A Three Monoclonal Antibody Combination Potently Neutralizes Multiple Botulinum Neurotoxin Serotype E Subtypes

 , ,
, ,
Abstract
:1. Introduction
2. Results
2.1. Characterization of Monoclonal Antibodies
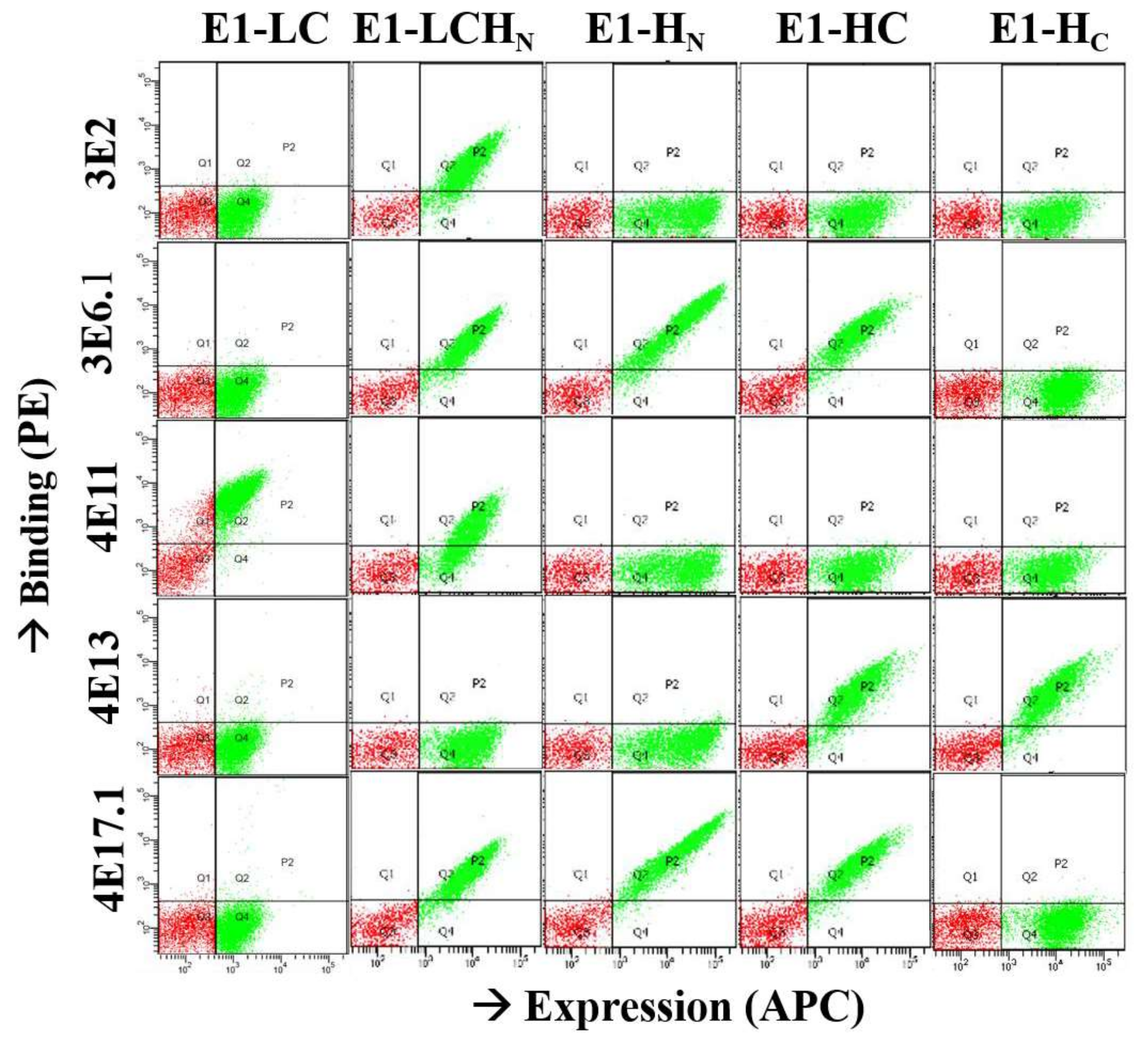
2.2. Epitope Mapping
2.3. Affinity Maturation of mAb 3E6
2.4. Mouse Neutralization Assays
3. Discussion
4. Conclusions
5. Materials and Methods
5.1. Ethics
5.2. Oligonucleotides for Library Construction
- 3E6.1mutH1rev GAC CCA GTG CAT AGC 521 542 542 511 532 GAA TCC AGA GGC TGC
- where:
- 1 = 70% A + 10% T + 10% G + 10% C
- 2 = 70% T + 10% A + 10% G + 10% C
- 3 = 70% G + 10% T + 10% A + 10% C
- 4 = 70% C + 10% T + 10% G + 10% A
- 5 = 50% G + 50% C
- PYDFor1: CCCCTCAACAACTAGCAAAGGCAGCCC
- PYDRev: GGTGATGGTGATGATGACCGGTACGCGTAG
- Link Rev: CGACCCGCCACCGCCAGAGCCACCTCCGCC
5.3. Strains, Media, Antibodies, and Toxin
5.4. Yeast-Displayed Human scFv Library Construction and Library Sorting
5.5. Measurement of KD Values of Yeast-Displayed scFv
5.6. Epitope Mapping
5.7. Identification of mAb Domain and Fine Epitopes
5.8. Site-Directed Mutagenesis
5.9. Affinity Maturation Library Construction and Sorting
5.10. Measurement of Solution Phase Affinity at Equilibrium
5.11. Measurement of In Vivo Toxin Neutralization
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Barash, J.R.; Arnon, S.S. A novel strain of Clostridium botulinum that produces type B and type H botulinum toxins. J. Infect. Dis. 2014, 209, 183–191. [Google Scholar] [CrossRef] [PubMed]
- Yao, G.; Lam, K.-H.; Perry, K.; Weisemann, J.; Rummel, A.; Jin, R. Crystal structure of the receptor-binding domain of botulinum neurotoxin type HA, also known as type FA or H. Toxins 2017, 9, 93. [Google Scholar] [CrossRef] [PubMed]
- Hatheway, C.L. Toxigenic clostridia. Clin. Microbiol. Rev. 1990, 3, 66–98. [Google Scholar] [CrossRef] [PubMed]
- Collins, M.D.; East, A.K. Phylogeny and taxonomy of the food-borne pathogen Clostridium botulinum and its neurotoxins. J. Appl. Microbiol. 1998, 84, 5–17. [Google Scholar] [CrossRef] [PubMed]
