Prodigiosin Promotes Nrf2 Activation to Inhibit Oxidative Stress Induced by Microcystin-LR in HepG2 Cells

 , , and
, , and 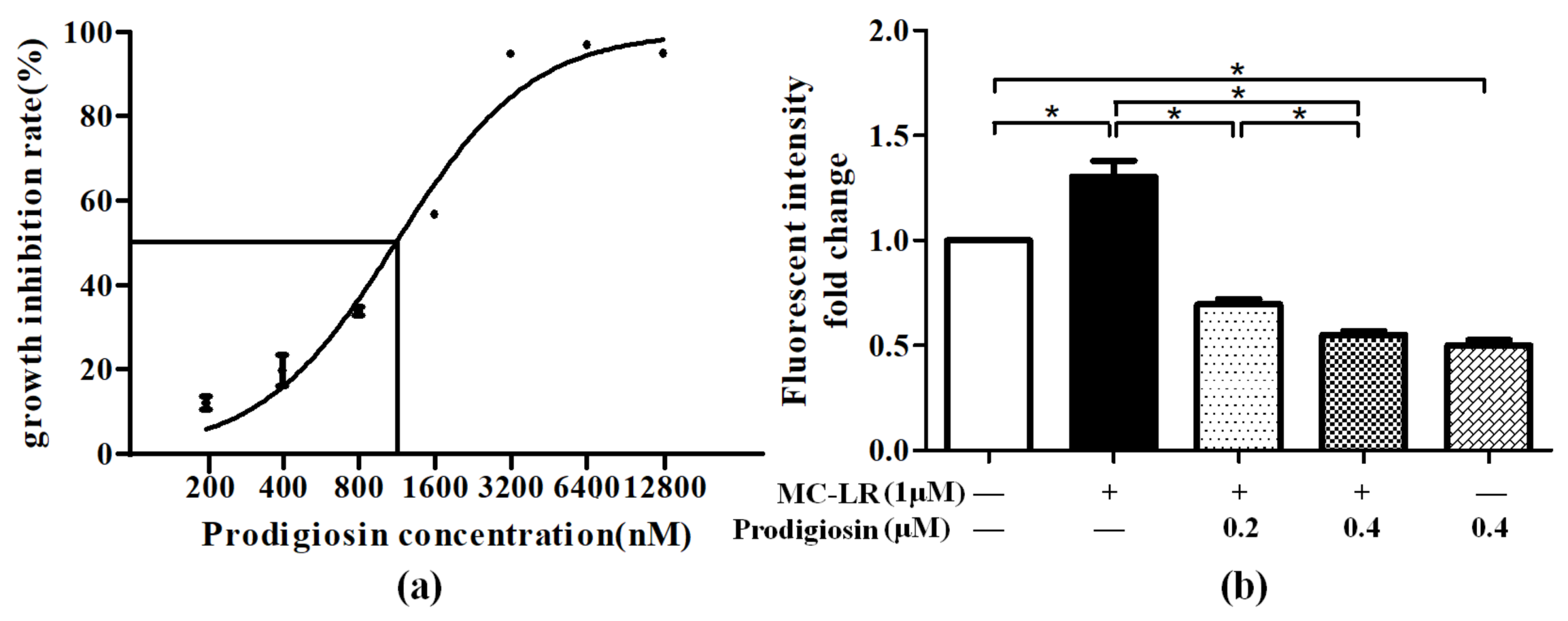{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
2.1. Effect of ROS Level Induced by MC-LR and Prodigiosin
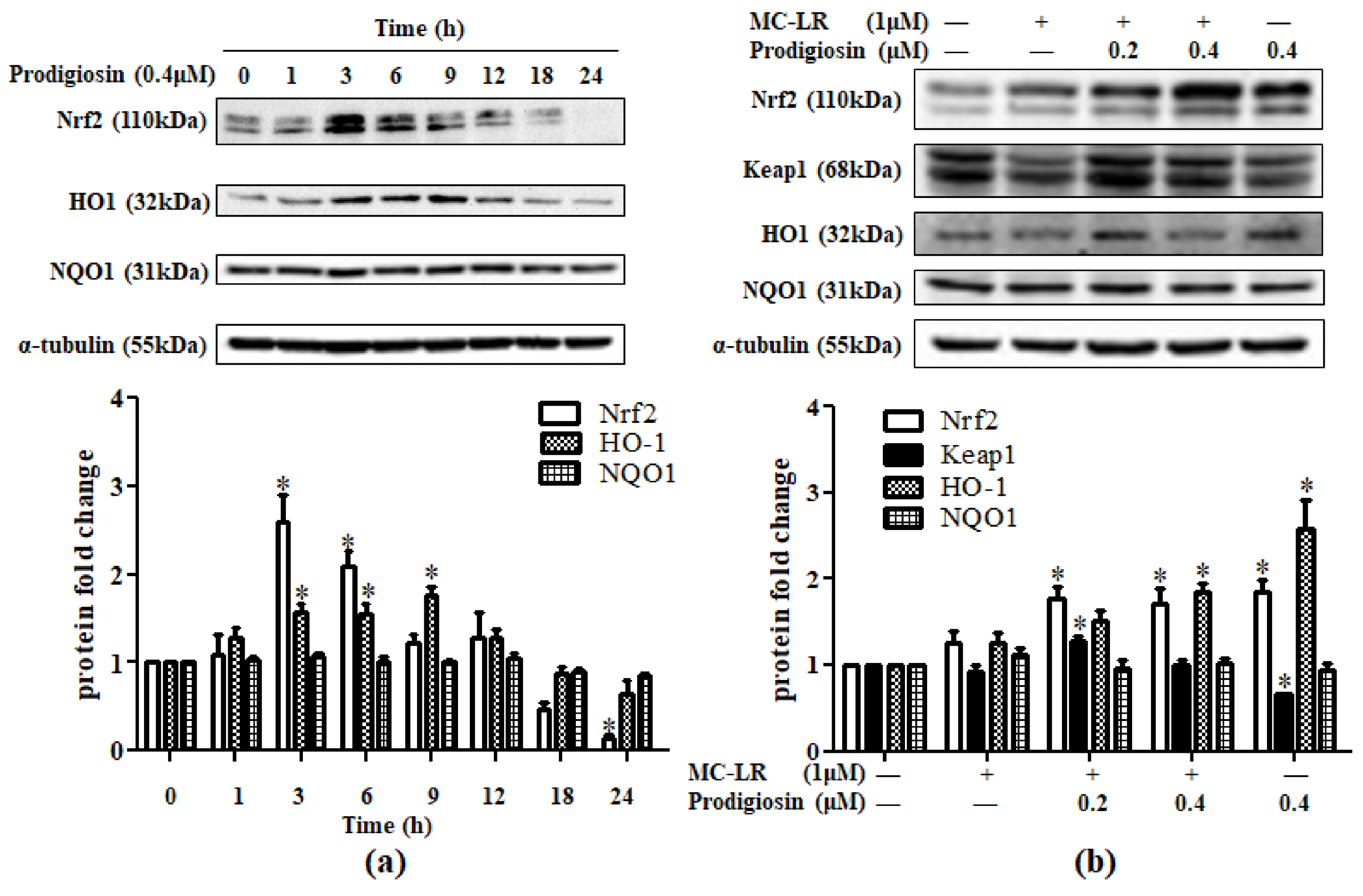
2.2. Effect of Prodigiosin and MC-LR in Nrf2-Related Protein
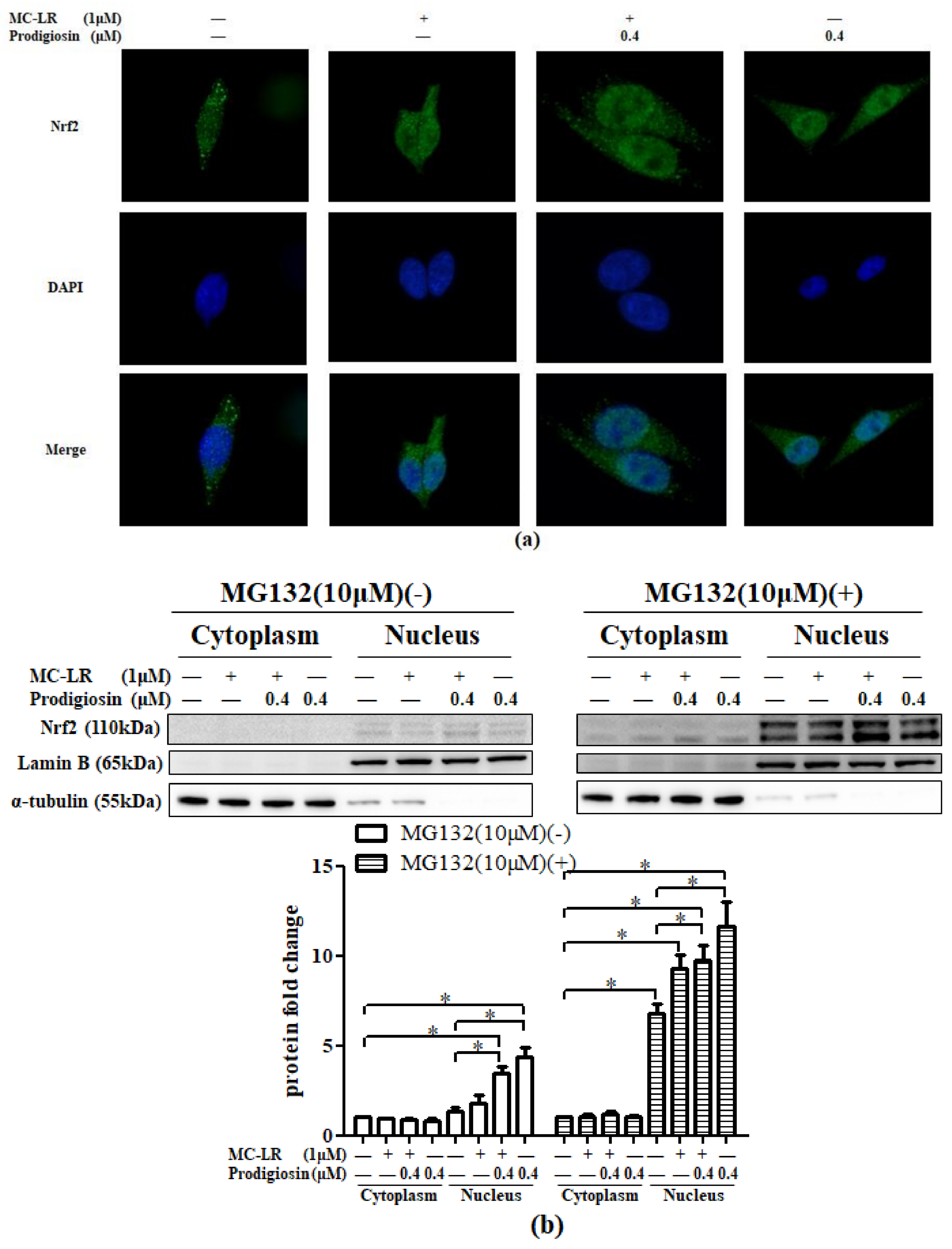
2.3. Prodigiosin and MC-LR Affect the Translocation of Nrf2
2.3.1. Activation of Nrf2 in the Nucleus
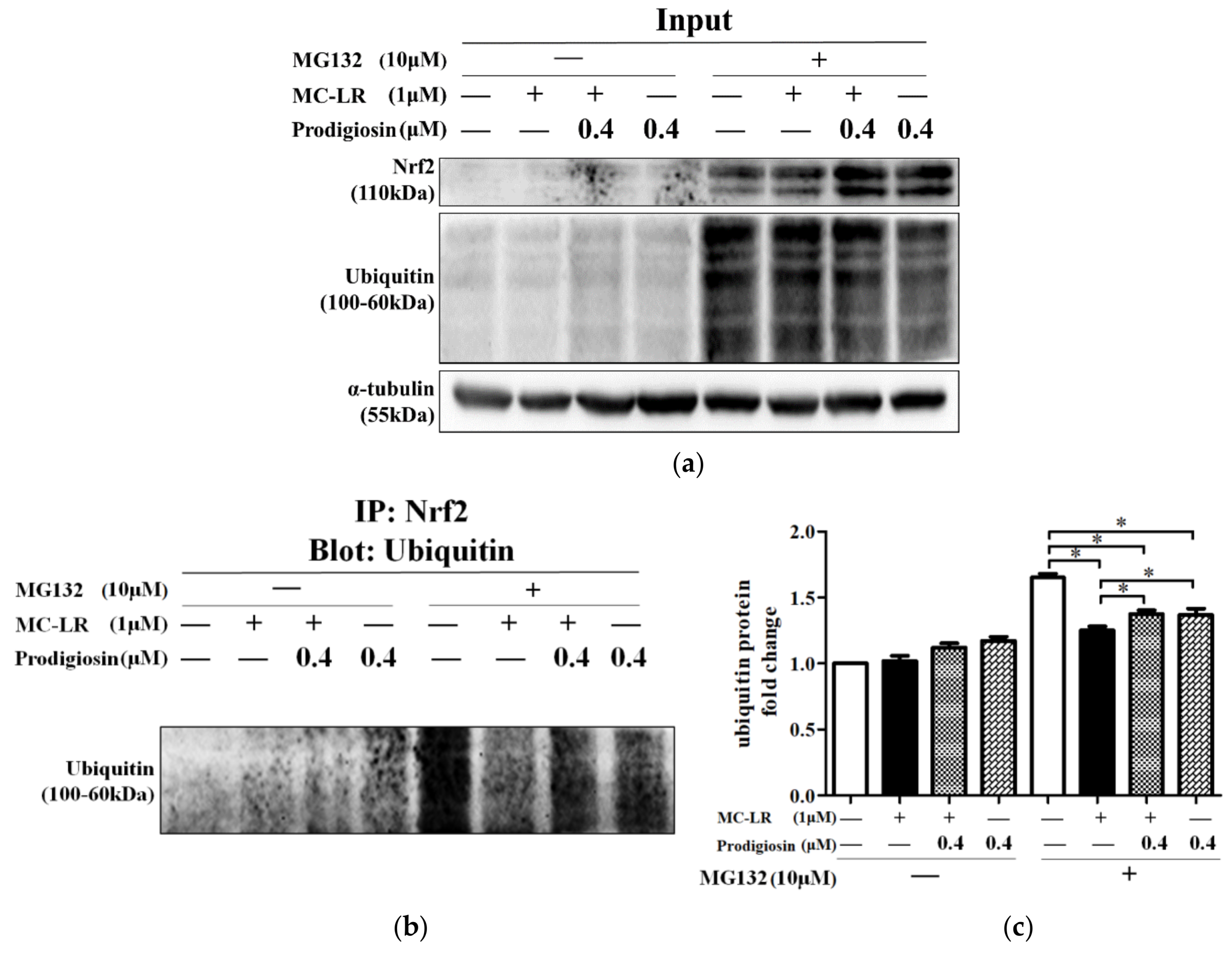
2.3.2. Inhibition of Ubiquitination by Prodigiosin and MC-LR
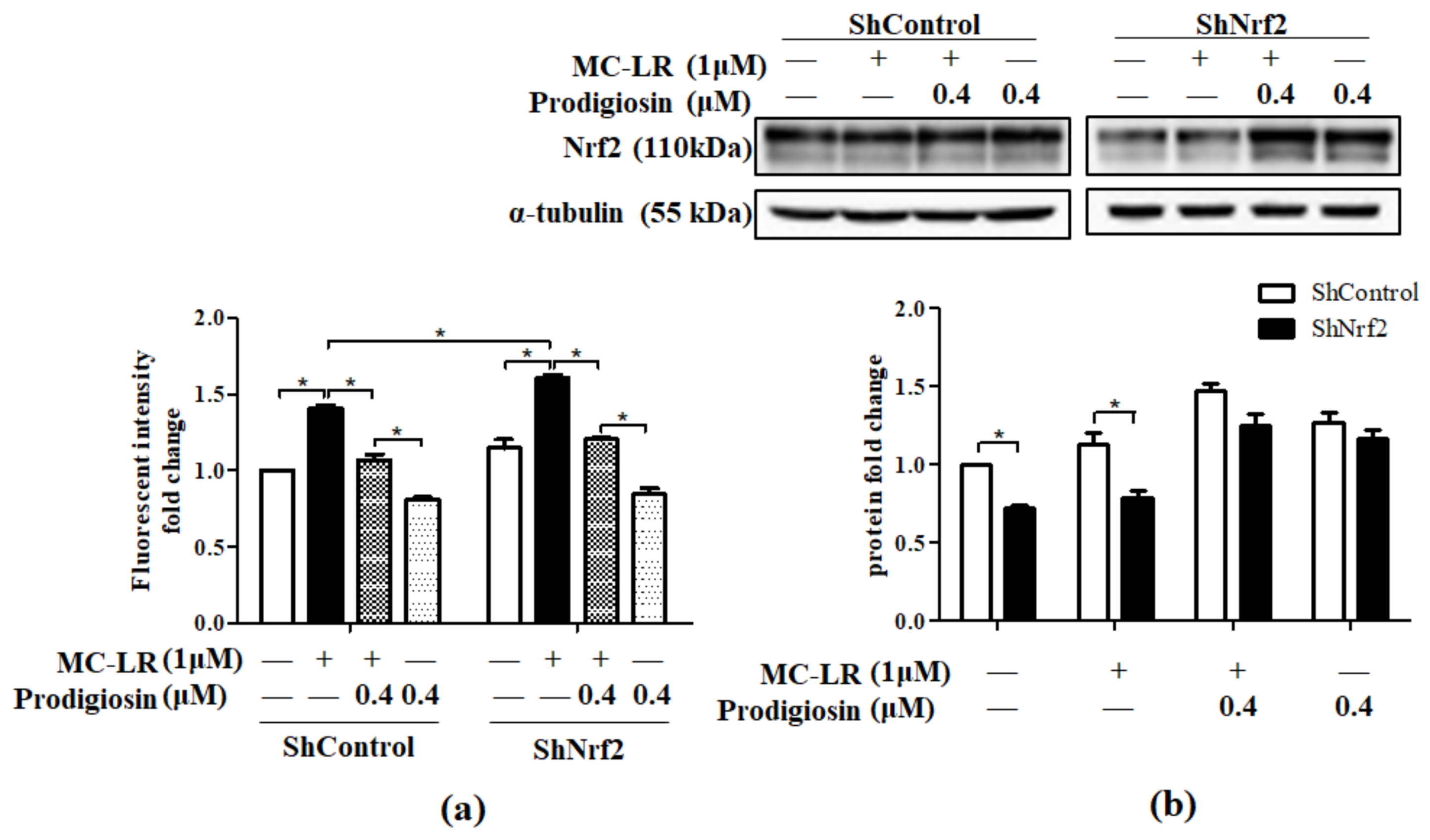
2.4. Effect of Prodigiosin and MC-LR on Nrf2 Knocked Down Cells.
3. Discussion
4. Materials and Methods
4.1. Cell Culture and Treatment
4.2. CCK8 Assay
4.3. ROS Assay
4.4. Western Blot Analysis
4.5. Immunofluorescence Assay
4.6. Immunoprecipitation
4.7. Transfection
4.8. Statistical Analysis
Author Contributions
Funding
Conflicts of Interest
References
- Codd, G.A.; Morrison, L.F.; Metcalf, J.S. Cyanobacterial toxins: Risk management for health protection. Toxicol. Appl. Pharm. 2005, 203, 264–272. [Google Scholar] [CrossRef] [PubMed]
- Massey, I.Y.; Yang, F.; Ding, Z.; Yang, S.; Guo, J.; Tezi, C.; Al-Osman, M.; Kamegni, R.B.; Zeng, W. Exposure routes and health effects of microcystins on animals and humans: A mini-review. Toxicon 2018, 151, 156–162. [Google Scholar] [CrossRef] [PubMed]
