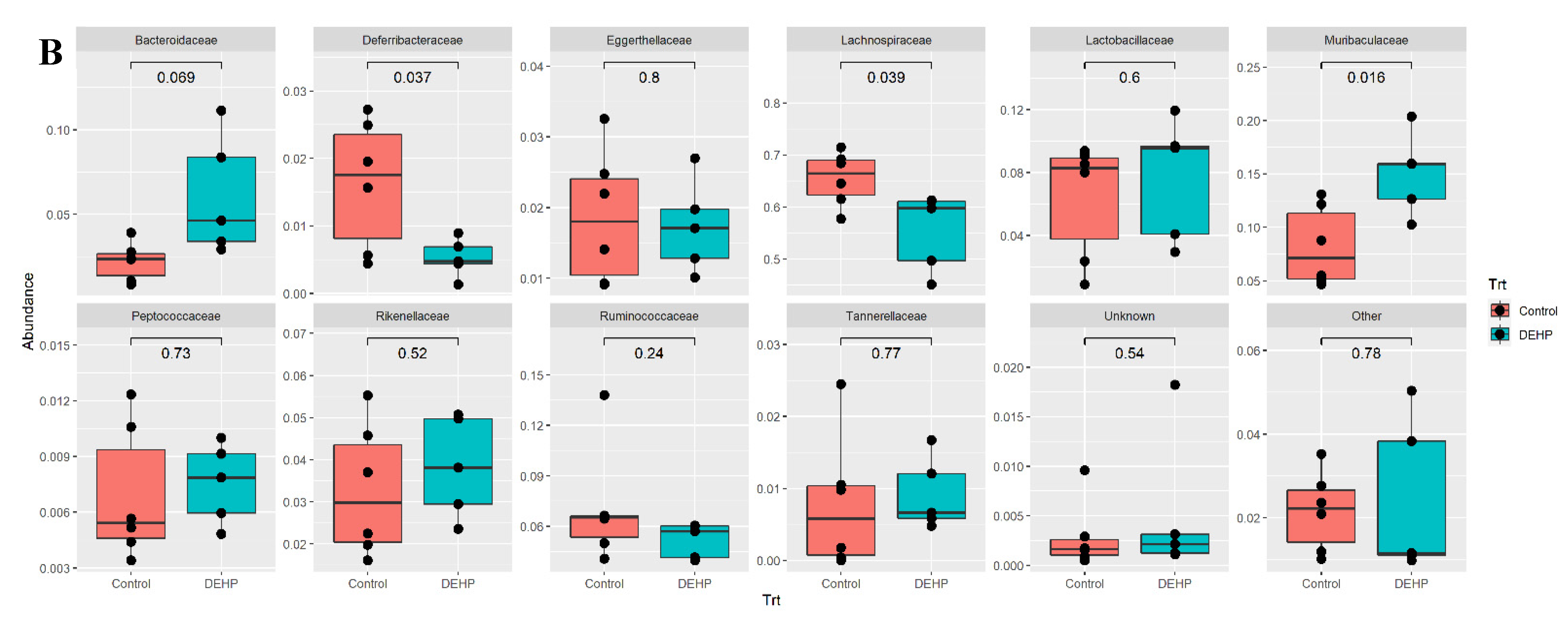
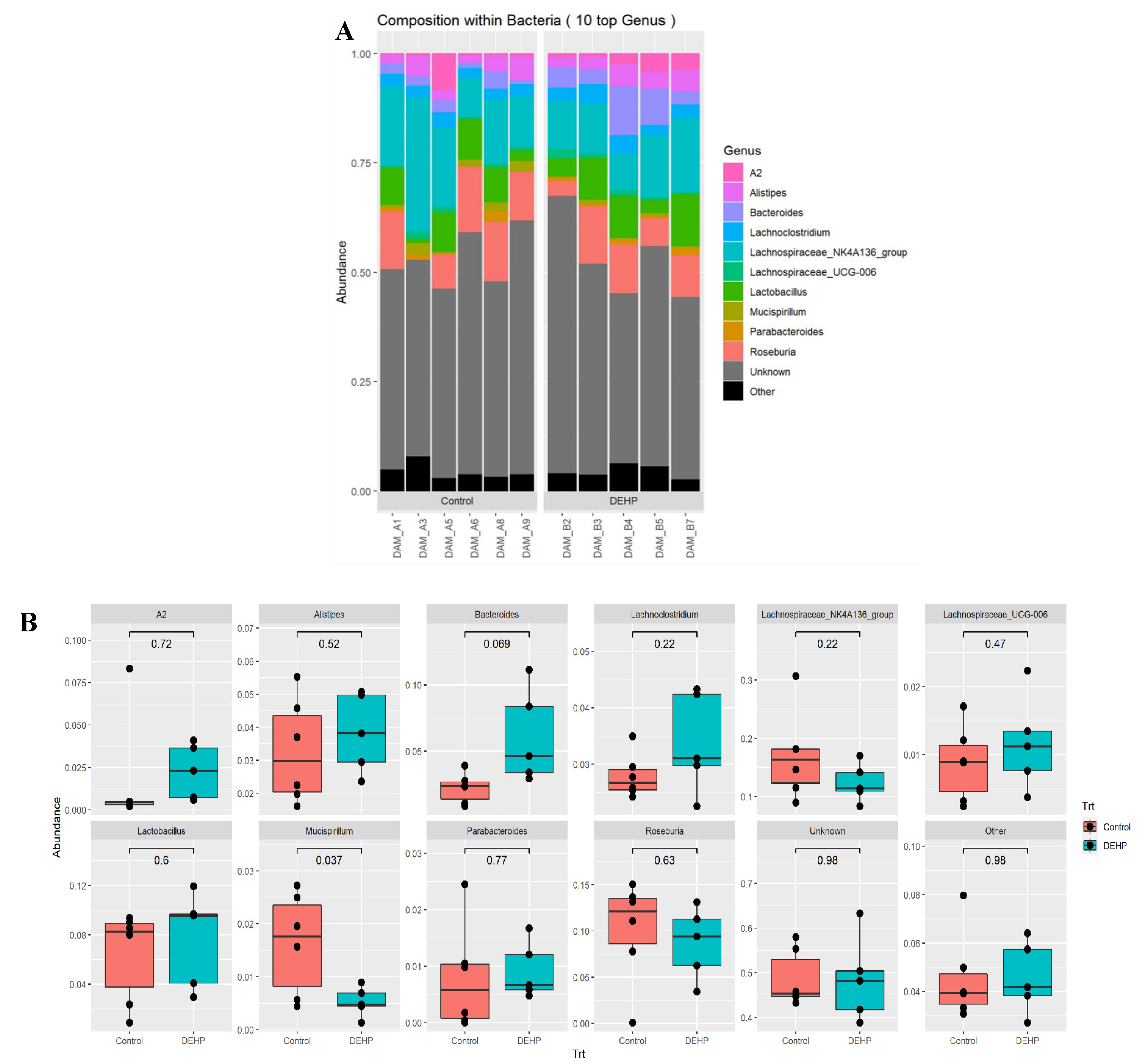
3.1.2. Taxonomic Results
3.2. Cecal Microbiome
3.2.1. Alpha and Beta Diversity
3.2.2. Results from Functional Analysis
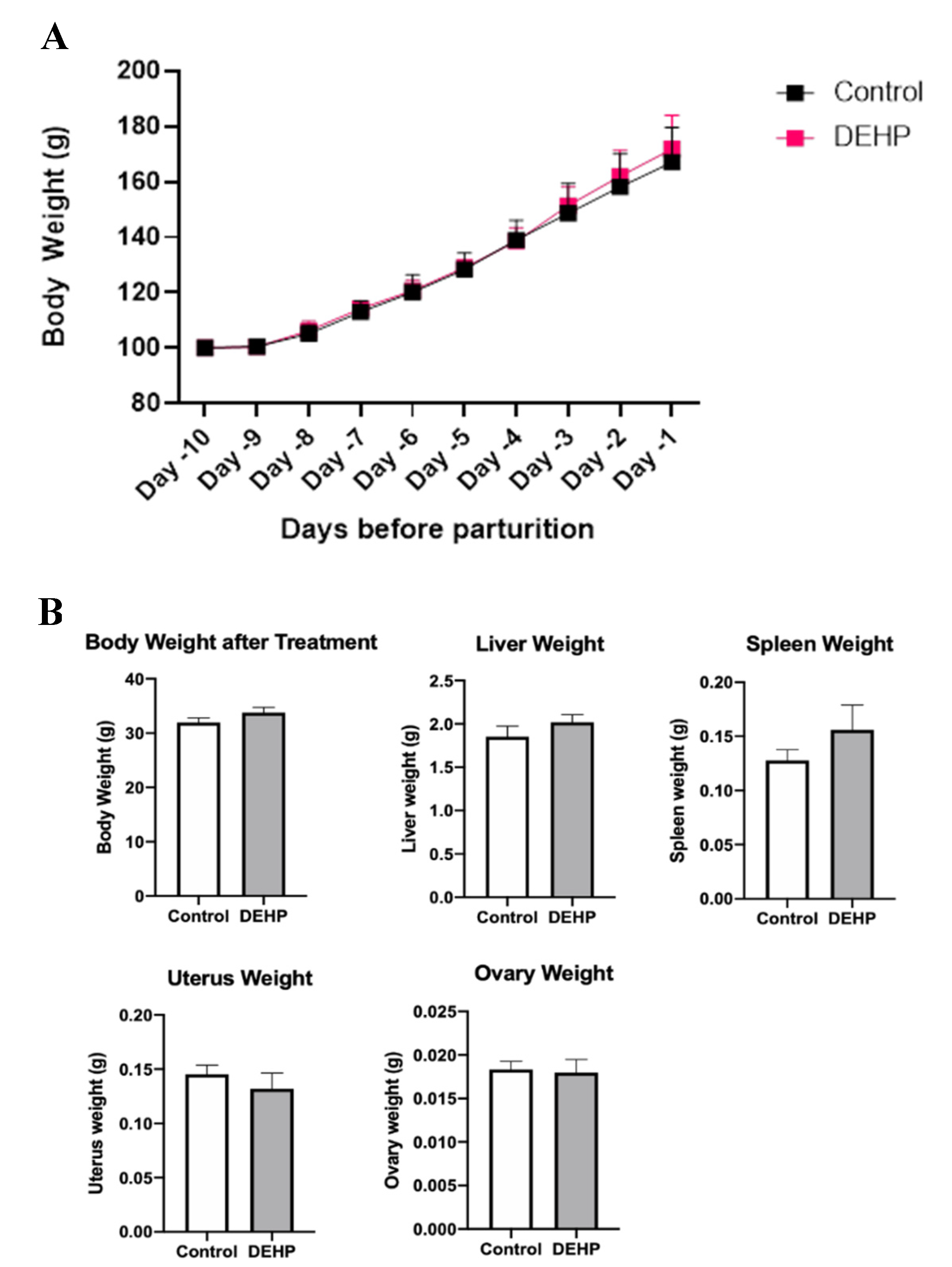
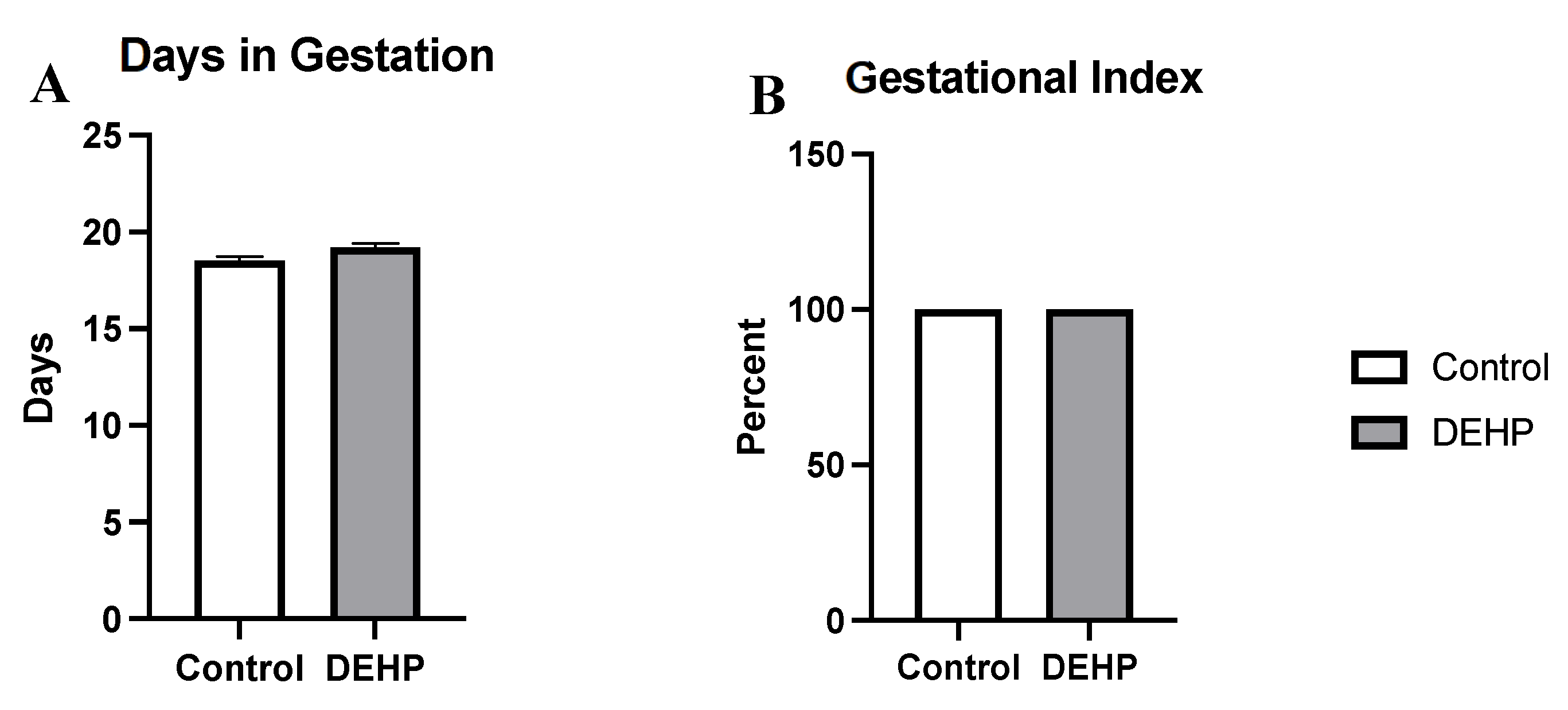
3.2.3. Gestational Outcomes and Weights in Dams
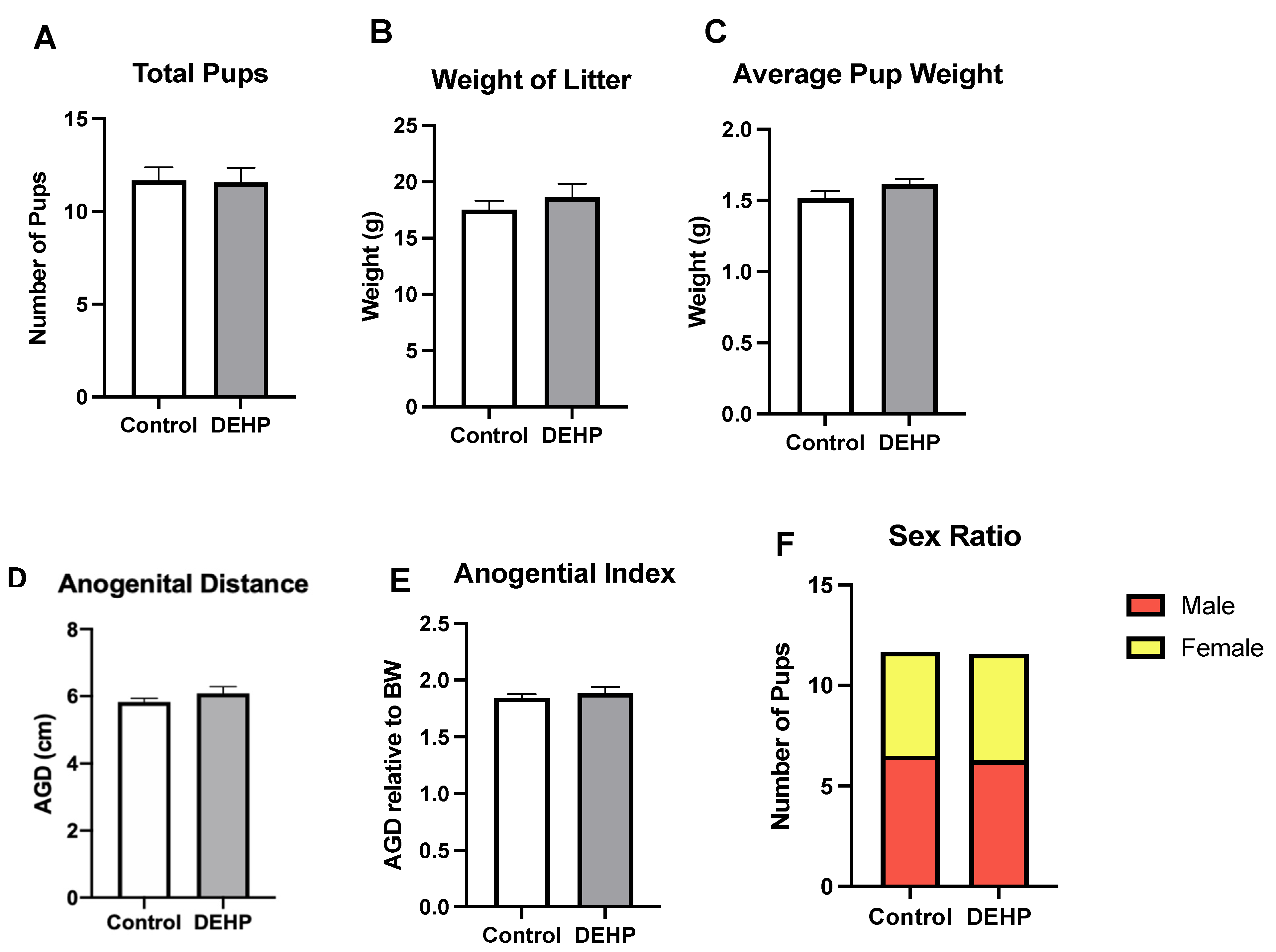
3.2.4. Outcomes in the F1 Pups
4. Discussion
5. Conclusions and Future Directions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Silva, M.J.; Barr, D.B.; Reidy, J.A.; Malek, N.A.; Hodge, C.C.; Caudill, S.P.; Brock, J.W.; Needham, L.L.; Calafat, A.M. Urinary levels of seven phthalate metabolites in the U.S. population from the National Health and Nutrition Examination Survey (NHANES) 1999–2000. Environ. Health Perspect. 2004, 112, 331–338. [Google Scholar] [CrossRef]
- Högberg, J.; Hanberg, A.; Berglund, M.; Skerfving, S.; Remberger, M.; Calafat, A.M.; Filipsson, A.F.; Jansson, B.; Johansson, N.; Appelgren, M.; et al. Phthalate Diesters and Their Metabolites in Human Breast Milk, Blood or Serum, and Urine as Biomarkers of Exposure in Vulnerable Populations. Environ. Health Perspect. 2008, 116, 334–339. [Google Scholar] [CrossRef] [Green Version]
- Marsee, K.; Woodruff, T.J.; Axelrad, D.A.; Calafat, A.M.; Swan, S.H. Estimated Daily Phthalate Exposures in a Population of Mothers of Male Infants Exhibiting Reduced Anogenital Distance. Environ. Health Perspect. 2006, 114, 805–809. [Google Scholar] [CrossRef] [PubMed]
- Sampson, J.; de Korte, D. DEHP-plasticised PVC: Relevance to blood services*. Transfus. Med. 2011, 21, 73–83. [Google Scholar] [CrossRef] [PubMed]
