Repeated Intrastriatal Botulinum Neurotoxin-A Injection in Hemiparkinsonian Rats Increased the Beneficial Effect on Rotational Behavior



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
2.1. Repetitive Intrastriatal BoNT-A Injection in Hemi-PD Rats
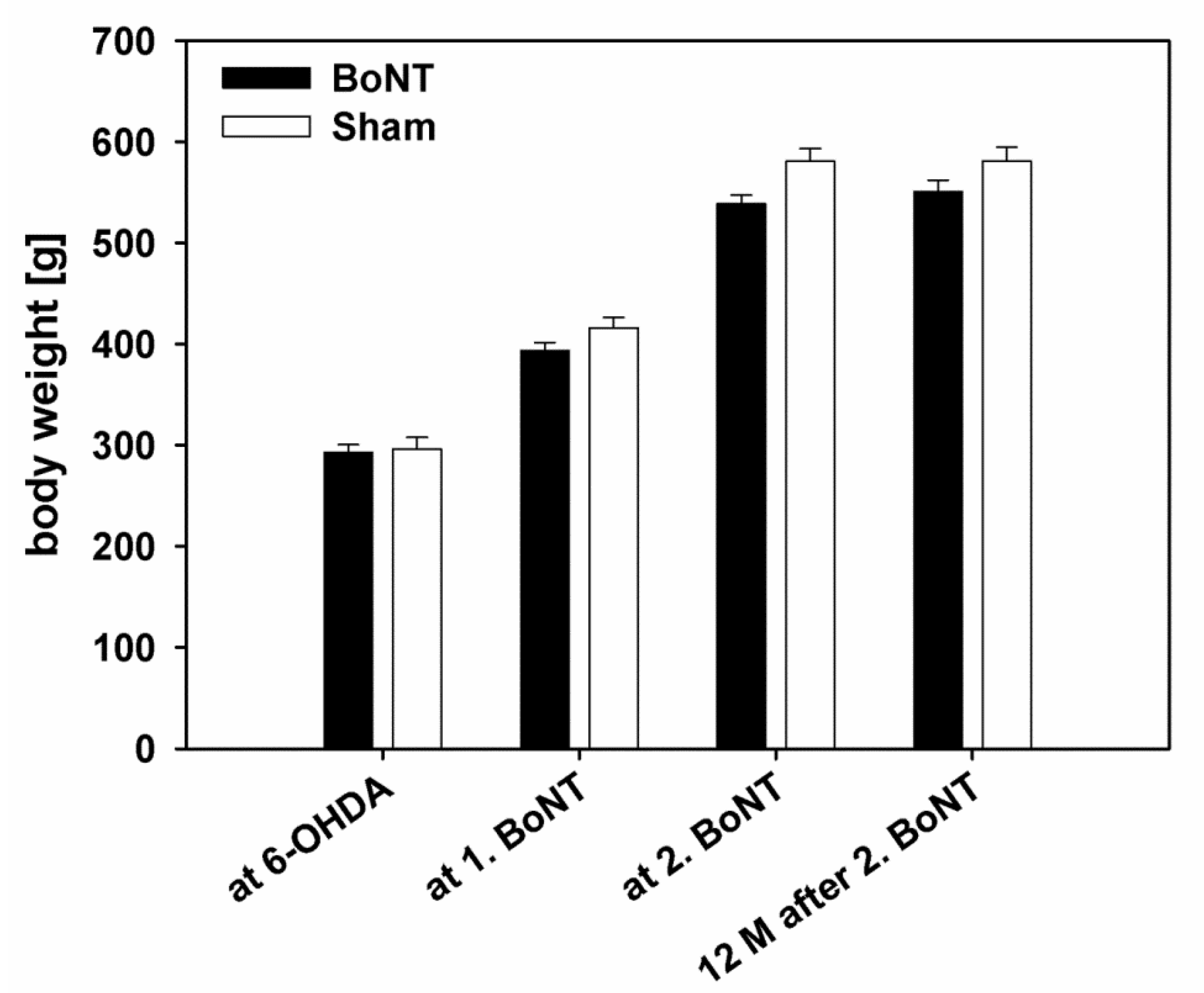
2.1.1. Body Weight
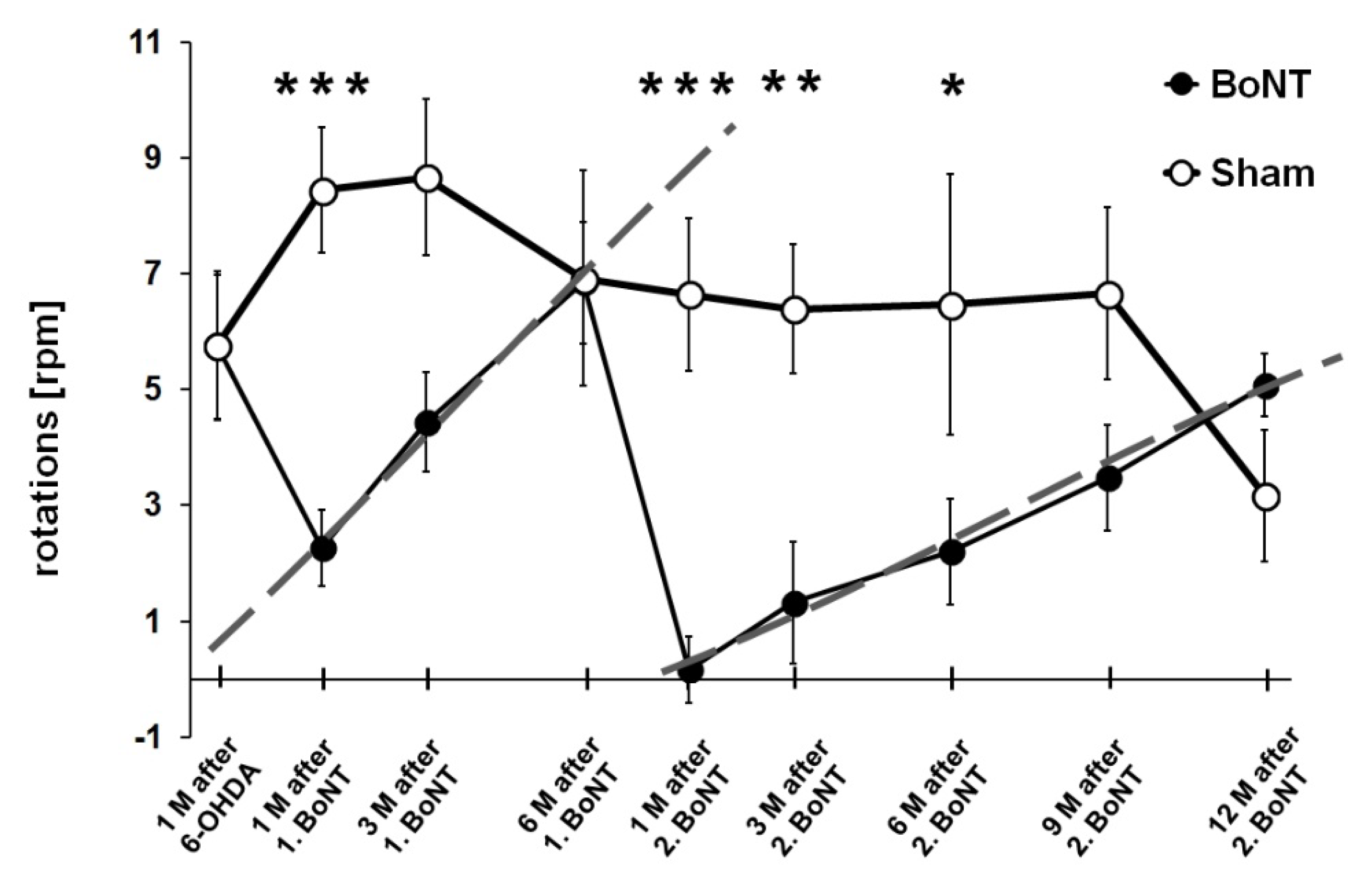
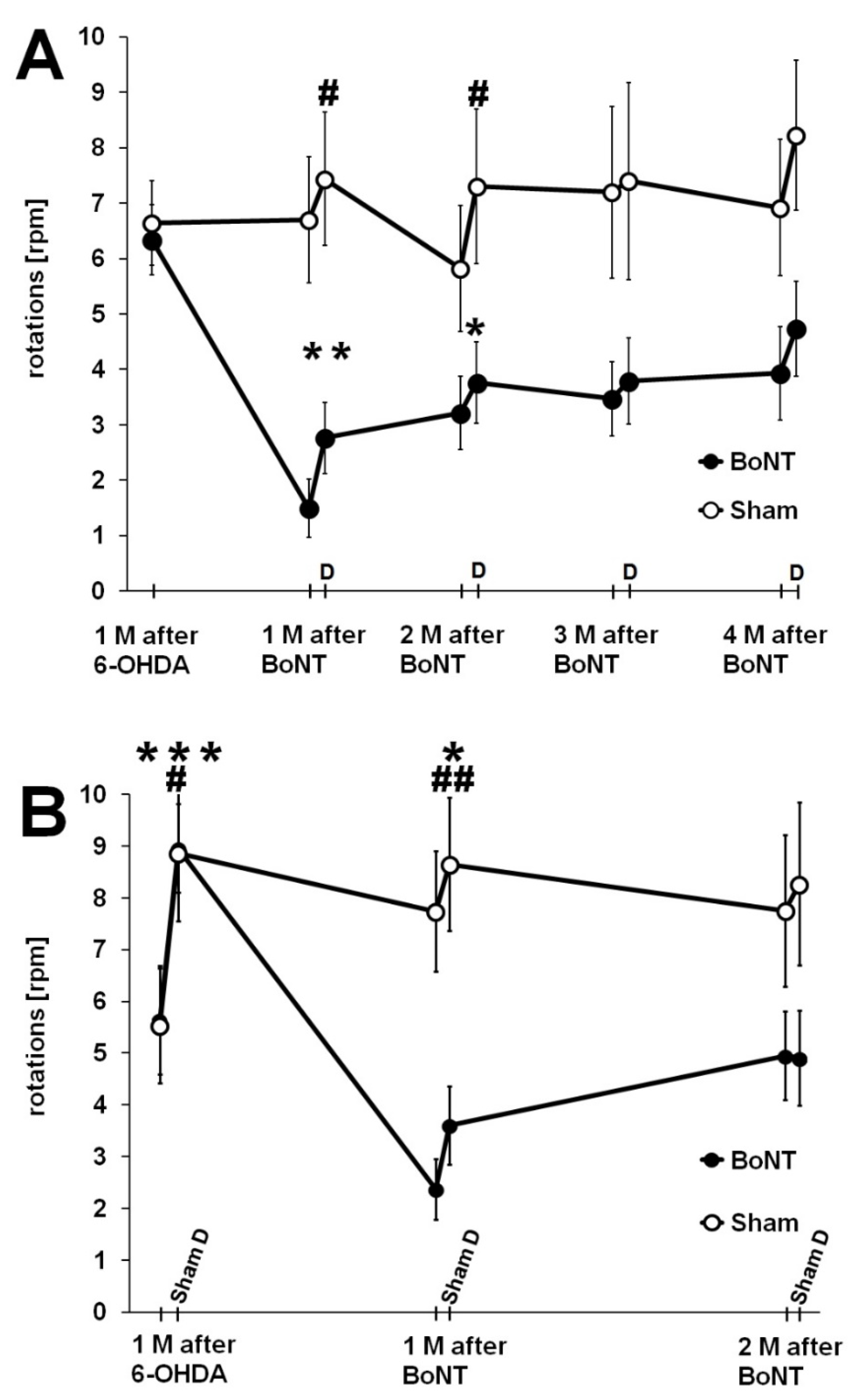
2.1.2. Apomorphine-Induced Rotation Test
2.1.3. Spontaneous Motor Tests
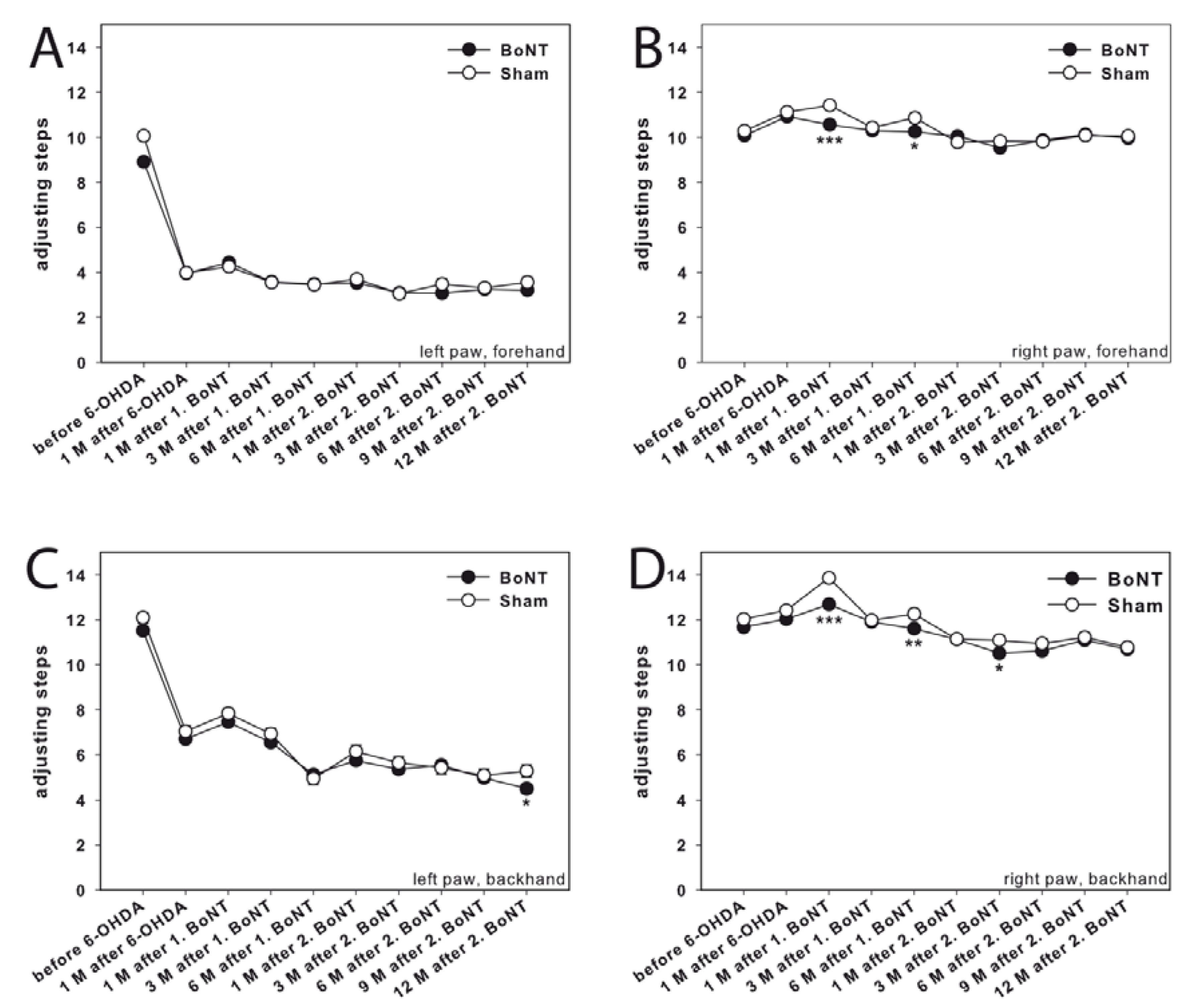
Adjusting Steps
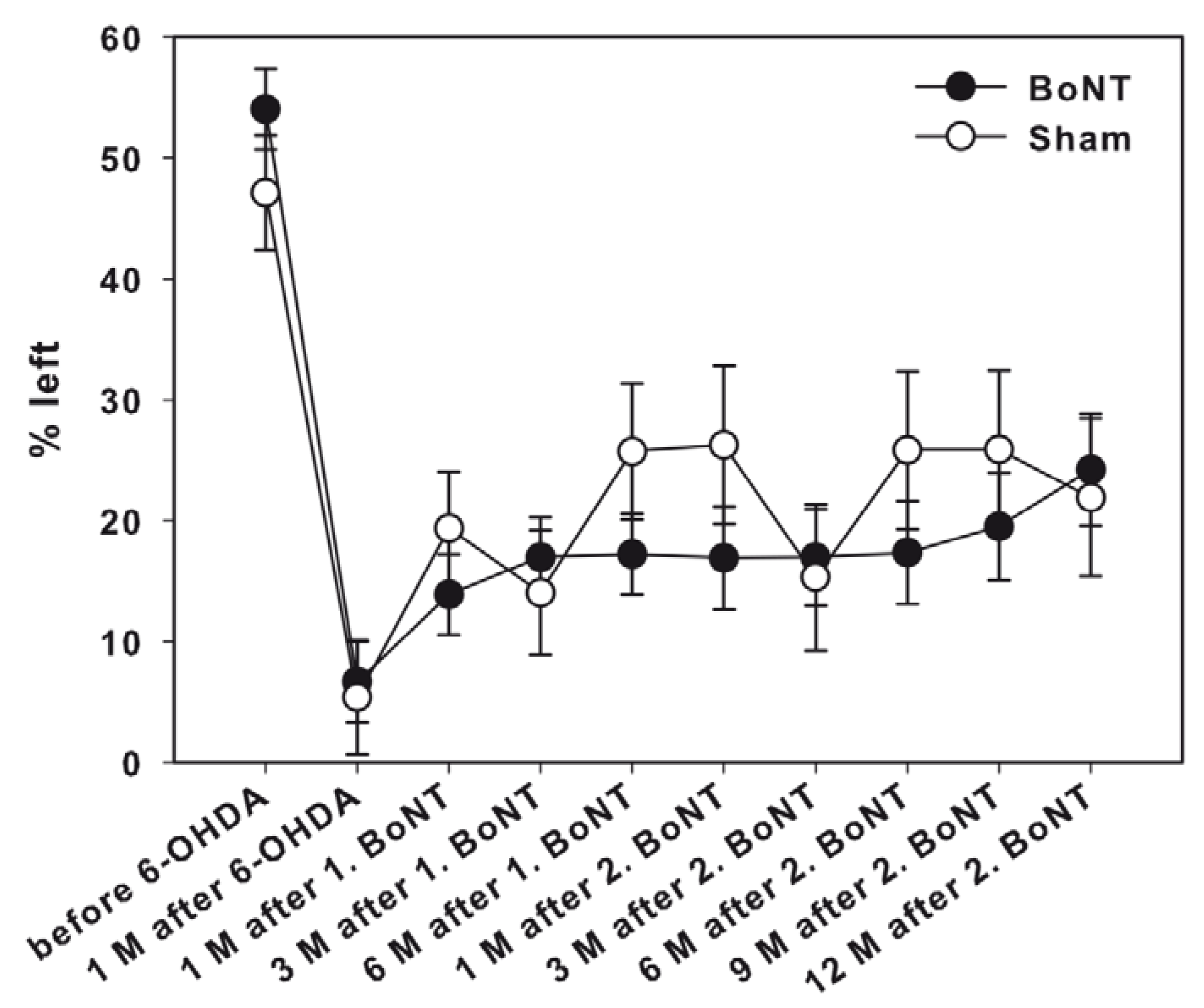
Corridor Task
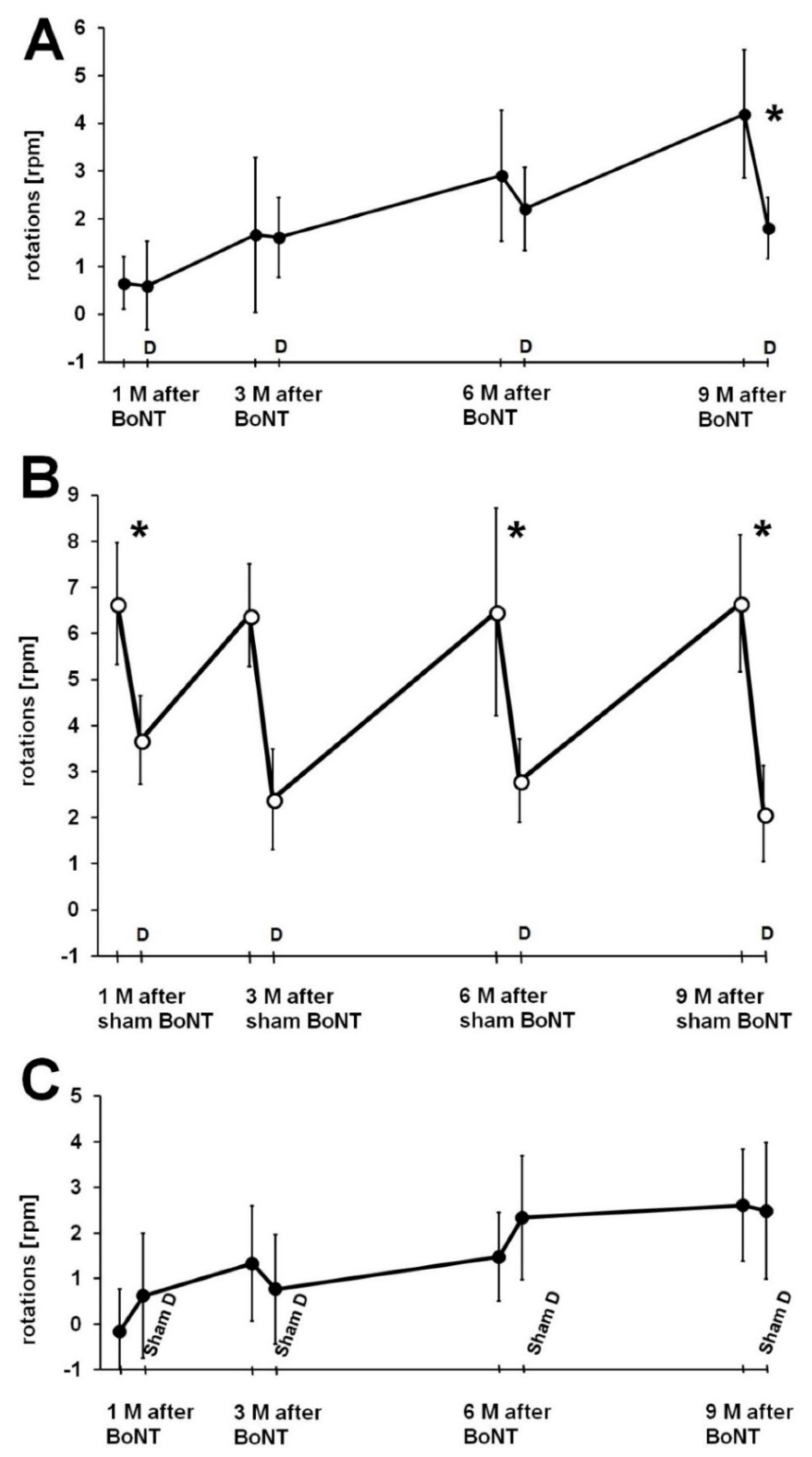
2.2. Influence of Donepezil on Apomorphine-Induced Rotation Behavior
2.2.1. Reaction of Rats to Donepezil
2.2.2. Donepezil Injection 24 h Prior to the Second Rotation Test
2.2.3. Sham Donepezil Injection 24 h Prior to the Second Rotation Test
2.2.4. Donepezil Injection 1 h Prior to the Second Rotation Test
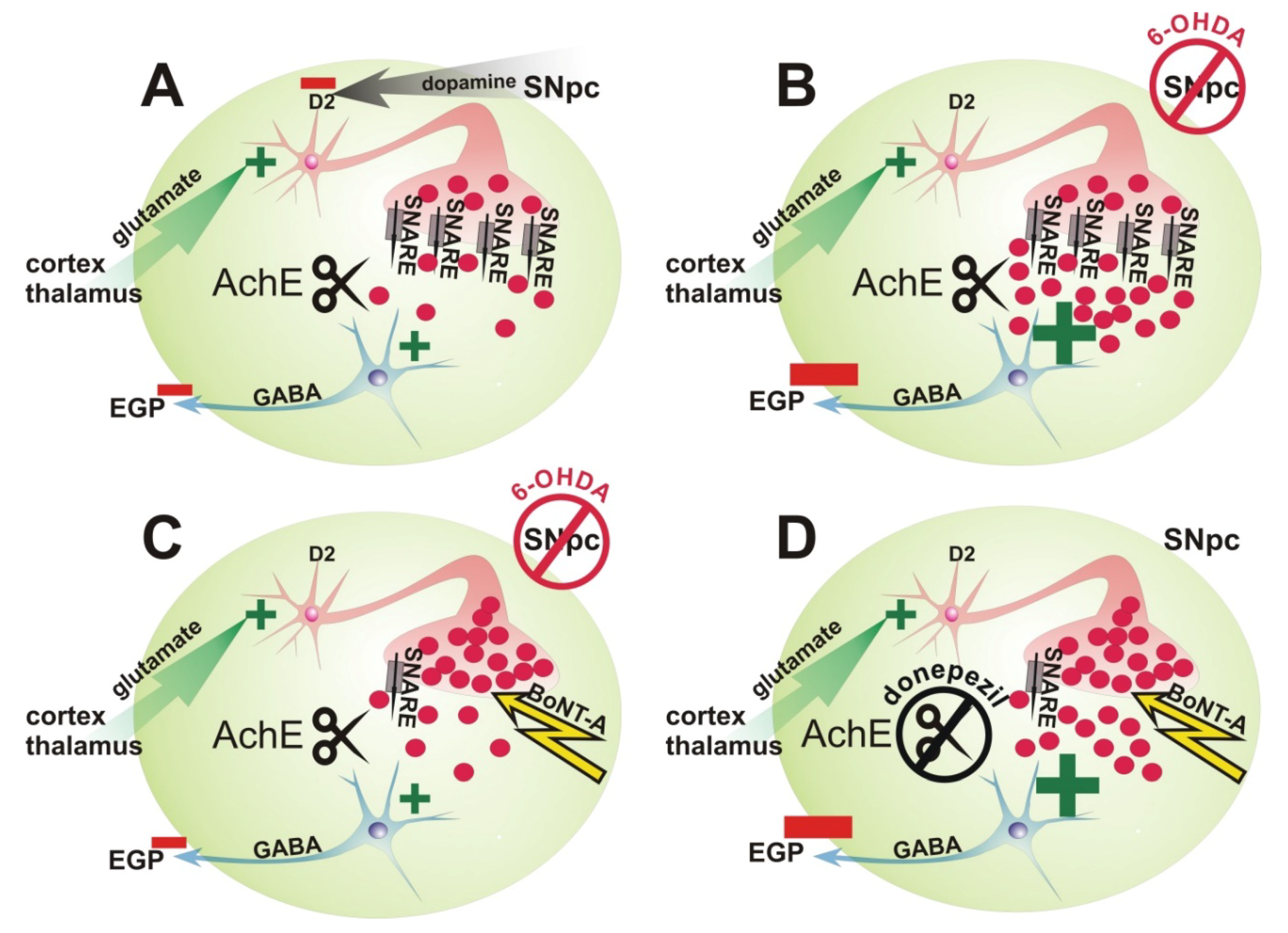
3. Discussion
3.1. Repetitive Intrastriatal BoNT-A Injection in Hemi-PD Rats
3.1.1. Body Weight
3.1.2. Apomorphine-Induced Rotation Test
3.1.3. Spontaneous Motor Tests
3.2. Influence of Application of 2 mg kg−1 BW Donepezil Prior to Apomorphine-Induced Rotation Test
4. Conclusions
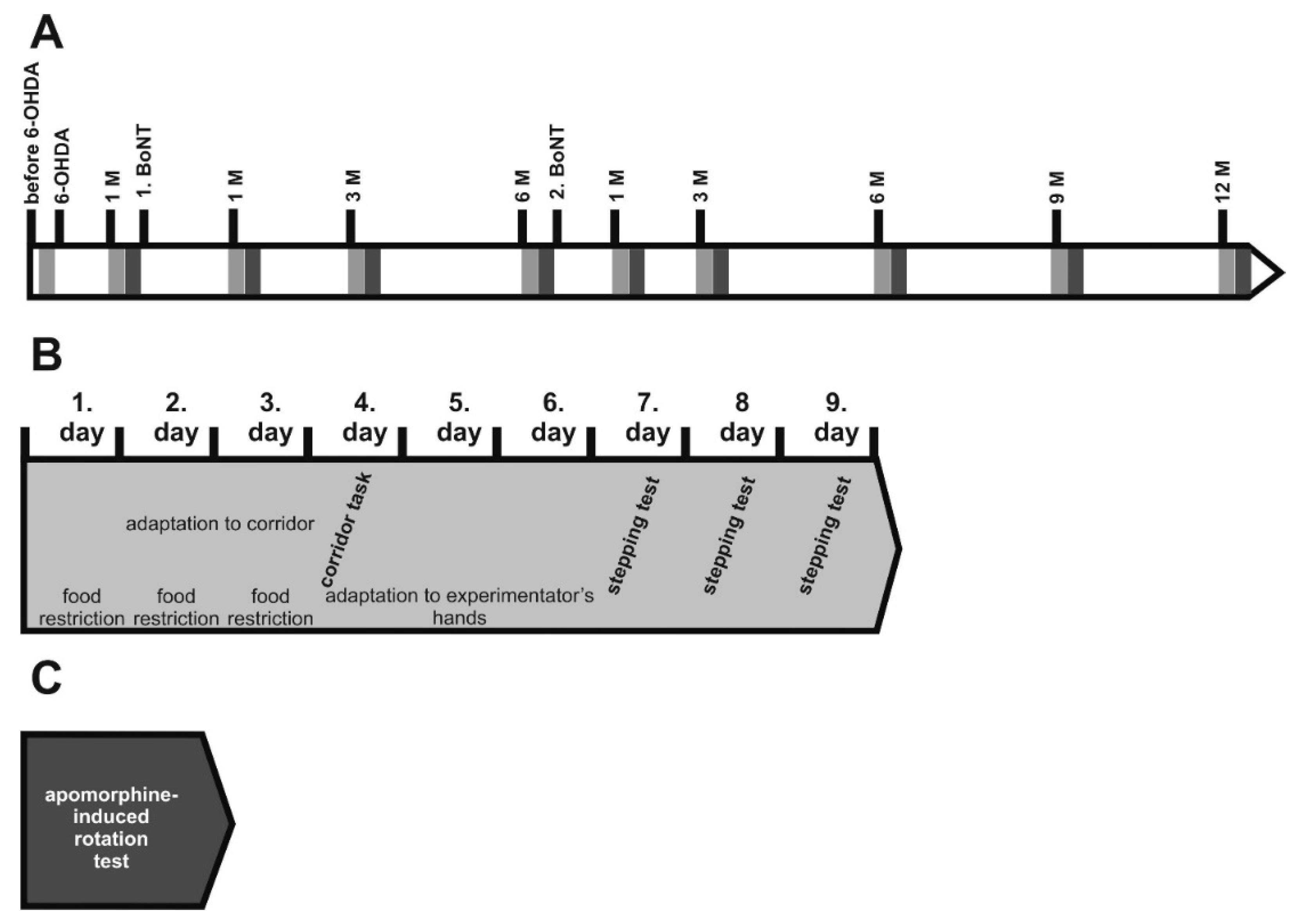
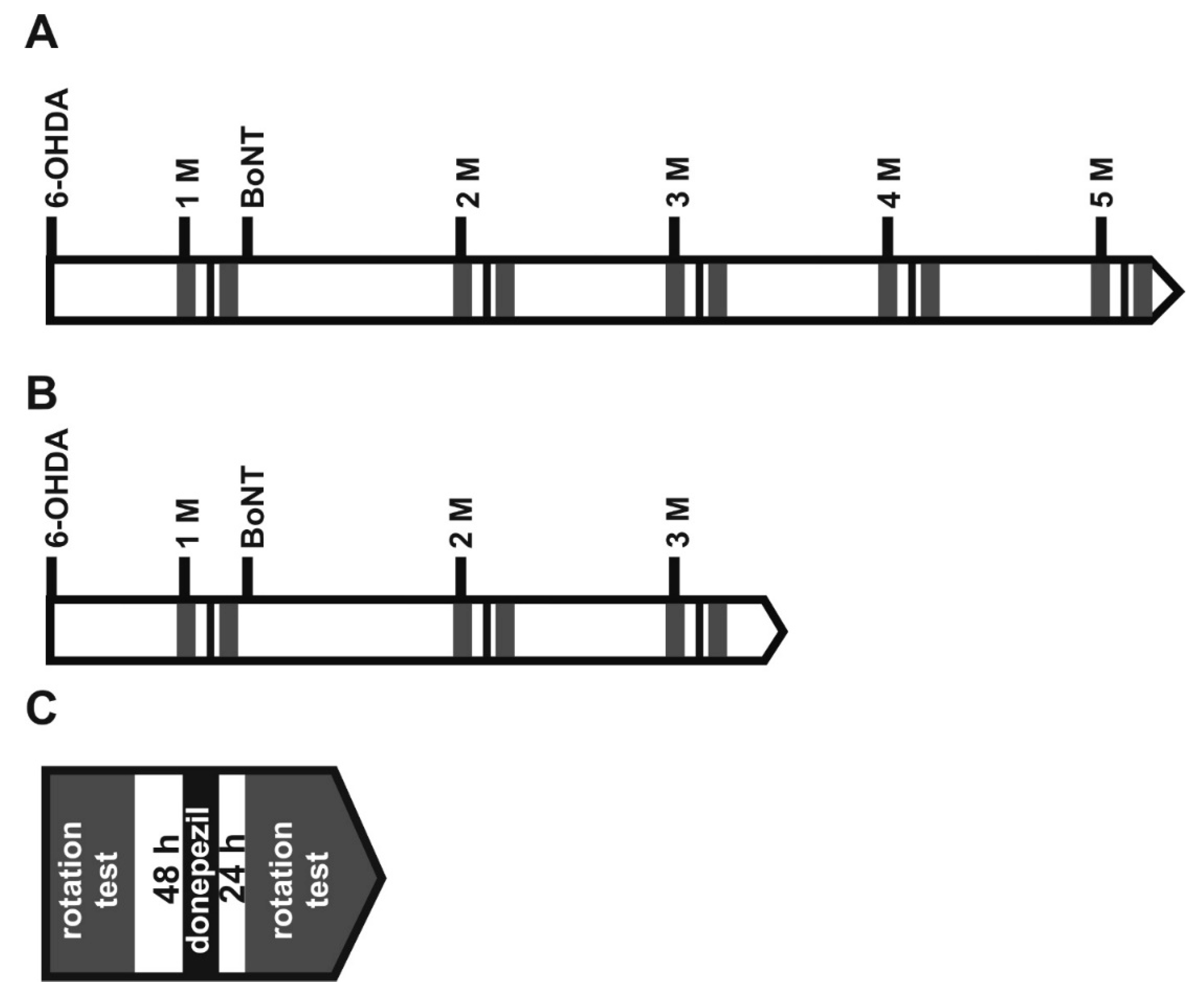
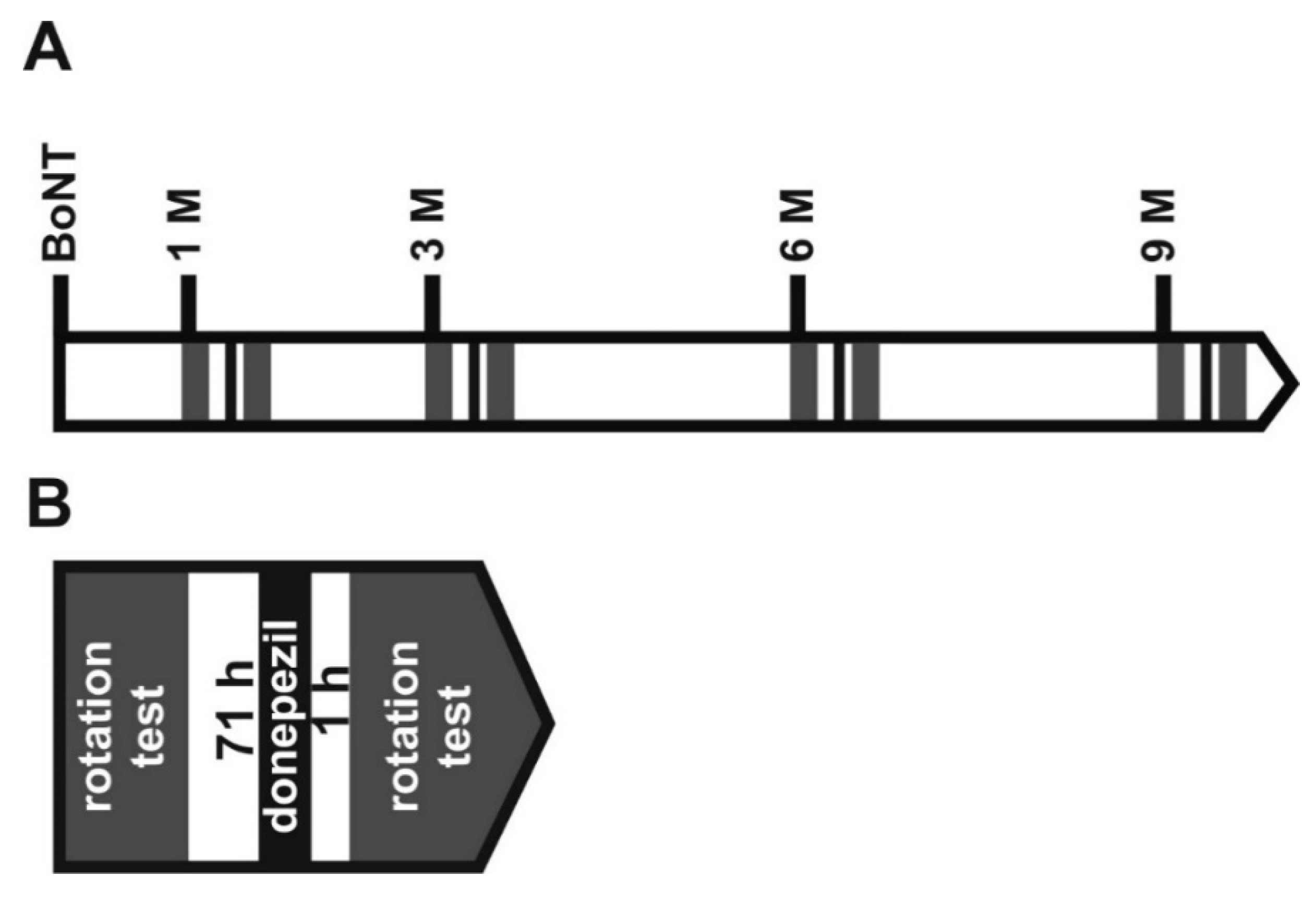
5. Materials and Methods
5.1. Animals
5.2. Induction of Hemiparkinsonism
5.3. Body Weight
5.4. Injection of BoNT-A into the Striatum
5.5. Apomorphine-Induced Rotation Test
5.6. Spontaneous Motor Tests
5.6.1. Adjusting Steps
5.6.2. Corridor Task
5.7. Donepezil Modifying Apomorphine-Induced Rotations
5.8. Data Analysis
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| 6-OHDA | 6-hydroxydopamine |
| ACh | acetylcholine |
| AChE | acetylcholinesterase |
| BoNT-A | botulinum neurotoxin-A |
| BW | body weight |
| CPu | caudate-putamen (striatum) |
| DA | dopamine |
| D2 | dopamine D2 receptor |
| EGP (=LGP) | external (lateral) globus pallidus |
| GABA | gamma-aminobutyric acid |
| hemi-PD | hemiparkinsonian |
| IGP (=MGP) | internal (medial) globus pallidus |
| M | slope/drawdown |
| MFB | medial forebrain bundle |
| MSN | medium spiny neuron |
| PD | Parkinson’s disease |
| PPN | pedunculopontine nucleus |
| RPM/d | (revolutions per minute)/day |
| SNAP-25 | synaptosomal-associated protein-25 |
| SNARE | N-ethylmaleimide-sensitive-factor attachment receptor |
| SNpc | substantia nigra pars compacta |
| STh | subthalamic nucleus |
| VL | ventrolateral thalamic nucleus |
References
- Day, M.; Wang, Z.; Ding, J.; An, X.; Ingham, C.A.; Shering, A.F.; Wokosin, D.; Ilijic, E.; Sun, Z.; Sampson, A.R.; et al. Selective elimination of glutamatergic synapses on striatopallidal neurons in Parkinson disease models. Nat. Neurosci. 2006, 9, 251–259. [Google Scholar] [CrossRef] [PubMed]
- Duvoisin, R.C. Cholinergic-Anticholinergic Antagonism in Parkinsonism. Arch. Neurol. 1967, 17, 124–136. [Google Scholar] [CrossRef] [PubMed]
- Pisani, A.; Bernardi, G.; Ding, J.; Surmeier, D.J. Re-emergence of striatal cholinergic interneurons in movement disorders. Trends Neurosci. 2007, 30, 545–553. [Google Scholar] [CrossRef] [PubMed]
- Obeso, J.Á.; Marin, C.; Rodriguez-Oroz, C.; Blesa, J.; Benitez-Temiño, B.; Mena-Segovia, J.; Rodríguez, M.; Olanow, C.W. The basal ganglia in Parkinson’s disease: Current concepts and unexplained observations. Ann. Neurol. 2008, 64, S30–S46. [Google Scholar] [CrossRef] [PubMed]