- Roegner, A.F.; Brena, B.; González-Sapienza, G.; Puschner, B. Microcystins in potable surface waters: Toxic effects and removal strategies. J. Appl. Toxicol. 2014, 34, 441–457. [Google Scholar] [CrossRef] [PubMed]
- Zimba, P.V.; Grimm, C.C. A synoptic survey of musty/muddy odor metabolites and microcystin toxin occurrence and concentration in southeastern USA channel catfish (Ictalurus punctatus Ralfinesque) production ponds. Aquaculture 2003, 218, 81–87. [Google Scholar] [CrossRef]
- Beaver, J.R.; Manis, E.E.; Loftin, K.A.; Graham, J.L.; Pollard, A.I.; Mitchell, R.M. Land use patterns, ecoregion, and microcystin relationships in U.S. lakes and reservoirs: A preliminary evaluation. Harmful Algae 2014, 36, 57–62. [Google Scholar] [CrossRef]
- Loftin, K.A.; Graham, J.L.; Hilborn, E.D.; Lehmann, S.C.; Meyer, M.T.; Dietze, J.E.; Griffith, C.B. Cyanotoxins in inland lakes of the United States: Occurrence and potential recreational health risks in the EPA National Lakes Assessment 2007. Harmful Algae 2016, 56, 77–90. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Carmichael, W.W. Assessment of blue-green algal toxins in raw and finished drinking water. AWWA Research Foundation and American Water Works Association; American Water Works Association: Denver, CO, USA, 2000. [Google Scholar]
