Chemical and Biological Characteristics of Antimicrobial α-Helical Peptides Found in Solitary Wasp Venoms and Their Interactions with Model Membranes


Abstract
:1. Introduction
2. Chemical and Biological Characterization
2.1. Mastoparans
2.2. Eumenitins
2.3. Protonectin
2.4. Decoralin
2.5. Anoplin
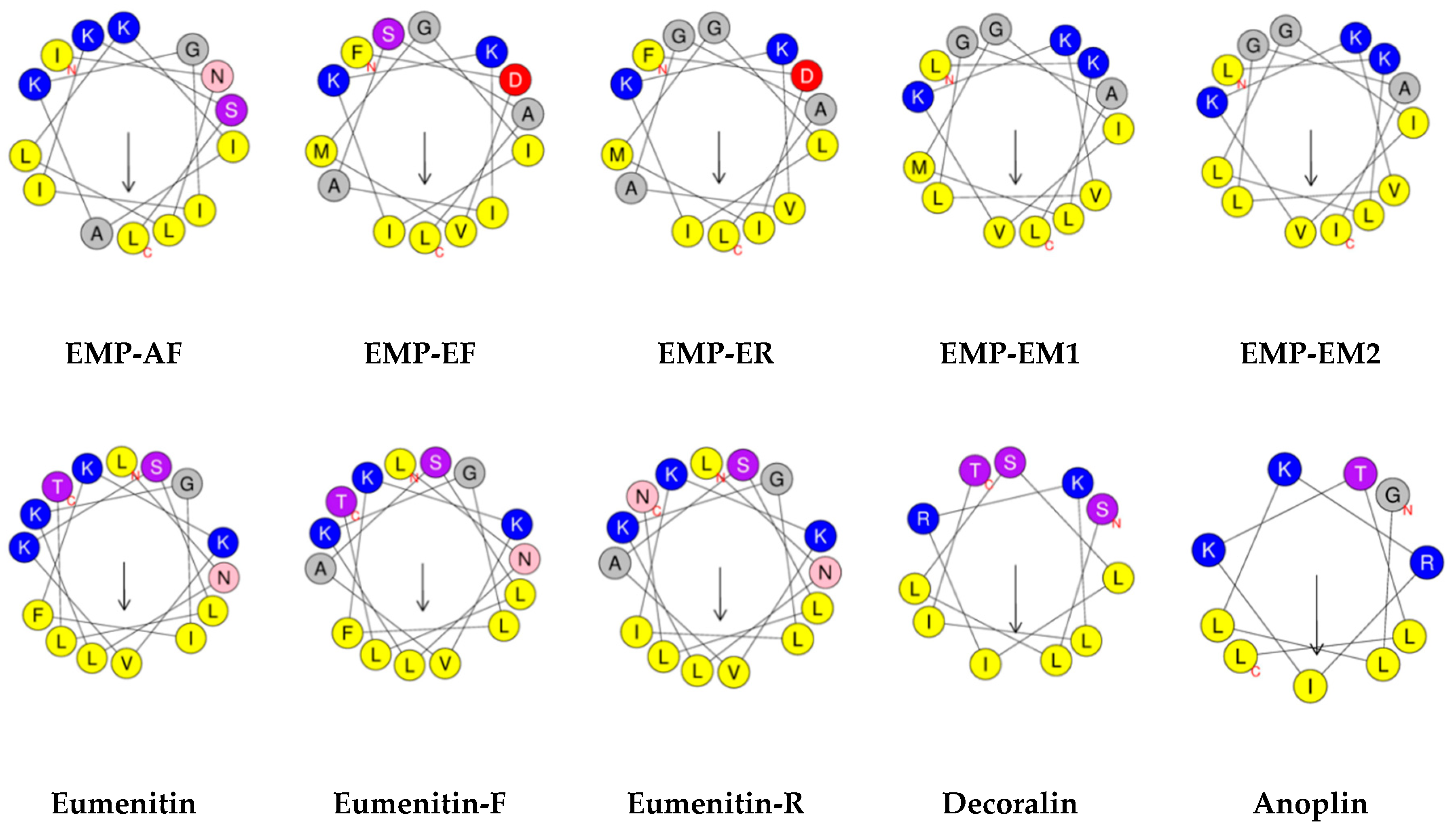
3. Physicochemical Properties and Secondary Structure
3.1. Sequence and Chain Length
3.2. N- and C-Termini
3.3. Charge and Helix Macro-Dipole
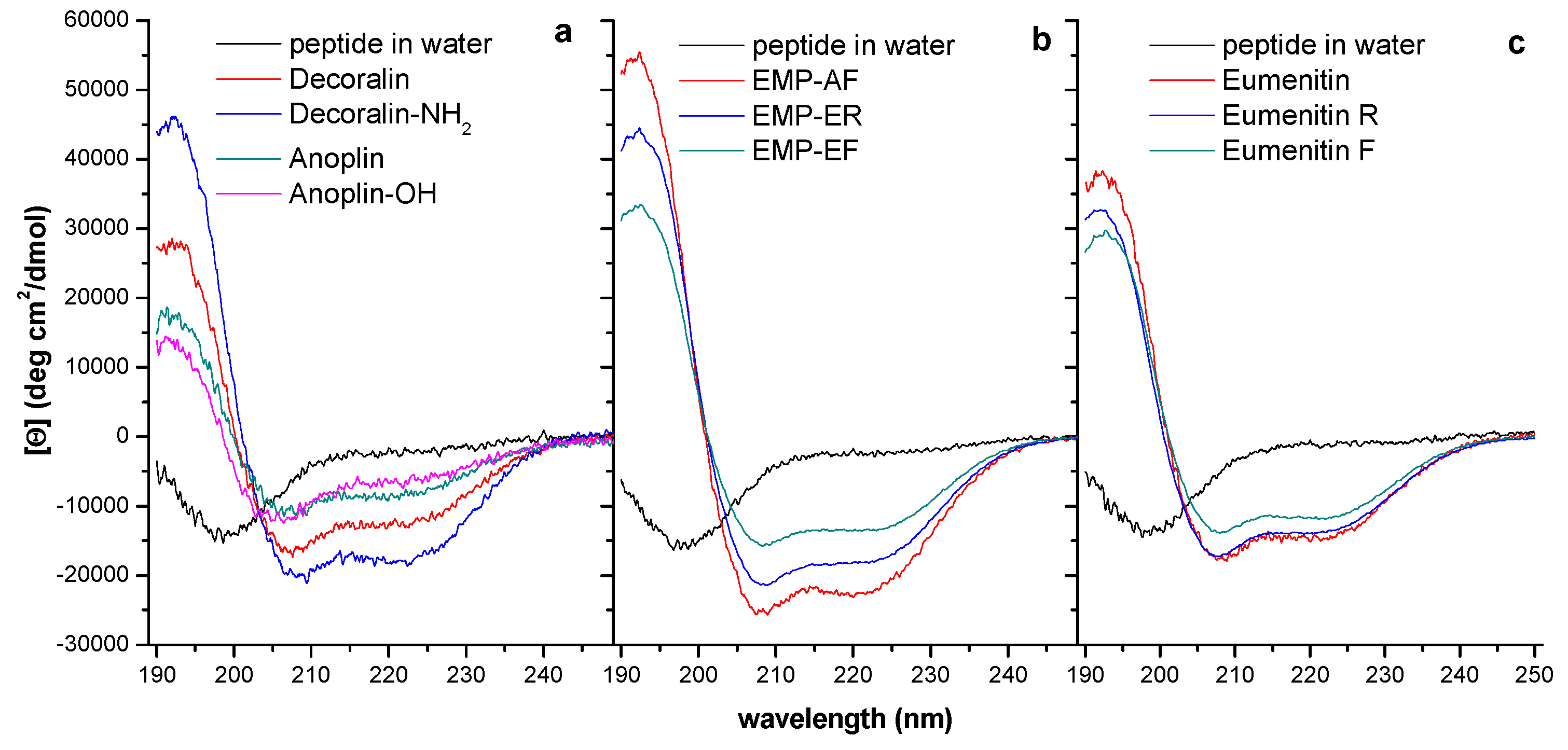
3.4. Helical Propensity and Helicity
3.5. Amphipathicity and Hydrophobic Moment
3.6. Hydrophobicity
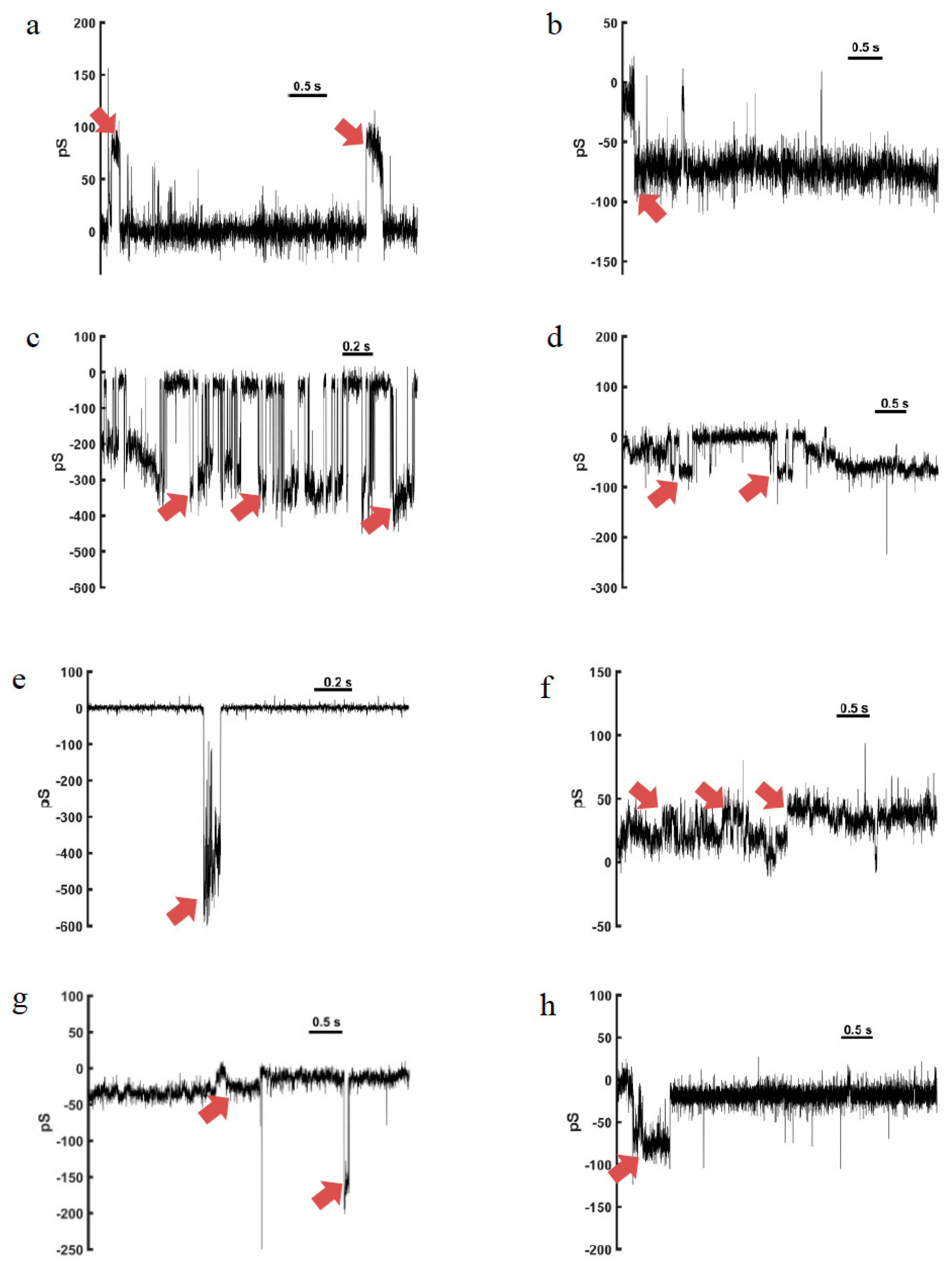
4. Channel-Like Pore-Forming Properties
4.1. Anoplin
4.2. Eumenitin
4.3. Eumenitin-F and Eumenitin-R
4.4. EMP-EF and EMP-ER
5. Concluding Remarks
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| EMP | eumenine mastoparan |
| PMTX | pompilidotoxin |
| HPLC | high performance liquid chromatography |
| CD | circular dichroism |
| TFE | 2,2,2-trifluoroethanol |
| SDS | sodium dodecylsulfate |
| NMR | nuclear magnetic resonance |
| KLA | lysine-leucine-alanine (amino acid sequence) |
| DPhPC | 1,2-diphytanoyl-sn-glycero-3-phosphocholine |
References
- Wang, G.; Li, X.; Wang, Z. APD3: The antimicrobial peptide database as a tool for research and education. Nucleic Acids Res. 2016, 44, D1087–D1093. [Google Scholar] [CrossRef] [PubMed]
- Koehbach, J.; Craik, D.J. The vast structural diversity of antimicrobial peptides. Trends Pharmacol. Sci. 2019, 40, 517–528. [Google Scholar] [CrossRef] [PubMed]
- Narayana, J.L.; Chen, J.-Y. Antimicrobial peptides: Possible anti-infective agents. Peptides 2015, 72, 88–94. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Dou, X.; Song, J.; Lyu, Y.; Zhu, X.; Xu, L.; Li, W.; Shan, A. Antimicrobial peptides: Promising alternatives in the post feeding antibiotic era. Med. Res. Rev. 2019, 39, 831–859. [Google Scholar] [CrossRef] [PubMed]
- Kuhn-Nentwig, L. Antimicrobial and cytolytic peptides of venomous arthropods. Cell. Mol. Life Sci. 2003, 60, 2651–2668. [Google Scholar] [CrossRef] [PubMed]
- Bulet, P.; Stöcklin, R.; Menin, L. Anti-microbial peptides: From invertebrates to vertebrates. Immunol. Rev. 2004, 198, 169–184. [Google Scholar] [CrossRef] [PubMed]
- Fratini, F.; Cilia, G.; Turchi, B.; Felicioli, A. Insects, arachnids and centipedes venom: A powerful weapon against bacteria. A literature review. Toxicon 2017, 130, 91–103. [Google Scholar] [CrossRef]
- Konno, K.; Hisada, M.; Naoki, H.; Itagaki, Y.; Kawai, N.; Miwa, A.; Yasuhara, T.; Morimoto, Y.; Nakata, Y. Structure and biological activities of eumenine mastoparan-AF (EMP-AF), a novel mast cell degranulating peptide in the venom of the solitary wasp (Anterhynchium flavomarginatum micado). Toxicon 2000, 38, 1505–1515. [Google Scholar] [CrossRef]
- Konno, K.; Hisada, M.; Fontana, R.; Lorenzi, C.C.B.; Naoki, H.; Itagaki, Y.; Miwa, A.; Kawai, N.; Nakata, Y.; Yasuhara, T.; et al. Anoplin, a novel antimicrobial peptide from the venom of the solitary wasp Anoplius samariensis. Biochim. Biophys. Acta 2001, 1550, 70–80. [Google Scholar] [CrossRef]
- Konno, K.; Hisada, M.; Naoki, H.; Itagaki, Y.; Fontana, R.; Rangel, M.; Oliveira, J.S.; Cabrera, M.P.; Ruggiero Neto, J.; Hide, I.; et al. Eumenitin, a novel antimicrobial peptide from the venom of the solitary eumenine wasp Eumenes rubronotatus. Peptides 2006, 27, 2624–2631. [Google Scholar] [CrossRef]