- Aureli, P.; Fenicia, L.; Pasolini, B.; Gianfranceschi, M.; McCroskey, L.M.; Hatheway, C.L. Two cases of type E infant botulism caused by neurotoxigenic Clostridium butyricum in Italy. J. Infect. Dis. 1986, 154, 207–211. [Google Scholar] [CrossRef] [PubMed]
- Suen, J.C.; Hatheway, C.L.; Steigerwalt, A.G.; Brenner, D.J. Genetic confirmation of identities of neurotoxigenic Clostridium baratii and Clostridium butyricum implicated as agents of infant botulism. J. Clin. Microbiol. 1988, 26, 2191–2192. [Google Scholar] [PubMed]
- Meng, X.; Karasawa, T.; Zou, K.; Kuang, X.; Wang, X.; Lu, C.; Wang, C.; Yamakawa, K.; Nakamura, S. Characterization of a neurotoxigenic Clostridium butyricum strain isolated from the food implicated in an outbreak of food-borne type E botulism. J. Clin. Microbiol. 1997, 35, 2160–2162. [Google Scholar] [PubMed]
- Wang, X.; Maegawa, T.; Karasawa, T.; Kozaki, S.; Tsukamoto, K.; Gyobu, Y.; Yamakawa, K.; Oguma, K.; Sakaguchi, Y.; Nakamura, S. Genetic analysis of type E botulinum toxin-producing Clostridium butyricum strains. App. Environ. Microbiol. 2000, 66, 4992–4997. [Google Scholar] [CrossRef]
- Hill, K.K.; Smith, T.J.; Helma, C.H.; Ticknor, L.O.; Foley, B.T.; Svensson, R.T.; Brown, J.L.; Johnson, E.A.; Smith, L.A.; Okinaka, R.T.; et al. Genetic diversity among Botulinum Neurotoxin-producing clostridial strains. J. Bacteriol. 2007, 189, 818–832. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.; Korkeala, H.; Aarnikunnas, J.; Lindström, M. Sequencing the botulinum neurotoxin gene and related genes in Clostridium botulinum type E strains reveals orfx3 and a novel type E neurotoxin subtype. J. Bacteriol. 2007, 189, 8643–8650. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Macdonald, T.E.; Helma, C.H.; Shou, Y.; Valdez, Y.E.; Ticknor, L.O.; Foley, B.T.; Davis, S.W.; Hannett, G.E.; Kelly-Cirino, C.D.; Barash, J.R.; et al. Analysis of Clostridium botulinum serotype E strains by using multilocus sequence typing, amplified fragment length polymorphism, variable-number tandem-repeat analysis, and botulinum neurotoxin gene sequencing. App. Environ. Microbiol. 2011, 77, 8625–8634. [Google Scholar] [CrossRef] [PubMed]