- Greer, B.; McNamee, S.E.; Boots, B.; Cimarelli, L.; Guillebault, D.; Helmi, K.; Marcheggiani, S.; Panaiotov, S.; Breitenbach, U.; Akçaalan, R.; et al. A validated UPLC–MS/MS method for the surveillance of ten aquatic biotoxins in European brackish and freshwater systems. Harmful Algae 2016, 55, 31–40. [Google Scholar] [CrossRef] [PubMed]
- Chen, L.; Yang, S.; Wen, C.; Zheng, S.; Yang, Y.; Feng, X.; Chen, J.; Luo, D.; Liu, R.; Yang, F. Regulation of Microcystin-LR-Induced DNA Damage by miR-451a in HL7702 Cells. Toxins 2019, 11, 164. [Google Scholar] [CrossRef] [PubMed]
- Wen, C.; Yang, S.; Zheng, S.; Feng, X.; Chen, J.; Yang, F. Analysis of long non-coding RNA profiled following MC-LR-induced hepatotoxicity using high-throughput sequencing. J. Toxicol. Environ. Health Part A 2018, 81, 1165–1172. [Google Scholar] [CrossRef]
- Yang, S.; Chen, L.; Wen, C.; Zhang, X.; Feng, X.; Yang, F. MicroRNA expression profiling involved in MC-LR-induced hepatotoxicity using high-throughput sequencing analysis. J. Toxicol. Environ. Health Part A 2018, 81, 89–97. [Google Scholar] [CrossRef] [PubMed]
- Eriksson, J.E.; Toivola, D.; Meriluoto, J.A.; Karaki, H.; Han, Y.G.; Hartshorne, D. Hepatocyte deformation induced by cyanobacterial toxins reflects inhibition of protein phosphatases. Biochem. Biophys. Res. Commun. 1990, 173, 1347–1353. [Google Scholar] [CrossRef]
- MacKintosh, C.; Beattie, K.A.; Klumpp, S.; Cohen, P.; Codd, G.A. Cyanobacterial microcystin-LR is a potent and specific inhibitor of protein phosphatases 1 and 2A from both mammals and higher plants. FEBS Lett. 1990, 264, 187–192. [Google Scholar] [CrossRef] [Green Version]