- Konno, K.; Rangel, M.; Oliveira, J.S.; Cabrera, M.P.S.; Fontana, R.; Hirata, I.Y.; Nakata, Y.; Mori, K.; Kawano, M.; Fuchino, H.; et al. Decoralin, a novel linear cationic α-helical peptide from the venom of the solitary eumenine wasp Oreumenes decoratus. Peptides 2007, 28, 2320–2327. [Google Scholar] [CrossRef] [PubMed]
- Rangel, M.; Cabrera, M.P.S.; Kazuma, K.; Ando, K.; Wang, X.; Kato, M.; Nihei, K.; Hirata, I.Y.; Cross, T.; Garcia, A.N.; et al. Chemical and biological characterization of four new antimicrobial and α-helical peptides from the venoms of two solitary eumenine wasps. Toxicon 2011, 57, 1081–1092. [Google Scholar] [CrossRef] [PubMed]
- Konno, K.; Rangel, M.; de Oliveira, J.S.; Fontana, R.; Kawano, M.; Fuchino, H.; Hide, I.; Yasuhara, T.; Nakata, Y. New mastoparan peptides in the venom of the solitary eumenine wasp Eumenes micado. Toxins 2019, 11, 155. [Google Scholar] [CrossRef] [PubMed]
- Cabrera, M.P.S.; Souza, B.M.; Fontana, R.; Konno, K.; Palma, M.S.; de Azevedo, W.F., Jr.; Ruggiero Neto, J. Conformation and lytic activity of eumenine mastoparan: A new antimicrobial peptide from wasp venom. J. Pept. Res. 2004, 64, 95–103. [Google Scholar] [CrossRef] [PubMed]
- Cabrera, M.P.S.; Arcisio-Miranda, M.; Costa, S.T.B.; Konno, K.; Ruggiero, J.R.; Procopio, J.; Ruggiero Neto, J. Study of the mechanism of action of Anoplin, a helical antimicrobial decapeptide with ion channel-like activity, and the role of the amidated C-terminus. J. Pept. Sci. 2008, 14, 661–669. [Google Scholar] [CrossRef] [PubMed]
- Arcisio-Miranda, M.; Cabrera, M.P.S.; Konno, K.; Rangel, M.; Procopio, J. Effects of the cationic antimicrobial peptide Eumenitin from the venom of solitary wasp Eumenes rubronotatus in planar lipid bilayers: Surface charge and pore formation activity. Toxicon 2008, 51, 736–745. [Google Scholar] [CrossRef] [PubMed]
- Konno, K.; Kazuma, K.; Nihei, K. Peptide toxins in solitary wasp venoms. Toxins 2016, 8, 114. [Google Scholar] [CrossRef] [PubMed]
- Piek, T. Neurotoxic kinins from wasp and ant venoms. Toxicon 1991, 29, 139–149. [Google Scholar] [CrossRef]
- Nakanishi, K.; Goodnow, R.; Konno, K.; Niwa, M.; Bukownik, R.; Kalimopoulos, T.A.; Usherwood, P.N.R.; Eldefrawi, A.T.; Eldefrawi, M.E. Philanthotoxin-433 (PhTX-433), a non-competitive glutamate receptor inhibitor. Pure Appl. Chem. 1990, 62, 1223–1230. [Google Scholar] [CrossRef]
- Konno, K.; Kawai, N. Pompilidotoxins: Novel peptide neurotoxins blocking sodium channel inactivation from solitary wasp venom. Curr. Med. Chem. Cent. Nerv. Syst. Agents 2004, 4, 139–146. [Google Scholar] [CrossRef]
- Hirai, Y.; Yasuhara, T.; Yoshida, H.; Nakajima, T.; Fujino, M.; Kitada, C. A new mast cell degranulating peptide “mastoparan” in the venom of Vespula lewisii. Chem. Pharm. Bull. 1979, 27, 1942–1944. [Google Scholar] [CrossRef] [PubMed]
- Murata, K.; Shinada, T.; Ohfune, Y.; Hisada, M.; Yasuda, A.; Naoki, H.; Nakajima, T. Novel biologically active peptides from the venom of Polistes rothneyi Iwatai. Biol. Pharm. Bull. 2006, 29, 2493–2497. [Google Scholar] [CrossRef] [PubMed]
- Nakajima, T. Pharmacological biochemistry of vespid venoms. In Venoms of the Hymenoptera: Biochemical, Pharmacological and Behavioural Aspects; Piek, T., Ed.; Academic Press: London, UK, 1986; pp. 309–327. ISBN 0-12-554771-4. [Google Scholar]
