Clinical S. aureus Isolates Vary in Their Virulence to Promote Adaptation to the Host

 ,
, 
 , ,
, , 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
2.1. The Genetic Analysis of Staphylococcal Isolates Reveals Only Minor Differences in Bacterial Origin
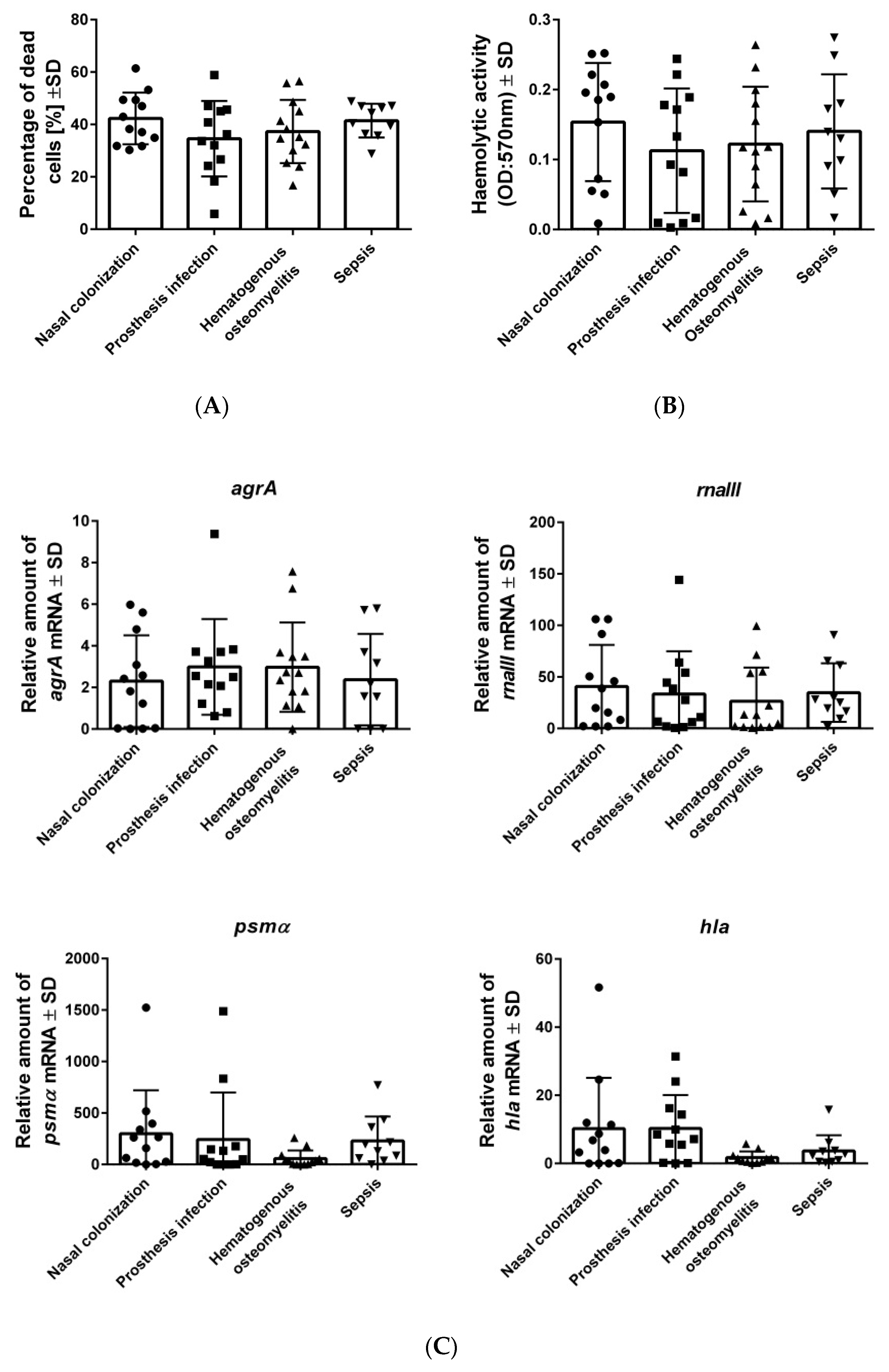
2.2. High- as Well as Low-Cytotoxicity Strains are Equally Distributed among Isolates from Different Bacterial Origins
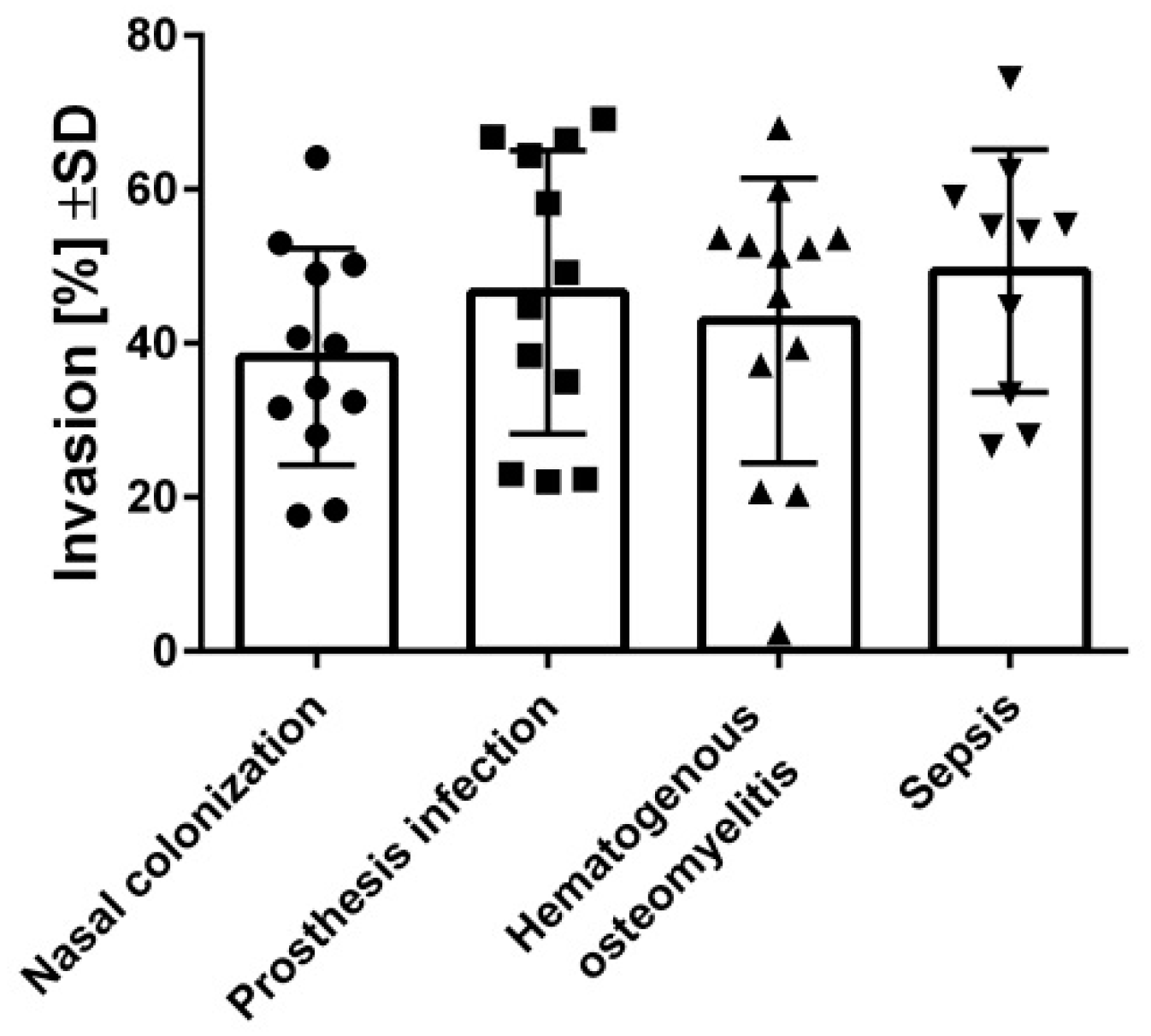
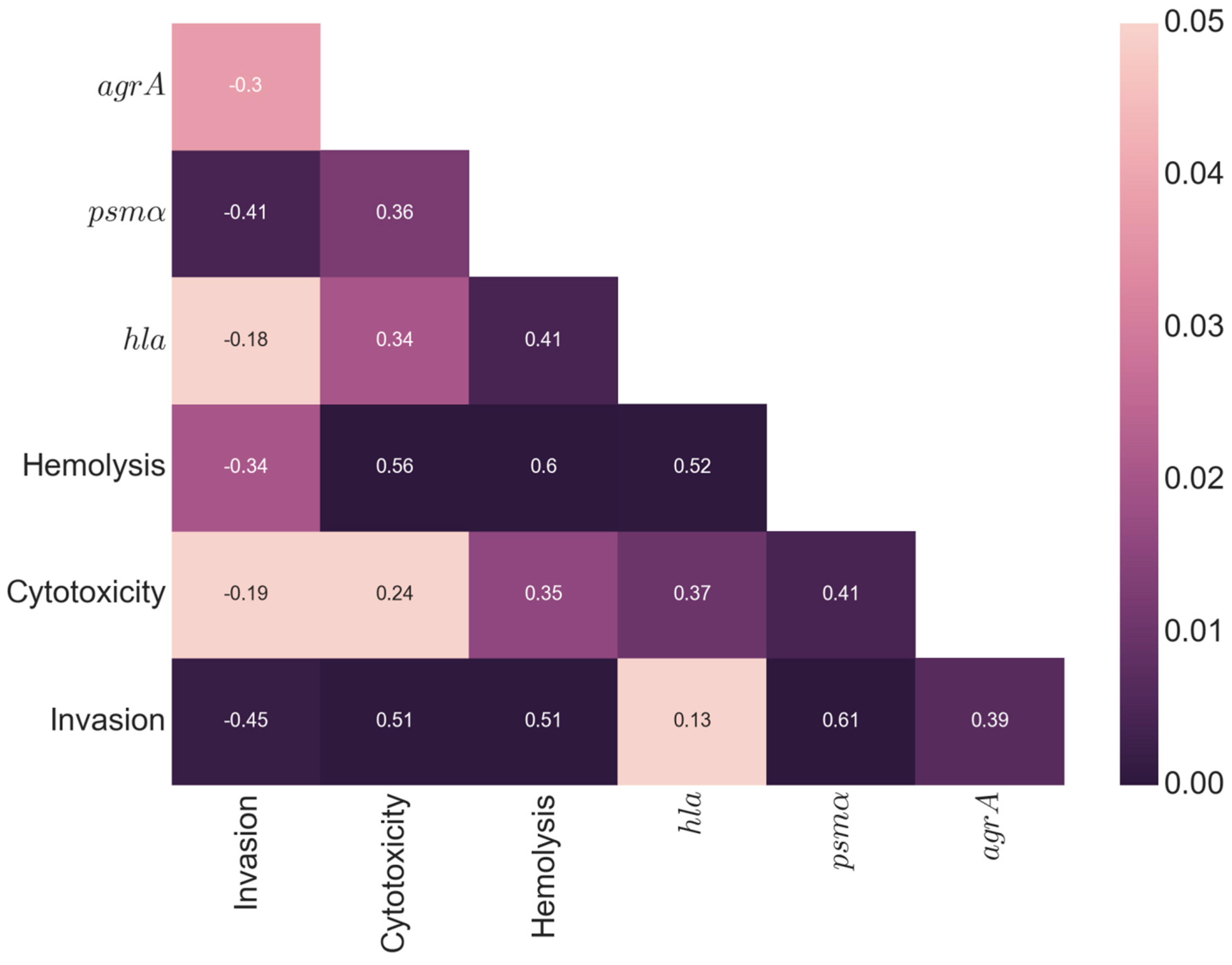
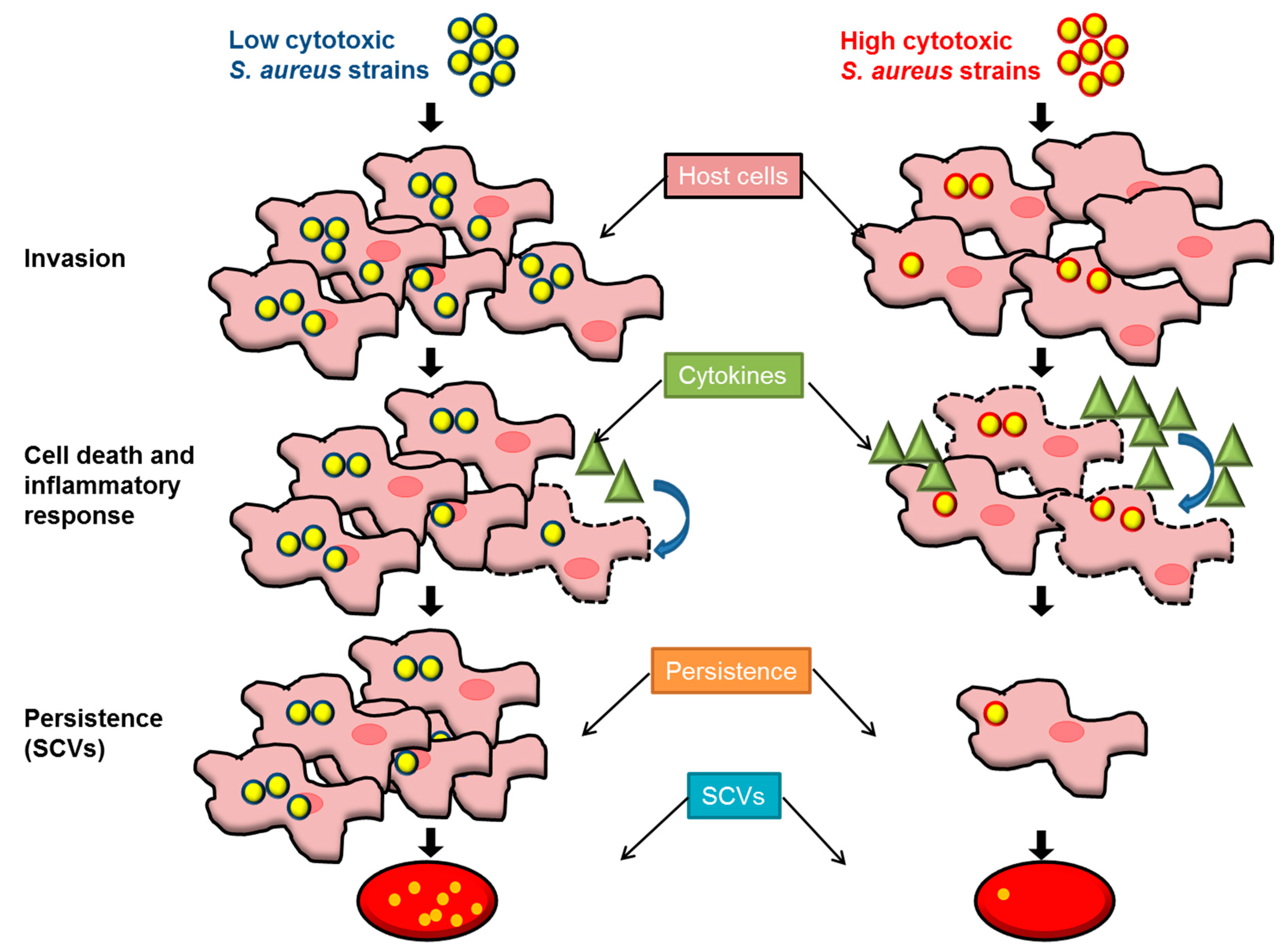
2.3. Highly Invasive but Low-Cytotoxicity Strains Persist at High Numbers within Host Cells
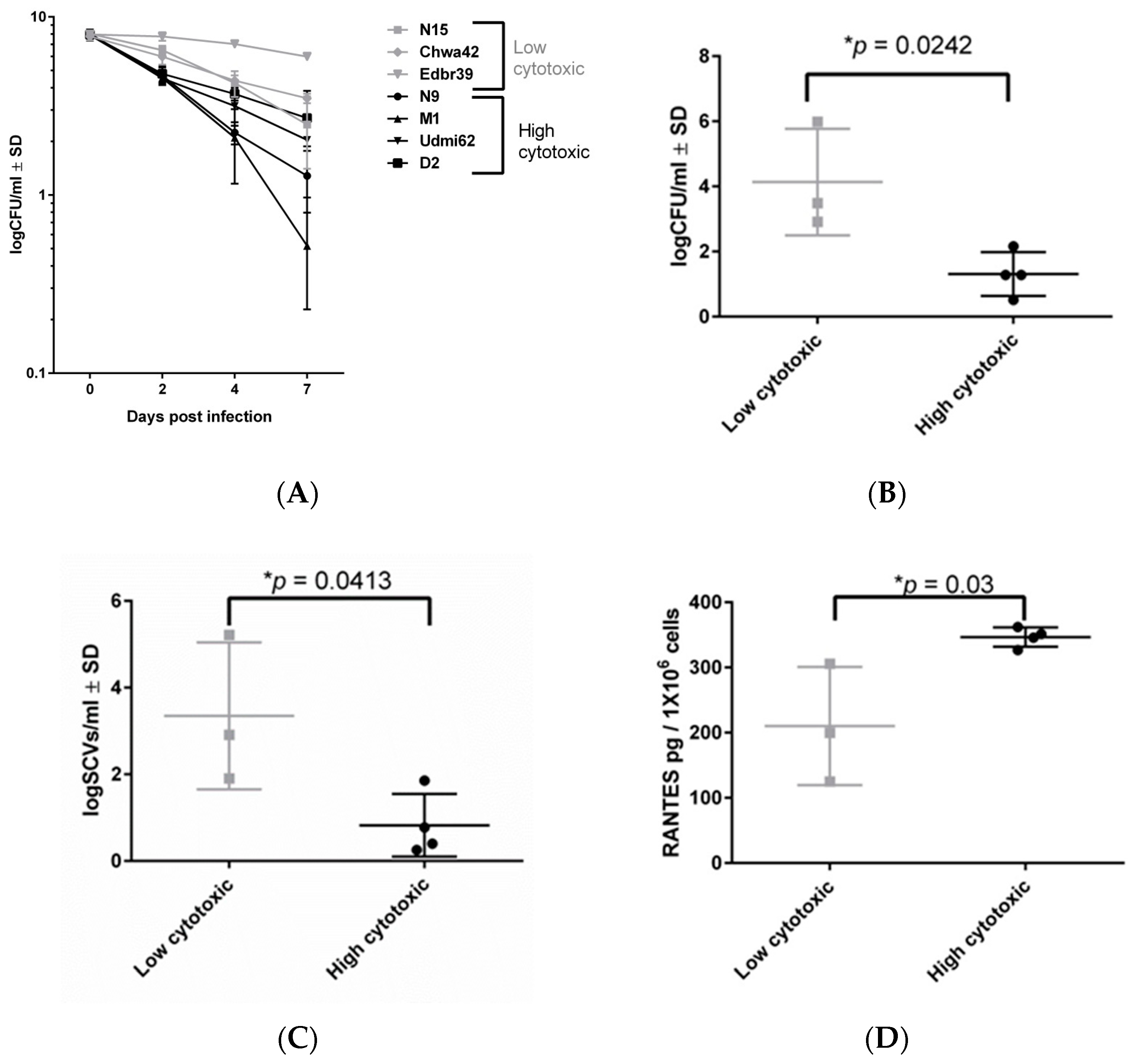
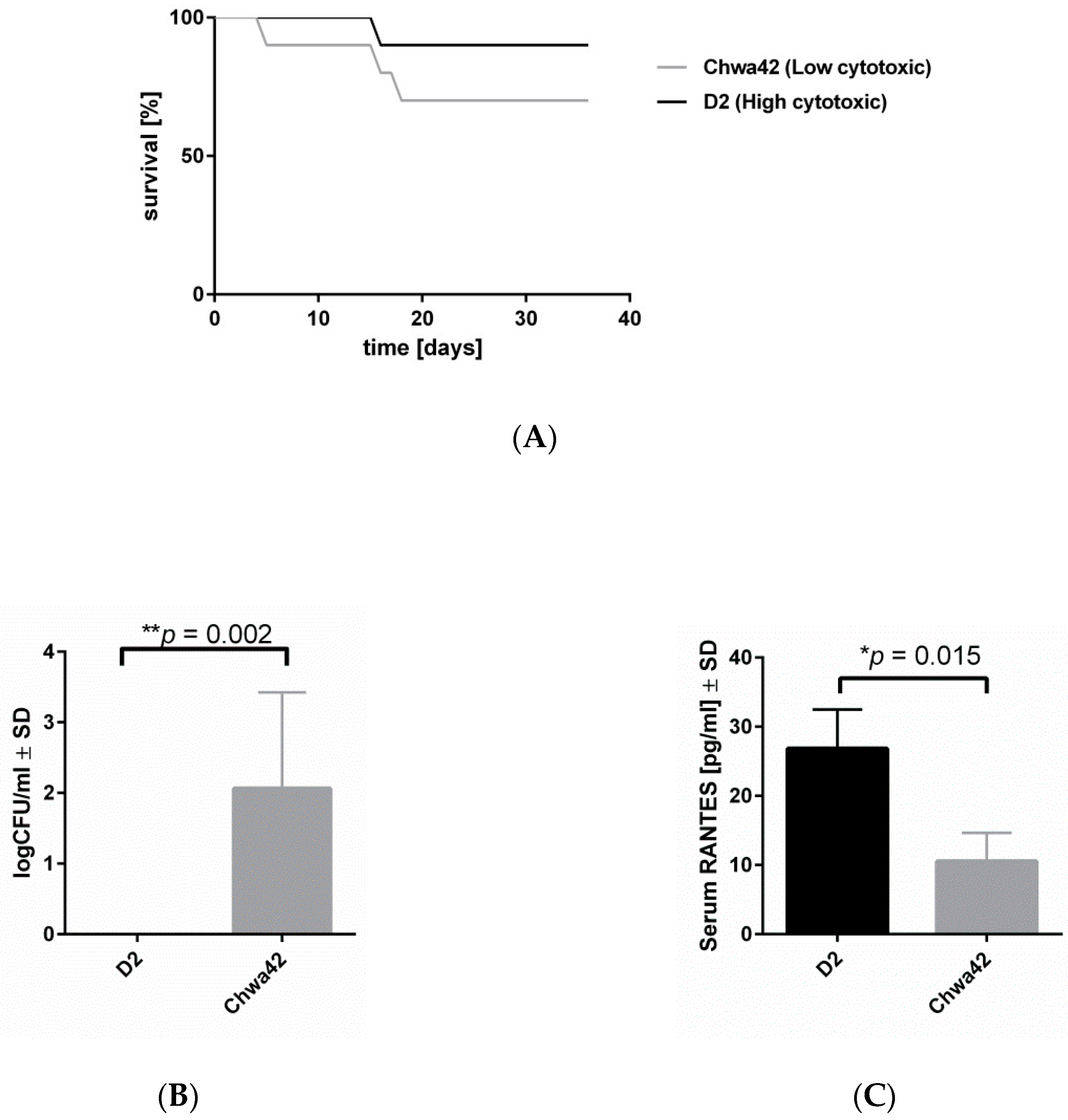
2.4. Low-Cytotoxicity Strains can Persist in High Numbers in a Murine Sepsis Model
3. Discussion
4. Material and Methods
4.1. Collection of S. aureus Isolates and Definitions
4.2. Genotypic Characterization
4.3. Growth Curve and Generation Time
4.4. Measurement of Haemolysis
4.5. Cell Death and Invasion Assay in Osteoblasts
4.6. Long-Term Persistence
4.7. RNA Isolation and Real-Time PCR
4.8. Murine Sepsis Model
4.9. Release of the Chemokine RANTES
4.10. Ethical Permissions
4.11. Statistics
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Kluytmans, J.; van Belkum, A.; Verbrugh, H. Nasal carriage of Staphylococcus aureus: Epidemiology, underlying mechanisms, and associated risks. Clin. Microbiol. Rev. 1997, 10, 505–520. [Google Scholar] [CrossRef] [PubMed]