- Raphael, B.H.; Lautenschlager, M.; Kalb, S.R.; de Jong, L.I.; Frace, M.; Lúquez, C.; Barr, J.R.; Fernández, R.A.; Maslanka, S.E. Analysis of a unique Clostridium botulinum strain from the Southern hemisphere producing a novel type E botulinum neurotoxin subtype. BMC Microbiol. 2012, 12, 245. [Google Scholar] [CrossRef] [PubMed]
- Weedmark, K.A.; Lambert, D.L.; Mabon, P.; Hayden, K.L.; Urfano, C.J.; Leclair, D.; Van Domselaar, G.; Austin, J.W.; Corbett, C.R. Two novel toxin variants revealed by whole-genome sequencing of 175 Clostridium botulinum type E strains. App. Environ. Microbiol. 2014, 80, 6334–6345. [Google Scholar] [CrossRef] [PubMed]
- Mazuet, C.; Sautereau, J.; Legeay, C.; Bouchier, C.; Bouvet, P.; Popoff, M.R. An atypical outbreak of food-borne botulism due to Clostridium botulinum types B and E from ham. J. Clin. Microbiol. 2015, 53, 722–726. [Google Scholar] [CrossRef] [PubMed]
- Peck, M.W.; Smith, T.J.; Anniballi, F.; Austin, J.W.; Bano, L.; Bradshaw, M.; Cuervo, P.; Cheng, L.W.; Derman, Y.; Dorner, B.G. Historical perspectives and guidelines for botulinum neurotoxin subtype nomenclature. Toxins 2017, 9, 38. [Google Scholar] [CrossRef] [PubMed]
- Montecucco, C.; Schiavo, G. Tetanus and botulism neurotoxins: a new group of zinc proteases. Trends Biochem. Sci. 1993, 18, 324–327. [Google Scholar] [CrossRef]
- Fischer, A.; Garcia-Rodriguez, C.; Geren, I.; Lou, J.; Marks, J.D.; Nakagawa, T.; Montal, M. Molecular architecture of botulinum neurotoxin E revealed by single particle electron microscopy. J. Biol. Chem. 2008, 283, 3997–4003. [Google Scholar] [CrossRef] [PubMed]
- Kumaran, D.; Eswaramoorthy, S.; Furey, W.; Navaza, J.; Sax, M.; Swaminathan, S. Domain organization in Clostridium botulinum neurotoxin type E is unique: its implication in faster translocation. J. Mol. Biol. 2009, 386, 233–245. [Google Scholar] [CrossRef] [PubMed]
