Occurrence and Identification of Aspergillus Section Flavi in the Context of the Emergence of Aflatoxins in French Maize

 , , , and
, , , and
Abstract
:1. Introduction
2. Results
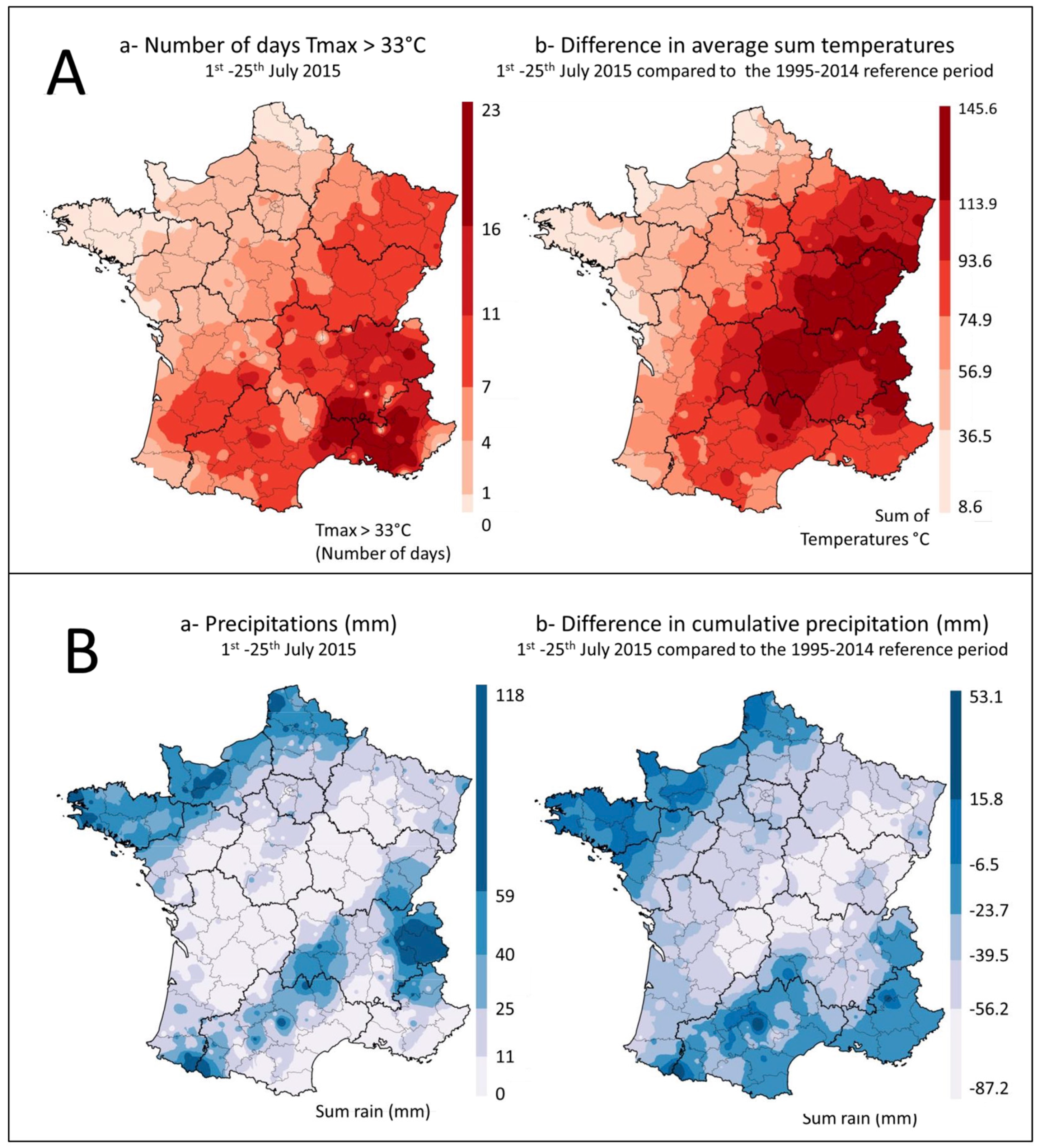
2.1. Climatic Data
2.2. Occurrence of Aflatoxins in Maize Samples
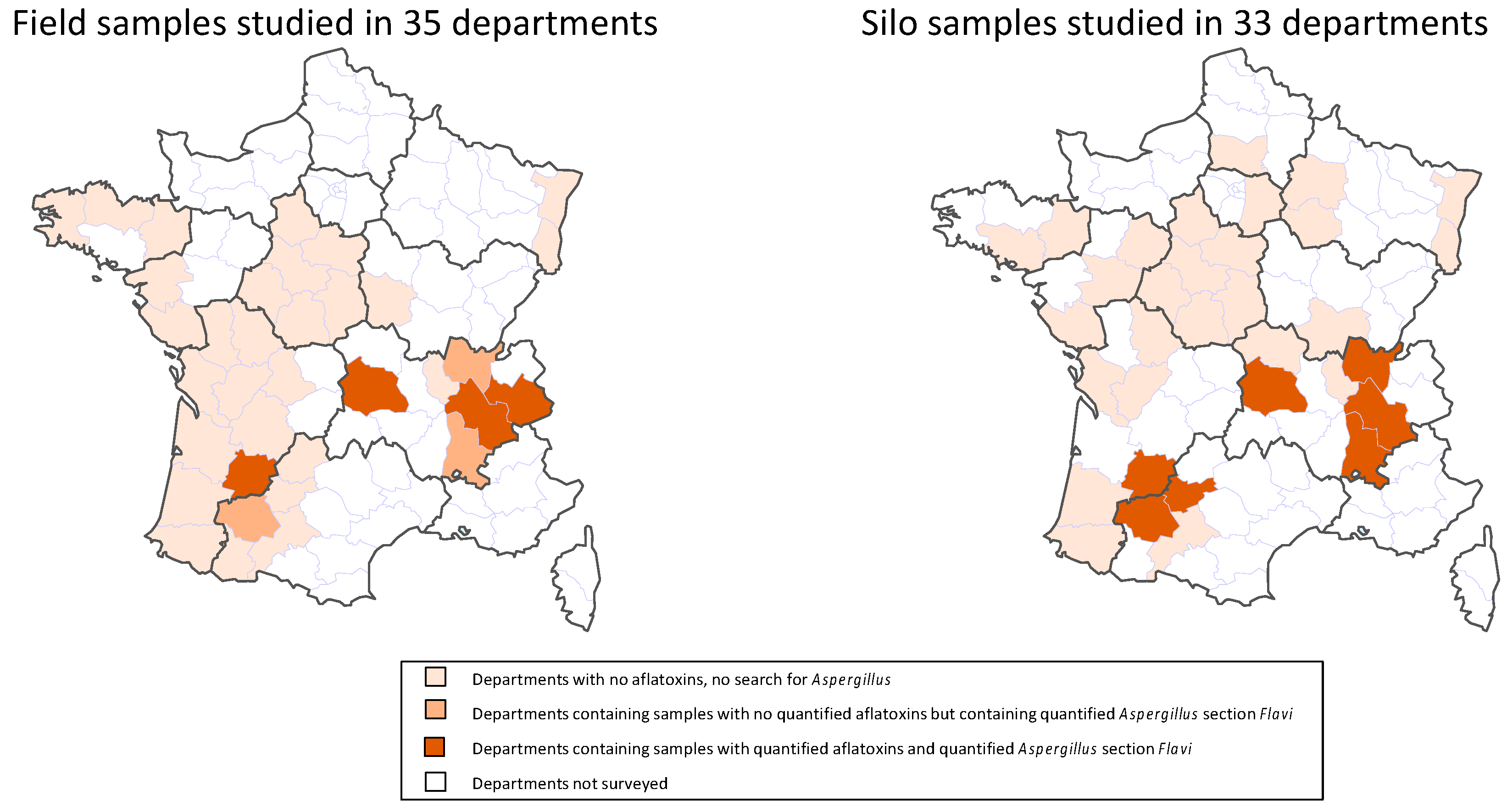
2.2.1. Number and Location of Contaminated Samples