- Hagenbuch, B.; Meier, P.J. The superfamily of organic anion transporting polypeptides. Biochim. Biophys. Acta 2003, 1609, 1–18. [Google Scholar] [CrossRef]
- Yang, F.; Wen, C.; Zheng, S.; Yang, S.; Chen, J.; Feng, X. Involvement of MAPK/ERK1/2 pathway in microcystin-induced microfilament reorganization in HL7702 hepatocytes. J. Toxicol. Environ. Health Part A 2018, 81, 1135–1141. [Google Scholar] [CrossRef] [PubMed]
- Fischer, W.J.; Dietrich, D.R. Pathological and Biochemical Characterization of Microcystin-Induced Hepatopancreas and Kidney Damage in Carp (Cyprinuscarpio). Toxicol. Appl. Pharmacol. 2000, 164, 73–81. [Google Scholar] [CrossRef] [PubMed]
- Gehringer, M.M. Microcystin-LR and okadaic acid-induced cellular effects: A dualistic response. FEBS Lett. 2004, 557, 1–8. [Google Scholar] [CrossRef]
- Stankovic, N.; Senerovic, L.; Ilic-Tomic, T.; Vasiljevic, B.; Nikodinovic-Runic, J. Properties and applications of undecylprodigiosin and other bacterial prodigiosins. Appl. Microbiol. Biot. 2014, 98, 3841–3858. [Google Scholar] [CrossRef] [PubMed]
- Darveau, R.P.; Lynch, D.L. The antibiotic activity of prodigiosins against certain blue-green and green algae. Phycologia 1977, 16, 349–351. [Google Scholar] [CrossRef]