- Lacy, D.B.; Tepp, W.; Cohen, A.C.; DasGupta, B.R.; Stevens, R.C. Crystal structure of botulinum neurotoxin type A and implications for toxicity. Nat. Struct. Biol. 1998, 5, 898–902. [Google Scholar] [CrossRef] [PubMed]
- Das Gupta, B.R.; Sugiyama, H. Role of a protease in natural activation of Clostridium botulinum neurotoxin. Infect. Immun. 1972, 6, 587–590. [Google Scholar] [PubMed]
- Schiavo, G.; Rossetto, O.; Montecucco, C. Clostridial neurotoxins as tools to investigate the molecular events of neurotransmitter release. Semin. Cell. Biol. 1994, 5, 221–229. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Meng, J.; Lawrence, G.W.; Zurawski, T.H.; Sasse, A.; Bodeker, M.O.; Gilmore, M.A.; Fernandez-Salas, E.; Francis, J.; Steward, L.E.; et al. Novel chimeras of botulinum neurotoxins A and E unveil contributions from the binding, translocation, and protease domains to their functional characteristics. J. Biol. Chem. 2008, 283, 16993–17002. [Google Scholar] [CrossRef] [PubMed]
- Dolman, C.E. Type E botulism: a hazard of the north. Arctic 1960, 13, 230–256. [Google Scholar] [CrossRef]
- Hammer, T.H.; Jespersen, S.; Kanstrup, J.; Ballegaard, V.C.; Kjerulf, A.; Gelvan, A. Fatal outbreak of botulism in Greenland. Infect. Dis. 2015, 47, 190–194. [Google Scholar] [CrossRef] [PubMed]
- Artin, I.; Bjorkman, P.; Cronqvist, J.; Radstrom, P.; Holst, E. First case of type E wound botulism diagnosed using real-time PCR. J. Clin. Microbiol. 2007, 45, 3589–3594. [Google Scholar] [CrossRef] [PubMed]
- Fenicia, L.; Anniballi, F.; Aureli, P. Intestinal toxemia botulism in Italy, 1984-2005. Eur. J. Clin. Microbiol. Infect. Dis. 2007, 26, 385–394. [Google Scholar] [CrossRef] [PubMed]
- Cangene Corp., BAT® [Botulism Antitoxin Heptavalent (A, B, C, D, E, F, G)-(Equine)] Sterile Solution for Injection. Available online: https://www.fda.gov/downloads/.../UCM345147.pdf (accessed on 11 October 2017).