- Obeso, J.Á.; Rodríguez-Oroz, M.C.; Benitez-Temino, B.; Blesa, F.J.; Guridi, J.; Marin, C.; Rodriguez, M. Functional organization of the basal ganglia: Therapeutic implications for Parkinson’s disease. Mov. Disord. 2008, 23, S548–S559. [Google Scholar] [CrossRef] [PubMed]
- Bohnen, N.I.; Albin, R.L. The cholinergic system and Parkinson disease. Behav. Brain Res. 2011, 221, 564–573. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Clarke, C.E. Medical Management of Parkinson’s Disease. J. Neurol. Neurosurg. Psychiatry 2002, 72, i22–i27. [Google Scholar] [CrossRef] [PubMed]
- Connolly, B.S.; Lang, A.E. Pharmacological treatment of Parkinson disease: A review. JAMA 2014, 311, 1670–1683. [Google Scholar] [CrossRef] [PubMed]
- Whitney, C.M. Medications for Parkinson’s disease. Neurologist 2007, 13, 387–388. [Google Scholar] [CrossRef] [PubMed]
- Montecucco, C.; Schiavo, G. Structure and Function of Tetanus and Botulinum Neurotoxins. Q. Rev. Biophys. 1995, 28, 423–472. [Google Scholar] [CrossRef] [PubMed]
- Coffeld, J.A.; Considine, R.V.; Simpson, L.L. Clostridial neurotoxins in the age of molecular medicine. Trends Microbiol. 1994, 2, 67–69. [Google Scholar] [CrossRef]
- Wree, A.; Mix, E.; Hawlitschka, A.; Antipova, V.; Witt, M.; Schmitt, O.; Benecke, R. Intrastriatal botulinum toxin abolishes pathologic rotational behaviour and induces axonal varicosities in the 6-OHDA rat model of Parkinson’s disease. Neurobiol. Dis. 2011, 41, 291–298. [Google Scholar] [CrossRef] [PubMed]
- Antipova, V.; Hawlitschka, A.; Mix, E.; Schmitt, O.; Dräger, D.; Benecke, R.; Wree, A. Behavioral and structural effects of unilateral intrastriatal injections of botulinum neurotoxin a in the rat model of Parkinson’s disease. J. Neurosci. Res. 2013, 91, 838–847. [Google Scholar] [CrossRef] [PubMed]
- Antipova, V.A.; Holzmann, C.; Schmitt, O.; Wree, A.; Hawlitschka, A. Botulinum Neurotoxin A Injected Ipsilaterally or Contralaterally into the Striatum in the Rat 6-OHDA Model of Unilateral Parkinson’s Disease Differently Affects Behavior. Front. Behav. Neurosci. 2017, 11, 119. [Google Scholar] [CrossRef] [PubMed]
- Hawlitschka, A.; Antipova, V.; Schmitt, O.; Witt, M.; Benecke, R.; Mix, E.; Wree, A. Intracerebrally applied botulinum neurotoxin in experimental neuroscience. Curr. Pharm. Biotechnol. 2013, 14, 124–130. [Google Scholar] [CrossRef] [PubMed]
- Holzmann, C.; Dräger, D.; Mix, E.; Hawlitschka, A.; Antipova, V.; Benecke, R.; Wree, A. Effects of intrastriatal botulinum neurotoxin A on the behavior of Wistar rats. Behav. Brain Res. 2012, 234, 107–116. [Google Scholar] [CrossRef] [PubMed]
- Mehlan, J.; Brosig, H.; Schmitt, O.; Mix, E.; Wree, A.; Hawlitschka, A. Intrastriatal injection of botulinum neurotoxin-A is not cytotoxic in rat brain—A histological and stereological analysis. Brain Res. 2016, 1630, 18–24. [Google Scholar] [CrossRef] [PubMed]
- Chen, S. Clinical uses of botulinum neurotoxins: Current indications, limitations and future developments. Toxins 2012, 4, 913–939. [Google Scholar] [CrossRef] [PubMed]
- Orsini, M.; Leite, M.A.A.; Chung, T.M.; Bocca, W.; de Souza, J.A.; de Souza, O.G.; Moreira, R.P.; Bastos, V.H.; Teixeira, S.; Oliveira, A.B.; et al. Botulinum neurotoxin type A in neurology: Update. Neurol. Int. 2015, 7, 79–84. [Google Scholar] [CrossRef] [PubMed]
- Tiseo, P.J.; Foley, K.; Friedhoff, L.T. An evaluation of the pharmacokinetics of donepezil HCl in patients with moderately to severely impaired renal function. Br. J. Clin. Pharmacol. 1998, 46 (Suppl. 1), 56–60. [Google Scholar] [CrossRef] [PubMed]
- Mihara, M.; Ohnishi, A.; Tomono, Y.; Hasegawa, J.; Shimamura, Y.; Yamazaki, K.; Morishita, N. Pharmacokinetics of E2020, a new compound for Alzheimer’s disease, in healthy male volunteers. Int. J. Clin. Pharmacol. Ther. Toxicol. 1993, 31, 223–229. [Google Scholar] [PubMed]