- Yang, F.; Wei, H.; Li, X.; Li, Y.; Li, X.; Yin, L.; Pu, Y. Isolation and characterization of an algicidal bacterium indigenous to lake Taihu with a red pigment able to lyse microcystis aeruginosa. Biomed. Environ. Sci. 2013, 26, 148–154. [Google Scholar]
- Elahian, F.; Moghimi, B.; Dinmohammadi, F.; Ghamghami, M.; Hamidi, M.; Mirzaei, S.A. The anticancer agent prodigiosin is not a multidrug resistance protein substrate. DNA Cell Biol. 2013, 32, 90–97. [Google Scholar] [CrossRef]
- Ibrahim, D.; Nazari, T.F.; Kassim, J.; Lim, S. Prodigiosin—An antibacterial red pigment produced by Serratia marcescens IBRL USM 84 associated with a marine sponge Xestospongia testudinaria. J. Appl. Pharmaceut. Sci. 2014, 4, 1–6. [Google Scholar]
- Papireddy, K.; Smilkstein, M.; Kelly, J.X.; Shweta; Salem, S.M.; Alhamadsheh, M.; Haynes, S.W.; Challis, G.L.; Reynolds, K.A. Antimalarial activity of natural and synthetic prodiginines. J. Med. Chem. 2011, 54, 5296–5306. [Google Scholar] [CrossRef] [PubMed]
- Arivizhivendhan, K.V.; Mahesh, M.; Boopathy, R.; Swarnalatha, S.; Mary, R.R.; Sekaran, G. Antioxidant and antimicrobial activity of bioactive prodigiosin produces from Serratia marcescens using agricultural waste as a substrate. J. Food Sci. Technol. Mysore 2018, 55, 2661–2670. [Google Scholar] [CrossRef] [PubMed]
- Lu, H.; Lei, X.; Zhang, Q. Moderate activation of IKK2-NF-kB in unstressed adult mouse liver induces cytoprotective genes and lipogenesis without apparent signs of inflammation or fibrosis. BMC Gastroenterol. 2015, 15, 94. [Google Scholar] [CrossRef] [PubMed]