- Fagan, R.P.; Neil, K.P.; Sasich, R.; Luquez, C.; Asaad, H.; Maslanka, S.; Khalil, W. Initial recovery and rebound of type F intestinal colonization botulism after administration of investigational heptavalent botulinum antitoxin. Clin. Infect. Dis. Off. Pub. Infect. Dis. Soc. Am. 2011, 53, e125–e128. [Google Scholar] [CrossRef] [PubMed]
- Meng, Q.; Garcia-Rodriguez, C.; Manzanarez, G.; Silberg, M.; Conrad, F.; Bettencourt, J.; Pan, X.; Breece, T.; To, R.; Li, M. Engineered domain-based assays to identify individual antibodies in oligoclonal combinations targeting the same protein. Anal. Biochem. 2012, 430, 141–150. [Google Scholar] [CrossRef] [PubMed]
- Meng, Q.; Li, M.; Silberg, M.A.; Conrad, F.; Bettencourt, J.; To, R.; Huang, C.; Ma, J.; Meyer, K.; Shimizu, R.; et al. Domain-based assays of individual antibody concentrations in an oligoclonal combination targeting a single protein. Anal. Biochem. 2012, 421, 351–361. [Google Scholar] [CrossRef] [PubMed]
- Nowakowski, A.; Wang, C.; Powers, D.; Amersdorfer, P.; Smith, T.; Montgomery, V.; Sheridan, R.; Blake, R.; Smith, L.; Marks, J. Potent neutralization of botulinum neurotoxin by recombinant oligoclonal antibody. Proc. Natnl. Acad. Sci. USA 2002, 99, 11346–11350. [Google Scholar] [CrossRef] [PubMed]
- Nayak, S.U.; Griffiss, J.M.; McKenzie, R.; Fuchs, E.J.; Jurao, R.A.; An, A.T.; Ahene, A.; Tomic, M.; Hendrix, C.W.; Zenilman, J.M. Safety and pharmacokinetics of XOMA 3AB, a novel mixture of three monoclonal antibodies against botulinum toxin A. Antimicrob. Agents Chemother. 2014, 58, 5047–5053. [Google Scholar] [CrossRef] [PubMed]
- Garcia-Rodriguez, C.; Geren, I.N.; Lou, J.; Conrad, F.; Forsyth, C.; Wen, W.; Chakraborti, S.; Zao, H.; Manzanarez, G.; Smith, T.J.; et al. Neutralizing human monoclonal antibodies binding multiple serotypes of botulinum neurotoxin. Prot. Eng. Des. Sel. PEDS 2011, 24, 321–331. [Google Scholar] [CrossRef] [PubMed]
- Mahrhold, S.; Strotmeier, J.; Garcia-Rodriguez, C.; Lou, J.; Marks, J.D.; Rummel, A.; Binz, T. Identification of the SV2 protein receptor-binding site of botulinum neurotoxin type E. Biochem. J. 2013, 453, 37–47. [Google Scholar] [CrossRef] [PubMed]
- Fan, Y.; Dong, J.; Lou, J.; Wen, W.; Conrad, F.; Geren, I.N.; Garcia-Rodriguez, C.; Smith, T.J.; Smith, L.A.; Ho, M.; et al. Monoclonal antibodies that inhibit the proteolytic activity of botulinum neurotoxin serotype/B. Toxins 2015, 7, 3405–3423. [Google Scholar] [CrossRef] [PubMed]
- Fan, Y.; Geren, I.N.; Dong, J.; Lou, J.; Wen, W.; Conrad, F.; Smith, T.J.; Smith, L.A.; Ho, M.; Pires-Alves, M.; et al. Monoclonal antibodies targeting the alpha-exosite of botulinum neurotoxin serotype/A inhibit catalytic activity. PloS ONE 2015, 10, e0135306. [Google Scholar] [CrossRef] [PubMed]