2.2.2. Types and Levels of AF Contamination
2.3. Fungal Flora of Maize Samples
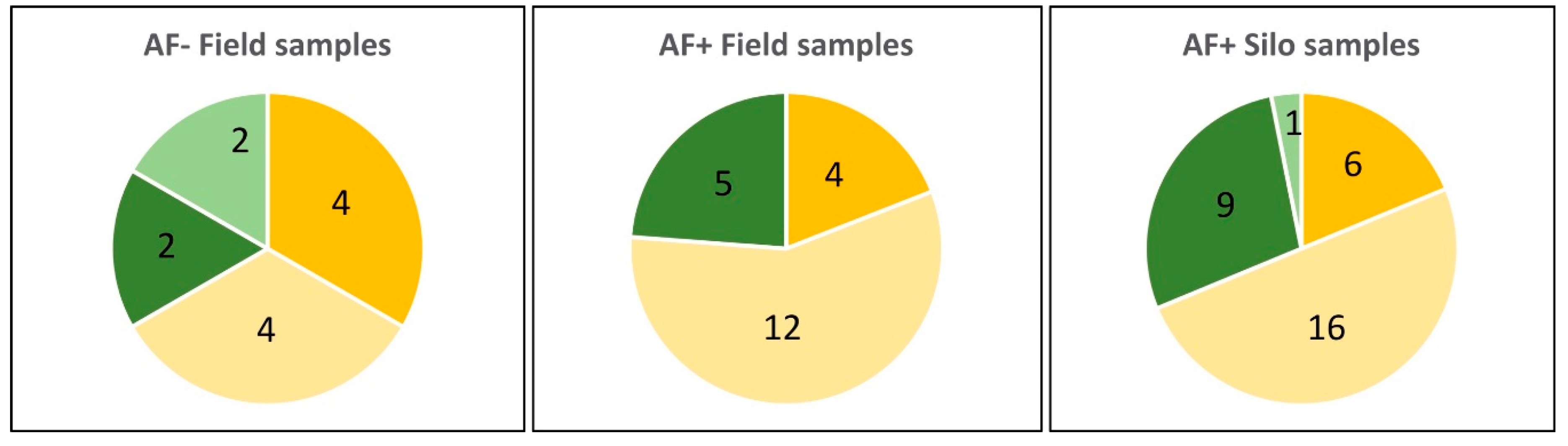
2.3.1. AF+ Samples
2.3.2. AF− Samples
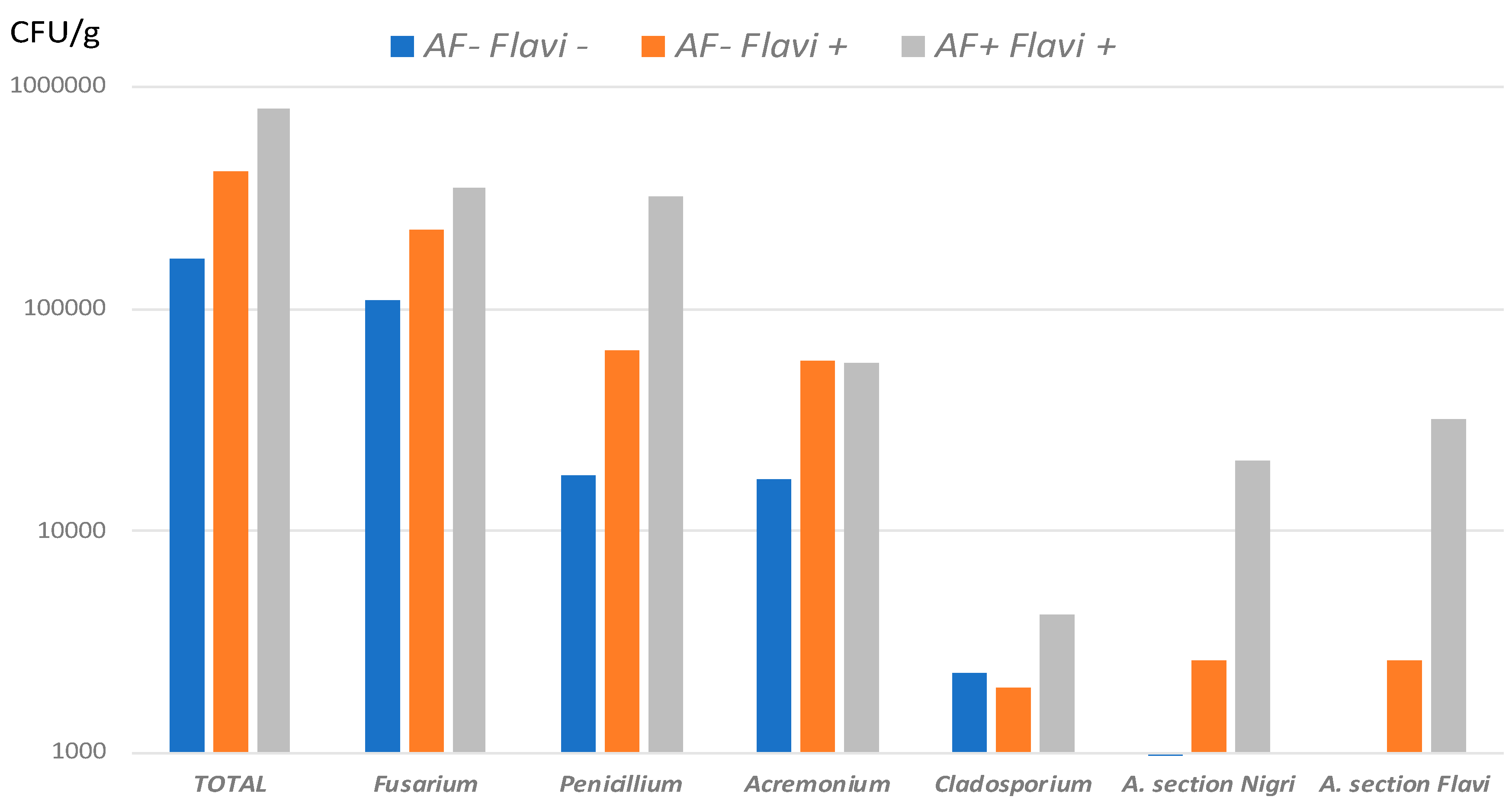
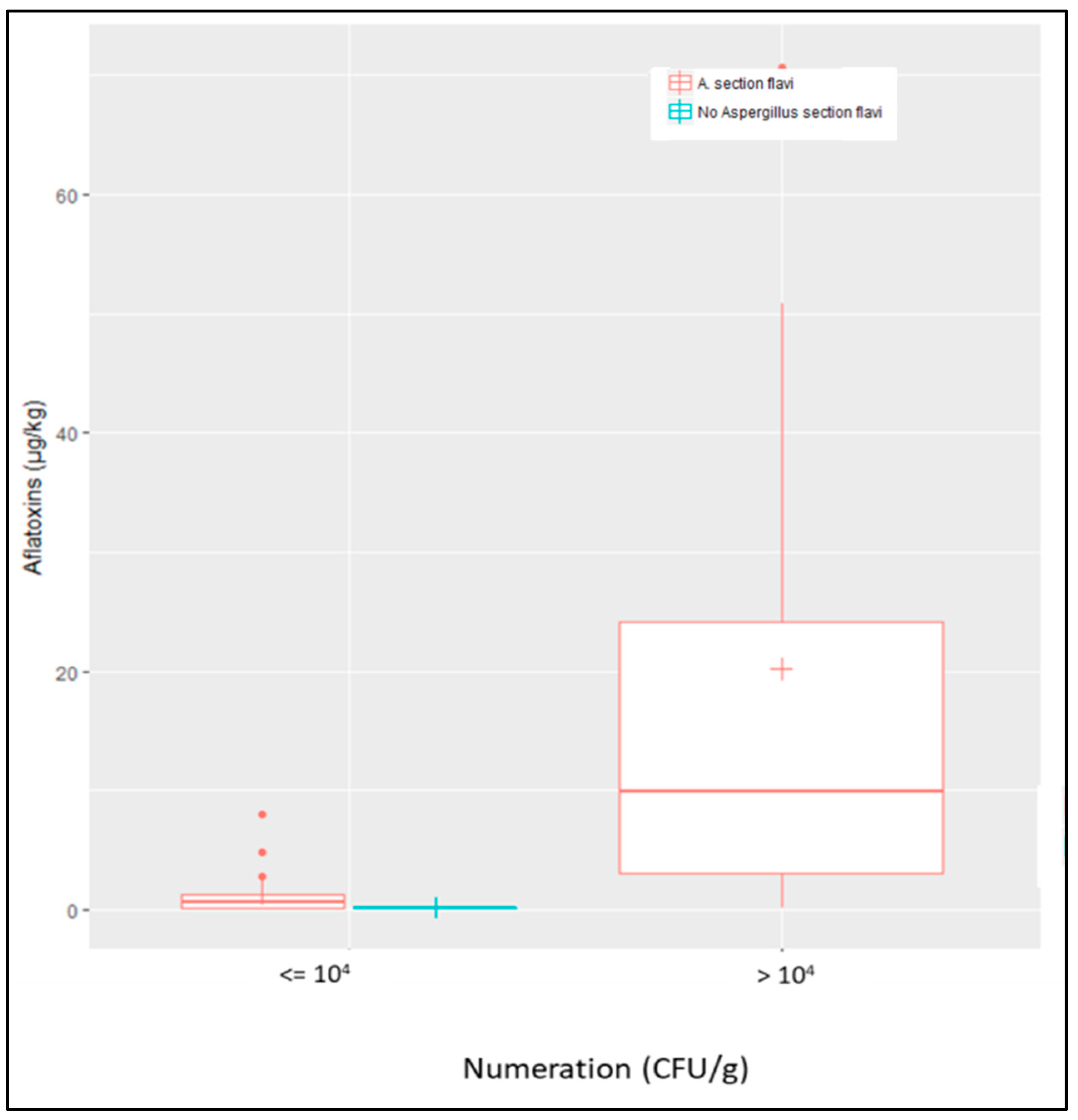
2.3.3. Quantitative Comparison
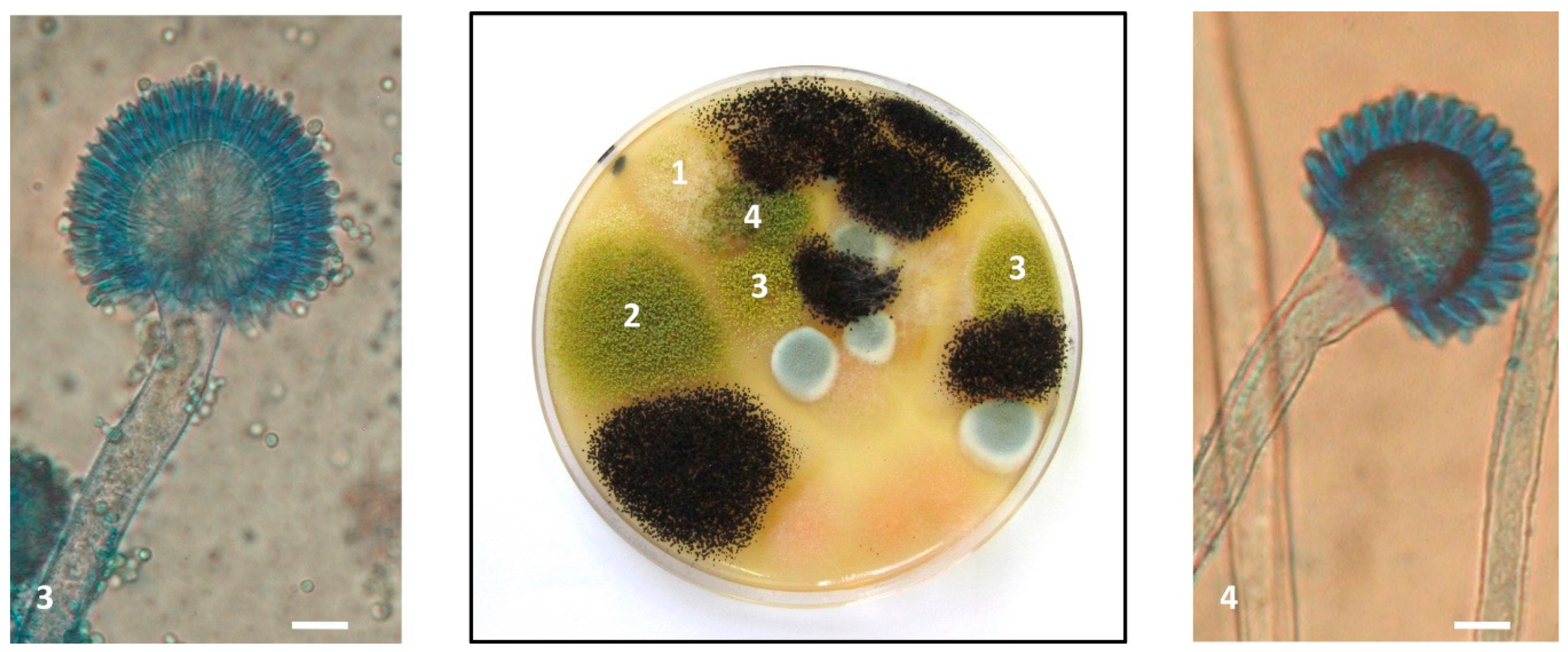
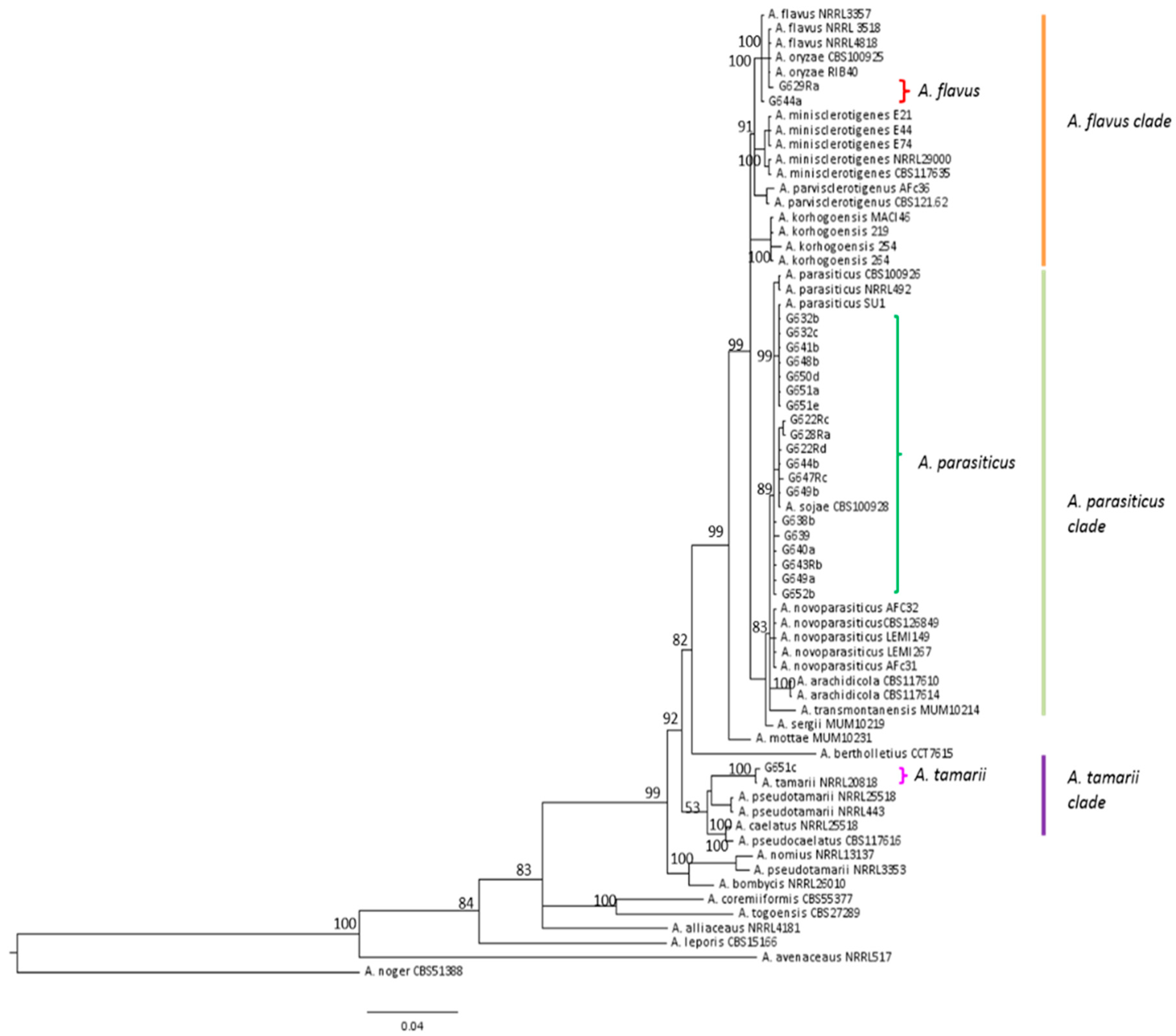
2.4. Characterization of Aspergillus Section Flavi Strains
3. Discussion
3.1. Impact of Climate on Aflatoxin Emergence in French Maize in 2015
3.2. Nature of Aspergillus Section Flavi Responsible for Aflatoxin Contamination of French Maize
4. Conclusions
5. Materials and Methods
5.1. Meteorological Data
5.2. Sampling
5.2.1. Sampling Strategy
5.2.2. Sample Collection
Field Samples
Silo Samples
5.2.3. Sample Preparation for Analysis
5.3. Aflatoxin and Cyclopiazonic Acid Quantification
5.4. Fungal Count and Identification
5.5. Characterization of Aspergillus Section Flavi
5.5.1. Morphological Identification
5.5.2. Molecular Identification
5.5.3. Mycotoxigenic Potential of Isolates
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- IARC Monographs on the evaluation of carcinogenic risks to humans. Aflatoxins 2012, 100F, 225–244.