- Abu-Qatouseh, L.F.; Chinni, S.V.; Seggewiss, J.; Proctor, R.A.; Brosius, J.; Rozhdestvensky, T.S.; Peters, G.; von Eiff, C.; Becker, K. Identification of differentially expressed small non-protein-coding RNAs in Staphylococcus aureus displaying both the normal and the small-colony variant phenotype. J. Mol. Med. 2010, 88, 565–575. [Google Scholar] [CrossRef] [PubMed]
- Lowy, F.D. Medical progress—Staphylococcus aureus infections. N. Engl. J. Med. 1998, 339, 520–532. [Google Scholar] [CrossRef] [PubMed]
- Von Eiff, C.; Becker, K.; Machka, K.; Stammer, H.; Peters, G. Nasal carriage as a source of Staphylococcus aureus bacteremia. Study Group. N. Engl. J. Med. 2001, 344, 11–16. [Google Scholar] [CrossRef] [PubMed]
- Brown, S.P.; Cornforth, D.M.; Mideo, N. Evolution of virulence in opportunistic pathogens: Generalism, plasticity, and control. Trends Microbiol. 2012, 20, 336–342. [Google Scholar] [CrossRef] [PubMed]
- Tuchscherr, L.; Bischoff, M.; Lattar, S.M.; Noto Llana, M.; Pfortner, H.; Niemann, S.; Geraci, J.; Van de Vyver, H.; Fraunholz, M.J.; Cheung, A.L.; et al. Sigma Factor SigB Is Crucial to Mediate Staphylococcus aureus Adaptation during Chronic Infections. PLoS Pathog. 2015, 11, e1004870. [Google Scholar] [CrossRef] [PubMed]
- Bischoff, M.; Dunman, P.; Kormanec, J.; Macapagal, D.; Murphy, E.; Mounts, W.; Berger-Bachi, B.; Projan, S. Microarray-based analysis of the Staphylococcus aureus sigma(B) regulon. J. Bacteriol. 2004, 186, 4085–4099. [Google Scholar] [CrossRef] [PubMed]