- Sforça, M.L.; Oyama, S., Jr.; Canduri, F.; Lorenzi, C.C.B.; Pertinez, T.A.; Konno, K.; Souza, B.M.; Palma, M.S.; Ruggiero Neto, J.; de Azevedo, W.F., Jr.; et al. How C-terminal carboxyamidation alters the mast cell degranulating activity of peptides from the venom of the eumenine solitary wasp. Biochemistry 2004, 43, 5608–5617. [Google Scholar] [CrossRef] [PubMed]
- Murata, K.; Shinada, T.; Ohfune, Y.; Hisada, M.; Yasuda, A.; Naoki, H.; Nakajima, T. Novel mastoparan and protonectin analogs isolated from a solitary wasp, Orancistrocerus drewseni drewseni. Amino Acids 2009, 37, 389–394. [Google Scholar] [CrossRef] [PubMed]
- Baek, J.H.; Lee, S.H. Isolation and molecular cloning of venom peptides from Orancistrocerus drewseni (Hymenoptera: Eumenidae). Toxicon 2010, 55, 711–718. [Google Scholar] [CrossRef] [PubMed]
- Baek, J.H.; Lee, S.H. Differential gene expression profiles in the venom gland/sac of Eumenes pomiformis (Hymenoptera: Eumenidae). Toxicon 2010, 55, 1147–1156. [Google Scholar] [CrossRef] [PubMed]
- Rodrigues Guerra, M.E.; Fadel, V.; Maltarollo, V.G.; Baldissera, G.; Honorio, K.M.; Ruggiero, J.R.; dos Santos Cabrera, M.P. MD simulations and multivariate studies for modeling the antileishmanial activity of peptides. Chem. Biol. Drug Des. 2017, 90, 501–510. [Google Scholar] [CrossRef] [PubMed]
- Konno, K.; Hisada, M.; Itagaki, Y.; Naoki, H.; Kawai, N.; Miwa, A.; Yasuhara, T.; Takayama, H. Isolation and structure of pompilidotoxins (PMTXs), novel neurotoxins in solitary wasp venoms. Biochem. Biophys. Res. Commun. 1998, 250, 612–616. [Google Scholar] [CrossRef]
- Ifrah, D.; Doisy, X.; Ryge, T.S.; Hansen, P.R. Structure-activity relationship study of anoplin. J. Pept. Sci. 2005, 11, 113–121. [Google Scholar] [CrossRef]
- Won, A.; Khan, M.; Gustin, S.; Akpawu, A.; Seebun, D.; Avis, T.J.; Leung, B.O.; Hitchcock, A.P.; Ianoul, A. Investigating the effects of L- to D-amino acid substitution and deamidation on the activity and membrane interactions of antimicrobial peptide anoplin. Biochim. Biophys. Acta 2011, 1808, 1592–1600. [Google Scholar] [CrossRef] [Green Version]
- Won, A.; Pripotnev, S.; Ruscito, A.; Ianoul, A. Effect of point mutations on the secondary structure and membrane interaction of antimicrobial peptide anoplin. J. Phys. Chem. B 2011, 115, 2371–2379. [Google Scholar] [CrossRef] [PubMed]
- Munk, J.K.; Uggerhøj, L.E.; Poulsen, T.J.; Frimodt-Møller, N.; Wimmer, R.; Nyberg, N.T.; Hansen, P.R. Synthetic analogs of anoplin show improved antimicrobial activities. J. Pept. Sci. 2013, 19, 669–675. [Google Scholar] [CrossRef] [PubMed]
- Slootweg, J.C.; van Schaik, T.B.; van Ufford, H.C.Q.; Breukink, E.; Liskamp, R.M.J.; Rijkers, D.T.S. Improving the biological activity of the antimicrobial peptide anoplin by membrane anchoring through a lipophilic amino acid derivative. Bioorgan. Med. Chem. Lett. 2013, 23, 3749–3752. [Google Scholar] [CrossRef] [PubMed]
- Jindrichova, B.; Burketova, L.; Novotna, Z. Novel properties of antimicrobial peptide anoplin. Biochem. Biophys. Res. Commun. 2014, 444, 520–524. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Chen, J.; Zheng, X.; Yang, X.; Ma, P.; Cai, Y.; Zhang, B.; Chen, Y. Design of novel analogues of short antimicrobial peptide anoplin with improved antimicrobial activity. J. Pept. Sci. 2014, 20, 945–951. [Google Scholar] [CrossRef] [PubMed]
- Munk, J.K.; Ritz, C.; Fliedner, F.P.; Frimodt-Møller, N.; Hansen, P.R. Novel method to identify the optimal antimicrobial peptide in a combination matrix, using anoplin as an example. Antimicrob. Agents Chemother. 2014, 58, 1063–1070. [Google Scholar] [CrossRef] [PubMed]