- Wu, F.; Groopman, J.D.; Pestka, J.J. Public health impacts of foodborne mycotoxins. Annu. Rev. Food. Sci. Technol. 2014, 5, 351–372. [Google Scholar] [CrossRef] [PubMed]
- Meissonnier, G.M.; Pinton, P.; Laffitte, J.; Cossalter, A.M.; Gong, Y.Y.; Wild, C.P.; Bertin, G.; Galtier, P.; Oswald, I.P. Immunotoxicity of aflatoxin B1: Impairment of the cell-mediated response to vaccine antigen and modulation of cytokine expression. Toxicol. Appl. Pharmacol. 2008, 231, 142–149. [Google Scholar] [CrossRef] [PubMed]
- Khlangwiset, P.; Shephard, G.S.; Wu, F. Aflatoxins and growth impairment: A review. Crit. Rev. Toxicol. 2011, 41, 740–755. [Google Scholar] [CrossRef] [PubMed]
- Gibb, H.; Devleesschauwer, B.; Bolger, B.M.; Wu, F.; Ezenman, J.; Cliff, J.; Zeilmaker, M.; Verger, P.; Pitt, J.; Baines, J. World Health Organization estimates of the global and regional disease burden of four foodborne chemical toxins, 2010: A data synthesis. F1000Research 2015, 4, 1393. [Google Scholar] [CrossRef] [PubMed]
- Pildain, M.B.; Frisvad, J.C.; Vaamonde, G.; Cabral, D.; Varga, J.; Samson, R.A. Two novel aflatoxin-producing Aspergillus species from Argentinean peanuts. Int. J. Syst. Evol. Microbiol. 2008, 58, 725–735. [Google Scholar] [CrossRef]
- Varga, J.; Frisvad, J.C.; Samson, R.A. Two new aflatoxin producing species and an overview of Aspergillus section. Flavi. Stud. Mycol. 2011, 69, 57–80. [Google Scholar] [CrossRef] [Green Version]
- Soares, C.; Rodrigues, P.; Peterson, S.W.; Lima, N.; Venancio, A. Three new species of Aspergillus section Flavi isolated from almonds and maize in Portugal. Mycologia 2012, 104, 682–697. [Google Scholar] [CrossRef]
- Taniwaki, M.H.; Pitt, J.I.; Iamanaka, B.T.; Sartori, D.; Copetti, M.V.; Balajee, A.; Fungaro, M.H.; Frisvad, J.C. Aspergillus bertholletius sp. nov. from Brazil nuts. PLoS ONE 2012, 7, e42480. [Google Scholar] [CrossRef] [Green Version]
- Carvajal-Campos, A.; Manizan, A.L.; Tadrist, S.; Koffi-Akaki, D.; Koffi-Nevry, R.; Moore, G.G.; Fapohunda, S.O.; Bailly, S.; Montet, D.; Oswald, I.P.; et al. Aspergillus korhogoensis, a novel aflatoxin producing species from the Côte d’Ivoire. Toxins 2017, 9, 353. [Google Scholar] [CrossRef]
- Frisvad, J.C.; Hubka, V.; Ezekiel, C.N.; Hong, S.B.; Novakova, A.; Chen, A.J.; Arzanlou, M.; Larsen, T.O.; Sklenar, F.; Mahakarnchanakul, W.; et al. Taxonomy of Aspergillus section Flavi and their production of aflatoxins, ochratoxins and other mycotoxins. Stud. Mycol. 2019, 93, 1–63. [Google Scholar] [CrossRef] [PubMed]
- Uka, V.; Moore, G.G.; Aroyo-Manzanares, N.; Nebija, D.; De Saeger, S.; Di Mavungu, J.D. Unravelling the diversity of the cyclopiazonic acid family of mycotoxins in Aspergillus flavus by UHPLC triple TOF HRMS. Toxins 2017, 9, 35. [Google Scholar] [CrossRef] [PubMed]
- Klich, M.A. Aspergillus flavus: The major producer of aflatoxin. Mol. Plant. Pathol. 2007, 8, 713–722. [Google Scholar] [CrossRef] [PubMed]
- Pitt, J.I.; Hocking, A.D. Fungi and Food Spoilage, 3rd ed.; Springer: London, UK, 2009; p. 519. ISBN 978-0-387-92206-5. [Google Scholar]
- Santin, E. Mould growth and mycotoxin production. In Mycotoxin Blue Book; Diaz, D.E., Ed.; Nottingham University Press: Nottingham, UK, 2005; pp. 225–234. ISBN 978-1-904761-19-8. [Google Scholar]
- Rustom, I.Y. Aflatoxin in food and feed: Occurrence, legislation and inactivation by physical methods. Food Chem. 1997, 59, 57–67. [Google Scholar] [CrossRef]
- Sharma, U.P.; Bhetaria, P.J.; Devi, P.; Varma, A. Aflatoxins: Implication on health. Ind. J. Biochem. 2017, 32, 124–133. [Google Scholar] [CrossRef] [PubMed]
- European Food Safety Authority. Opinion of the Scientific Panel on Contaminants in the Food Chain on a request from the Commission related to Aflatoxin B1 as undesirable substance in animal feed. EFSA J. 2004, 39, 1–27. [Google Scholar]
- European Food Safety Authority. Opinion of the Scientific Panel on Contaminants in the Food Chain on a request from the Commission related to the potential increase of consumer health risk by a possible increase of the existing maximum levels for aflatoxins in almonds, hazelnuts and pistachios and derived products. EFSA J. 2007, 446, 1–127. [Google Scholar]
- Tabuc, C.; Marin, D.; Guerre, P.; Sesan, T.; Bailly, J.D. Molds and mycotoxin content of cereals in southeastern Romania. J. Food Prot. 2009, 72, 662–665. [Google Scholar] [CrossRef]
- Covarelli, L.; Beccari, G.; Salvi, S. Infection by mycotoxigenic fungal species and mycotoxin contamination of maize grain in Umbria, central Italy. Food Chem. Toxicol. 2011, 49, 2365–2369. [Google Scholar] [CrossRef]
- Amorini, S.; Altafini, A.; Zaghini, A.; Roncada, P. Occurrence of aflatoxin B1 in conventional and organic flour in Italy and the role of sampling. Food Control. 2015, 50, 858–863. [Google Scholar] [CrossRef]