- Ohnishi, A.; Mihara, M.; Kamakura, H.; Tomono, Y.; Hasegawa, J.; Yamazaki, K.; Morishita, N.; Tanaka, T. Comparison of the pharmacokinetics of E2020, a new compound for Alzheimer’s disease, in healthy young and elderly subjects. J. Clin. Pharmacol. 1993, 33, 1086–1091. [Google Scholar] [CrossRef] [PubMed]
- Snape, M.F.; Misra, A.; Murray, T.K.; De Souza, R.J.; Williams, J.L.; Cross, A.J.; Green, A.R. A comparative study in rats of the in vitro and in vivo pharmacology of the acetylcholinesterase inhibitors tacrine, donepezil and NXX-066. Neuropharmacology 1999, 38, 181–193. [Google Scholar] [CrossRef]
- Barnes, C.A.; Meltzer, J.; Houston, F.; Orr, G.; McGann, K.; Wenk, G.L. Chronic treatment of old rats with donepezil or galantamine: Effects on memory, hippocampal plasticity and nicotinic receptors. Neuroscience 2000, 99, 17–23. [Google Scholar] [CrossRef]
- Mumenthaler, M.S.; Yesavage, J.A.; Taylor, J.L.; O’Hara, R.; Friedman, L.; Lee, H.; Kraemer, H.C. Psychoactive drugs and pilot performance: A comparison of nicotine, donepezil, and alcohol effects. Neuropsychopharmacology 2003, 28, 1366–1373. [Google Scholar] [CrossRef] [PubMed]
- Geerts, H.; Guillaumat, P.O.; Grantham, C.; Bode, W.; Anciaux, K.; Sachak, S. Brain levels and acetylcholinesterase inhibition with galantamine and donepezil in rats, mice, and rabbits. Brain Res. 2005, 1033, 186–193. [Google Scholar] [CrossRef] [PubMed]
- Goh, C.W.; Aw, C.C.; Lee, J.H.; Chen, C.P.; Browne, E.R. Pharmacokinetic and pharmacodynamic properties of cholinesterase inhibitors donepezil, tacrine, and galantamine in aged and young lister hooded rats. Drug Metab. Dispos. 2011, 39, 402–411. [Google Scholar] [CrossRef] [PubMed]
- Nirogi, R.; Bhyrapuneni, G.; Kandikere, V.; Benade, V.; Muddana, N.; Saralaya, R.; Irappanavar, S.; Ponnamaneni, R.; Mukkanti, K. Concurrent administration of atypical antipsychotics and donepezil: Drug interaction study in rats. Eur. J. Drug Metab. Pharmacokinet. 2012, 37, 155–161. [Google Scholar] [CrossRef] [PubMed]
- Matsui, K.; Mishima, M.; Nagai, Y.; Yuzuriha, T.; Yoshimura, T. Absorption, distribution, metabolism, and excretion of donepezil (Aricept) after a single oral administration to Rat. Drug Metab. Dispos. 1999, 27, 1406–1414. [Google Scholar] [PubMed]
- Kirik, D.; Georgievska, B.; Burger, C.; Winkler, C.; Muzyczka, N.; Mandel, R.J.; Björklund, A. Reversal of motor impairments in parkinsonian rats by continuous intrastriatal delivery of L-dopa using rAAV-mediated gene transfer. Proc. Natl. Acad. Sci. USA 2002, 99, 4708–4713. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kirik, D.; Rosenblad, C.; Bjorklund, A. Preservation of a functional nigrostriatal dopamine pathway by GDNF in the intrastriatal 6-OHDA lesion model depends on the site of administration of the trophic factor. Eur. J. Neurosci. 2000, 12, 3871–3882. [Google Scholar] [CrossRef] [PubMed]
- Kirik, D.; Rosenblad, C.; Bjorklund, A.; Mandel, R.J. Long-term rAAV-mediated gene transfer of GDNF in the rat Parkinson’s model: Intrastriatal but not intranigral transduction promotes functional regeneration in the lesioned nigrostriatal system. J. Neurosci. 2000, 20, 4686–4700. [Google Scholar] [CrossRef] [PubMed]
- Olsson, M.; Nikkhah, G.; Bentlage, C.; Björklund, A. Forelimb akinesia in the rat Parkinson model: Differential effects of dopamine agonists and nigral transplants as assessed by a new stepping test. J. Neurosci. 1995, 15, 3863–3875. [Google Scholar] [CrossRef] [PubMed]