- Uggerhøj, L.E.; Poulsen, T.J.; Munk, J.K.; Fredborg, M.; Sondergaard, T.E.; Frimodt-Møller, N.; Hansen, P.R.; Wimmer, R. Rational design of alpha-helical antimicrobial peptides: do’s and don’ts. ChemBioChem 2015, 16, 242–253. [Google Scholar] [CrossRef] [PubMed]
- Aschi, M.; Luzi, C.; Fiorillo, A.; Bozzi, A. Folding propensity of anoplin: A molecular dynamics study of the native peptide and four mutated isoforms. Biopolymers 2015, 103, 692–701. [Google Scholar] [CrossRef] [PubMed]
- Sahariah, P.; Sørensen, K.K.; HjáLlmarsdóttir, M.A.; Sigurjónsson, Ó.E.; Jensen, K.J.; Másson, M.; Thygesen, M.B. Antimicrobial peptide shows enhanced activity and reduced toxicity upon grafting to chitosan polymers. Chem. Commun. 2015, 51, 11611–11614. [Google Scholar] [CrossRef]
- Daben, M.; Libardo, J.; Nagella, S.; Lugo, A.; Pierce, S.; Angeles-Boza, A.M. Copper-binding tripeptide motif increases potency of the antimicrobial peptide Anoplin via Reactive Oxygen Species generation. Biochem. Biophys. Res. Commun. 2015, 456, 446–451. [Google Scholar] [CrossRef]
- Chionis, K.; Krikorian, D.; Koukkou, A.I.; Sakarellos-Daitsiotis, M.; Panou-Pomonis, E. Synthesis and biological activity of lipophilic analogs of the cationic antimicrobial active peptides anoplin. J. Pept. Sci. 2016, 22, 731–736. [Google Scholar] [CrossRef]
- Salas, R.L.; Garcia, J.K.D.L.; Miranda, A.C.R.; Rivera, W.L.; Nellas, R.B.; Sabido, P.M.G. Effects of truncation of the peptide chain on the secondary structure and bioactivities of palmitoylated anoplin. Peptides 2018, 104, 7–14. [Google Scholar] [CrossRef] [PubMed]
- Baek, J.H.; Ji, Y.; Shin, J.-S.; Lee, S.; Lee, S.H. Venom peptides from solitary hunting wasps induce feeding disorder in lepidopteran larvae. Peptides 2011, 32, 568–572. [Google Scholar] [CrossRef] [PubMed]
- Tossi, A.; Sandri, L.; Giangaspero, A. Amphipathic, α-helical antimicrobial peptides. Biopolymers 2000, 55, 4–30. [Google Scholar] [CrossRef]
- Giangaspero, A.; Sandri, L.; Tossi, A. Amphipathic α-helical antimicrobial peptides—A systematic study of the effects of structural and physical properties on biological activity. Eur. J. Biochem. 2001, 268, 5589–5600. [Google Scholar] [CrossRef] [PubMed]
- Scholtz, J.M.; Qian, H.; York, E.J.; Stewart, J.M.; Baldwin, R.L. Parameters of helix-coil transition theory for alanine-based peptides of varying chain lengths in water. Biopolymers 1991, 31, 1463–1470. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Andreu, D.; Rivas, L. Animal antimicrobial peptides: An overview. Biopolymers 1998, 47, 415–433. [Google Scholar] [CrossRef]
- Schulz, G.E.; Schirmer, R.H. Principles of Protein Structure; Springer: New York, NY, USA, 1978; pp. 10–16. [Google Scholar]
- Sitaram, N.; Nagaraj, R. Interaction of antimicrobial peptides with biological and model membranes: Structural and charge requirements for activity. Biochim. Biophys. Acta 1999, 1462, 29–54. [Google Scholar] [CrossRef]
- Matsuzaki, K.; Sugishita, K.; Harada, M.; Fujii, N.; Miyajima, K. Interactions of an antimicrobial peptide, magainin 2, with outer and inner membranes of Gram-negative bacteria. Biochim. Biophys. Acta 1997, 1327, 119–130. [Google Scholar] [CrossRef] [Green Version]
- Dathe, M.; Wieprecht, T.; Nikolenko, H.; Handel, L.; Maloy, W.L.; MacDonald, D.L.; Beyermann, M.; Bienert, M. Hydrophobicity, hydrophobic moment and angle subtended by charged residues modulate antibacterial and haemolytic activity of amphipathic helical peptides. FEBS Lett. 1997, 403, 208–212. [Google Scholar] [CrossRef] [Green Version]