- Alborch, L.; Bragulat, M.R.; Castella, G.; Abarca, M.L.; Cabanes, F.J. Mycobiota and mycotoxin contamination of maize flours and popcorn kernels for human consumption commercialized in Spain. Food Microbiol. 2012, 32, 97–103. [Google Scholar] [CrossRef] [PubMed]
- Pleadin, J.; Vulic, A.; Persi, N.; Skrivanko, M.; Capek, B.; Cvetnic, Z. Aflatoxin B1 occurrence in maize sampled from Croatian farms and feed factories during 2013. Food Control. 2014, 40, 286–291. [Google Scholar] [CrossRef]
- Pleadin, J.; Vulic, A.; Persi, N.; Skrivanko, M.; Capek, B.; Cvetnic, Z. Annual and regional variations of aflatoxin B1 levels seen in grains and feed coming from Croatian dairy farms over a 5-year period. Food Control. 2015, 47, 221–225. [Google Scholar] [CrossRef]
- Kos, J.; Skrinjar, M.M.; Mandic, A.I.; Misan, A.C.; Bursic, V.P.; Saric, B.M.; Janic-Hajnal, E.P. Presence of aflatoxins in cereals from Serbia. Food Feed. Res. 2014, 41, 31–38. [Google Scholar] [CrossRef] [Green Version]
- Ketney, O.; Santini, A.; Oancea, S. Recent aflatoxin survey data in milk products: A review. Int. J. Dairy Technol. 2017, 70, 320–331. [Google Scholar] [CrossRef]
- Kos, J.; Levic, J.; Duragic, O.; Kokic, B.; Miladinovic, I. Occurrence and estimation of aflatoxin M1 exposure un milk in Serbia. Food Control. 2014, 38, 41–46. [Google Scholar] [CrossRef]
- Skrbic, B.; Zivancev, J.; Antic, I.; Godula, M. Levels of aflatoxin M1 in different types of milk collected in Serbia: Assessment of human and animal exposure. Food Control. 2014, 40, 113–119. [Google Scholar] [CrossRef]
- Tomasevic, I.; Petrovic, J.; Jovetic, M.; Raicevic, S.; Milojevic, M.; Miocinovic, J. Two year survey on the occurrence and seasonal variation of aflatoxin M1 in milk and milk products in Serbia. Food Control. 2015, 56, 64–70. [Google Scholar] [CrossRef]
- European Union. Commission regulation 1881/2006 setting maximum levels for certain contaminants in foodstuffs. Off. J. Eur. Union 2006, L364, 5–24. [Google Scholar]
- Mikusova, P.; Sulyok, M.; Santini, A.; Srobarova, A. Aspergillus spp and their secondary metabolite production in grape berries from Slovakia. Phytopathol. Mediterr. 2014, 53, 109–114. [Google Scholar] [CrossRef]
- Mikusova, P.; Ritieni, A.; Santini, A.; Juhasova, G.; Srobarova, A. Contamination by moulds of grape berries in Slovakia. Food Addit. Contam. 2010, 27, 738–747. [Google Scholar] [CrossRef] [PubMed]
- Udovicki, B.; Audenaert, K.; De Saeger, S.; Rajkovic, A. Overview on the mycotoxin incidence in Serbia in the period 2004-2016. Toxins 2018, 10, 279. [Google Scholar] [CrossRef] [PubMed]
- Medina, A.; Rodriguez, A.; Magan, N. Effect of climate change on Aspergillus flavus and aflatoxin B1 production. Front. Microbiol. 2014, 5, 348. [Google Scholar] [CrossRef] [PubMed]
- Battilani, P.; Toscano, P.; Van der Fels-Klerx, H.J.; Moretti, A.; Carmado Leggieri, M.; Brera, C.; Rortais, A.; Goumperis, T.; Robinson, T. Aflatoxin B1 contamination in maize in Europe increases due to climate change. Sci. Rep. 2016, 6, 24328. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- European Commission. RAFF—Food and Feed Safety Alerts. Available online: https://ec.europa.eu/food/safety/rasff_en (accessed on 6 December 2018).
- Janic Hajnal, E.; Kos, J.; Krulj, J.; Krstovic, S.; Jajic, V.; Pezo, L.; Saric, B.; Nedeljkovic, N. Aflatoxins contamination of maize in Serbia: The impact of weather conditions in 2015. Food Addit. Contam. Part A 2017, 34, 1999–2010. [Google Scholar] [CrossRef] [PubMed]
- Payne, G.A. Process of contamination by aflatoxin-producing fungi and their impact on crops. In Mycotoxins in Agriculture and Food Safety; Sinha, K.K., Bhatnagar, D., Eds.; Marcel Dekker Inc.: New York, NY, USA, 1998; pp. 279–300. [Google Scholar]
- Cotty, P.J.; Jaime-Garcia, R. Influences of climate change on aflatoxin producing fungi and aflatoxin contamination. Int. J. Food Microbiol. 2007, 119, 109–115. [Google Scholar] [CrossRef] [PubMed]
- Le Bras, A.; Niquet, G. Le nettoyage des grains devrait prendre du poids. Persp. Agric. 2006, 327, 22–24. [Google Scholar]
- Battilani, P.; Barbano, C.; Piva, G. Aflatoxin B1 contamination in maize related to the aridity index in North Italy. World Mycotoxin J. 2008, 1, 449–456. [Google Scholar] [CrossRef]
- Martins, M.L.; Martins, H.M. Natural and in vitro coproduction of cyclopiazonic acid and aflatoxins. J. Food Protect. 1999, 62, 292–294. [Google Scholar] [CrossRef]
- Fernandez Pinto, V.; Patriarca, A.; Locani, O.; Vaamonde, G. Natural co-occurrence of aflatoxin and cyclopiazonic acid in peanuts grown in Argentina. Food Addit. Contam. 2001, 18, 1017–1020. [Google Scholar] [CrossRef]