- Butterfield, D.A. Amyloid beta-peptide (1-42)-induced oxidative stress and neurotoxicity: Implications for neurodegeneration in Alzheimer’s disease brain. A review. Free Radical Res. Commun. 2002, 36, 1307–1313. [Google Scholar] [CrossRef]
- Zandalinas, S.I.; Mittler, R. ROS-induced ROS release in plant and animal cells. Free Radical Bio. Med. 2018, 122, 21–27. [Google Scholar] [CrossRef]
- Wang, Z.; Li, G.; Wu, Q.; Liu, C.; Shen, J.; Yan, W. Microcystin-LR exposure induced nephrotoxicity by triggering apoptosis in female zebrafish. Chemosphere 2019, 214, 598–605. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.; Wu, Y.; Fang, W.; Wang, D. Regulatory effect of quercetin on hazardous microcystin-LR-induced apoptosis of Carassius auratus lymphocytes in vitro. Fish. Shellfish Immun. 2014, 37, 278–285. [Google Scholar] [CrossRef]
- Takumi, S.; Ikema, S.; Hanyu, T.; Shima, Y.; Kurimoto, T.; Shiozaki, K.; Sugiyama, Y.; Park, H.; Ando, S.; Furukawa, T.; et al. Naringin attenuates the cytotoxicity of hepatotoxin microcystin-LR by the curious mechanisms to OATP1B1- and OATP1B3-expressing cells. Environ. Toxicol. Phar. 2015, 39, 974–981. [Google Scholar] [CrossRef]
- Žegura, B.; Lah, T.; Filipič, M. The role of reactive oxygen species in microcystin-LR-induced DNA damage. Toxicology 2004, 200, 59–68. [Google Scholar]