- Wahl, H.G.; Hong, Q.; Hildenbrand, S.; Risler, T.; Luft, D.; Liebich, H. 4-Heptanone is a metabolite of the plasticizer di(2-ethylhexyl) phthalate (DEHP) in haemodialysis patients. Nephrol. Dial. Transplant. 2004, 19, 2576–2583. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tsai, Y.-A.; Tsai, M.-S.; Hou, J.-W.; Lin, C.-L.; Chen, C.-Y.; Chang, C.-H.; Chen, M.-L. Evidence of high di(2-ethylhexyl) phthalate (DEHP) exposure due to tainted food intake in Taiwanese pregnant women and the health effects on birth outcomes. Sci. Total Environ. 2018, 618, 635–644. [Google Scholar] [CrossRef]
- Adibi, J.J.; Hauser, R.; Williams, P.L.; Whyatt, R.M.; Calafat, A.M.; Nelson, H.; Swan, S.H. Maternal Urinary Metabolites of Di-(2-Ethylhexyl) Phthalate in Relation to the Timing of Labor in a US Multicenter Pregnancy Cohort Study. Am. J. Epidemiol. 2009, 169, 1015–1024. [Google Scholar] [CrossRef]
- Caserta, D.; Pegoraro, S.; Mallozzi, M.; Di Benedetto, L.; Colicino, E.; Lionetto, L.; Simmaco, M. Maternal exposure to endocrine disruptors and placental transmission: A pilot study. Gynecol. Endocrinol. 2018, 34, 1001–1004. [Google Scholar] [CrossRef]
- Li, R.; Yu, C.; Gao, R.; Liu, X.; Lu, J.; Zhao, L.; He, J. Effects of DEHP on endometrial receptivity and embryo implantation in pregnant mice. J. Hazard. Mater. 2012, 241, 231–240. [Google Scholar] [CrossRef]
- Niermann, S.; Rattan, S.; Brehm, E.; Flaws, J.A. Prenatal exposure to di-(2-ethylhexyl) phthalate (DEHP) affects reproductive outcomes in female mice. Reprod. Toxicol. 2015, 53, 23–32. [Google Scholar] [CrossRef] [Green Version]