- Alassane-Kpembi, I.; Schatzmayr, G.; Marin, D.; Taranu, I.; Puel, O.; Oswald, I.P. Mycotoxins co-contamination: Methodological aspects and biological relevance of combined toxicity studies. Crit. Rev. Food Sci. 2017, 16, 3489–3507. [Google Scholar] [CrossRef] [PubMed]
- Diao, E.; Dong, H.; Hou, H.; Zhang, Z.; Ji, N.; Ma, W. Factors influencing aflatoxin contamination in before and after harvest peanuts: A review. J. Food Res. 2015, 4, 148–154. [Google Scholar] [CrossRef]
- Atehnkeng, J.; Ojiambo, P.S.; Ikotun, T.; Sikora, R.A.; Cotty, P.J.; Bandyopadhay, R. Evaluation of atoxigenic isolates of Aspergillus flavus as potential biocontrol agents for aflatoxin in maize. Food Addit. Contam. 2008, 25, 1264–1271. [Google Scholar] [CrossRef] [PubMed]
- Perrone, G.; Haidukowski, M.; Stea, G.; Epifani, F.; Bandyopadhyay, R.; Leslie, J.F.; Logrieco, A. Population structure and aflatoxin production by Aspergillus sect. Flavi from maize in Nigeria and Ghana. Food Microbiol. 2014, 41, 52–59. [Google Scholar] [CrossRef] [PubMed]
- Horn, B.W. Ecology and population biology of aflatoxigenic fungi in soil. In Aflatoxin and Food Safety; Abbas, H.K., Ed.; Taylor & Francis: Boca Raton, FL, USA, 2005; pp. 95–116. [Google Scholar]
- Mauro, A.; Battilani, P.; Callicott, A.; Giorni, P.; Pietri, A.; Cotty, P. Structure pf an Aspergillus flavus population from maize kernels in northern Italy. Int. J. Food Microbiol. 2013, 162, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Giorni, P.; Magan, N.; Pietri, A.; Bertuzzi, T.; Battilani, P. Studies on Aspergillus section Flavi isolated from maize in northern Italy. Int. J. Food Microbiol. 2007, 113, 330–338. [Google Scholar] [CrossRef] [PubMed]
- Horn, B.W. Biodiversity of Aspergillus section Flavi in the United states: A review. Food Addit. Contam. 2007, 24, 1088–1101. [Google Scholar] [CrossRef]
- Sarrocco, S.; Vannacci, G. Preharvest application of beneficial fungi as a strategy to prevent postharvest mycotoxin contamination: A review. Crop Protect. 2018, 110, 160–170. [Google Scholar] [CrossRef]
- Ehrlich, K.C. Non-aflatoxigenic Aspergillus flavus to prevent aflatoxin contamination in crops: Advantages and limitation. Front. Microbiol. 2014, 5, 50. [Google Scholar] [CrossRef]
- Deudon, O.; Le Bris, X.; Piraux, F. Interest and implementation of a spatialization method of meteorological data used in Agricultural Decision Support Tools. In Proceedings of the 2017 EFITA Congress, Montpellier, France, 2–6 July 2017. [Google Scholar]
- International Organisation for Standardization. ISO 7954:1987. Microbiology, general guidance for enumeration of yeasts and moulds—Colony count technique at 25 °C; ISO, Ed.; International Organisation for Standardization: Geneva, Switzerland, 1987. [Google Scholar]
- Samson, R.A.; Houbraken, J.; Thrane, U.; Frisvad, J.C.; Andersen, B. Food and Indoor Fungi; CBS Knaw: Uthrecht, The Netherlands, 2010. [Google Scholar]
- Pitt, J.I.; Hocking, A.D.; Glenn, D.R. An improved medium for detection of Aspergillus flavus and A. Parasiticus. J. Appl. Bacteriol. 1983, 54, 109–114. [Google Scholar] [CrossRef]
- Adjovi, Y.; Bailly, S.; Gnonlonfin, B.J.G.; Tadrist, S.; Querin, A.; Sanni, A.; Oswald, I.P.; Puel, O.; Bailly, J.D. Contrast between natural occurrence of toxigenic Aspergillii of the Flavi section and Aflatoxin B1 in cassava: Possible explanation. Food Microbiol. 2014, 38, 151–159. [Google Scholar] [CrossRef] [PubMed]
- BioEdit v7.0.5. Available online: http://www.mbio.ncsu.edu/BioEdit/page2.html (accessed on 15 November 2018).
- Maddison, W.P.; Maddison, D.R. Mesquite: A Modular System for Evolutionary Analysis. Version 3.2. 2017. Available online: http://mesquiteproject.org (accessed on 15 November 2018).
- Lanfear, R.; Frandsen, P.B.; Wright, A.M.; Senfeld, T.; Calcott, B. PartitionFinder 2: New methods for selecting partitioned models of evolution for molecular and morphological phylogenetic analyses. Mol. Biol. Evol. 2017, 34, 772–773. [Google Scholar] [CrossRef] [PubMed]
- Ronquist, F.; Teslenko, M.; van der Mark, P.; Ayres, D.L.; Darling, A.; Höhna, S.; Larget, B.; Liu, L.; Suchard, M.A.; Huelsenbeck, J.P. MrBayes 3.2: Efficient Bayesian Phylogenetic Inference and Model Choice Across a Large Model Space. Syst. Biol. 2012, 61, 539–542. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rambaut, A.; Suchard, M.A.; Xie, D.; Drummond, A.J. Tracer v1.6. 2014. Available online: http://tree.bio.ed.ac.uk/software/tracer (accessed on 15 November 2018).
- Rambaut, A. FigTree v1.4.2, A Graphical Viewer of Phylogenetic Trees. 2014. Available online: http://tree.bio.ed.ac.uk/software/figtree/ (accessed on 15 November 2018).