- Levy, R.; Forsyth, C.; LaPorte, S.; Geren, I.; Smith, L.; Marks, J. Fine and domain-level epitope mapping of botulinum neurotoxin type A neutralizing antibodies by yeast surface display. J. Mol. Biol. 2007, 365, 196–210. [Google Scholar] [CrossRef] [PubMed]
- Swaminathan, S.; Eswaramoorthy, S. Structural analysis of the catalytic and binding sites of Clostridium botulinum neurotoxin B. Nat. Struct. Biol. 2000, 7, 693–699. [Google Scholar] [CrossRef] [PubMed]
- Schier, R.; Bye, J.; Apell, G.; McCall, A.; Adams, G.P.; Malmqvist, M.; Weiner, L.M.; Marks, J.D. Isolation of high-affinity monomeric human anti-c-erbB-2 single chain Fv using affinity-driven selection. J. Mol. Biol. 1996, 255, 28–43. [Google Scholar] [CrossRef] [PubMed]
- Razai, A.; Garcia-Rodriguez, C.; Lou, J.; Geren, I.N.; Forsyth, C.M.; Robles, Y.; Tsai, R.; Smith, T.J.; Smith, L.A.; Siegel, R.W.; et al. Molecular evolution of antibody affinity for sensitive detection of botulinum neurotoxin type A. J. Mol. Biol. 2005, 351, 158–169. [Google Scholar] [CrossRef] [PubMed]
- Smith, T.J.; Lou, J.; Geren, I.N.; Forsyth, C.M.; Tsai, R.; Laporte, S.L.; Tepp, W.H.; Bradshaw, M.; Johnson, E.A.; Smith, L.A.; et al. Sequence variation within botulinum neurotoxin serotypes impacts antibody binding and neutralization. Infect. Immun. 2005, 73, 5450–5457. [Google Scholar] [CrossRef] [PubMed]
- Adekar, S.P.; Takahashi, T.; Jones, R.M.; Al-Saleem, F.H.; Ancharski, D.M.; Root, M.J.; Kapadnis, B.P.; Simpson, L.L.; Dessain, S.K. Neutralization of botulinum neurotoxin by a human monoclonal antibody specific for the catalytic light chain. PloS ONE 2008, 3, e3023. [Google Scholar] [CrossRef] [PubMed]
- Takahashi, T.; Joshi, S.G.; Al-Saleem, F.; Ancharski, D.; Singh, A.; Nasser, Z.; Simpson, L.L. Localization of the sites and characterization of the mechanisms by which anti-light chain antibodies neutralize the actions of the botulinum holotoxin. Vaccine 2009, 27, 2616–2624. [Google Scholar] [CrossRef] [PubMed]
- Al-Saleem, F.H.; Nasser, Z.; Olson, R.M.; Cao, L.; Simpson, L.L. Identification of the factors that govern the ability of therapeutic antibodies to provide postchallenge protection against botulinum toxin: A model for assessing postchallenge efficacy of medical countermeasures against agents of bioterrorism and biological warfare. J. Pharmacol. Exp. Therap. 2011, 338, 503–517. [Google Scholar]
- Diamant, E.; Lachmi, B.-E.; Keren, A.; Barnea, A.; Marcus, H.; Cohen, S.; David, A.B.; Zichel, R. Evaluating the synergistic neutralizing effect of anti-botulinum oligoclonal antibody preparations. PLoS ONE 2014, 9, e87089. [Google Scholar] [CrossRef] [PubMed]
- Bakherad, H.; Gargari, S.L.M.; Rasooli, I.; RajabiBazl, M.; Mohammadi, M.; Ebrahimizadeh, W.; Ardakani, L.S.; Zare, H. In vivo neutralization of botulinum neurotoxins serotype E with heavy-chain camelid antibodies (VHH). Mol. Biotechnol. 2013, 55, 159–167. [Google Scholar] [CrossRef] [PubMed]