- Adamovsky, O.; Buerger, A.N.; Vespalcova, H.; Sohag, S.R.; Hanlon, A.T.; Ginn, P.E.; Craft, S.L.; Smatana, S.; Budinska, E.; Persico, M.; et al. Evaluation of Microbiome-Host Relationships in the Zebrafish Gastrointestinal System Reveals Adaptive Immunity Is a Target. of Bis(2-ethylhexyl) Phthalate (DEHP) Exposure. Environ. Sci. Technol. 2020, 54, 5719–5728. [Google Scholar] [CrossRef]
- Buerger, A.N.; Dillon, D.T.; Schmidt, J.; Yang, T.; Zubcevic, J.; Martyniuk, C.J.; Bisesi, J.H., Jr. Gastrointestinal dysbiosis following diethylhexyl phthalate exposure in zebrafish (Danio rerio): Altered microbial diversity, functionality, and network connectivity. Environ. Pollut. 2020, 265, 114496. [Google Scholar] [CrossRef]
- Edwards, S.M.; Cunningham, S.A.; Dunlop, A.L.; Corwin, E.J. The Maternal Gut Microbiome During Pregnancy. MCN Am. J. Matern. Child Nurs. 2017, 42, 310–317. [Google Scholar] [CrossRef] [PubMed]
- Fox, C.; Eichelberger, K. Maternal microbiome and pregnancy outcomes. Fertil. Steril. 2015, 104, 1358–1363. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, H.; Guo, R.; Li, S.; Liang, F.; Tian, C.; Zhao, X.; Long, Y.; Liu, F.; Jiang, M.; Zhang, Y.; et al. Systematic analysis of gut microbiota in pregnant women and its correlations with individual heterogeneity. NPJ Biofilms Microbiomes 2020, 6, 32. [Google Scholar] [CrossRef]
- Dieterich, W.; Schink, M.; Zopf, Y. Microbiota in the Gastrointestinal Tract. Med. Sci. 2018, 6, 116. [Google Scholar] [CrossRef] [Green Version]
- Shelby, M.D. NTP-CERHR monograph on the potential human reproductive and developmental effects of di (2-ethylhexyl) phthalate (DEHP). NTP CERHR Monogr. 2006, 18, v–vii. [Google Scholar]