- Dathe, M.; Nikolenko, H.; Meyer, J.; Beyermann, M.; Bienert, M. Optimization of the antimicrobial activity of Magainin peptides by modification of charge. FEBS Lett. 2001, 501, 146–150. [Google Scholar] [CrossRef]
- Yeaman, M.R.; Yount, N.Y. Mechanisms of antimicrobial peptide action and resistance. Pharmacol. Rev. 2003, 55, 27–55. [Google Scholar] [CrossRef] [PubMed]
- Matsuzaki, K. Magainins as paradigm for the mode of action of pore forming polypeptides Biochim. Biophys. Acta 1998, 1376, 391–400. [Google Scholar] [CrossRef]
- Lin, Y.-C.; Lim, Y.F.; Russo, E.; Schneider, P.; Bolliger, L.; Edenharter, A.; Altmann, K.H.; Halin, C.; Hiss, J.A.; Schneider, G. Multidimensional design of anticancer peptides. Angew. Chem. Int. Ed. 2015, 54, 10370–10374. [Google Scholar] [CrossRef] [PubMed]
- Torres, M.D.T.; Pedron, C.N.; Araujo, I.; Silva, P.I., Jr.; Silva, F.D.; Oliveira, V.X. Decoralin analogs with increased resistance to degradation and lower hemolytic activity. ChemistrySelect 2017, 2, 18–23. [Google Scholar] [CrossRef]
- Torres, M.D.T.; Pedron, C.N.; Silva Lima, J.A.; Silva, P.I., Jr.; Silva, F.D.; Oliveira, V.X. Antimicrobial activity of leucine-substituted decoralin analogs with lower hemolytic activity. J. Pept. Sci. 2017, 23, 818–823. [Google Scholar] [CrossRef] [PubMed]
- Torres, M.D.T.; Andrade, G.T.; Sato, R.H.; Pedron, C.N.; Manieri, T.M.; Cerchiaro, G.; Ribeiro, A.O.; Fuente-Nunez, C.; Oliveira, V.X., Jr. Natural and redesigned wasp venom peptides with selective antitumoral activity. Beilstein J. Org. Chem. 2018, 14, 1693–1703. [Google Scholar] [CrossRef] [Green Version]
- Torres, M.D.T.; Silva, F.D.; Pedron, C.N.; Capurro, M.L.; Fuente-Nunez, C.; Oliveira, V.X., Jr. Peptide design enables reengineering of an inactive wasp venom peptide into synthetic antiplasmodial agents. ChemistrySelect 2018, 3, 5859–5863. [Google Scholar] [CrossRef]
- Hol, W.G.; van Duijnen, P.T.; Berendsen, H.J.C. The α-helix dipole and the properties of proteins. Nature 1978, 273, 443–446. [Google Scholar] [CrossRef]
- Joshi, H.V.; Meier, M.S. The effect of a peptide helix macrodipole on the pKa of an Asp side chain carboxylate. J. Am. Chem. Soc. 1996, 118, 12038–12044. [Google Scholar] [CrossRef]
- Rohl, C.A.; Baldwin, R.L. Deciphering rules of helix stability in peptides. Methods Enzymol. 1998, 295, 1–26. [Google Scholar] [CrossRef] [PubMed]
- Chou, P.Y.; Fasman, G.D. Conformational parameters for amino acids in helical, β-sheet, and random coil regions calculated from proteins. Biochemistry 1974, 13, 211–222. [Google Scholar] [CrossRef] [PubMed]
- Gans, P.J.; Lyu, P.C.; Manning, M.C.; Woody, R.W. The helix-coil transition in heterogeneous peptides with specific side-chain interactions: Theory and comparison with CD spectral data. Biopolymers 1991, 31, 1605–1614. [Google Scholar] [CrossRef] [PubMed]
- O’Neil, K.T.; DeGrado, W.F. A thermodynamic scale for the helix-forming tendencies of the commonly occurring amino acids. Science 1990, 250, 646–651. [Google Scholar] [CrossRef] [PubMed]
- Marqusee, S.; Robbins, H.V.; Baldwin, R.L. Unusually stable helix formation in short alanine-based peptides. Proc. Natl. Acad. Sci. USA 1989, 86, 5286–5290. [Google Scholar] [CrossRef] [PubMed]
- Blondelle, S.E.; Forood, B.; Houghten, R.A.; Pérez-Payá, E. Secondary structure induction in aqueous vs membrane-like environments. Biopolymers 1997, 42, 489–498. [Google Scholar] [CrossRef]