- Novick, R.P.; Geisinger, E. Quorum Sensing in Staphylococci. Annu. Rev. Genet. 2008, 42, 541–564. [Google Scholar] [CrossRef] [PubMed]
- Sonnen, A.F.P.; Henneke, P. Role of Pore-Forming Toxins in Neonatal Sepsis. Clin. Dev. Immunol. 2013. [Google Scholar] [CrossRef] [PubMed]
- Foster, T.J.; Geoghegan, J.A.; Ganesh, V.K.; Hook, M. Adhesion, invasion and evasion: The many functions of the surface proteins of Staphylococcus aureus. Nat. Rev. Microbiol. 2014, 12, 49–62. [Google Scholar] [CrossRef] [PubMed]
- Proctor, R.A.; von Eiff, C.; Kahl, B.C.; Becker, K.; McNamara, P.; Herrmann, M.; Peters, G. Small colony variants: A pathogenic form of bacteria that facilitates persistent and recurrent infections. Nat. Rev. Microbiol. 2006, 4, 295–305. [Google Scholar] [CrossRef] [PubMed]
- Tuchscherr, L.; Heitmann, V.; Hussain, M.; Viemann, D.; Roth, J.; von Eiff, C.; Peters, G.; Becker, K.; Loffler, B. Staphylococcus aureus Small-Colony Variants Are Adapted Phenotypes for Intracellular Persistence. J. Infect. Dis. 2010, 202, 1031–1040. [Google Scholar] [CrossRef] [PubMed]
- Tuchscherr, L.; Geraci, J.; Loffler, B. Staphylococcus aureus Regulator Sigma B is Important to Develop Chronic Infections in Hematogenous Murine Osteomyelitis Model. Pathogens 2017, 6, 31. [Google Scholar] [CrossRef] [PubMed]
- Cheung, A.L.; Eberhardt, K.J.; Chung, E.; Yeaman, M.R.; Sullam, P.M.; Ramos, M.; Bayer, A.S. Diminished virulence of a sar-/agr- mutant of Staphylococcus aureus in the rabbit model of endocarditis. J. Clin. Investig. 1994, 94, 1815–1822. [Google Scholar] [CrossRef] [PubMed]