- Tuddenham, S.; Sears, C.L. The intestinal microbiome and health. Curr. Opin. Infect. Dis. 2015, 28, 464–470. [Google Scholar] [CrossRef] [Green Version]
- Callahan, B.J.; McMurdie, P.J.; Rosen, M.J.; Han, A.W.; Johnson, A.J.; Holmes, S.P. DADA2: High-resolution sample inference from Illumina amplicon data. Nat. Methods 2016, 13, 581–583. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lan, Y.; Wang, Q.; Cole, J.R.; Rosen, G.L. Using the RDP classifier to predict taxonomic novelty and reduce the search space for finding novel organisms. PLoS ONE 2012, 7, e32491. [Google Scholar] [CrossRef] [Green Version]
- Quast, C.; Pruesse, E.; Yilmaz, P.; Gerken, J.; Schweer, T.; Yarza, P.; Peplies, J.; Glöckner, F.O. The SILVA ribosomal RNA gene database project: Improved data processing and web-based tools. Nucleic Acids Res. 2013, 41, D590–D596. [Google Scholar] [CrossRef] [PubMed]
- Price, M.N.; Dehal, P.S.; Arkin, A.P. FastTree 2--approximately maximum-likelihood trees for large alignments. PLoS ONE 2010, 5, e9490. [Google Scholar] [CrossRef]
- Kim, B.R.; Shin, J.; Guevarra, R.; Lee, J.H.; Kim, D.W.; Seol, K.H.; Isaacson, R. Deciphering Diversity Indices for a Better Understanding of Microbial Communities. J. Microbiol. Biotechnol. 2017, 27, 2089–2093. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fukuyama, J. Emphasis on the deep or shallow parts of the tree provides a new characterization of phylogenetic distances. Genome Biol. 2019, 20, 131. [Google Scholar] [CrossRef]
- Di Tommaso, P.; Chatzou, M.; Floden, E.W.; Barja, P.P.; Palumbo, E.; Notredame, C. Nextflow enables reproducible computational workflows. Nat. Biotechnol. 2017, 35, 316–319. [Google Scholar] [CrossRef] [PubMed]