- Dronfield, S.; Egan, K.; Marsden, C.A.; Green, A.R. Comparison of donepezil-, tacrine-, rivastigmine-and metrifonateinduced central and peripheral cholinergically mediated responses in the rat. J. Psychopharmacol. 2000, 14, 275–279. [Google Scholar] [CrossRef] [PubMed]
- Haug, K.H.; Bogen, I.L.; Osmundsen, H.; Walaas, I.; Fonnum, F. Effects on cholinergic markers in rat brain and blood after short and prolonged administration of donepezil. Neurochem. Res. 2005, 30, 1511–1520. [Google Scholar] [CrossRef] [PubMed]
- Hatip-Al-Khatib, I.; Takashi, A.; Egashira, N.; Iwasaki, K.; Fujiwara, M. Comparison of the effect of TAK-147 (zanapezil) and E-2020 (donepezil) on extracellular acetylcholine level and blood flow in the ventral hippocampus of freely moving rats. Brain Res. 2004, 1012, 169–176. [Google Scholar] [CrossRef] [PubMed]
- Kosasa, T.; Kuriya, Y.; Yamanishi, Y. Effect of donepezil hydrochloride (E2020) on extracellular acetylcholine concentration in the cerebral cortex of rats. Jpn. J. Pharmacol. 1999, 81, 216–222. [Google Scholar] [CrossRef] [PubMed]
- Mann, T.; Zilles, K.; Dikow, H.; Hellfritsch, A.; Cremer, M.; Piel, M.; Rösch, F.; Hawlitschka, A.; Schmitt, O.; Wree, A. Dopamine, Noradrenaline and Serotonin Receptor Densities in the Striatum of Hemiparkinsonian Rats following Botulinum Neurotoxin-A Injection. Neuroscience 2018, 374, 187–204. [Google Scholar] [CrossRef] [PubMed]
- Wedekind, F.; Oskamp, A.; Lang, M.; Hawlitschka, A.; Zilles, K.; Wree, A.; Bauer, A. Intrastriatal administration of botulinum neurotoxin A normalizes striatal D2R binding and reduces striatal D1R binding in male hemiparkinsonian rats. J. Neurosci. Res. 2017. [Google Scholar] [CrossRef]
- Rogozhin, A.A.; Pang, K.K.; Bukharaeva, E.; Young, C.; Slater, C.R. Recovery of mouse neuromuscular junctions from single and repeated injections of botulinum neurotoxin A. J. Physiol. 2008, 586, 3163–3182. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Şen, A.; Arpaci, B. Effects of Repeated Botulinum Toxin Treatment for Sialorrhea in Patients with Parkinson’s disease. Noro Psikiyatr. Ars. 2015, 52, 69–72. [Google Scholar] [CrossRef] [PubMed]
- Brashear, A.; Hogan, P.; Wooten-Watts, M.; Marchetti, A.; Magar, R.; Martin, J. Longitudinal assessment of the dose consistency of botulinum toxin type A (BOTOX®) for cervical dystonia. Adv. Ther. 2005, 22, 49–55. [Google Scholar] [CrossRef] [PubMed]
- Gordon, M.F.; Barron, R. Effectiveness of repeated treatment with botulinum toxin type A across different conditions. South. Med. J. 2006, 99, 853–861. [Google Scholar] [CrossRef] [PubMed]
- Colosimo, C.; Tiple, D.; Berardelli, A. Efficacy and safety of long-term botulinum toxin treatment in craniocervical dystonia: A systematic review. Neurotox. Res. 2012, 22, 265–273. [Google Scholar] [CrossRef] [PubMed]
- Flynn, T.C. Botulinum toxin: Examining duration of effect in facial aesthetic applications. Am. J. Clin. Dermatol. 2010, 11, 183–199. [Google Scholar] [CrossRef] [PubMed]
- Spirduso, W.W.; Gilliam, P.E.; Schallert, T.; Upchurch, M.; Vaughn, D.M.; Wilcox, R.E. Reactive capacity: A sensitive behavioral marker of movement initiation and nigrostriatal dopamine function. Brain Res. 1985, 335, 45–54. [Google Scholar] [CrossRef]
- Miklyaeva, E.I.; Martens, D.J.; Whishaw, I.Q. Impairments and compensatory adjustments in spontaneous movement after unilateral dopamine depletion in rats. Brain Res. 1995, 681, 23–40. [Google Scholar] [CrossRef]
- Tillerson, J.L.; Cohen, A.D.; Philhower, J.; Miller, G.W.; Zigmond, M.J.; Schallert, T. Forced limb-use effects on the behavioral and neurochemical effects of 6-hydroxydopamine. J. Neurosci. 2001, 21, 4427–4435. [Google Scholar] [CrossRef] [PubMed]