- Seidl, K.; Bayer, A.S.; McKinnell, J.A.; Ellison, S.; Filler, S.G.; Xiong, Y.Q. In vitro endothelial cell damage is positively correlated with enhanced virulence and poor vancomycin responsiveness in experimental endocarditis due to methicillin-resistant Staphylococcus aureus. Cell. Microbiol. 2011, 13, 1530–1541. [Google Scholar] [CrossRef] [PubMed]
- Laabei, M.; Uhlemann, A.C.; Lowy, F.D.; Austin, E.D.; Yokoyama, M.; Ouadi, K.; Feil, E.; Thorpe, H.A.; Williams, B.; Perkins, M.; et al. Evolutionary Trade-Offs Underlie the Multi-faceted Virulence of Staphylococcus aureus. PLoS Biol. 2015, 13, e1002229. [Google Scholar] [CrossRef] [PubMed]
- Suligoy, C.M.; Lattar, S.M.; Noto Llana, M.; Gonzalez, C.D.; Alvarez, L.P.; Robinson, D.A.; Gomez, M.I.; Buzzola, F.R.; Sordelli, D.O. Mutation of Agr Is Associated with the Adaptation of Staphylococcus aureus to the Host during Chronic Osteomyelitis. Front. Cell. Infect. Microbiol. 2018, 8, 18. [Google Scholar] [CrossRef] [PubMed]
- Joo, E.J.; Choi, J.Y.; Chung, D.R.; Song, J.H.; Ko, K.S. Characteristics of the community-genotype sequence type 72 methicillin-resistant Staphylococcus aureus isolates that underlie their persistence in hospitals. J. Microbiol. 2016, 54, 445–450. [Google Scholar] [CrossRef] [PubMed]
- Holtfreter, S.; Grumann, D.; Balau, V.; Barwich, A.; Kolata, J.; Goehler, A.; Weiss, S.; Holtfreter, B.; Bauerfeind, S.S.; Doring, P.; et al. Molecular Epidemiology of Staphylococcus aureus in the General Population in Northeast Germany: Results of the Study of Health in Pomerania (SHIP-TREND-0). J. Clin. Microbiol. 2016, 54, 2774–2785. [Google Scholar] [CrossRef] [PubMed]