- Andrews, S. FastQC: A Quality Control Tool for High Throughput Sequence Data. 2016. Available online: http://www.bioinformatics.babraham.ac.uk/projects/fastqc/ (accessed on 3 September 2021).
- Bolger, A.M.; Lohse, M.; Usadel, B. Trimmomatic: A flexible trimmer for Illumina sequence data. Bioinformatics 2014, 30, 2114–2120. [Google Scholar] [CrossRef] [Green Version]
- Rognes, T.; Flouri, T.; Nichols, B.; Quince, C.; Mahe, F. VSEARCH: A versatile open source tool for metagenomics. PeerJ 2016, 4, e2584. [Google Scholar] [CrossRef] [PubMed]
- The Huttenhower Lab. KneadData. 2018; 0.6.1. Available online: http://huttenhower.sph.harvard.edu/kneaddata (accessed on 3 September 2021).
- Buchfink, B.; Xie, C.; Huson, D.H. Fast and sensitive protein alignment using DIAMOND. Nat. Methods 2015, 12, 59–60. [Google Scholar] [CrossRef]
- Huson, D.H.; Beier, S.; Flade, I.; Górska, A.; El-Hadidi, M.; Mitra, S.; Ruscheweyh, H.-J.; Tappu, R. MEGAN Community Edition—Interactive Exploration and Analysis of Large-Scale Microbiome Sequencing Data. PLoS Comput. Biol. 2016, 12, e1004957. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2019. [Google Scholar]
- McMurdie, P.J.; Holmes, S. phyloseq: An R package for reproducible interactive analysis and graphics of microbiome census data. PLoS ONE 2013, 8, e61217. [Google Scholar] [CrossRef] [Green Version]
- Oksanen, J.; Blanchet, F.G.; Friendly, M.; Kindt, R.; Legendre, P.; McGlinn, D.; Wagner, H. Vegan: Community Ecology Package. 2017. Available online: https://CRAN.R-project.org/package=vegan (accessed on 3 September 2021).