- Cho, J.; Duke, D.; Manzino, L.; Sonsalla, P.K.; West, M.O. Dopamine depletion causes fragmented clustering of neurons in the sensorimotor striatum: Evidence of lasting reorganization of corticostriatal input. J. Comp. Neurol. 2002, 452, 24–37. [Google Scholar] [CrossRef] [PubMed]
- Fitzsimmons, D.F.; Moloney, T.C.; Dowd, E. Further validation of the corridor task for assessing deficit and recovery in the hemi-Parkinsonian rat: Restoration of bilateral food retrieval by dopamine receptor agonism. Behav. Brain Res. 2006, 169, 352–355. [Google Scholar] [CrossRef] [PubMed]
- Plowman, E.K.; Thomas, N.J.; Kleim, J.A. Striatal dopamine depletion induces forelimb motor impairments and disrupts forelimb movement representations within the motor cortex. J. Parkinsons. Dis. 2011, 1, 93–100. [Google Scholar] [CrossRef] [PubMed]
- Oudega, M.; Perez, M.A. Corticospinal reorganization after spinal cord injury. J. Physiol. 2012, 590, 3647–3663. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Takeuchi, Y.; Takahashi, O.; Satoda, T.; Matsushima, R. Long dendrites of the parabrachial nucleus neurons projecting to the amygdala: A horseradish peroxidase and Golgi study in the cat. Exp. Neurol. 1987, 96, 203–207. [Google Scholar] [CrossRef]
- Kathe, C.; Hutson, T.H.; Chen, Q.; Shine, H.D.; McMahon, S.B.; Moon, L.D.F. Unilateral pyramidotomy of the corticospinal tract in rats for assessment of neuroplasticity-inducing therapies. J. Vis. Exp. 2014. [Google Scholar] [CrossRef] [PubMed]
- Wree, A.; Schmitt, O. Basal Ganglia. In Brain Mapping; Toga, A.W., Ed.; Elsevier Academic: Cambridge, MA, USA, 2015; pp. 217–227. ISBN 9780123973160. [Google Scholar]
- Huang, W.; Foster, J.A.; Rogachefsky, A.S. Pharmacology of botulinum toxin. J. Am. Acad. Dermatol. 2000, 43, 249–259. [Google Scholar] [CrossRef] [PubMed]
- Rosales, R.L.; Bigalke, H.; Dressler, D. Pharmacology of botulinum toxin: Differences between type A preparations. Eur. J. Neurol. 2006, 13 (Suppl. 1), 2–10. [Google Scholar] [CrossRef] [PubMed]
- Nigam, P.K.; Nigam, A. Botulinum toxin. Indian J. Dermatol. 2010, 55, 8–14. [Google Scholar] [CrossRef] [PubMed]
- Ashton, A.C.; Dolly, J.O. Characterization of the Inhibitory Action of Botulinum Neurotoxin Type A on the Release of Several Transmitters from Rat Cerebrocortical Synaptosomes. J. Neurochem. 1988, 50, 1808–1816. [Google Scholar] [CrossRef] [PubMed]
- Bigalke, H.; Heller, I.; Bizzini, B.; Habermann, E. Tetanus toxin and botulinum A toxin inhibit release and uptake of various transmitters, as studied with particulate preparations from rat brain and spinal cord. Naunyn. Schmiedebergs. Arch. Pharmacol. 1981, 316, 244–251. [Google Scholar] [CrossRef] [PubMed]
- Pearce, L.B.; First, E.R.; MacCallum, R.D.; Gupta, A. Pharmacologic characterization of botulinum toxin for basic science and medicine. Toxicon 1997, 35, 1373–1412. [Google Scholar] [CrossRef]
- Kirik, D.; Rosenblad, C.; Björklund, A. Characterization of behavioral and neurodegenerative changes following partial lesions of the nigrostriatal dopamine system induced by intrastriatal 6-hydroxydopamine in the rat. Exp. Neurol. 1998, 152, 259–277. [Google Scholar] [CrossRef] [PubMed]
- Fang, X.; Sugiyama, K.; Akamine, S.; Namba, H. The stepping test and its learning process in different degrees of unilateral striatal lesions by 6-hydroxydopamine in rats. Neurosci. Res. 2006, 55, 403–409. [Google Scholar] [CrossRef] [PubMed]
- Whishaw, I.Q.; Woodward, N.C.; Miklyaeva, E.; Pellis, S.M. Analysis of limb use by control rats and unilateral DA-depleted rats in the Montoya staircase test: Movements, impairments and compensatory strategies. Behav. Brain Res. 1997, 89, 167–177. [Google Scholar] [CrossRef]