- Otto, M. Staphylococcus aureus toxins. Curr. Opin. Microbiol. 2014, 17, 32–37. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cheung, G.Y.; Joo, H.S.; Chatterjee, S.S.; Otto, M. Phenol-soluble modulins--critical determinants of staphylococcal virulence. FEMS Microbiol. Rev. 2014, 38, 698–719. [Google Scholar] [CrossRef] [PubMed]
- Tuchscherr, L.; Medina, E.; Hussain, M.; Volker, W.; Heitmann, V.; Niemann, S.; Holzinger, D.; Roth, J.; Proctor, R.A.; Becker, K.; et al. Staphylococcus aureus phenotype switching: An effective bacterial strategy to escape host immune response and establish a chronic infection. EMBO Mol. Med. 2011, 3, 129–141. [Google Scholar] [CrossRef] [PubMed]
- Horst, S.A.; Hoerr, V.; Beineke, A.; Kreis, C.; Tuchscherr, L.; Kalinka, J.; Lehne, S.; Schleicher, I.; Kohler, G.; Fuchs, T.; et al. A novel mouse model of Staphylococcus aureus chronic osteomyelitis that closely mimics the human infection: An integrated view of disease pathogenesis. Am. J. Pathol. 2012, 181, 1206–1214. [Google Scholar] [CrossRef] [PubMed]
- Recker, M.; Laabei, M.; Toleman, M.S.; Reuter, S.; Saunderson, R.B.; Blane, B.; Torok, M.E.; Ouadi, K.; Stevens, E.; Yokoyama, M.; et al. Clonal differences in Staphylococcus aureus bacteraemia-associated mortality. Nat. Microbiol. 2017, 2, 1381–1388. [Google Scholar] [CrossRef] [PubMed]