- Waterhous, D.V.; Johnson, W.C., Jr. Importance of environment in determining secondary structure in proteins. Biochemistry 1994, 33, 2121–2128. [Google Scholar] [CrossRef] [PubMed]
- Zhong, L.; Johnson, C., Jr. Environment affects amino acid preference for secondary structure. Proc. Natl. Acad. Sci. USA 1992, 89, 4462–4465. [Google Scholar] [CrossRef]
- Deber, C.M.; Li, S.C. Peptides in membranes: Helicity and hydrophobicity. Biopolymers 1995, 37, 295–318. [Google Scholar] [CrossRef] [PubMed]
- Blondelle, S.E.; Houghten, R.A. Design of model amphipathic peptides having potent antimicrobial activities. Biochemistry 1992, 31, 12688–12694. [Google Scholar] [CrossRef] [PubMed]
- Dathe, M.; Wieprecht, T. Structural features of helical antimicrobial peptides: Their potential to modulate activity on model membranes and biological cells. Biochim. Biophys. Acta 1999, 1462, 71–87. [Google Scholar] [CrossRef]
- Dathe, M.; Schümann, M.; Wieprecht, T.; Winkler, A.; Beyermann, M.; Krause, E.; Matsuzaki, K.; Murase, O.; Bienert, M. Peptide helicity and membrane surface charge modulate the balance of electrostatic and hydrophobic interactions with lipid bilayers and biological membranes. Biochemistry 1996, 35, 12612–12622. [Google Scholar] [CrossRef] [PubMed]
- Eisenberg, D.; Schwarz, E.; Komaromy, M.; Wall, R. Analysis of membrane and surface protein sequences with the hydrophobic moment plot. J. Mol. Biol. 1984, 179, 125–142. [Google Scholar] [CrossRef]
- Dathe, M.; Meyer, J.; Beyermann, M.; Maul, B.; Hoischen, C.; Bienert, M. General aspects of peptide selectivity towards lipid bilayers and cell membranes studied by variation of the structural parameters of amphipathic helical model peptides. Biochim. Biophys. Acta 2002, 1558, 171–186. [Google Scholar] [CrossRef]
- Pathak, N.; Salas-Auvert, R.; Ruche, G.; Janna, M.H.; McCarthy, D.; Harrison, R.G. Comparison of the effects of hydrophobicity, amphiphilicity, and α-helicity on the activities of antimicrobial Peptides. Proteins Struct. Funct. Bioinf. 1995, 22, 182–186. [Google Scholar] [CrossRef] [PubMed]
- Wimley, W.C. Describing the mechanism of antimicrobial peptide action with the interfacial activity model. ACS Chem. Biol. 2010, 5, 905–917. [Google Scholar] [CrossRef] [PubMed]
- Lum, K.; Chandler, D.; Weeks, J.D. Hydrophobicity at small and large length scales. J. Phys. Chem. B 1999, 103, 4570–4577. [Google Scholar] [CrossRef]
- Wieprecht, T.; Dathe, M.; Beyermann, M.; Krause, E.; Maloy, W.L.; MacDonald, D.L.; Bienert, M. Peptide hydrophobicity controls the activity and selectivity of Magainin 2 amide in interaction with membranes. Biochemistry 1997, 36, 6124–6132. [Google Scholar] [CrossRef]
- Leite, N.B.; da Costa, L.C.; Dos Santos Alvares, D.; Dos Santos Cabrera, M.P.; De Souza, B.M.; Palma, M.S.; Ruggiero Neto, J. The effect of acidic residues and amphipathicity on the lytic activities of mastoparan peptides studied by fluorescence and CD spectroscopy. Amino Acids 2011, 40, 91–100. [Google Scholar] [CrossRef]
- Neville, F.; Cahuzac, M.; Konovalov, O.; Ishitsuka, Y.; Lee, K.Y.C.; Kuzmenko, I.; Kale, G.M.; Gidalevitz, D. Lipid head group discrimination by antimicrobial peptide LL-37: Insight into mechanism of action. Biophys. J. 2006, 90, 1275–1287. [Google Scholar] [CrossRef]
- Leontiadou, H.; Mark, A.E.; Marrink, S.J. Antimicrobial peptides an action. J. Am. Chem. Soc. 2006, 128, 12156–12161. [Google Scholar] [CrossRef]