- Paulson, J.N.; Stine, O.C.; Bravo, H.C.; Pop, M. Differential abundance analysis for microbial marker-gene surveys. Nat. Methods 2013, 10, 1200–1202. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Love, M.I.; Huber, W.; Anders, S. Moderated estimation of fold change and dispersion for RNA-seq data with DESeq2. Genome Biol. 2014, 15, 550. [Google Scholar] [CrossRef] [Green Version]
- Ritchie, M.E.; Phipson, B.; Wu, D.; Hu, Y.; Law, C.W.; Shi, W.; Smyth, G.K. Limma powers differential expression analyses for RNA-sequencing and microarray studies. Nucleic Acids Res. 2015, 43, e47. [Google Scholar] [CrossRef] [PubMed]
- Liaw, A.; Wiener, M. Classification and Regression by randomForest. R News 2002, 2, 18–22. [Google Scholar]
- Chiu, K.; Warner, G.; Nowak, R.A.; Flaws, J.A.; Mei, W. The Impact of Environmental Chemicals on the Gut Microbiome. Toxicol. Sci. 2020, 176, 253–284. [Google Scholar] [CrossRef]
- Ferguson, K.K.; McElrath, T.F.; Meeker, J.D. Environmental phthalate exposure and preterm birth. JAMA Pediatrics 2014, 168, 61–67. [Google Scholar] [CrossRef] [Green Version]
- Jurewicz, J.; Hanke, W. Exposure to phthalates: Reproductive outcome and children health. A review of epidemiological studies. Int. J. Occup. Med. Environ. Health 2011, 24, 115–141. [Google Scholar] [CrossRef]
- Lei, M.; Menon, R.; Manteiga, S.; Alden, N.; Hunt, C.; Alaniz, R.C.; Jayaraman, A. Environmental chemical diethylhexyl phthalate alters intestinal microbiota community structure and metabolite profile in mice. Msystems 2019, 4, e00724-19. [Google Scholar] [CrossRef] [Green Version]
- Wang, C.; Yue, S.; Hao, Z.; Ren, G.; Lu, D.; Zhang, Q.; Zhao, M. Pubertal exposure to the endocrine disruptor mono-2-ethylhexyl ester at body burden level caused cholesterol imbalance in mice. Environ. Pollut. 2019, 244, 657–666. [Google Scholar] [CrossRef]
- Hu, J.; Raikhel, V.; Gopalakrishnan, K.; Fernandez-Hernandez, H.; Lambertini, L.; Manservisi, F.; Chen, J. Effect of postnatal low-dose exposure to environmental chemicals on the gut microbiome in a rodent model. Microbiome 2016, 4, 26. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, Y.N.; Yang, Y.S.H.; Lin, I.H.; Chen, Y.Y.; Lin, H.Y.; Wu, C.Y.; Suen, J.L. Phthalate exposure alters gut microbiota composition and IgM vaccine response in human newborns. Food Chem. Toxicol. 2019, 132, 110700. [Google Scholar] [CrossRef] [PubMed]
- Wang, G.; Chen, Q.; Tian, P.; Wang, L.; Li, X.; Lee, Y.K.; Chen, W. Gut microbiota dysbiosis might be responsible to different toxicity caused by Di-(2-ethylhexyl) phthalate exposure in murine rodents. Environ. Pollut. 2020, 261, 114164. [Google Scholar] [CrossRef] [PubMed]
- Bondia-Pons, I.; Maukonen, J.; Mattila, I.; Rissanen, A.; Saarela, M.; Kaprio, J.; Orešič, M. Metabolome and fecal microbiota in monozygotic twin pairs discordant for weight: A Big Mac challenge. FASEB J. 2014, 28, 4169–4179. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hu, H.J.; Park, S.G.; Jang, H.B.; Choi, M.K.; Park, K.H.; Kang, J.H.; Cho, S.H. Obesity Alters the Microbial Community Profile in Korean Adolescents. PLoS ONE 2015, 10, e0134333. [Google Scholar] [CrossRef] [Green Version]
- Tian, X.; Yu, Z.; Feng, P.; Ye, Z.; Li, R.; Liu, J.; Hu, J.; Kakade, A.; Liu, P.; Li, X. Lactobacillus plantarum TW1-1 Alleviates Diethylhexylphthalate-Induced Testicular Damage in Mice by Modulating Gut Microbiota and Decreasing Inflammation. Front. Cell. Infect. Microbiol. 2019, 9, 221. [Google Scholar] [CrossRef] [Green Version]
- Loy, A.; Pfann, C.; Steinberger, M.; Hanson, B.; Herp, S.; Brugiroux, S.; Berry, D. Lifestyle and Horizontal Gene Transfer-Mediated Evolution of Mucispirillum schaedleri, a Core Member of the Murine Gut Microbiota. Msystems 2017, 2, e00171-16. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bunker, J.J.; Flynn, T.M.; Koval, J.C.; Shaw, D.G.; Meisel, M.; McDonald, B.D.; Ishizuka, I.E.; Dent, A.L.; Wilson, P.C.; Jabri, B.; et al. Innate and Adaptive Humoral Responses Coat. Distinct Commensal Bacteria with Immunoglobulin A. Immunity 2015, 43, 541–553. [Google Scholar] [CrossRef] [Green Version]
- Lagkouvardos, I.; Lesker, T.R.; Hitch, T.C.A.; Gálvez, E.J.C.; Smit, N.; Neuhaus, K.; Wang, J.; Baines, J.F.; Abt, B.; Stecher, B.; et al. Sequence and cultivation study of Muribaculaceae reveals novel species, host preference, and functional potential of this yet undescribed family. Microbiome 2019, 7, 28. [Google Scholar] [CrossRef] [Green Version]