- Tseng, K.Y.; Kargieman, L.; Gacio, S.; Riquelme, L.A.; Murer, M.G. Consequences of partial and severe dopaminergic lesion on basal ganglia oscillatory activity and akinesia. Eur. J. Neurosci. 2005, 22, 2579–2586. [Google Scholar] [CrossRef] [PubMed]
- Kelsey, J.E.; Mague, S.D.; Pijanowski, R.S.; Harris, R.C.; Kleckner, N.W.; Matthews, R.T. NMDA receptor antagonists ameliorate the stepping deficits produced by unilateral medial forebrain bundle injections of 6-OHDA in rats. Psychopharmacology 2004, 175, 179–188. [Google Scholar] [CrossRef] [PubMed]
- Barnéoud, P.; Descombris, E.; Aubin, N.; Abrous, D.N. Evaluation of simple and complex sensorimotor behaviours in rats with a partial lesion of the dopaminergic nigrostriatal system. Eur. J. Neurosci. 2000, 12, 322–336. [Google Scholar] [CrossRef] [PubMed]
- Chang, J.W.; Wachtel, S.R.; Young, D.; Kang, U.J. Biochemical and anatomical characterization of forepaw adjusting steps in rat models of Parkinson’s disease: Studies on medial forebrain bundle and striatal lesions. Neuroscience 1999, 88, 617–628. [Google Scholar] [CrossRef]
- Deumens, R.; Blokland, A.; Prickaerts, J. Modeling Parkinson’s disease in rats: An evaluation of 6-OHDA lesions of the nigrostriatal pathway. Exp. Neurol. 2002, 175, 303–317. [Google Scholar] [CrossRef] [PubMed]
- Seeger-Armbruster, S.; Von Ameln-Mayerhofer, A. Short- and long-term unilateral 6-hydroxydopamine lesions in rats show different changes in characteristics of spontaneous firing of substantia Nigra pars reticulata neurons. Exp. Brain Res. 2013, 224, 15–24. [Google Scholar] [CrossRef] [PubMed]
- Solari, N.; Bonito-Oliva, A.; Fisone, G.; Brambilla, R. Understanding cognitive deficits in Parkinson’s disease: Lessons from preclinical animal models. Learn. Mem. 2013, 20, 592–600. [Google Scholar] [CrossRef] [PubMed]
- Kawashima, K.; Sato, A.; Yoshizawa, M.; Fujii, T.; Fujimoto, K.; Suzuki, T. Effects of the centrally acting cholinesterase inhibitors tetrahydroaminoacridine and E2020 on the basal concentration of extracellular acetylcholine in the hippocampus of freely moving rats. Naunyn. Schmiedebergs. Arch. Pharmacol. 1994, 350, 523–528. [Google Scholar] [CrossRef] [PubMed]
- Liang, Y.Q.; Tang, X.C. Comparative studies of huperzine A, donepezil, and rivastigmine on brain acetylcholine, dopamine, norepinephrine, and 5-hydroxytryptamine levels in freely-moving rats. Acta Pharmacol. Sin. 2006, 27, 1127–1136. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liang, Y.Q.; Tang, X.C. Comparative effects of huperzine A, donepezil and rivastigmine on cortical acetylcholine level and acetylcholinesterase activity in rats. Neurosci. Lett. 2004, 361, 56–59. [Google Scholar] [CrossRef] [PubMed]
- Castro, R.; Abreu, P.; Calzadilla, C.H.; Rodriguez, M. Increased or decreased locomotor response in rats following repeated administration of apomorphine depends on dosage interval. Psychopharmacology 1985, 85, 333–339. [Google Scholar] [CrossRef] [PubMed]
- Gancher, S.; Crabbe, J.; Garland, A.; Lea, E.; Woodward, W. Dose- and duration-dependent tolerance to rotational effects of apomorphine in a rat model of Parkinsonism. J. Pharmacol. Exp. Ther. 1995, 272, 275–281. [Google Scholar] [PubMed]
- Naik, R.S.; Hartmann, J.; Kiewert, C.; Duysen, E.G.; Lockridge, O.; Klein, J. Effects of rivastigmine and donepezil on brain acetylcholine levels in acetylcholinesterase-deficient mice. J. Pharm. Pharm. Sci. 2009, 12, 79–85. [Google Scholar] [CrossRef] [PubMed]