- Rzepiela, A.J.; Sengupta, D.; Goga, N.; Marrink, S.J. Membrane poration by antimicrobial peptides combining atomistic and coarse-grained descriptions. Faraday Discuss. 2010, 144, 431–443. [Google Scholar] [CrossRef]
- Giladevitz, D.; Ishitzuka, Y.; Murean, A.S.; Konovalov, O.; Waring, A.L.; Lehrer, R.I.; Lee, K.Y.C. Interaction of antimicrobial peptide protegrin with biomembranes. Proc. Natl. Acad. Sci. USA 2003, 100, 6302–6307. [Google Scholar] [CrossRef] [Green Version]
- Andreev, K.; Martynowycz, M.W.; Huang, M.L.; Kuzmenko, I.; Bu, W.; Kirshenbaum, K.; Gidalevitz, D. Hydrophobic interactions modulate antimicrobial peptoid selectivity toward anionic lipid membranes. Biochim. Biophys. Acta 2018, 1860, 1414–1423. [Google Scholar] [CrossRef]
- Rangel, M.; Konno, K.; Brunaldi, K.; Procopio, J.; Freitas, J.C. Neurotoxic activity induced by a haemolytic substance in the extract of the marine sponge Geodia corticostylifera. Comp. Biochem. Physiol. C Toxicol. Pharmacol. 2005, 141, 207–215. [Google Scholar] [CrossRef]
- Boland, M.P.; Separovic, F. Membrane interactions of antimicrobial peptides from Australian tree frogs. Biochim. Biophys. Acta 2006, 1758, 1178–1183. [Google Scholar] [CrossRef] [Green Version]
- Shai, Y. Mechanism of the binding, insertion and destabilization of phospholipid bilayer membranes by α-helical antimicrobial and cell non-selective membrane-lytic peptides. Biochim. Biophys. Acta 1999, 1462, 55–70. [Google Scholar] [CrossRef]
- Dos Santos Cabrera, M.P.; Arcisio-Miranda, M.; da Costa, L.C.; De Souza, B.M.; Broggio Costa, S.T.; Palma, M.S.; Ruggiero Neto, J.; Procopio, J. Interactions of mast cell degranulating peptides with model membranes: A comparative biophysical study. Arch. Biochem Biophys. 2009, 486, 1–11. [Google Scholar] [CrossRef]
- Matsuzaki, K.; Murase, O.; Fujii, N.; Miyajima, K. An antimicrobial peptide, magainin 2, induced rapid flip-flop of phospholipids coupled with pore formation and peptide translocation. Biochemistry 1996, 35, 11361–11368. [Google Scholar] [CrossRef]
- Yang, L.; Harroun, T.A.; Weiss, T.M.; Ding, L.; Huang, H.W. Barrel-stave model or toroidal model? A case study on melittin pores. Biophys. J. 2001, 81, 1475–1485. [Google Scholar] [CrossRef]
- Strandberg, K.; Westerberg, S. Composition of phospholipids and phospholipid fatty acids in rat mast cells. Mol. Cell. Biochem. 1976, 11, 103–107. [Google Scholar] [CrossRef] [PubMed]
- Christensen, B.; Fink, J.; Merrifield, R.B.; Mauzerall, D. Channel-forming properties of cecropins and related model compounds incorporated into planar lipid membranes. Proc. Natl. Acad. Sci. USA 1988, 85, 5072–5076. [Google Scholar] [CrossRef] [PubMed]
- Bechinger, B.; Lohner, K. Detergent-like actions of linear amphipathic cationic antimicrobial peptides. Biochim. Biophys. Acta 2006, 1758, 1529–1539. [Google Scholar] [CrossRef] [PubMed] [Green Version]




{kind=link}
{kind=link}
{kind=link}
| Peptide | Sequence | Peptide | Sequence |
|---|---|---|---|
| EMP-AF | INLLKIAKGIIKSL-NH2 | Eumenitin | LNLKGIFKKVKSLLT |
| EMP-EF | FDVMGIIKKIASAL-NH2 | Eumenitin-F | LNLKGLFKKVASLLT |
| EMP-ER | FDIMGLIKKVAGAL-NH2 | Eumenitin-R | LNLKGLIKKVASLLN |
| EMP-EM1 | LKLMGIVKKVLGAL-NH2 | EpVP1 | INLKGLIKKVASLLT |
| EMP-EM2 | LKLLGIVKKVLGAI-NH2 | Decoralin | SLLSLIRKLIT |
| EpVP2a | FDLLGLVKKVASAL-NH2 | OdVP3 | KDLHTVVSAILQAL-NH2 |
| EpVP2b | FDLLGLVKSVVSAL-NH2 | EMP-OD (OdVP1) | GRILSFIKGLAEHL-NH2 |