- Pereira, F.C.; Wasmund, K.; Cobankovic, I.; Jehmlich, N.; Herbold, C.W.; Lee, K.S.; Sziranyi, B.; Vesely, C.; Decker, T.; Stocker, R.; et al. Rational design of a microbial consortium of mucosal sugar utilizers reduces Clostridiodes difficile colonization. Nat. Commun. 2020, 11, 5104. [Google Scholar] [CrossRef]
- Vacca, M.; Celano, G.; Calabrese, F.M.; Portincasa, P.; Gobbetti, M.; De Angelis, M. The Controversial Role of Human Gut Lachnospiraceae. Microorganisms 2020, 8, 573. [Google Scholar] [CrossRef]
- Hugenholtz, F.; de Vos, W.M. Mouse models for human intestinal microbiota research: A critical evaluation. Cell. Mol. Life Sci. CMLS 2018, 75, 149–160. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cullender, T.C.; Chassaing, B.; Janzon, A.; Kumar, K.; Muller, C.E.; Werner, J.J.; Angenent, L.T.; Bell, M.E.; Hay, A.G.; Peterson, D.A.; et al. Innate and adaptive immunity interact to quench microbiome flagellar motility in the gut. Cell Host Microbe 2013, 14, 571–581. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Parkunan, S.M.; Astley, R.; Callegan, M.C. Role of TLR5 and flagella in bacillus intraocular infection. PLoS ONE 2014, 9, e100543. [Google Scholar] [CrossRef] [PubMed]
- Chiu, K.; Bashir, S.T.; Nowak, R.A.; Mei, W.; Flaws, J.A. Subacute exposure to di-isononyl phthalate alters the morphology, endocrine function, and immune system in the colon of adult female mice. Sci. Rep. 2020, 10, 18788. [Google Scholar] [CrossRef] [PubMed]
- Williams, S.M.; Chen, Y.-T.; Andermann, T.M.; Carter, J.E.; McGee, D.J.; Ottemann, K.M. Helicobacter pylori chemotaxis modulates inflammation and bacterium-gastric epithelium interactions in infected mice. Infect. Immun. 2007, 75, 3747–3757. [Google Scholar] [CrossRef] [Green Version]
- Johnson, K.S.; Ottemann, K.M. Colonization, localization, and inflammation: The roles of H. pylori chemotaxis in vivo. Curr. Opin. Microbiol. 2018, 41, 51–57. [Google Scholar] [CrossRef]
- Tran, H.Q.; Ley, R.E.; Gewirtz, A.T.; Chassaing, B. Flagellin-elicited adaptive immunity suppresses flagellated microbiota and vaccinates against chronic inflammatory diseases. Nat. Commun. 2019, 10, 5650. [Google Scholar] [CrossRef] [Green Version]
- Simpson, B.W.; May, J.M.; Sherman, D.J.; Kahne, D.; Ruiz, N. Lipopolysaccharide transport to the cell surface: Biosynthesis and extraction from the inner membrane. Philos. Trans. R. Soc. Lond. Ser. B Biol. Sci. 2015, 370, 20150029. [Google Scholar] [CrossRef]
- Whitfield, C.; Trent, M.S. Biosynthesis and Export of Bacterial Lipopolysaccharides. Annu. Rev. Biochem. 2014, 83, 99–128. [Google Scholar] [CrossRef] [PubMed]
- Rosadini, C.V.; Kagan, J.C. Early innate immune responses to bacterial LPS. Curr. Opin. Immunol. 2017, 44, 14–19. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rattan, S.; Brehm, E.; Gao, L.; Flaws, J.A. Di(2-Ethylhexyl) Phthalate Exposure During Prenatal Development Causes Adverse Transgenerational Effects on Female Fertility in Mice. Toxicol. Sci. 2018, 163, 420–429. [Google Scholar] [CrossRef] [Green Version]
- Zhou, C.; Gao, L.; Flaws, J.A. Prenatal exposure to an environmentally relevant phthalate mixture disrupts reproduction in F1 female mice. Toxicol. Appl. Pharmacol. 2017, 318, 49–57. [Google Scholar] [CrossRef] [Green Version]
- Chiang, C.; Flaws, J.A. Subchronic Exposure to Di(2-ethylhexyl) Phthalate and Diisononyl Phthalate During Adulthood Has Immediate and Long-Term Reproductive Consequences in Female Mice. Toxicol. Sci. 2019, 168, 620–631. [Google Scholar] [CrossRef] [PubMed]
- Li, K.; Liszka, M.; Zhou, C.; Brehm, E.; Flaws, J.A.; Nowak, R.A. Prenatal exposure to a phthalate mixture leads to multigenerational and transgenerational effects on uterine morphology and function in mice. Reprod. Toxicol. 2020, 93, 178–190. [Google Scholar] [CrossRef] [PubMed]
- Latini, G.; De Felice, C.; Presta, G.; Del Vecchio, A.; Paris, I.; Ruggieri, F.; Mazzeo, P. In utero exposure to di-(2-ethylhexyl)phthalate and duration of human pregnancy. Environ. Health Perspect. 2003, 111, 1783–1785. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Polańska, K.; Ligocka, D.; Sobala, W.; Hanke, W. Effect of environmental phthalate exposure on pregnancy duration and birth outcomes. Int. J. Occup. Med. Environ. Health 2016, 29, 683–697. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Deng, T.; Du, Y.; Wang, Y.; Teng, X.; Hua, X.; Yuan, X.; Yao, Y.; Guo, N.; Li, Y. The associations of urinary phthalate metabolites with the intermediate and pregnancy outcomes of women receiving IVF/ICSI treatments: A prospective single-center study. Ecotoxicol. Environ. Saf. 2020, 188, 109884. [Google Scholar] [CrossRef] [PubMed]