- Babiloni, C.; Del Percio, C.; Infarinato, F.; Messano, F.P.; Forloni, G.; Kelley, J.; Dix, S.; Bordet, R.; Richardson, J.; Limatola, C.; et al. Effects of Donepezil on EEG-EMG markres in wild type (WT) and triple transgenic (TAUPS2APP) mice. Alzheimer’s Dement. 2014, 10, P661. [Google Scholar] [CrossRef]
- Shaw, K.E.; Bondi, C.O.; Light, S.H.; Massimino, L.A.; McAloon, R.L.; Monaco, C.M.; Kline, A.E. Donepezil Is Ineffective in Promoting Motor and Cognitive Benefits after Controlled Cortical Impact Injury in Male Rats. J. Neurotrauma 2013, 30, 557–564. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dooley, M.; Lamb, H.M. Donepezil. A review of its use in Alzheimer’s disease. Drugs Aging 2000, 16, 199–226. [Google Scholar] [CrossRef] [PubMed]
- Rogers, S.L.; Friedhoff, L.T. The efficacy and safety of donepezil in patients with Alzheimer’s disease: Results of a US Multicentre, Randomized, Double-Blind, Placebo-Controlled Trial. The Donepezil Study Group. Dementia 1996, 7, 293–303. [Google Scholar] [CrossRef] [PubMed]
- Evans, M.; Ellis, A.; Watson, D.; Chowdhury, T. Sustained cognitive improvement following treatment of Alzheimer’s disease with donepezil. Int. J. Geriatr. Psychiatry 2000, 15, 50–53. [Google Scholar] [CrossRef]
- Wilkinson, D.; Doody, R.; Helme, R.; Taubman, K.; Mintzer, J.; Kertesz, A.; Pratt, R.D. Donepezil in vascular dementia: A randomized, placebo-controlled study. Neurology 2003, 61, 479–486. [Google Scholar] [CrossRef] [PubMed]
- Bohnen, N.; Kaufer, D.; Hendrickson, R.; Ivanco, L.; Moore, R.; DeKosky, S. Effects of donepezil on motor function in patients with Alzheimer disease. J. Clin. Psychopharmacol. 2004, 24, 354–356. [Google Scholar] [CrossRef] [PubMed]
- Ravina, B.; Putt, M.; Siderowf, A.; Farrar, J.T.; Gillespie, M.; Crawley, A.; Fernandez, H.H.; Trieschmann, M.M.; Reichwein, S.; Simuni, T. Donepezil for dementia in Parkinson’s disease: A randomised, double blind, placebo controlled, crossover study. J. Neurol. Neurosurg. Psychiatry 2005, 76, 934–939. [Google Scholar] [CrossRef] [PubMed]
- Mori, E.; Ikeda, M.; Nakagawa, M.; Miyagishi, H.; Yamaguchi, H.; Kosaka, K. Effects of donepezil on extrapyramidal symptoms in patients with dementia with lewy bodies: A secondary pooled analysis of two randomized-controlled and two open-label long-term extension studies. Dement. Geriatr. Cogn. Disord. 2015, 40, 186–198. [Google Scholar] [CrossRef] [PubMed]
- Bourke, D.; Druckenbrod, R.W. Possible association between donepezil and worsening Parkinson’s disease. Ann. Pharmacother. 1998, 32, 610–611. [Google Scholar] [CrossRef] [PubMed]
- Shea, C.; MacKnight, C.; Rockwood, K. Donepezil for treatment of dementia with Lewy bodies: A case series of nine patients. Int. Psychogeriatr. 1998, 10, 229–238. [Google Scholar] [CrossRef] [PubMed]
- Carcenac, D.; Martin-Hunyadi, C.; Kiesmann, M.; Demuynck-Roegel, C.; Alt, M.; Kuntzmann, F. Extra-pyramidal syndrome induced by donepezil. Press. Méd. (Paris Fr. 1983) 2000, 29, 992–993. [Google Scholar]
- Fabbrini, G.; Barbanti, P.; Aurilia, C.; Pauletti, C.; Lenzi, G.L.; Meco, G. Donepezil in the treatment of hallucinations and delusions in Parkinson’s disease. Neurol. Sci. 2002, 23, 41–43. [Google Scholar] [CrossRef] [PubMed]
- Litvan, I.; Phipps, M.; Pharr, V.L.; Hallett, M.; Grafman, J.; Salazar, A. Randomized placebo-controlled trial of donepezil in patients with progressive supranuclear palsy. Neurology 2001, 57, 467–473. [Google Scholar] [CrossRef] [PubMed]
- Kwak, Y.T.; Han, I.W.; Baik, J.; Koo, M.S. Relation between cholinesterase inhibitor and Pisa syndrome. Lancet 2000, 355, 2222. [Google Scholar] [CrossRef]