- Sandulescu, O.; Bleotu, C.; Matei, L.; Streinu-Cercel, A.; Oprea, M.; Dragulescu, E.C.; Chifiriuc, M.C.; Rafila, A.; Pirici, D.; Talapan, D.; et al. Comparative evaluation of aggressiveness traits in staphylococcal strains from severe infections versus nasopharyngeal carriage. Microb. Pathog. 2017, 102, 45–53. [Google Scholar] [CrossRef] [PubMed]
- Casadevall, A.; Pirofski, L.A. Host-pathogen interactions: Redefining the basic concepts of virulence and pathogenicity. Infect. Immun. 1999, 67, 3703–3713. [Google Scholar] [PubMed]
- Oliveira, D.; Borges, A.; Simoes, M. Staphylococcus aureus Toxins and Their Molecular Activity in Infectious Diseases. Toxins 2018, 10, 252. [Google Scholar] [CrossRef] [PubMed]
- Rudkin, J.K.; McLoughlin, R.M.; Preston, A.; Massey, R.C. Bacterial toxins: Offensive, defensive, or something else altogether? PLoS Pathog. 2017, 13, e1006452. [Google Scholar] [CrossRef] [PubMed]
- Davis, K.M.; Isberg, R.R. One for All, but Not All for One: Social Behavior during Bacterial Diseases. Trends Microbiol. 2018. [Google Scholar] [CrossRef] [PubMed]
- Priest, N.K.; Rudkin, J.K.; Feil, E.J.; van den Elsen, J.M.; Cheung, A.; Peacock, S.J.; Laabei, M.; Lucks, D.A.; Recker, M.; Massey, R.C. From genotype to phenotype: Can systems biology be used to predict Staphylococcus aureus virulence? Nat. Rev. Microbiol. 2012, 10, 791–797. [Google Scholar] [CrossRef] [PubMed]
- Davis, K.M.; Isberg, R.R. Defining heterogeneity within bacterial populations via single cell approaches. Bioessays News Rev. Mol. Cell. Dev. Biol. 2016, 38, 782–790. [Google Scholar] [CrossRef] [PubMed]