- El Mahgubi, A.; Puel, O.; Bailly, S.; Tadrist, S.; Querin, A.; Ouadia, A.; Oswald, I.P.; Bailly, J.D. Distribution and toxigenicity of Aspergillus section Flavi in spices marketed in Morocco. Food Cont. 2013, 32, 143–148. [Google Scholar] [CrossRef]
- Caceres, I.; El Khoury, R.; Medina, A.; Lippi, Y.; Naylis, C.; Atoui, A.; El Khoury, A.; Oswald, I.P.; Bailly, J.D.; Puel, O. Deciphering the anti-aflatoxinogenic properties of eugenol using a large scale q-PCR approach. Toxins 2016, 8, 123. [Google Scholar] [CrossRef] [PubMed] [Green Version]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Origin | Contaminated Samples (%) | Aflatoxin Content (µg/kg) | ||||
|---|---|---|---|---|---|---|
| AFB1 | AFB2 | AFG1 | AFG2 | Total AFs | ||
| 1.1 | ND | 0.4 | ND | 1.5 | ||
| 0.1 | ND | 0.2 | ND | 0.3 | ||
| 4.0 | 0.4 | 9.7 | 1.2 | 15.3 | ||
| FIELD | 7 samples/118 | 0.3 | ND | 2.0 | 0.3 | 2.6 |
| 6% | 0.7 | ND | 0.5 | ND | 1.2 | |
| 20.4 | 2.1 | 24.8 | 3.5 | 50.8 | ||
| 66.0 | 3.1 | 0.9 | ND | 70 | ||
| 0.1 | ND | 0.7 | ND | 0.8 | ||
| 3.2 | 0.2 | 4.5 | ND | 7.9 | ||
| 0.6 | ND | ND | ND | 0.6 | ||
| 3.3 | 0.2 | ND | ND | 3.5 | ||
| 2.4 | 0.2 | ND | ND | 2.6 | ||
| SILOS | 12 samples/81 | 1.4 | ND | 0.1 | ND | 1.5 |
| 15% | 3.8 | 0.2 | 0.8 | ND | 4.8 | |
| 0.6 | ND | ND | ND | 0.6 | ||
| 1.9 | 0.2 | 3.9 | 0.6 | 6.6 | ||
| 7.2 | 0.3 | 5.6 | 0.2 | 13.3 | ||
| 0.4 | ND | 0.1 | ND | 0.5 | ||
| 0.7 | ND | ND | ND | 0.7 | ||
| Mycoflora | Mean Fungal Load (CFU/g) (Mini–Maxi) % Contaminated Samples | ||
|---|---|---|---|
| AF− Field Samples (n = 24) | AF+ Field Samples (n = 7) | AF+ Silo Samples (n = 12) | |
| Total flora | 2.3 × 105 | 7.9 × 105 | 2 × 105 |
| (1.2 × 104–1.2 × 106) | (6.9 × 104–2.5 × 106) | (1.4 × 104–7.7 × 105) | |
| Fusarium sp. | 1.6 × 105 | 3.5 × 105 | 1.7 × 105 |
| (103–5 × 105) | (104–2 × 106) | (104–7 × 105) | |
| 100% | 100% | 100% | |
| Penicillium sp. | 3.5 × 104 | 3.2 × 105 | 6.1 × 103 |
| (2 × 102–4 × 105) | (5 × 103–2 × 106) | (5 × 102–3 × 104) | |
| 100% | 100% | 100% | |
| Acremonium sp. | 3.3 × 104 | 5.7 × 104 | 1.1 × 104 |
| (0–3 × 105) | (0–2 × 105) | (0–3 × 104) | |
| 67% | 86% | 58% | |
| Cladosporium sp. | 2.2 × 103 | 4.2 × 103 | 1.3 × 102 |
| (0–104) | (0–2 × 104) | (0–103) | |
| 79% | 86% | 25% | |
| Aspergillus sp. | 2.2 × 103 | 5.3 × 104 | 9.2 × 103 |
| (0–2 × 104) | (2 × 103–1.7 × 105) | (102–4 × 104) | |
| 54% | 100% | 100% | |
| Aspergillus section Nigri | 103 | 2.1 × 104 | 5.8 × 102 |
| (0–2 × 104) | (102–7 × 104) | (0–4 × 103) | |
| 46% | 100% | 83% | |
| Aspergillus section Flavi | 9.9 × 102 | 3.2 × 104 | 6.2 × 103 |
| (0–2 × 104) | (6 × 102–105) | (102–3 × 104) | |
| 38% | 100% | 100% | |
| Gene | Gene Name | Length (bp) | Primers | Sequence (Nucleotides: 5’→3’) | |
|---|---|---|---|---|---|
| Forward | Reverse | ||||
| ITS (4–5) | Internal transcribed spacer | 300–330 | ITS5 | 5′-GGAAGTAAAAGTCGTAACAAGG | |
| ITS 4 | 5′-TCCTCCGCTTATTGATATGC | ||||
| benA-2 | ß-tubulin | 1125 | benA 2a | 5′-GGTAACCAAATCGGTGCTGC | |
| benA 2b | 5′-ACCCTCAGTGTAGTGACCCTTGGC | ||||
| cmdA | Calmodulin | 543 | Cmd5 | 5′-CCGAGTACAAGGAGGCCTTC-3’ | |
| Cmd6 | 5′-CCGATAGAGGTCATAACGTGG-3’ | ||||
| Species and Strain Number | Accession Number | ||
|---|---|---|---|
| A. parasiticus | ITS | cmdA | benA |
| G622Rc | MK165710 | MK165730 | MK172077 |
| G622Rd | MK165711 | MK165731 | MK172078 |
| G628Ra | MK165712 | MK165732 | MK172079 |
| G632b | MK165713 | MK165733 | MK172080 |
| G632c | MK165714 | MK165734 | MK172081 |
| G638b | MK165709 | MK165735 | MK172082 |
| G639 | MK165726 | MK165736 | MK172083 |
| G640a | MK165715 | MK165737 | MK172084 |
| G641b | MK165716 | MK165738 | MK172085 |
| G643Rb | MK165717 | MK165739 | MK172086 |
| G644b | MK165718 | MK165740 | MK172087 |
| G647Rc | MK165719 | MK165741 | MK172088 |
| G648b | MK165720 | MK172072 | MK172089 |
| G649a | MK165708 | MK165742 | MK172090 |
| G649b | MK165721 | MK165743 | MK172091 |
| G650d | MK165722 | MK165744 | MK172092 |
| G651a | MK165723 | MK165745 | MK172093 |
| G651e | MK165724 | MK172073 | MK172094 |
| G652b | MK165725 | MK172096 | MK172095 |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bailly, S.; Mahgubi, A.E.; Carvajal-Campos, A.; Lorber, S.; Puel, O.; Oswald, I.P.; Bailly, J.-D.; Orlando, B. Occurrence and Identification of Aspergillus Section Flavi in the Context of the Emergence of Aflatoxins in French Maize. Toxins 2018, 10, 525. https://0-doi-org.brum.beds.ac.uk/10.3390/toxins10120525
Bailly S, Mahgubi AE, Carvajal-Campos A, Lorber S, Puel O, Oswald IP, Bailly J-D, Orlando B. Occurrence and Identification of Aspergillus Section Flavi in the Context of the Emergence of Aflatoxins in French Maize. Toxins. 2018; 10(12):525. https://0-doi-org.brum.beds.ac.uk/10.3390/toxins10120525
Chicago/Turabian StyleBailly, Sylviane, Anwar El Mahgubi, Amaranta Carvajal-Campos, Sophie Lorber, Olivier Puel, Isabelle P. Oswald, Jean-Denis Bailly, and Béatrice Orlando. 2018. "Occurrence and Identification of Aspergillus Section Flavi in the Context of the Emergence of Aflatoxins in French Maize" Toxins 10, no. 12: 525. https://0-doi-org.brum.beds.ac.uk/10.3390/toxins10120525