- Paxinos, G.; Watson, C. The Rat Brain in Stereotaxic Coordinates, 5th ed.; Elsevier Academic Press: Amsterdam, The Netherlands; London, UK, 2005; ISBN 0120885727. [Google Scholar]
- Schwarting, R.K.W.; Huston, J.P. The unilateral 6-hydroxydopamine lesion model in behavioral brain research. Analysis of functional deficits, recovery and treatments. Prog. Neurobiol. 1996, 50, 275–331. [Google Scholar] [CrossRef]
- Ungerstedt, U.; Arbuthnott, G.W. Quantitative recording of rotational behavior in rats after 6-hydroxy-dopamine lesions of the nigrostriatal dopamine system. Brain Res. 1970, 24, 485–493. [Google Scholar] [CrossRef]
- Schallert, T.; De Ryck, M.; Whishaw, I.Q.; Ramirez, V.D.; Teitelbaum, P. Excessive bracing reactions and their control by atropine and L-DOPA in an animal analog of Parkinsonism. Exp. Neurol. 1979, 64, 33–43. [Google Scholar] [CrossRef]
- Abedi, P.M.; Delaville, C.; De Deurwaerdere, P.; Benjelloun, W.; Benazzouz, A. Intrapallidal administration of 6-hydroxydopamine mimics in large part the electrophysiological and behavioral consequences of major dopamine depletion in the rat. Neuroscience 2013, 236, 289–297. [Google Scholar] [CrossRef] [PubMed]
- Grealish, S.; Mattsson, B.; Draxler, P.; Björklund, A. Characterisation of behavioural and neurodegenerative changes induced by intranigral 6-hydroxydopamine lesions in a mouse model of Parkinson’s disease. Eur. J. Neurosci. 2010, 31, 2266–2278. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schackel, S.; Pauly, M.-C.; Piroth, T.; Nikkhah, G.; Döbrössy, M.D. Donor age dependent graft development and recovery in a rat model of Huntington’s disease: Histological and behavioral analysis. Behav. Brain Res. 2013, 256, 56–63. [Google Scholar] [CrossRef] [PubMed]
- Jouve, L.; Salin, P.; Melon, C.; Goff, L.K.-L. Deep Brain Stimulation of the Center Median-Parafascicular Complex of the Thalamus Has Efficient Anti-Parkinsonian Action Associated with Widespread Cellular Responses in the Basal Ganglia Network in a Rat Model of Parkinson’s Disease. J. Neurosci. 2010, 30, 9919–9928. [Google Scholar] [CrossRef] [PubMed]
- Dowd, E.; Monville, C.; Torres, E.M.; Dunnett, S.B. The Corridor Task: A simple test of lateralised response selection sensitive to unilateral dopamine deafferentation and graft-derived dopamine replacement in the striatum. Brain Res. Bull. 2005, 68, 24–30. [Google Scholar] [CrossRef] [PubMed]
- Kaindlstorfer, C.; García, J.; Winkler, C.; Wenning, G.K.; Nikkhah, G.; Döbrössy, M.D. Behavioral and histological analysis of a partial double-lesion model of parkinson-variant multiple system atrophy. J. Neurosci. Res. 2012, 90, 1284–1295. [Google Scholar] [CrossRef] [PubMed]










© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hawlitschka, A.; Holzmann, C.; Wree, A.; Antipova, V. Repeated Intrastriatal Botulinum Neurotoxin-A Injection in Hemiparkinsonian Rats Increased the Beneficial Effect on Rotational Behavior. Toxins 2018, 10, 368. https://0-doi-org.brum.beds.ac.uk/10.3390/toxins10090368
Hawlitschka A, Holzmann C, Wree A, Antipova V. Repeated Intrastriatal Botulinum Neurotoxin-A Injection in Hemiparkinsonian Rats Increased the Beneficial Effect on Rotational Behavior. Toxins. 2018; 10(9):368. https://0-doi-org.brum.beds.ac.uk/10.3390/toxins10090368
Chicago/Turabian StyleHawlitschka, Alexander, Carsten Holzmann, Andreas Wree, and Veronica Antipova. 2018. "Repeated Intrastriatal Botulinum Neurotoxin-A Injection in Hemiparkinsonian Rats Increased the Beneficial Effect on Rotational Behavior" Toxins 10, no. 9: 368. https://0-doi-org.brum.beds.ac.uk/10.3390/toxins10090368