- Rasetti-Escargueil, C.; Avril, A.; Miethe, S.; Mazuet, C.; Derman, Y.; Selby, K.; Thullier, P.; Pelat, T.; Urbain, R.; Fontayne, A. The European AntibotABE Framework Program and Its Update: Development of Innovative Botulinum Antibodies. Toxins 2017, 9, 309. [Google Scholar] [CrossRef] [PubMed]
- Derman, Y.; Selby, K.; Miethe, S.; Frenzel, A.; Liu, Y.; Rasetti-Escargueil, C.; Avril, A.; Pelat, T.; Urbain, R.; Fontayne, A. Neutralization of botulinum neurotoxin type E by a humanized antibody. Toxins 2016, 8, 257. [Google Scholar] [CrossRef] [PubMed]
- Marks, J.D.; Hoogenboom, H.R.; Bonnert, T.P.; McCafferty, J.; Griffiths, A.D.; Winter, G. By-passing immunization. Human antibodies from V-gene libraries displayed on phage. J. Mol. Biol. 1991, 222, 581–597. [Google Scholar] [CrossRef]
- Dong, J.; Thompson, A.A.; Fan, Y.; Lou, J.; Conrad, F.; Ho, M.; Pires-Alves, M.; Wilson, B.A.; Stevens, R.C.; Marks, J.D. A single-domain llama antibody potently inhibits the enzymatic activity of botulinum neurotoxin by binding to the non-catalytic alpha-exosite binding region. J. Mol. Biol. 2010, 397, 1106–1118. [Google Scholar] [CrossRef] [PubMed]
- Sheets, M.D.; Amersdorfer, P.; Finnern, R.; Sargent, P.; Lindqvist, E.; Schier, R.; Hemingsen, G.; Wong, C.; Gerhart, J.C.; Marks, J.D. Efficient construction of a large nonimmune phage antibody library: the production of high-affinity human single-chain antibodies to protein antigens. Proc. Natl. Acad. Sci. USA 1998, 95, 6157–6162. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| mAb Name | Domain Bound | KD (×10−9 M) | ||||
|---|---|---|---|---|---|---|
| BoNT/E1, scFv | BoNTE2, scFv | BoNT/E3, scFv | BoNT/E4, scFv | BoNT/E3, IgG | ||
| 3E1 | LC | NB | NB | 0.70 | NB | ND |
| 3E2 | LC-HN | 0.62 | 1.16 | 0.15 | 0.16 | 0.002 |
| 3E3 | LC | NB | NB | 0.6 | NB | ND |
| 3E4 | LC-HN | 4.7 | >100 | 4.7 | NB | 0.70 |
| 3E5 | LC | 24.10 | 1.5 | 4.90 | NB | 1.4 |
| 3E6 | HN | 15.40 | >100 | 15.4 | >100 | ND |
| 3E6.1 | HN | 0.83 | 0.57 | 0.28 | >10.0 | 0.04 |
| 3E6.2 | HN | 0.55 | 0.64 | 0.17 | 1.42 | 0.009 |
| 4E11 | LC | 12.67 | 12.39 | 14.20 | NB | ND |
| 4E13 | HC | 4.38 | NB | 9.90 | NB | 7.24 |
| 4E16 | LC | 2.22 | 1.43 | 0.20 | NB | 0.15 |
| 4E17 | HN | 18.37 | >100 | 14.09 | >100 | ND |
| 4E17.1 | HN | 0.16 | 1.27 | 0.1 | 0.60 | 0.24 |
| Mean Fluorescence Intensity (MFI) of IgG Binding to Yeast scFv-Captured BoNT/E3 | |||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| Capture mAb | 3E1 | 3E2 | 3E3 | 3E4 | 3E5 | 3E6 | 4E11 | 4E13 | 4E16 | 4E17.1 | No IgG |
| 3E1 | 196 | 2916 | 283 | 134 | 6486 | 1558 | 5357 | 509 | 1919 | 3742 | 98 |
| 3E2 | 2492 | 276 | 5503 | 10,394 | 8503 | 6729 | 10,425 | 5843 | 11,564 | 6870 | 89 |