| Anoplin | GLLKRIKTLL-NH2 | Orancis-Protonectin (OdVP2) | ILGIITSLLKSL-NH2 |
| Peptides | Antimicrobial Activity (MIC, μM) | Hemolysis (EC50, μM) | Degranulation (EC50, μM) | References | ||
|---|---|---|---|---|---|---|
| S. aureus | E. coli | C. albicans | ||||
| EMP-AF | 3 | 33 | - | 50 | 30 | [8,14] |
| EMP-EF | 30 | 30 | 7.5 | 181 | 60 | [12] |
| EMP-ER | 30 | 30 | 7.5 | 200 | 70 | [12] |
| EMP-EM1 | 7 | 7 | >60 | na | 70 | [13] |
| EMP-EM2 | 3 | 3 | >60 | na | 60 | [13] |
| Eumenitin | 6 | 6 | - | na | 70 | [10] |
| Eumenitin-F | >60 | 30 | 7.5 | 353 | na | [12] |
| Eumenitin-R | 60 | 30 | 7.5 | 530 | na | [12] |
| Decoralin | 40 | 80 | 40 | na | na | [11] |
| Anoplin | 4 | 43 | - | na | 30 | [9] |
| Peptides | N | Q | C-Term | <H> | µ | fH | References | ||
|---|---|---|---|---|---|---|---|---|---|
| TFE | SDS | PC | |||||||
| EMP-AF | 14 | +4 | amide | 0.051 | 0.342 | 0.55 | 0.72 | 0.16 | [14] |
| EMP-EF | 14 | +2 | amide | 0.115 | 0.279 | 0.41 | 0.44 | nd | [12] |
| EMP-ER | 14 | +2 | amide | 0.131 | 0.251 | 0.53 | 0.59 | nd | [12] |
| EMP-EM1 | 14 | +4 | amide | 0.104 | 0.258 | 0.31 | 0.33 | 0.11 | [13] |
| EMP-EM2 | 14 | +4 | amide | 0.138 | 0.278 | 0.37 | 0.41 | < 0.03 | [13] |
| Eumenitin | 15 | +3 | carboxyl | 0.002 | 0.265 | 0.43 | 0.50 | 0.07/0.11 | [10] |
| Eumenitin-F | 15 | +3 | carboxyl | - | 0.256 | 0.34 | 0.44 | nd | [12] |
| Eumenitin-R | 15 | +3 | carboxyl | - | 0.281 | 0.43 | 0.48 | nd | [12] |
| Decoralin | 11 | +3 | carboxyl | 0.028 | 0.393 | 0.38 | 0.50 | rc | [11] |
| Anoplin | 10 | +4 | amide | - | 0.366 | 0.24 | 0.26 | rc | [9] |
| Peptides | Lipid | Vhold (mV) | Conductance (pS) | SEM | References |
|---|---|---|---|---|---|
| Eumenitin | asolectin | −50 | 118.0/160.0 | 3.67/7.07 | [16] |
| asolectin | +50 | 118.0/160.0 | 3.67/7.07 | [16] | |
| DPhPC | +50 | 61.13 | 7.57 | [16] | |
| DPhPC-cholesterol | −100 | None | - | [16] | |
| DPhPC-cholesterol | +100 | None | - | [16] | |
| Eumenitin-R | asolectin | −100 | 82.5 | 17.1 | [12] |
| asolectin | +100 | 118.8 | 44 | [12] | |
| Eumenitin-F | asolectin | −100 | 298.6 | 51 | [12] |
| asolectin | +100 | 187.1 | 67.7 | [12] | |
| EMP-ER | asolectin | −100 | 68.2 | 4 | [12] |
| asolectin | +100 | 61.4 | 3.7 | [12] | |
| EMP-EF | asolectin | −100 | 33.6 | 8.9 | [12] |
| asolectin | +100 | 32.2 | 6.9 | [12] | |
| Anoplin | asolectin | +100 | 50.7 | 3.8 | [15] |
| asolectin | +130 | 58.4 | 2.7 | [15] | |
| asolectin | +150 | 66.8 | 6.7 | [15] |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
dos Santos Cabrera, M.P.; Rangel, M.; Ruggiero Neto, J.; Konno, K. Chemical and Biological Characteristics of Antimicrobial α-Helical Peptides Found in Solitary Wasp Venoms and Their Interactions with Model Membranes. Toxins 2019, 11, 559. https://0-doi-org.brum.beds.ac.uk/10.3390/toxins11100559
dos Santos Cabrera MP, Rangel M, Ruggiero Neto J, Konno K. Chemical and Biological Characteristics of Antimicrobial α-Helical Peptides Found in Solitary Wasp Venoms and Their Interactions with Model Membranes. Toxins. 2019; 11(10):559. https://0-doi-org.brum.beds.ac.uk/10.3390/toxins11100559
Chicago/Turabian Styledos Santos Cabrera, Marcia Perez, Marisa Rangel, João Ruggiero Neto, and Katsuhiro Konno. 2019. "Chemical and Biological Characteristics of Antimicrobial α-Helical Peptides Found in Solitary Wasp Venoms and Their Interactions with Model Membranes" Toxins 11, no. 10: 559. https://0-doi-org.brum.beds.ac.uk/10.3390/toxins11100559