- Chang, C.; Wang, Y.; Chern, C.; Liou, K.; Hou, Y.; Peng, Y.; Shen, Y. Prodigiosin inhibits gp91phox and iNOX expression to protect mice against the oxidative/nitrosative brain injury induced by hypoxia-ischemia. Toxicol. Appl. Pharmacol. 2011, 257, 137–147. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.; Zheng, W.; Feng, X.; Yang, F.; Qin, H.; Wu, S.; Hou, D.X.; Chen, J. Nrf2–ARE signaling acts as master pathway for the cellular antioxidant activity of fisetin. Molecules 2019, 24, 708. [Google Scholar] [CrossRef] [PubMed]
- Jaramillo, M.C.; Zhang, D.D. The emerging role of the Nrf2-Keap1 signaling pathway in cancer. Gene. Dev. 2013, 27, 2179–2191. [Google Scholar] [CrossRef] [PubMed]
- Yao, J.; Zhang, B.; Ge, C.; Peng, S.; Fang, J. Xanthohumol, a polyphenol chalcone present in hops, activating nrf2 enzymes to confer protection against oxidative damage in PC12 Cells. J. Agr. Food Chem. 2015, 63, 1521–1531. [Google Scholar] [CrossRef] [PubMed]
- Na, H.; Surh, Y. Oncogenic potential of Nrf2 and its principal target protein heme oxygenase-1. Free Radical Biol. Med. 2014, 67, 353–365. [Google Scholar] [CrossRef] [PubMed]
- Lu, Y.F.; Liu, J.; Wu, K.C.; Qu, Q.; Fan, F.; Klaassen, C.D. Overexpression of Nrf2 protects against microcystin-induced hepatotoxicity in mice. PLoS ONE 2014, 9, e93013. [Google Scholar] [CrossRef] [PubMed]
- Gan, N.; Sun, X.; Song, L. Activation of Nrf2 by microcystin-LR provides advantages for liver cancer cell growth. Chem. Res. Toxicol. 2010, 23, 1477–1484. [Google Scholar] [CrossRef]