| 3E3 | 143 | 1255 | 252 | 138 | 1966 | 482 | 2712 | 188 | 681 | 646 | 105 |
| 3E4.1 | 509 | 17,993 | 359 | 236 | 12,747 | 14,858 | 301 | 11,952 | 10,433 | 3262 | 104 |
| 3E5 | 2059 | 6026 | 3175 | 3910 | 103 | 1455 | 100 | 1460 | 104 | 1429 | 103 |
| 3E6.1 | 6795 | 18,622 | 13,081 | 17,313 | 7654 | 230 | 9309 | 10,982 | 10,185 | 4270 | 105 |
| 4E11 | 844 | 4807 | 802 | 237 | 139 | 2346 | 241 | 2201 | 210 | 5708 | 95 |
| 4E13 | 739 | 3646 | 1141 | 1334 | 1306 | 1472 | 1176 | 238 | 1976 | 3602 | 99 |
| 4E16 | 2093 | 7133 | 5165 | 9466 | 232 | 4641 | 101 | 7109 | 163 | 4659 | 85 |
| 4E17.1 | 8975 | 9039 | 13,085 | 12,820 | 7247 | 14,713 | 9442 | 6911 | 9667 | 128 | 93 |
| mAb | KD (×10−12 M) | kon (106 M−1 s−1) | koff (10−6 s−1) | |||
|---|---|---|---|---|---|---|
| BoNT/E1 | BoNT/E2 | BoNT/E3 | BoNT/E4 | BoNT/E3 | BoNT/E3 | |
| 3E2 | 11.52 | 15.34 | 2.28 | 17.53 | 1.080 | 2.461 |
| 3E6.1 | 35.11 | 50.93 | 40.32 | 9720 | 0.5272 | 21.26 |
| 3E6.2 | 6.41 | 7.07 | 8.55 | 542 | 2.136 | 18.27 |
| 4E16 | 312.13 | 383.12 | 153.64 | NB | 0.4444 | 68.28 |
| 4E17.1 | 730.30 | 204.74 | 239.58 | 492.04 | 0.7968 | 190.90 |
| Number of Mice Surviving/10 Mice Treated | ||||
|---|---|---|---|---|
| Antibody Dose | 50 μg | 10 μg | 5 μg | 2 μg |
| Antibody combination | BoNT/E1: 40,000 LD50 | |||
| 3E2, 3E6.1, 4E16 | 9/10 | 9/10 | 9/10 | 0/10 |
| 3E2, 3E6.1, 4E17.1 | 10/10 | 10/10 | 0/10 | |
| BoNT/E3: 40,000 LD50 | ||||
| 3E2, 3E6.1, 4E16 | 9/10 | 9/10 | 1/10 | |
| 3E2, 3E6.1, 4E17.1 | 9/10 | 10/10 | 0/10 | |
| 3E2, 3E6.2, 4E17.1 | 9/10 | 10/10 | 0/10 | |
| BoNT/E4: 40,000 LD50 | ||||
| 3E2, 3E6.1, 4E16 | 0/10 | |||
| BoNT/E4: 10,000 LD50 | ||||
| 3E2, 3E6.1, 4E17.1 | 10/10 | 1/10 | ||
| 3E2, 3E6.2, 4E17.1 | 8/10 | 5/10 | ||
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Garcia-Rodriguez, C.; Razai, A.; Geren, I.N.; Lou, J.; Conrad, F.; Wen, W.-H.; Farr-Jones, S.; Smith, T.J.; Brown, J.L.; Skerry, J.C.; et al. A Three Monoclonal Antibody Combination Potently Neutralizes Multiple Botulinum Neurotoxin Serotype E Subtypes. Toxins 2018, 10, 105. https://0-doi-org.brum.beds.ac.uk/10.3390/toxins10030105
Garcia-Rodriguez C, Razai A, Geren IN, Lou J, Conrad F, Wen W-H, Farr-Jones S, Smith TJ, Brown JL, Skerry JC, et al. A Three Monoclonal Antibody Combination Potently Neutralizes Multiple Botulinum Neurotoxin Serotype E Subtypes. Toxins. 2018; 10(3):105. https://0-doi-org.brum.beds.ac.uk/10.3390/toxins10030105
Chicago/Turabian StyleGarcia-Rodriguez, Consuelo, Ali Razai, Isin N. Geren, Jianlong Lou, Fraser Conrad, Wei-Hua Wen, Shauna Farr-Jones, Theresa J. Smith, Jennifer L. Brown, Janet C. Skerry, and et al. 2018. "A Three Monoclonal Antibody Combination Potently Neutralizes Multiple Botulinum Neurotoxin Serotype E Subtypes" Toxins 10, no. 3: 105. https://0-doi-org.brum.beds.ac.uk/10.3390/toxins10030105