- Avery, S.V. Microbial cell individuality and the underlying sources of heterogeneity. Nat. Rev. Microbiol. 2006, 4, 577–587. [Google Scholar] [CrossRef] [PubMed]
- Abajy, M.Y.; Kopec, J.; Schiwon, K.; Burzynski, M.; Doring, M.; Bohn, C.; Grohmann, E. A type IV-secretion-like system is required for conjugative DNA transport of broad-host-range plasmid pIP501 in gram-positive bacteria. J. Bacteriol. 2007, 189, 2487–2496. [Google Scholar] [CrossRef] [PubMed]
- Deinhardt-Emmer, S.; Sachse, S.; Geraci, J.; Fischer, C.; Kwetkat, A.; Dawczynski, K.; Tuchscherr, L.; Loffler, B. Virulence patterns of Staphylococcus aureus strains from nasopharyngeal colonization. J. Hosp. Infect. 2018, 100, 309–315. [Google Scholar] [CrossRef] [PubMed]
- Schijffelen, M.J.; Boel, C.E.; van Strijp, J.A.; Fluit, A.C. Whole genome analysis of a livestock-associated methicillin-resistant Staphylococcus aureus ST398 isolate from a case of human endocarditis. BMC Genom. 2010, 11, 376. [Google Scholar] [CrossRef] [PubMed]
- Tuchscherr, L.; Korpos, E.; van de Vyver, H.; Findeisen, C.; Kherkheulidze, S.; Siegmund, A.; Deinhardt-Emmer, S.; Bach, O.; Rindert, M.; Mellmann, A.; et al. Staphylococcus aureus requires less virulence to establish an infection in diabetic hosts. Int. J. Med. Microbiol. 2018, 308, 761–769. [Google Scholar] [CrossRef] [PubMed]