- Kyun, H.K.; Ruxana, T.S.; Ji, Y.L.; Han, S.J.; Yu, K.O.; Timothy, S.B.; Myungsoo, S. Suppressed ubiquitination of Nrf2 by p47phox contributes to Nrf2 activation. Free Radical Biol. Med. 2017, 113, 48–58. [Google Scholar]
- Gan, N.; Mi, L.; Sun, X.; Dai, G.; Chung, F.; Song, L. Sulforaphane protects Microcystin-LR-induced toxicity through activation of the Nrf2-mediated defensive response. Toxicol. Appl. Pharm. 2010, 247, 129–137. [Google Scholar] [CrossRef] [Green Version]





© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chen, J.; Li, Y.; Liu, F.; Hou, D.-X.; Xu, J.; Zhao, X.; Yang, F.; Feng, X. Prodigiosin Promotes Nrf2 Activation to Inhibit Oxidative Stress Induced by Microcystin-LR in HepG2 Cells. Toxins 2019, 11, 403. https://0-doi-org.brum.beds.ac.uk/10.3390/toxins11070403
Chen J, Li Y, Liu F, Hou D-X, Xu J, Zhao X, Yang F, Feng X. Prodigiosin Promotes Nrf2 Activation to Inhibit Oxidative Stress Induced by Microcystin-LR in HepG2 Cells. Toxins. 2019; 11(7):403. https://0-doi-org.brum.beds.ac.uk/10.3390/toxins11070403
Chicago/Turabian StyleChen, Jihua, Yuji Li, Fuqiang Liu, De-Xing Hou, Jingjing Xu, Xinying Zhao, Fei Yang, and Xiangling Feng. 2019. "Prodigiosin Promotes Nrf2 Activation to Inhibit Oxidative Stress Induced by Microcystin-LR in HepG2 Cells" Toxins 11, no. 7: 403. https://0-doi-org.brum.beds.ac.uk/10.3390/toxins11070403