{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Classification | Number of Taxa |
|---|---|
| Kingdom | 178 |
| Phylum | 178 |
| Class | 178 |
| Order | 178 |
| Family | 168 |
| Genus | 66 |
| Species | 4 |
| Variable | Bray–Curtis | UniFrac | Weighted UniFrac |
|---|---|---|---|
| Trt | 0.012 | 0.009 | 0.017 |
| Body weight | 0.452 | 0.433 | 0.082 |
| Ovary weight | 0.483 | 0.461 | 0.585 |
| Uterus weight | 0.649 | 0.057 | 0.872 |
| Liver weight | 0.291 | 0.673 | 0.046 |
| Spleen weight | 0.156 | 0.190 | 0.072 |
| Library size | 0.477 | 0.624 | 0.108 |
| Trt + Body Weight | NA | NA | 0.012 |
| Trt + Uterus Weight | NA | 0.007 | NA |
| Trt + Liver Weight | NA | NA | 0.008 |
| Trt + Spleen Weight | NA | NA | 0.016 |
| Pathway ID | Pathway | n | DE | P.DE |
|---|---|---|---|---|
| path:ko02040 | Flagellar assembly | 30 | 24 | 4.34 × 10−8 |
| path:ko02030 | Bacterial chemotaxis | 21 | 16 | 2.70 × 10−5 |
| path:ko03060 | Protein export | 18 | 12 | 2.02 × 10−3 |
| path:ko00190 | Oxidative phosphorylation | 45 | 23 | 3.94 × 10−3 |
| path:ko00720 | Carbon fixation pathways in prokaryotes | 53 | 26 | 4.60 × 10−3 |
| path:ko02024 | Quorum sensing | 86 | 38 | 6.89 × 10−3 |
| path:ko03440 | Homologous recombination | 21 | 12 | 1.19 × 10−2 |
| path:ko00340 | Histidine metabolism | 20 | 11 | 2.26 × 10−2 |
| path:ko01100 | Metabolic pathways | 898 | 304 | 2.45 × 10−2 |
| path:ko00130 | Ubiquinone and other terpenoid-quinone biosynthesis | 18 | 10 | 2.71 × 10−2 |
| path:ko00121 | Secondary bile acid biosynthesis | 3 | 3 | 3.02 × 10−2 |
| path:ko01502 | Vancomycin resistance | 16 | 9 | 3.25 × 10−2 |
| path:ko01200 | Carbon metabolism | 152 | 58 | 3.65 × 10−2 |
| path:ko00020 | Citrate cycle (TCA cycle) | 37 | 17 | 4.11 × 10−2 |
| path:ko00061 | Fatty acid biosynthesis | 17 | 9 | 5.04 × 10−2 |
| path:ko00540 | Lipopolysaccharide biosynthesis | 25 | 12 | 5.75 × 10−2 |
| path:ko03430 | Mismatch repair | 20 | 10 | 6.05 × 10−2 |
| path:ko00250 | Alanine, aspartate, and glutamate metabolism | 36 | 16 | 6.37 × 10−2 |
| path:ko01210 | 2-Oxocarboxylic acid metabolism | 28 | 13 | 6.43 × 10−2 |
| path:ko01110 | Biosynthesis of secondary metabolites | 350 | 122 | 6.64 × 10−2 |
| path:ko04142 | Lysosome | 8 | 5 | 6.73 × 10−2 |
| path:ko00780 | Biotin metabolism | 13 | 7 | 7.48 × 10−2 |
| path:ko00290 | Valine, leucine and isoleucine biosynthesis | 13 | 7 | 7.48 × 10−2 |
| path:ko00531 | Glycosaminoglycan degradation | 6 | 4 | 7.99 × 10−2 |
| path:ko00401 | Novobiocin biosynthesis | 6 | 4 | 7.99 × 10−2 |
| path:ko03018 | RNA degradation | 16 | 8 | 9.01 x 10-2 |
| path:ko00550 | Peptidoglycan biosynthesis | 32 | 14 | 9.06 × 10−2 |
| path:ko03013 | RNA transport | 4 | 3 | 9.27 × 10−2 |
| path:ko04214 | Apoptosis—fly | 2 | 2 | 9.71 × 10−2 |
| path:ko00332 | Carbapenem biosynthesis | 2 | 2 | 9.71 × 10−2 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chiu, K.; Bashir, S.T.; Gao, L.; Gutierrez, J.; de Godoy, M.R.C.; Drnevich, J.; Fields, C.J.; Cann, I.; Flaws, J.A.; Nowak, R.A. Subacute Exposure to an Environmentally Relevant Dose of Di-(2-ethylhexyl) Phthalate during Gestation Alters the Cecal Microbiome, but Not Pregnancy Outcomes in Mice. Toxics 2021, 9, 215. https://0-doi-org.brum.beds.ac.uk/10.3390/toxics9090215
Chiu K, Bashir ST, Gao L, Gutierrez J, de Godoy MRC, Drnevich J, Fields CJ, Cann I, Flaws JA, Nowak RA. Subacute Exposure to an Environmentally Relevant Dose of Di-(2-ethylhexyl) Phthalate during Gestation Alters the Cecal Microbiome, but Not Pregnancy Outcomes in Mice. Toxics. 2021; 9(9):215. https://0-doi-org.brum.beds.ac.uk/10.3390/toxics9090215
Chicago/Turabian StyleChiu, Karen, Shah Tauseef Bashir, Liying Gao, Jessica Gutierrez, Maria R. C. de Godoy, Jenny Drnevich, Christopher J. Fields, Isaac Cann, Jodi A. Flaws, and Romana A. Nowak. 2021. "Subacute Exposure to an Environmentally Relevant Dose of Di-(2-ethylhexyl) Phthalate during Gestation Alters the Cecal Microbiome, but Not Pregnancy Outcomes in Mice" Toxics 9, no. 9: 215. https://0-doi-org.brum.beds.ac.uk/10.3390/toxics9090215