- Haslinger, B.; Strangfeld, K.; Peters, G.; Schulze-Osthoff, K.; Sinha, B. Staphylococcus aureus alpha-toxin induces apoptosis in peripheral blood mononuclear cells: Role of endogenous tumour necrosis factor-alpha and the mitochondrial death pathway. Cell. Microbiol. 2003, 5, 729–741. [Google Scholar] [CrossRef] [PubMed]
- Pfaffl, M.W. A new mathematical model for relative quantification in real-time RT-PCR. Nucleic Acids Res. 2001, 29, e45. [Google Scholar] [CrossRef] [PubMed]
- Oliphant, T.E. Python for scientific computing. Comput. Sci. Eng. 2007, 9, 10–20. [Google Scholar] [CrossRef]






© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tuchscherr, L.; Pöllath, C.; Siegmund, A.; Deinhardt-Emmer, S.; Hoerr, V.; Svensson, C.-M.; Thilo Figge, M.; Monecke, S.; Löffler, B. Clinical S. aureus Isolates Vary in Their Virulence to Promote Adaptation to the Host. Toxins 2019, 11, 135. https://0-doi-org.brum.beds.ac.uk/10.3390/toxins11030135
Tuchscherr L, Pöllath C, Siegmund A, Deinhardt-Emmer S, Hoerr V, Svensson C-M, Thilo Figge M, Monecke S, Löffler B. Clinical S. aureus Isolates Vary in Their Virulence to Promote Adaptation to the Host. Toxins. 2019; 11(3):135. https://0-doi-org.brum.beds.ac.uk/10.3390/toxins11030135
Chicago/Turabian StyleTuchscherr, Lorena, Christine Pöllath, Anke Siegmund, Stefanie Deinhardt-Emmer, Verena Hoerr, Carl-Magnus Svensson, Marc Thilo Figge, Stefan Monecke, and Bettina Löffler. 2019. "Clinical S. aureus Isolates Vary in Their Virulence to Promote Adaptation to the Host" Toxins 11, no. 3: 135. https://0-doi-org.brum.beds.ac.uk/10.3390/toxins11030135